

Биология внутренних вод, 2019, № 1, стр. 51-55
Окончательно избираемая температура молоди трехиглой колюшки Gasterosteus aculeatus (Linnaeus, 1758)
А. К. Смирнов 1, *, Е. С. Смирнова 1
1 Институт биологии внутренних вод им. И.Д. Папанина РАН
152742 Ярославская обл., Некоузский р-н, пос. Борок, Россия
* E-mail: smirnov@ibiw.yaroslavl.ru
Поступила в редакцию 10.12.2016
После доработки 12.05.2017
Принята к публикации 22.03.2017
Аннотация
В экспериментальных условиях исследованы температуры, предпочитаемые молодью трехиглой колюшки из пресных водоемов прибрежья Белого моря. Окончательная избираемая температура для сеголеток была 15.2°С независимо от температуры предварительного содержания (11, 16 и 20°С). Увеличенная продолжительность опытов (10 сут) позволила отразить весь процесс выбора рыбами температурного оптимума, а не его отдельные этапы. Сопоставление результатов с литературными данными выявило возможность существования различий в температурных характеристиках, как географически удаленных популяций колюшки, так и между ее пресноводной и морской формами.
ВВЕДЕНИЕ
Трехиглая колюшка Gasterosteus aculeatus (Linnaeus, 1758) – вид с очень широким ареалом, включающим в себя бассейны северной части Атлантического и Тихого океанов. В европейской части России колюшка распространена от Кольского полуострова и Новой Земли до Черного моря. Обитает в бассейне Баренцева, Белого (оз. Имандра, все реки Карелии и Архангельской обл., включая р. Северную Двину), Балтийского (Ладожское и Онежское озера) и Черного морей (р. Дон, устье р. Кубань). Однако отсутствует в бассейне Каспийского и Аральского морей и в реках Сибири [1]. Вид адаптирован к таким факторам среды как температура и соленость. Трехиглая колюшка населяет сильно различающиеся водоемы – от ультраолиготрофных с минерализацией <10 г/л до супергалинных с соленостью до 100 г/л. Температурные пределы жизнедеятельности вида также очень широки и варьируют от 0°С (и даже ниже в зимний период в северных морях) до 32°С в горячих источниках Камчатки [1].
Вопросы, затрагивающие изучение температурных границ жизнедеятельности трехиглой колюшки, а также ее температурный оптимум весьма актуальны. Однако, работы посвященные исследованию температурных характеристик G. aculeatus, относительно малочисленны [13–16, 18]. Имеющиеся в них данные различны и не позволяют сделать однозначных выводов о температурах, предпочитаемых видом в целом, а также его отдельными формами (морской, проходной и пресноводной). Также следует отметить кратковременность таких экспериментов, как правило, не превышавшую нескольких часов и не позволяющую в полной мере оценить весь процесс выбора рыбами температурного оптимума [2, 4, 6].
Цель работы – определение окончательно избираемой температуры (ОИТ) у молоди пресноводной формы трехиглой колюшки в условиях длительных экспериментов после предварительной акклимации рыб к различным температурам.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
В работе использовали молодь трехиглой колюшки, выращенную в лабораторных условиях в течение лета и осени 2015 г. Половые продукты отбирали у особей, отловленных в пресноводных озерах прибрежья Кандалакшского залива Белого моря в районе пос. Чкаловский в начале лета 2015 г. После искусственного оплодотворения икру транспортировали в лабораторию Института биологии внутренних вод РАН, где проходила ее инкубация и выклев личинок. Вылупившуюся молодь содержали в аквариумах объемом 30–70 л с ежедневной заменой 30–50% воды. Поскольку температуру в аквариумах не регулировали, она соответствовала таковой в помещении (18–22°С). На начальных этапах развития молоди колюшку кормили науплиями артемии (Artemia salina Linnaeus, 1758), на заключительных – личинками хирономид (Chironomus spp.).
Экспериментальную работу по изучению температурных предпочтений молоди колюшки проводили с ноября по декабрь 2015 г. К началу опытов возраст рыб был 0+, средняя длина тела – 49.7 ± 1.5 мм, средняя масса – 1.3 ± 0.1 г. Перед экспериментом молодь адаптировали к трем различным температурам (11, 16 и 20°С) в течение 10 сут. После этого рыб (по 7 особей) помещали в экспериментальную горизонтальную термоградиентную установку в отсек с температурой, равной температуре акклимации [8]. Во всех опытах температурный градиент охватывал диапазон от 10 до 25°С. Жесткость градиента равнялась 3.5°С · м–1, световой режим – 12 : 12 ч. Каждый опыт продолжался 10 сут. Наблюдения за распределением рыб по отсекам установки осуществляли 9 раз в течение светлого времени суток. На основе этих данных рассчитывали среднесуточную и окончательно избираемую (отсутствие достоверных различий в течение нескольких последовательных суток) температуры. Кормление колюшек проводили ежедневно, используя сухой полнорационный комбинированный корм (SERA vipan). Его вносили только в отсеки установки, где на момент кормления присутствовала молодь, с целью исключить влияние данного фактора на терморегуляционное поведение рыб [8].
При статистической обработке данных для внутригрупповых (в пределах одной температуры акклимации) сравнений величин избираемых температур использовали критерий Вилкоксона. Для межгруппового (различные температуры акклимации) сравнения применяли ранговый дискриминантный анализ Краскела–Уоллиса.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Молодь колюшки, помещенная в температурно-неоднородную среду, достаточно быстро покидала начальные отсеки экспериментальной установки и выбирала температуры, отличные от акклимационных (рис. 1). Выраженность подобного поведения зависела, прежде всего, от степени различия акклимационной и определенной впоследствии ОИТ. Так, рыб, акклимированных к температуре 11°С, уже в течение первых суток чаще отмечали в отсеках с температурой 16–22°С, со средним значением избираемой температуры 18.1°С. Схожим образом колюшки, адаптированные к 20°С, также демонстрировали достаточно резкий уход из начальных отсеков. Уже на вторые сутки опыта их среднесуточная избираемая температура составила 15.4°С, т.е. значительно отличалась от акклимационного значения. Только при температуре акклимации 16°С (достаточно близкой к определенной впоследствии ОИТ), предпочитаемый рыбами температурный диапазон не претерпевал значительных изменений.
Рис. 1.
Динамика среднесуточных избираемых температур (°С) молоди трехиглой колюшки акклимированной к различным температурам: 1 – 11°С, 2 – 16°С, 3 – 20°С (вертикальная линия – ошибка среднего).
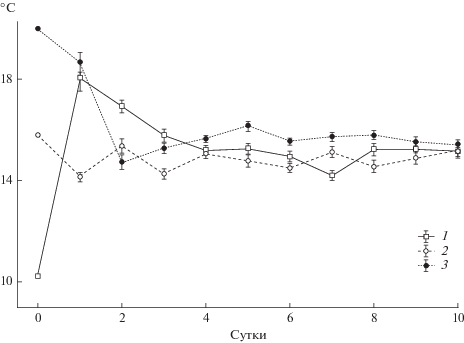
Статистическое сравнение среднесуточных избираемых температур для рыб из различных акклимационных групп выявило в каждой из них отсутствие достоверных различий (p > 0.05) в течение двух последних суток опытов. При этом сами значения избираемых температур за этот период времени были относительно близки – 15.2°С (температура акклимации 11°С), 15.0°С (температура акклимации 16°С) и 15.5°С (температура акклимации 20°С). Сравнение между группами показало, что схожим образом различия также отсутствовали в последние двое суток опытов (p > 0.05). Вследствие этого, полученную среднюю величину (15.2°С) можно считать ОИТ для исследованной молоди, так как она удовлетворяет одному из важнейших ее критериев – независимости от температуры предварительного содержания [12].
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Как и большинство организмов, способных к самоперемещению, молодь трехиглой колюшки, попадая в температурно-неоднородную среду, постепенно скапливается в зоне, наиболее благоприятной для своей жизнедеятельности [3]. Такая зона обычно характеризуется как температурный оптимум для роста и развития особей на текущем этапе онтогенеза [14]. Считается, что при этих температурах затраты на адаптационные нужды минимальны, и все доступные ресурсы расходуются организмом “только на фундаментальные жизненные процессы” [10]. Выбор температуры особями чаще протекал с реакцией избыточного реагирования или перерегулирования, особенно выраженной при заметном различии температур акклимации и ОИТ (рис. 1). Подобная схема поведения характерна для различных видов рыб при помещении их в температурно-неоднородную среду [4, 6].
Ранее в работах [2, 4, 6, 17 и др.] неоднократно поднимался вопрос о понятии “окончательно избираемая” или “предпочитаемая температура”, введенном Фраем [12]. Данный термин был предложен для того, чтобы отличать температуру, предпочитаемую в период переходного процесса, от таковой на момент стабильного выбора. Одним из наиболее важных критериев данного температурного показателя указывается его независимость от температуры предварительного содержания [12, 17]. Именно это условие соблюдалось в настоящей работе. Независимо от температуры акклимации молодь колюшки к концу опытов скапливалась в температурной зоне со средним значением 15.2°С (рис. 1).
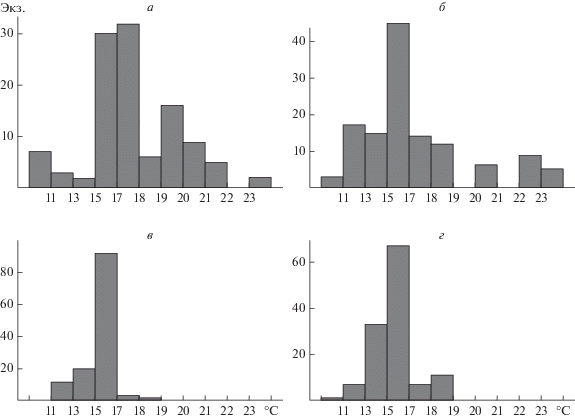
Процесс температурного выбора сопровождался сменой характера распределения рыб в отсеках экспериментальной термоградиентной установки (рис. 2). На этапе переходного процесса рыбы широко распределялись в предоставленном пространстве, посещая и исследуя фактически все отсеки. Однако на момент окончания опытов распределение выраженно сужалось вокруг максимума, расположенного в зоне ОИТ. Это дополнительно подтверждает комфортность выбранной температуры для жизнедеятельности молоди колюшки. Поскольку корма было в достаточном количестве, то посещаемость отсеков, удаленных от температурного оптимума, резко сокращалась.
Рис. 2.
Распределение молоди колюшки в градиенте температур (ось абсцисс, °С) в различные периоды эксперимента: для рыб акклимированных к 11°С (а – начало, в – конец эксперимента), к 20°С (б – начало, г – конец эксперимента).
В экспериментах Гарсайда и др. [13] в качестве ОИТ колюшек, отловленных в приливных водоемах канадской провинции Новая Шотландия, указывались значения 16 и 18°С для пресной и соленой воды соответственно. Следует отметить, что эти величины расчетные (на основе определенного алгоритма), а продолжительность экспериментов не превышала нескольких часов. Лачанце и др. [16] показали, температурный диапазон, предпочитаемый трехиглой колюшкой из приливных водоемов провинции Квебек (9–12°С), эксперименты также были непродолжительны по времени (~2 ч). Также в работе [16] показано, что предпочитаемый температурный диапазон у симпатрических видов колюшек либо схож – 11–14°С (для двухиглой колюшки Gasterosteus wheatlandi Putnam, 1867), либо несколько отличался и носил бимодальный характер – 9–10 и 15–16°С (для девятииглой колюшки Pungitius pungitius (Linnaeus, 1758)). Подробное исследование сезонной динамики температурных предпочтений трехиглой колюшки проведено норвежскими исследователями [18]. Рыб отлавливали в морской воде Осло-фьорда в течение всего года и исследовали их избираемую температуру. Независимо от времени года колюшки выбирали температуры <11°С, чаще всего они встречались в диапазоне температур 4–8°С. Как и в предыдущих исследованиях, продолжительность опытов была невелика (~1 сут).
Таким образом, даже у исследованных ранее морских форм трехиглой колюшки наблюдается большое разнообразие значений избираемых температур. По-видимому, это может быть напрямую связано с различием в методических подходах к изучению данного поведенческого показателя, и прежде всего, с продолжительностью опытов. Ранее было показано [4, 6, 7], что для достижения ОИТ рыбы затрачивают от 5 до 10 сут. Такая длительность связана со сложным характером реакции термопреферендума. Известно, что в процессе температурного выбора затрагиваются различные уровни организации от организменного до клеточного. И если поведенческие реакции достаточно скоротечны, то физиолого-биохимические адаптационные процессы требуют существенных временных затрат [5, 9, 11]. Поэтому более продолжительные эксперименты наглядно демонстрируют, что только спустя несколько суток после помещения в температурный градиент рыбы выбирают температуры, близкие к ОИТ (рис. 1).
Выводы. Выявлено, что ОИТ для сеголетков колюшки из пресноводной популяции бассейна Белого моря (прибрежье Кандалакшского залива) равна 15.2°С. Это значение ближе всего соответствует расчетной величине, приведенной для морской формы колюшки из Канады при помещении ее в пресноводный температурный градиент [13]. Имеющихся на сегодняшний день экспериментальных данных явно недостаточно для того, чтобы однозначно оценить существование различий в температурных характеристиках пресноводной и морской форм трехиглой колюшки, а также среди различных популяций этого широко распространенного вида.
Авторы благодарны А.А. Болотовскому за предоставленный для исследования живой материал.
Работа выполнена в рамках государственного задания № АААА-А18-118012690102-9 при частичной поддержке темы Президиума РАН: 0122-2018-0001.
Список литературы
Атлас пресноводных рыб России. М.: Наука, 2002. Т. 2. 253 с.
Голованов В.К. Эколого-физиологические закономерности распределения и поведения пресноводных рыб в термоградиентных условиях // Вопр. ихтиологии. 2013. Т. 53. № 3. С. 286–314.
Ивлев В.С. Элементы физиологической гидробиологии // Физиология морских животных. М.: Наука, 1966. С. 3–45.
Лапкин В.В., Свирский А.М., Сопов Ю.Н. Избираемая температура и температура акклимации рыб // Зоол. журн. 1979. Т. 58. Вып. 11. С. 1659–1670.
Озернюк Н.Д. Температурные адаптации. М.: Изд-во Москов. ун-та, 2000. 205 с.
Свирский А.М. Поведение рыб в гетеротермальных условиях // Поведение и распределение рыб: Докл. 2-го Всерос. совещ. Борок, 1996. С. 140–152.
Свирский А.М., Голованов В.К. Влияние температуры акклимации на терморегуляционное поведение молоди леща Abramis brama (L.) в различные сезоны года // Вопр. ихтиологии. 1991. Т. 31. Вып. 6. С. 974–980.
Смирнов А.К. Влияние наличия пищи в зоне температурного оптимума на поведение молоди речного окуня Perca fluviatilis L. // Вестн. Астрахан. гос. тех. ун-та. Сер. Рыб. хоз-во. 2013. № 1. С. 75–82.
Хлебович В.В. Акклимация животных организмов. Л.: Наука, 1981. 136 с.
Шилов И.А. Экология. М.: Высш. шк., 2001. 512 с.
Bicego K.C., Barros R.C.H., Branco L.G.S. Physiology of temperature regulation: comparative aspects // Comp. Biochem. Physiol. – Part A: Molecular & Integrative Physiology. 2007. V. 147. P. 616–639.
Fry F.E.J. Effect of the environment on animal activity // Univ. Toronto Studies, Biol. Ser., Publ. Ont. Fish. Res. Lab., 1947. № 68. 62 p.
Garside E.T., Heinze D.G., Barbour S.E. Thermal preference in relation to salinity in the threespine stickleback, Gasterosteus aculeatus L., with an interpretation of its significance // Can. J. Zool. 1977. V. 55(3). P. 590–594.
Jobling M. Temperature tolerance and the final preferendum – rapid methods for the assessment of optimum growth temperatures // J. Fish Biol. 1981. V. 19. P. 439–455.
Jordan C.M., Garside E.T. Upper lethal temperatures of threespine stickleback, Gasterosteus aculeatus (L.) in relation to thermal and osmotic acclimation, ambient salinity, and size // Can. J. Zool. 1972. V. 50. P. 1405–1411.
Lachance S., Magnan P., Fitz Gerald G.J. Temperature preferences of three sympatric sticklebacks (Gasterosteidae) // Can. J. Zool. 1987. V. 65(6). P. 1573–1576.
Reynolds W.W., Casterlin M.E. Behavioral thermoregulation and the “final preferendum” paradigm. Thermoregulation in ectotherms. Symp. Richmond. 1978 // Amer. Zool. 1979. V. 19. № 1. P. 211–224.
Røed K.H. The Temperature Preference of the Three-spined Stickleback, Gasterosteus aculeatus L. (Pisces), Collected at Different Seasons // Sarsia. 1979. V. 64. P. 137–141.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод