

Биология внутренних вод, 2019, № 2-1, стр. 62-70
Влияние моллюска Dreissena polymorpha (Pallas, 1771) (Mollusca, Dreissenidae) и разных возрастных групп окуня Perca fluviatilis (L.) (Pisces, Perciidae) на структуру и основные характеристики макрозообентоса экспериментальных мезокосмов
Г. Х. Щербина 1, *, В. В. Безматерных 2
1 Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
152742 Ярославская обл., Некоузский р-н, пос. Борок, Россия
2 Пермское отделение ФГБНУ “ГосНИОРХ” им. Л.С. Берга
614002 Пермь, ул. Чернышевского, д. 3, Россия
* E-mail: gregory@ibiw.yaroslavl.ru
Поступила в редакцию 30.08.2016
После доработки 02.02.2018
Принята к публикации 05.04.2018
Аннотация
Исследован состав макрозообентоса экспериментальных мезокосмов из 116 таксонов, наиболее широко представлены хирономиды (50 видов), моллюски (25) и олигохеты (21). Большинство таксонов (61%) – обычные для биоценоза Dreissena polymorpha (Pallas, 1771) Рыбинского водохранилища, остальные вселились за период проведения эксперимента. В мезокосмах с личинками окуня отмечено минимальное количество вселенцев, в вариантах с дрейссеной – максимальное. Показано, что в отсутствии дрейссены возрастные группы окуня, потребляющие макрозообентос, снижают его основные структурные характеристики. В бассейнах с годовиками окуня и дрейссеной отмечена максимальная сумма основных структурных характеристик, что указывает на позитивную средообразующую роль Dreissena polymorpha в бентосных сообществах и возможность использования ее друз в качестве убежищ.
ВВЕДЕНИЕ
Влияние Dreissena polymorpha на бентоценозы водоемов неоднократно показано [3, 8, 18, 24 и др.]. Однако материалов, отражающих совместное воздействие этого моллюска и пищевой активности рыб-бентофагов на состояние сообществ донных макробеспозвоночных, недостаточно. В 1993 г. на экспериментальной базе “Сунога” Института биологии внутренних вод им. И.Д. Папанина РАН проведен эксперимент о влиянии дрейссены полиморфной и различных возрастных групп окуня на видовой состав и структуру макрозообентоса экспериментальных мезокосмов. Вопросы развития и роста окуня в условиях проведенного эксперимента освещены в работе [27].
Цель работы – выявление совокупного влияния дрейссены и разных возрастных групп окуня на основные структурные характеристики макрозообентоса и его отдельных компонентов.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Эксперимент проводили в бетонных прямоугольных бассейнах размером 4 × 4 м объемом 15 м3. На дно каждого бассейна (площадь 16 м2) укладывали суглинок толщиной ~3 см, заливали небольшой слой воды (20 см), затем закладывали ил из Волжского плеса Рыбинского водохранилища (по 120 дночерпателей ДАК-250) и поднимали уровень воды до 90 см. Было установлено 5 вариантов (по 3 бассейна каждый): K – в бассейны помещали только ил; ГО – помещали ил и выпускали в бассейн 12 годовиков окуня со средней массой 3.7 г и средней длиной 6.3 см; ЛО – помещали ил и выпускали в бассейн 1100 личинок окуня со средней массой 3.9 мг и длиной 8.5 мм; ГО + Д – помещали ил, равномерно распределяли по дну бассейна друзы дрейссены из расчета 0.75 кг/м2 и выпускали в бассейн 12 годовиков окуня; ЛО + Д – помещали ил, друзы дрейссены из расчета 0.75 кг/м2 и выпускали в бассейн 1100 личинок окуня. Окунь выбран в связи с широким распространением в прибрежной зоне Рыбинского водохранилища и легким приспособлением к разным экологическим условиям. Его годовики в открытой части водоема – типичные бентофаги, основу пищи которых составляют крупные личинки хирономид – Lipiniella araenicola Shilova и Chironomus muratensis Ryser et al. (> 90% массы пищевого комка).
Пробы макрозообентоса отбирали еженедельно с 10 мая по 11 августа 1993 г. в каждом бассейне по восемь дночерпателей площадью захвата 1/400 м2 (конструкция дночерпателя Г.Х. Щербины), промывали через мельничный газ с размером ячеи 210 мкм. Организмы из остатков грунта выбирали живыми, затем моллюсков фиксировали в 70%-ном спирте, остальных беспозвоночных – в 8%-ном формалине и после трехмесячной фиксации, отведенной на стабилизацию массы фиксированных организмов [1], приступали к их камеральной обработке. Всего собрано и обработано 195 проб макрозообентоса.
Для определения количества вылетевших видов хирономид с поверхности воды собирали экзувии куколок хирономид в те же сроки, что и макрозообентос. Собранные экзувии фиксировали в жидкости Удеманса и после приготовления тотальных препаратов определяли их видовую принадлежность. Для анализа вылета хирономид, кроме количества вылетевших особей каждого вида и динамики вылета, рассчитывали введенный авторами впервые индекс вылета хирономид (Dx). Индекс представляет собой отношение суммы всех вылетевших особей имаго хирономид в данном варианте за весь период эксперимента (W) к общему числу вылетевших в данном варианте видов хирономид (S): Dx = W/S.
В тексте статьи употребляются термины “аборигены” – виды, попавшие в экспериментальные мезокосмы вместе с грунтом из Волжского плеса Рыбинского водохранилища и “вселенцы” – виды, отсутствовавшие в изначальном составе (в иле, наполнявшем бассейны в начале опыта), чаще всего это насекомые, отложившие кладки в бассейнах в период проведения эксперимента [24].
Камеральную обработку материала проводили согласно общепринятым методикам [1, 9, 19]. Для идентификации видовой принадлежности донных макробеспозвоночных использовали определители [4–7, 10–17, 20–22, 25, 28, 29]. При составлении списка обнаруженных видов придерживались системы и сокращений, изложенных в работе [26] и серии определителей [11–14].
Структуру макрозообентоса оценивали по количеству таксонов, численности, биомассе, встречаемости и доле отдельных таксономических групп в общей численности и биомассе. Индекс доминирования рассчитан по работе [2], индекс Шеннона [23].
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
В составе макрозообентоса экспериментальных мезокосмов обнаружено 116 таксонов, из которых наиболее широко представлены хирономиды (50 видов), моллюски (25) и олигохеты (21). Из всех обнаруженных видов 73 обычны в составе макрозообентоса биоценоза дрейссены Рыбинского водохранилища (аборигены). Остальные 43 вида вселились за период проведения эксперимента (вселенцы). Подавляющее большинство видов (69) были редкими (частота встречаемости в составе макрозообентоса < 10%), в то время как постоянных видов (с частотой встречаемости ≥ 50%) зарегистрировано всего девять: личинки хирономид (7) и олигохеты (2) (табл. 1). Наибольшая частота встречаемости в различных вариантах отмечена у следующих трех видов: Limnodrilus hoffmeisteri (84.6–97.4%), Potamothrix moldaviensis (71.8–92.3%) и Cladotanytarsus wexionensis (66.7–79.5%).
Таблица 1.
Основные структурные характеристики доминирующих видов донных макробеспозвоночных в различных вариантах экспериментальных мезокосмов
| Вид | Вариант эксперимента | ||||
|---|---|---|---|---|---|
| К | ЛО | ЛО + Д | ГО | ГО + Д | |
| Численность, экз./м2 | |||||
| Limnodrilus hoffmeisteri Claparède | 362 | 295 | 373 | 206 | 263 |
| Potamothrix moldaviensis (Vejdovsky et Mrazek) | 247 | 176 | 264 | 115 | 178 |
| Psectrocladius fabricus Zelentzov | 108 | 50 | 133 | 27 | 147 |
| Chironomus cingulatus (Meigen) | 41 | 371 | 569 | 206 | 1063 |
| Cladotanytarsus wexionensis Brundin | 410 | 341 | 795 | 333 | 968 |
| Tanytarsus pallidicornis (Walker) | 306 | 496 | 699 | 51 | 262 |
| T. mendax Kieffer | 36 | 132 | 219 | 65 | 323 |
| Polypedilum nubeculosum (Meigen) | 49 | 59 | 23 | 44 | 71 |
| Биомасса, г/м2 | |||||
| Limnodrilus hoffmeisteri | 1.05 | 0.86 | 1.37 | 0.38 | 0.69 |
| Potamothrix moldaviensis | 0.52 | 0.39 | 0.58 | 0.21 | 0.39 |
| Psectrocladius fabricus | 0.12 | 0.06 | 0.14 | 0.02 | 0.14 |
| Chironomus cingulatus | 0.23 | 1.85 | 2.99 | 1.25 | 3.57 |
| Cladotanytarsus wexionensis | 0.15 | 0.11 | 0.31 | 0.11 | 0.36 |
| Tanytarsus pallidicornis | 0.17 | 0.23 | 0.40 | 0.03 | 0.17 |
| T. mendax | 0.03 | 0.08 | 0.14 | 0.04 | 0.24 |
| Polypedilum nubeculosum | 0.07 | 0.10 | 0.04 | 0.08 | 0.11 |
| Встречаемость, % | |||||
| Limnodrilus hoffmeisteri | 97.4 | 84.6 | 94.9 | 82.1 | 89.7 |
| Potamothrix moldaviensis | 89.7 | 89.7 | 89.7 | 71.8 | 92.3 |
| Psectrocladius fabricus | 41.0 | 35.9 | 46.2 | 20.5 | 35.9 |
| Chironomus cingulatus | 25.6 | 56.4 | 61.5 | 46.2 | 51.3 |
| Cladotanytarsus wexionensis | 66.7 | 69.2 | 69.2 | 69.2 | 79.5 |
| Tanytarsus pallidicornis | 69.2 | 74.4 | 76.9 | 35.9 | 69.2 |
| T. mendax | 30.8 | 56.4 | 69.2 | 43.6 | 71.8 |
| Polypedilum nubeculosum | 43.6 | 41.0 | 30.8 | 38.5 | 48.7 |
| Индекс доминирования | |||||
| Limnodrilus hoffmeisteri | 7.37 | 5.29 | 5.24 | 3.57 | 4.23 |
| Potamothrix moldaviensis | 5.00 | 3.51 | 3.44 | 2.29 | 3.10 |
| Psectrocladius fabricus | 1.77 | 0.88 | 1.37 | 0.42 | 1.50 |
| Chironomus cingulatus | 1.37 | 6.44 | 6.78 | 3.96 | 10.70 |
| Cladotanytarsus wexionensis | 3.53 | 2.65 | 3.69 | 2.59 | 5.07 |
| Tanytarsus pallidicornis | 3.42 | 3.89 | 4.01 | 0.70 | 2.44 |
| T. mendax | 0.68 | 1.63 | 1.83 | 0.94 | 2.98 |
| Polypedilum nubeculosum | 1.19 | 1.17 | 0.42 | 0.97 | 1.19 |
Примечание. Здесь и в табл. 2 полужирным шрифтом выделены значимые различия количественных характеристик.
В начале эксперимента по частоте встречаемости незначительно лидировали представители олигохет, но с вселением в лотки массовых полицикличных видов хирономид последние стали преобладать по этому показателю почти до конца эксперимента. В конце эксперимента повысилась роль двух представителей моллюсков – Lymnaea peregra (Müeller) и Cincina piscinalis (Müeller).
В первую неделю эксперимента число видов во всех вариантах варьировало от 13 до 18, причем основу составляли хирономиды и олигохеты. Затем видовое богатство возросло во всех вариантах, при этом более существенный рост наблюдался в вариантах с дрейссеной (ЛО + Д и ГО + Д), а менее значительный в вариантах с рыбой (ЛО и ГО), контрольные мезокосмы (К) занимали промежуточное положение. В свою очередь в мезокосмах с годовиками окуня количество обнаруженных видов было меньше, чем в вариантах с личинками. Сходная динамика наблюдалась и у числа обнаруженных видов хирономид. Меньшее влияние оказывает присутствие дрейссены в мезокосмах на число обнаруженных олигохет и моллюсков.
Всего за период исследований зарегистрировано 29 видов постоянных (встреченных во всех мезокосмах) видов. Из них хирономиды – 19, олигохеты – 6, моллюски – 3, пиявок – 3. Динамика количества постоянных видов фактически не отличается от таковой предыдущих структурных характеристик. Следует отметить, что в вариантах с годовиками окуня (ГО и ГО + Д) наличие дрейссены способствовало более значительному росту числа постоянных видов, чем в вариантах с личинками окуня (ЛО и ЛО + Д).
Анализ динамики количества видов-вселенцев и вылетевших имаго хирономид показал, что максимальное их количество отмечено в мезокосмах с годовиками окуня и дрейссеной, несколько ниже аналогичный показатель в мезокосмах с личинками окуня и дрейссеной. Наибольшие значения средней за период проведения эксперимента численности и биомассы макробеспозвоночных наблюдали в обоих вариантах с дрейссеной, наименьший аналогичный показатель в мезокосмах с годовиками окуня и контроле (рис. 1, табл. 2).
Рис. 1.
Средняя численность, тыс. экз./м2 (а) и биомасса, г/м2 (б) основных групп макрозообентоса в различных вариантах экспериментальных мезокосмов: 1 – моллюски, 2 – олигохеты, 3 – хирономиды, 4 – прочие.
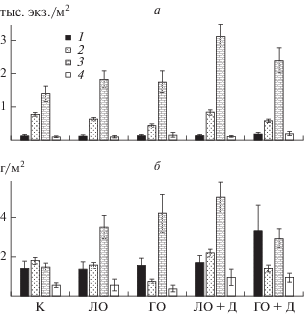
Таблица 2.
Структурные характеристики макрозообентоса в различных вариантах экспериментальных мезокосмов
| Структурная характеристика | Вариант | ||||
|---|---|---|---|---|---|
| К | ЛО | ЛО + Д | ГО | ГО + Д | |
| Всего обнаружено видов | 69 | 65 | 79 | 61 | 75 |
| из них: хирономид | 36 | 30 | 38 | 29 | 33 |
| олигохет | 14 | 15 | 15 | 12 | 15 |
| моллюсков | 15 | 16 | 15 | 13 | 15 |
| прочих | 4 | 4 | 11 | 7 | 12 |
| Число аборигенных видов* | 48 | 48 | 48 | 40 | 46 |
| Число видов-вселенцев* | 21 | 17 | 31 | 21 | 29 |
| Среднее число обнаруженных видов* | 21 ± 1 | 21 ± 1 | 26 ± 1 | 18 ± 1 | 25 ± 1 |
| Число вылетевших видов хирономид* | 22 | 15 | 17 | 10 | 23 |
| Индекс вылета хирономид, $D_{x}^{{\text{*}}}$ | 3.4 | 5.5 | 7.1 | 3.0 | 4.6 |
| Число постоянных видов, P ≥ 50%* | 15 | 14 | 17 | 9 | 22 |
| Средняя численность, тыс. экз./м2* | 2.4 ± 0.2 | 2.7 ± 0.3 | 4.2 ± 0.4 | 2.5 ± 0.3 | 3.4 ± 0.4 |
| Средняя биомасса, г/м2* | 5.2 ± 0.4 | 7.0 ± 0.8 | 9.9 ± 1.1 | 6.9 ± 1 | 8.6 ± 0.9 |
| Сумма основных структурных характеристик | 138 | 130.2 | 160.2 | 110.4 | 161.6 |
При анализе динамики численности и биомассы основных групп макрозообентоса выявлено значительное преобладание по численности хирономид-вселенцев, которые доминировали в течение всего эксперимента (общее для всех пяти вариантов). Роль аборигенов (основа – олигохеты, пиявки и моллюски) незначительна. Сначала общая численность во всех вариантах возрастала, а после достижения максимума убывала. Причем, колебание численности аборигенов в различных вариантах несущественны, а основные изменения тесно связаны с появлением в составе макрозообентоса молоди хирономид-вселенцев и вылетом их имаго (рис. 2).
Рис. 2.
Динамика численности (тыс. экз./м2) и биомассы (г/м2) аборигенов (1) и вселенцев (2) в различных вариантах: К (а, б), ЛО (в, г), ЛО + Д (д, е), ГО (ж, з), ГО + Д (и, к).
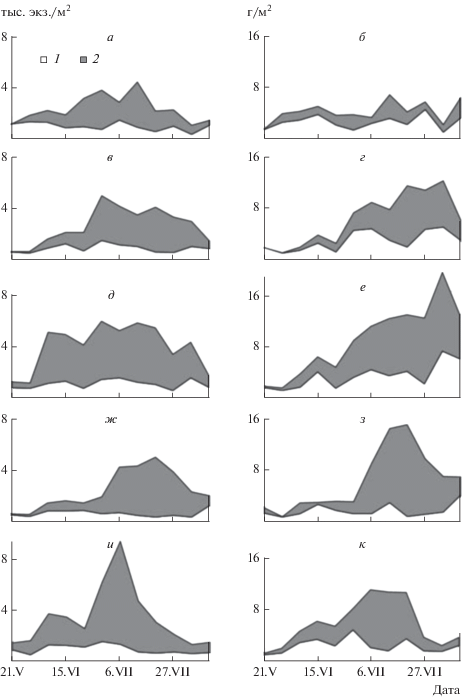
В контрольных мезокосмах численность макрозообентоса изменялась незначительно, с двумя небольшими максимумами в конце июня и середине июля, далее происходило постепенное ее снижение до первоначального значения.
В варианте с годовиками окуня без дрейссены численность вселенцев постепенно возрастала и достигала максимума на 62-e сут эксперимента, после чего постепенно снижалась (рис. 2). Наиболее существенные колебания численности наблюдали в мезокосмах с годовиками окуня и дрейссеной (два максимума). Уже на 20-e сут проведения эксперимента численность донных беспозвоночных здесь достигала 3.5 тыс.экз./м2. Второй максимум численности был значительно выше первого – 9.4 тыс. экз./м2.
В мезокосмах с личинками окуня без дрейссены динамика численности сходна с таковой у годовиков, но пик численности наступил почти на месяц раньше. С переходом личинок на 40-e сут эксперимента к активному потреблению донных беспозвоночных [27] произошла убыль численности. В варианте с личинками окуня и дрейссеной динамика численности значительно отличалась от предыдущих вариантов. Для нее характерен резкий рост уже после 10 сут эксперимента, хотя 70% пищевого комка личинок окуня составили донные беспозвоночные [27]. В последующий период численность макрозообентоса колебалась незначительно, и резкая ее убыль произошла за неделю до конца эксперимента (рис. 2).
Биомасса макрозообентоса в контроле в течение всего эксперимента существенно не изменялась, основу ее составляли олигохеты, хирономиды и в отдельные периоды моллюски. В целом биомасса близка к 4 г/м2, а ее пики носят скорее случайный, чем закономерный характер. Следует отметить, что доля аборигенов была несколько выше, чем видов-вселенцев (рис. 2).
Динамика биомассы аборигенов и вселенцев в обоих вариантах с годовиками окуня носила сходный характер. В первые 5 нед эксперимента биомасса хирономид-вселенцев была незначительной (рис. 2). В последующий период биомасса макрозообентоса возрастала и на 8–10 cут эксперимента в мезокосмах с годовиками окуня составила 10–15 г/м2, с годовиками окуня и дрейссеной – 11–12 г/м2.
В обоих вариантах с личинками окуня динамика биомассы аборигенов и вселенцев имела сходный характер и существенно отличалась от других вариантов. Максимальных величин биомасса достигла в начале августа, после чего происходила резкая ее убыль (рис. 2).
Минимальная доля численности хирономид (58%) и биомассы (28%) отмечена в контроле, а максимальные аналогичные показатели (71 и 61% соответственно) в мезокосмах с годовиками окуня (рис. 1).
Всего из экспериментальных мезокосмов зарегистрирован вылет 33 видов имаго хирономид. Минимальное количество вылетевших имаго отмечено в мезокосмах с годовиками окуня (10), значительно выше оно в вариантах с личинками окуня и дрейсcеной и контроле (23 и 22 вида, соответственно) (табл. 2). Из 33 видов имаго хирономид, вылетевших за время проведения эксперимента, 13 отложили свои кладки повторно, а 11 из них к концу эксперимента осуществили как минимум по два вылета.
За время проведения эксперимента максимальное число обнаруженных видов отмечено в вариантах с дрейссеной, а минимальное – с рыбами. Контрольные мезокосмы по данному показателю занимали промежуточное положение. Причем, увеличение видового богатства в вариантах с дрейссеной было обеспечено, прежде всего, за счет группы прочих, основу которых составляли поденки, ручейники, хаобориды и др., в то время как число обнаруженных видов хирономид, олигохет и моллюсков по вариантам существенно не различалось. Следует отметить, что основную роль в увеличении числа обнаруженных видов в вариантах с дрейссеной вносили виды-вселенцы, в то время как число аборигенных видов различалось незначительно. Расчет среднего числа обнаруженных видов и числа постоянных видов также показал, что в вариантах с дрейссеной его значение значимо выше, чем в остальных трех вариантах (табл. 2).
Индекс вылета также выше в вариантах с Dreissena polymorpha: как в случае с личинками, так и с годовиками его значение возрастает в присутствии моллюска на 1.6. В то же время этот показатель в вариантах с годовиками ниже на 2.5 по сравнению с вариантами с личинками окуня (табл. 2).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Анализ динамики числа обнаруженных видов показал, что максимальное их количество отмечено в вариантах с дрейссеной, а минимальное – в мезокосмах с годовиками окуня (табл. 2). В свою очередь в мезокосмах с годовиками окуня количество обнаруженных видов было меньше, чем в вариантах с личинками. Такая закономерность вполне объяснима, так как годовики окуня – бентофаги, в то время как его личинки в первый месяц эксперимента потребляли в основном зоопланктон [27]. Повышенное видовое богатство в вариантах с дрейссеной связано с ее жизнедеятельностью, в результате которой на дно осаждаются фекалии и агглютинаты, служащие пищей для многих собирателей и глотателей, составляющие основу видового состава макрозообентоса экспериментальных мезокосмов.
Из всех обнаруженных видов 73 обычны в составе макрозообентоса биоценоза дрейссены Рыбинского водохранилища, а остальные 43 вида вселились за период проведения эксперимента (25 видов – хирономиды). Из всех вселенцев 30 видов встречались очень редко и в небольших количествах. Такие виды как моллюск Lymnaea peregra (Müeller) и личинки хирономид: Chironomus cingulatus, Сh. obtusidens Goetghebuer, Microtendipes pedellus (De Geer), Polypedilum nubeculosum, Tanytarsus pallidicornis и Paratanytarsus confusus Palmen отмечены во всех пяти вариантах, в отдельные периоды они довольно многочисленны. Подавляющее большинство вселенцев являются гетеротопами, достигающими массового развития в расположенных недалеко прудах и временных водоемах. Динамика вселенцев несколько отличается от таковой всех предыдущих характеристик. Минимальное количество вселенцев наблюдалось в мезокосмах с личинками окуня, несколько выше у годовиков окуня и контроле и значительно больше в обоих вариантах с дрейссеной (табл. 2). Так как основу вселенцев составляли личинки хирономид из трибы Tanytarsini (подавляющее большинство из которых по типу питания собиратели), фильтрационная деятельность дрейссены создавала хорошие трофические условия для них, что привело к повышенному их видовому богатству.
Как было отмечено ранее, колебание численности аборигенов в различных вариантах незначительны, а основные изменения тесно связаны с появлением в составе макрозообентоса молоди хирономид-вселенцев и вылетом их имаго (рис. 2). Наиболее существенные колебания численности наблюдались в вариантах с дрейссеной, где у ГО + Д отмечено два максимума, а у ЛО + Д – четыре.
Снижение численности в варианте ГО + Д во второй половине июня связано с первым пиком вылета имаго хирономид. Второй максимум приурочен к массовому появлению второго поколения личинок хирономид (90% всего макрозообентоса) и особенно их мелких представителей из трибы Tanytarsini. Резкое снижение численности в последующий период эксперимента вызвано, по-видимому, двумя причинами: во-первых, в конце июня–начале июля наблюдался второй пик вылета имаго хирономид; во-вторых, личинки хирономид испытывали двойной пресс хищников – годовиков окуня и хищных донных беспозвоночных, численность и биомасса которых к концу эксперимента здесь была > 20% всего макрозообентоса. Существенное превышение численности макробеспозвоночных (особенно хирономид в варианте ГО + Д над вариантом ГО) связано с тем, что многочисленные мелкие личинки хирономид находили свое убежище среди друз дрейссены, в то время как в бассейнах без нее они были более доступны при питании немногочисленных годовиков окуня.
Как было отмечено ранее, динамика биомассы аборигенов и вселенцев в обоих вариантах с годовиками окуня носила сходный характер. Несколько повышенная биомасса аборигенов в мезокосмах с дрейссеной связана с ее фильтрационной деятельностью. По-видимому, на относительно слабый рост биомассы хирономид в первые 5 нед эксперимента оказывало наличие большой доли пиявок, для которых хирономиды были основой пищи [6]. После их убыли рост биомассы шел значительно быстрее и в июле достиг максимального значения. С появлением в составе макрозообентоса доминирующего здесь до конца эксперимента крупного представителя хирономид – Chironomus cingulatus произошел резкий скачок биомассы. Дальнейшее ее снижение вызвали, по-видимому, две причины. Во-первых, с 20 июля начался вылет имаго Ch. cingulatus, особенно ярко выраженный в мезокосмах с дрейссеной, где популяция достигала состояния предкуколки через 3 нед. Во-вторых, в мезокосмах без дрейссены годовики окуня, в первую очередь, потребляли более крупных особей данного вида, о чем свидетельствовал незначительный рост средней индивидуальной массы личинок мотыля в данном варианте (рис. 3). В мезокосмах с дрейссеной крупные личинки находили убежище среди друз дрейссены, продукты жизнедеятельности которой способствовали такому быстрому темпу росту мотыля.
Рис. 3.
Средняя индивидуальная масса личинок Chironomus cingulatus в различных вариантах экспериментальных мезокосмов: 1 – ГО, 2 – ГО + Д, 3 – ЛО, 4 – ЛО + Д.
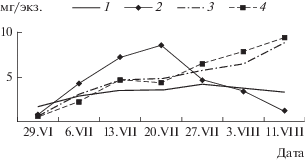
Резкое снижение биомассы в конце июля вызвано, по-видимому, несколькими причинами. Во-первых, значительно возросла биомасса пиявок. Во-вторых, трофические ресурсы планктонных сообществ были основательно истощены, что делало фильтрационную деятельность дрейссены энергетически невыгодной для нее, а значит, препятствовало поступлению в грунт пищи для собирателей и глотателей. В-третьих, эти мезокосмы были наиболее заросшие нитчаткой, которая в конце июля покрывала значительную часть друз дрейссены, что, по-видимому, влияло на развитие видов-аборигенов.
В обоих вариантах с личинками окуня динамика биомассы аборигенов и вселенцев имела довольно сходный характер и существенно отличалась от других вариантов. Максимальных величин она достигала в начале августа, после чего происходила резкая убыль биомассы (рис. 2). Причем, в мезокосмах с дрейссеной она росла более существенно до конца эксперимента, хотя именно здесь у личинок окуня уже на 10-е сут эксперимента 70% пищевого комка составляли хирономиды [27]. Хотя в вариантах с личинками окуня численность макрозообентоса во второй половине эксперимента убывала, его биомасса в этот период существенно увеличилась (рис. 2). Это связано с основным доминирующим видом во второй половине эксперимента – личинками Ch. cingulatus. В мезокосмах с личинками окуня при переходе последних на питание донными беспозвоночными основными жертвами становились личинки мотыля II и III возрастов (динамика роста средней индивидуальной массы с 0.61–0.62 мг в конце июня до 8.61–9.32 мг в конце эксперимента) (рис. 3). Средние за весь период эксперимента значения численности, биомассы и индекса Шеннона были наибольшими в мезокосмах с личинками окуня и дрейссеной, а минимальные – в контроле. Причем, велика роль хирономид во всех вариантах, за исключением контроля, где доминировали моллюски (рис. 1, табл. 2).
Как было отмечено выше, наибольший интерес из всех структурных характеристик представляет индекс вылета хирономид. Причем, Dreissena polymorpha в обоих случаях увеличивает его значение на 1.6 благодаря улучшению трофических условий их обитания, а возраст окуня – в 2.5 (табл. 2), так как личинки окуня не могут питаться куколками во время вылета имаго хирономид из-за их крупных размеров. Годовики окуня даже в естественных водоемах предпочитают питаться куколками хирономид, которые во время вылета некоторое время вынуждены находиться в толще воды, где становятся легкой добычей не только для бентофагов, но и для рыб-планктофагов.
Следует также отметить, что в мезокосмах с годовиками окуня без дрейссены большинство структурных характеристик, как правило, меньше, чем таковые с личинками. Такая закономерность вполне объяснима, так как годовики окуня уже в начале эксперимента – бентофаги, в то время как его личинки в первый месяц эксперимента потребляли в основном зоопланктон, и только в дальнейшем донные беспозвоночные стали по биомассе преобладать в их рационе [27]. В бассейнах с годовиками окуня и дрейссеной, наоборот, отмечены максимальные значения числа вылетевших видов хирономид и числа постоянных видов (табл. 2). Это связано с тем, что многие крупные беспозвоночные (основная пища годовиков окуня) могут успешно прятаться среди друз дрейссены, тем самым становясь менее доступными при питании рыб.
Косвенное подтверждение данного предположения – рост доминирующих по численности и биомассе в составе макрозообентоса экспериментальных мезокосмов личинок Chironomus cingulatus. В обоих вариантах с личинками окуня (ЛО и ЛО + Д) рост средней индивидуальной массы популяции Ch. cingulatus в период с 29 июня по 11 августа 1993 г. фактически не различался, и к концу эксперимента личинки IV-го возраста были на стадии предкуколки (рис. 3). Это можно объяснить тем, что личинкам окуня доступны только младшие возраста (I или II) популяции хирономид, поедая которые, они способствовали росту средней индивидуальной массы личинок Ch. cingulatus. В мезокосмах с годовиками окуня рост средней индивидуальной массы популяции Ch. cingulatus по вариантам существенно различался. В бассейнах, где Dreissena polymorpha отсутствовала (ГО), некоторый рост средней индивидуальной массы наблюдался, но был незначителен. В среднем популяция хирономид к концу эксперимента не достигала состояния предкуколки, так как годовики окуня в первую очередь выедали более крупных личинок IV возраста, тем самым, снижая среднюю индивидуальную массу всей популяции. В бассейнах с дрейссеной личинки очень быстро росли и через три недели достигали состояния предкуколки. Несомненно, что основная причина такого быстрого роста личинок – хорошие трофические условия в мезокосмах с дрейссеной и возможность взрослым личинкам использовать друзы дрейссены в качестве убежищ. О хороших трофических условиях в бассейнах с годовиками окуня и дрейссеной (ГО + Д) свидетельствует повторное отложение кладок популяцией Ch. cingulatus, в результате чего средняя индивидуальная масса личинок к концу эксперимента продолжала снижаться (рис. 3).
Наибольшие значения средней за период проведения эксперимента численности и биомассы макробеспозвоночных были в обоих вариантах с дрейссеной, наименьший аналогичный показатель – в мезокосмах с годовиками окуня и контроле (рис. 1, табл. 2).
Выводы. Из 116 таксонов, обнаруженных в экспериментальных мезокосмах, 61% – обычные для биоценоза Dreissena polymorpha Рыбинского водохранилища, остальные виды вселились за период проведения эксперимента. В мезокосмах с личинками окуня отмечено минимальное количество видов-вселенцев, в вариантах с дрейссеной – максимальное. В результате проведенных исследований показано, что совокупность исследованных факторов (наличие и возраст окуня и дрейссены) воздействует на численность, биомассу и разнообразие как всего макрозообентоса, так и его основных компонентов. В мезокосмах с годовиками окуня без дрейссены биомасса, численность и видовое разнообразие макрозообентоса меньше, чем в варианте с личинками. Это свидетельствует, что в отсутствие дрейссены возрастные группы окуня, потребляющие бентос, снижают его основные структурные характеристики. В бассейнах с окунем и дрейссеной, наоборот, отмечены максимальные значения основных структурных характеристик, что указывает на позитивную средообразующую роль D. polymorpha в бентосных сообществах и возможность использовать ее друзы в качестве убежищ.
Список литературы
Боруцкий Е.В. К вопросу о технике количественного учета донной фауны. К методике обработки количественных проб озерного бентоса. Методика определения сырого веса // Тр. Лимнол. ст. в Косине. 1934. Вып. 17. С. 119–136.
Броцкая В.А., Зенкевич Л.А. Количественный учет донной фауны Баренцева моря // Тр. Всесоюз. НИИ ин-та мор. рыб. хоз-ва и океаногр. 1939. Т. 4. С. 5–98.
Дрейссена Dreissena polymorpha (Pallas) (Bivalvia, Dreissenidae): Систематика, экология, практическое значение. М.: Наука, 1994. 240 с.
Жадин В.И. Моллюски пресных вод СССР. М.; Л.: Изд-во АН СССР, 1952. Вып. 46. 376 с.
Кикнадзе И.И., Шилова А.И., Керкис И.Е. и др. Кариотипы и морфология личинок трибы Chironomini. Атлас. Новосибирск: Наука, 1991. 115 с.
Липина А.Н. Пресные воды и их жизнь. М.: Учпедгиз, 1950. 348 с.
Лукин Е.И. Пиявки. Т. 1: Пиявки пресных и солоноватых водоемов. Фауна СССР. М.: Наука, 1976. Т. 109. 484 с.
Ляхнович В.П., Каратаев А.Ю., Митрахович П.А. Влияние Dreissena polymorpha Pallas на экосистему евтрофного озера // Биология внутренних вод: Информ. бюл. Л., 1983. № 60. С. 25–28.
Методика изучения биогеоценозов внутренних водоемов. М.: Наука, 1975. 254 с.
Определитель пресноводных беспозвоночных Европейской части СССР. Л.: Гидрометеоиздат, 1977. 511 с.
Определитель пресноводных беспозвоночных России и сопредельных территорий. Т. 1: Низшие беспозвоночные. СПб.: Наука, 1994. 439 с.
Определитель пресноводных беспозвоночных России и сопредельных территорий. Т. 2: Ракообразные. СПб.: Наука, 1995. 628 с.
Определитель пресноводных беспозвоночных России и сопредельных территорий. Т. 4: Высшие насекомые, двукрылые. СПб.: Наука, 1999. 998 с.
Определитель пресноводных беспозвоночных России и сопредельных территорий. Т. 6: Моллюски, полихеты, немертины. СПб.: Наука, 2004. 526 с.
Панкратова В.Я. Личинки и куколки комаров подсем. Orthocladiinae фауны СССР (Diptera, Chironomidae). Л.: Наука, 1970. 344 с.
Панкратова В.Я. Личинки и куколки комаров подсем. Podonominae и Tanypodinae фауны СССР (Diptera, Chironomidae). Л.: Наука, 1977. 152 с.
Панкратова В.Я. Личинки и куколки комаров подсемейства Chironomidae фауны СССР (Diptera, Chironomidae). Л.: Наука, 1983. 295 с.
Перова С.Н., Щербина Г.Х. Влияние массовых инвазионных видов на продуктивность макрозообентоса Горьковского водохранилища // Инвазии чужеродных видов в Голарктике. Борок: Изд-во Рыбинский печатный двор, 2003. С. 148–152.
Соколова Н.Ю., Баканов А.И. Методика количественного учета и выявления пространственного распределения бентоса (хирономид) // Методическое пособие по изучению хирономид. Душанбе: Дониш, 1982. С. 3–19.
Чекановская О.В. Водные малощетинковые черви фауны СССР. М.; Л.: АН СССР, 1962. 411 с.
Черновский А.А. Определитель личинок комаров семейства Tendipedidae. М.; Л.: Изд-во АН СССР, 1949. 240 с.
Шилова А.И. Хирономиды Рыбинского водохранилища. Л.: Наука, 1976. 253 с.
Шеннон К. Математическая теория связи // Работы по теории информатики и кибернетики. М.: Ин. лит., 1963. С. 243–332. (Shannon C.E. A Mathematical Theory of Communication (англ.) // Bell System Technical Journal: журнал. 1948. V. 27. P. 379–423.)
Щербина Г.Х. Влияние моллюска Dreissena polymorpha (Pall.) на структуру макрозообентоса экспериментальных мезокосмов // Биология внутр. вод. 2001. № 1. С. 63–70.
Cranston P.S. A key to the Larvae of British Orthocladiinae (Chironomidae) // Freshwater Biol. Ass. 1982. № 45. 152 p.
Limnofauna Europaea. Stuttgart; N.Y.; Amsterdam: Gustav Fischer Verlag; Swets & Zeitlinger B.V., 1978. 532 p.
Mavrin A.S., Strel’nikova A.P. Feeding, development, and growth of juvenile perch perca fluviatilis in mesocosms in the presence of filter-feeding zebra mussel Dreissena polymorpha Pallas // Inland Water Biol. 2011. V. 4. № 2. pp. 232–241.
Wiederholm T. Chironomidae of the Holarctic region // Keys and diagnoses. Part. 1: Larvae. Entomol. Scand. Suppl. 1983. № 19. 451 p.
Wiederholm T. Chironomidae of the Holarctic region // Keys and diagnoses. Part. 2: Pupae. Entomol. Scand. Suppl. 1986. № 28. 482 p.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод