

Биология внутренних вод, 2019, № 2-1, стр. 3-9
Первая находка морского вида Prorocentrum leve Faust, Vandersea, Kibler, Tester & Litaker (Dinophyceae) в континентальном водоеме
Г. Ю. Глущенко 1, *, О. Л. Лужняк 2, К. В. Двадненко 1, М. С. Селина 3
1 Южный научный центр Российской академии наук
344006 д. 41, Ростов-на-Дону, пр. Чехова, Россия
2 Азовский научно-исследовательский институт рыбного хозяйства
344002 Ростов-на-Дону, ул. Береговая, д. 21в, Россия
3 Национальный научный центр морской биологии им. А.В. Жирмунского Дальневосточного отделения
Российской академии наук
690041 Владивосток, ул. Пальчевского, д. 17, Россия
* E-mail: gluschenko_gala@mail.ru
Поступила в редакцию 29.05.2017
После доработки 23.10.2017
Принята к публикации 07.12.2017
Аннотация
Приведены данные о первой находке морской динофитовой водоросли Prorocentrum leve Faust, Vandersea, Kibler, Tester & Litaker в гипергалинном оз. Маныч-Гудило. В результате исследований в мае, июле, октябре 2016 г. и феврале 2017 г. выявлено, что вид встречается в небольших количествах преимущественно в северо-западной части озера. Его морфология, изученная с помощью электронного сканирующего микроскопа, в основном соответствует описанной в литературе. Нахождение морского вида P. leve может служить косвенным доказательством фрагментации ареалов морских водорослей в результате регрессии палеобассейнов. Полученные результаты расширяют сведения о составе альгофлоры внутриконтинентальных водоемов России, а также об экологических возможностях и общем распространении этого вида.
ВВЕДЕНИЕ
Динофитовые водоросли (Dinophyceae) или динофлагелляты – одна из наиболее разнообразных и широко распространенных в Мировом океане групп микроводорослей. К настоящему времени известно 4000 ископаемых и >2500 рецентных динофлагеллят, 90% из которых обитает в морях, остальные – в пресных водах [17]. Однако изучение континентальных гипергалинных водоемов является не менее актуальным, а данные о динофлагеллятах таких водоемов юга европейской части России и сопредельных территорий крайне спорадичны [2, 12, 24]. Наибольший теоретический интерес, особенно в плане флорогенетических и палеогеографических реконструкций, представляют сведения о динофлагеллятах континентальных водоемов Понто-Каспия, исторически связанных с эпиконтинентальными морями – остатками древнего океана Восточный Паратетис. Один из таких водоемов – гипергалинное оз. Маныч-Гудило, расположенное в центральной части Кумо-Манычской депрессии, возникновение которой исторически связано с существованием морских проливов (последний из них – хвалынский пролив) в позднем плейстоцене. Окончательная изоляция водоема от морских бассейнов с развитой речной сетью установилась в позднем голоцене [15]. Фауна и флора водоемов Манычской депрессии крайне неоднородны, что связано не только с последующей их фрагментацией в позднем плейстоцене и голоцене, но и с современной деятельностью человека – постройкой каскада водохранилищ и ирригационной сети каналов, а также интродукцией промысловых видов гидробионтов [7]. Свидетельством существования, по меньшей мере, четырех разнонаправленных морских проливов в Манычской депрессии служат многочисленные ископаемые средиземноморские или каспийские моллюски, бентосные и планктонные фораминиферы [11, 15]. В отличие от многих бентосных организмов, на основе которых ведется палеонтологическая летопись Понто-Каспия, нахождение общих для Азовского и Черного морей микроводорослей морского происхождения в водоемах Манычской депрессии [5] может служить косвенным доказательством фрагментации их ареалов в результате регрессии палеобассейнов.
Один из таких примеров – обнаружение в оз. Маныч-Гудило морской водоросли рода Prorocentrum, по морфологическим признакам полностью соответствующей виду P. leve Faust, Vandersea, Kibler, Tester & Litaker, недавно описанному (как P. levis Faust, Vandersea, Kibler, Tester & Litaker) из Карибского моря [22]. Род Prorocentrum Ehrenberg выделен в 1834 г. [21] и в настоящее время насчитывает ~80 видов [26], его представители – преимущественно морские планктонные или бентосные обитатели, за исключением двух видов, найденных в пресных водоемах [25]. Десять видов рода известны как потенциально токсичные [25, 28]. Для вида Prorocentrum leve, собранного и описанного с плавающего детрита у побережья Белиза в Карибском море, также указана способность продуцировать окадаевую кислоту и динофизис токсин-2 [22]. Позже этот вид обнаружили в северной части Эгейского моря, западной и северо-западной частях Адриатического моря [18, 27, 29] и юго-восточной части Бискайского залива [19], но тесты данных штаммов P. leve не подтвердили наличие у них токсичности [18, 19, 29].
Как показали предыдущие исследования [4, 7, 10, 16], для оз. Маныч-Гудило характерно изменение таксономического состава фитопланктона в зависимости от трансформации его гидрохимического режима. В период наибольшего обводнения, когда уровень минерализации вод не превышал 20 г/л, отмечен лишь один вид динофитовой водоросли Gymnodinium sp. [4]. С последующим повышением минерализации озера состав динофитовых увеличился до семи видов, характерных преимущественно для морских водоемов [5–7, 16].
В оз. Маныч-Гудило Prorocentrum leve впервые найден в 2016 г. в фиксированных пробах в ходе комплексных мониторинговых исследований водоемов Манычской депрессии.
Цель работы – описание морфологии Prorocentrum leve в условиях гипергалинного континентального водоема (с использованием методов световой и сканирующей электронной микроскопии) и его встречаемости на данной акватории.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Физико-географическая характеристика и гидрохимический режим водоема. Озеро Маныч-Гудило относится к внутриконтинентальному мелководному (средняя глубина 2.6 м) гипергалинному водоему, расположенному в зоне неустойчивого и недостаточного увлажнения умеренного климатического пояса [7, 14]. Характерные для региона небольшое количество выпадающих осадков, высокие летние температуры и продолжительные суховеи приводят к сильному испарению влаги, что влечет за собой еще большее повышение солености озера. Минерализация данного водоема также зависит от объема пресной воды питающих его рек – Большой Егорлык и Калаус [7, 9, 14]. В настоящее время общая минерализация оз. Маныч-Гудило колеблется от 5.2 (в местах впадения рек) до ≥50 г/л. Воды озера относят к мезо- и полигалинным, а по соотношению основных солесодержащих ионов они занимают промежуточное положение между карбонатно-сульфатно-кальциевым типом вод континентального засоления и хлоридно-натриевым типом морских вод [9, 14].
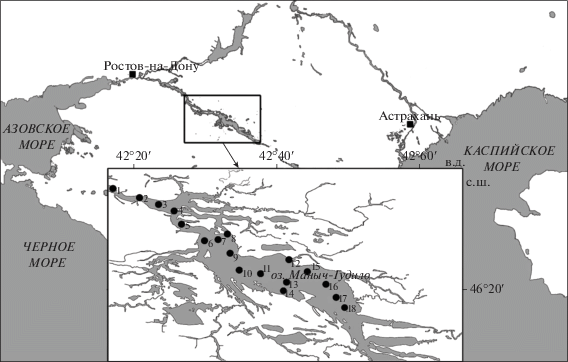
Методы исследования. В ходе экспедиционных исследований на водоемах Манычской системы сотрудниками Южного научного центра РАН в 2016 г. отобраны пробы воды в мае на 5 станциях, в июле ‒ на 3, в октябре ‒ на 13 станциях (с поверхностного горизонта). Кроме того, в феврале 2017 г. на двух станциях взяты подледные пробы воды и остатки водной растительности, осевшие на дно (рис. 1).
Рис. 1.
Карта-схема водоемов Кумо-Манычской депрессии. В выноске – местонахождения станций отбора проб воды на оз. Маныч-Гудило: в 2016 г. – май (ст. № 1–5), июль (ст. № 6–8), октябрь (ст. № 6–18) и в 2017 г. ‒ февраль (№ 8, 12).
Для изучения морфологических особенностей организмов использовали световую микроскопию (микроскоп “Микмед-5”), эпилюминесцентную (флуоресцентный микроскоп “Микмед-2-Вар-11”) и сканирующую электронную микроскопию (СЭМ EVO-40 XVP). Пробы воды фиксировали кислым раствором Люголя и 25%-ным глютаровым альдегидом с конечной концентрацией в пробе 1% [13].
Для электронной микроскопии клетки водорослей осаждали на мембранные ядерные фильтры с диаметром пор 1 мкм, дважды промывали дистиллированной водой, дегидратировали через серию растворов этилового спирта восходящей концентрации (25, 40, 55, 70 и 96%) и высушивали на воздухе при комнатной температуре. Затем фильтры прикрепляли с помощью двусторонней проводящей углеродной ленты к столикам. Напыление поверхности образцов проводили с помощью Au/Pd-мишени в установке вакуумного напыления Mini Sputter Coater SC7620 (QuorumTechnologies).
Клетки (24 экз.) и их детали измеряли по электронным микрофотографиям, используя программное обеспечение Scandium (версия 5.0), а также с помощью светового микроскопа и видеоокуляра ToupСam UCMOS 10 MP. В качестве показателя варьирования признаков приведено стандартное отклонение. Для обозначения текальных пластин микроводоросли применяли терминологию, предложенную М. Хоппенрат с соавт. [25].
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Морфология Prorocentrum leve. Клетки широкоовальной формы, симметричные, вогнутые/двояковогнутые со стороны пояска (рис. 2а–2ж). Длина клеток 43.74–50.37 мкм (47.99 ± 1.52 мкм), ширина – 39.73–45.31 мкм (42.85 ± 1.38 мкм), отношение длины к ширине – 1.05–1.18 (1.12 ± 0.03). Клетки содержат хлоропласты, в центре располагается крупный, хорошо заметный пиреноид, овальное ядро находится в нижней части клетки (рис. 2б, 2в). Клетка покрыта двумя большими пластинками, которые соединяются сагиттальным швом (рис. 2и). На апикальном конце левой пластинки видна небольшая вмятина (рис. 2д), а на правой располагается узкое V-образное углубление, 5.56 мкм шириной и 4.01 мкм глубиной (рис. 2г–2ж). Это прижгутиковая (periflagellar) зона, в которой находятся жгутиковая и апикальная поры, а также ряд маленьких пластинок (рис. 2к–2м). Большие текальные пластинки гладкие, с разбросанными по ним небольшими углублениями диаметром 0.45–0.93 мкм (0.64 ± 0.12 мкм, n = 48), в которых располагаются два типа пор: преобладающие большие, диаметром 0.12–0.21 мкм (0.16 ± 0.03, n = 59), и маленькие, диаметром 0.06–0.11 мкм (0.09 ± 0.02, n = 13) (рис. 2з). Углубления с порами разбросаны почти по всей поверхности больших пластинок, более плотно располагаются на краях и в большинстве случаев отсутствуют в их центральной части. Количество пор на пластинке 203–310 (267 ± 34.9), в краевой части зоне 134–164 (151 ± 9.2, n = 7). Вставочный поясок гладкий (рис. 2и). В верхней части прижгутиковой зоны размещаются четыре маленькие пластинки, из них вторая – самая маленькая (рис. 2к–2м). Под ними располагаются большая овальная жгутиковая и маленькая узкая апикальная поры (рис. 2к–2м).
Рис. 2.
Световые (а–в) и электронные (г–м) микрофотографии Prorocentrum leve: а, б – вид разных клеток с правой стороны (фиксация глютаром (а), раствором Люголя (б)); в – вид клетки с левой стороны (люминесцентный микроскоп); г – вид с правой стороны; д – вид с левой стороны; е, ж – вид клеток сбоку; з – поры; и – вид вставочного пояска; к–м – прижгутиковая зона. ап – апикальная пора, жг – жгутиковая пора, п – пиреноид, я – ядро.

Встречаемость водоросли Prorocentrum leve в оз. Маныч-Гудило. При исследовании фитопланктона в мае 2016 г. (ст. 1–5) (рис. 1) клетки Prorocentrum leve не обнаружены. В июле пробы планктона отбирали в северо-западной части озера (ст. 6–8) при сильном волнении, P. leve присутствовал на этих станциях при численности 1300–3826 кл./л (2–4% общей численности фитопланктона). Температура воды составляла 25.3–26.8°С, а минерализация – 44 г/л. В планктоне доминировали мелкие зеленые водоросли (83–94% общей численности фитопланктона). На долю динофитовых и диатомовых водорослей приходилось до 11 и 6% соответственно. Наряду с P. leve найдены другие морские виды динофлагеллят – Heterocapsa rotundata (Lohmann) Hansen и Akashiwo sanguinea (Hirasaka) Hansen et Moestrup.
В октябре была исследована почти вся акватория озера (ст. 6–18). Однако P. leve, как и в июле, встречен лишь на ст. 6–8 (северо-западная часть озера). Численность вида достигала 1111–1342 кл./л, доминировал вид Akashiwo sanguinea. Температура поверхности воды в этот период достигала 7–10°С, минерализация – 38–44 г/л.
В феврале 2017 г. при температуре 0.2–0.3°C и минерализации 6–38 г/л Prorocentrum leve в планктоне отсутствовал, и лишь несколько неподвижных его клеток co сжатым содержимым были отмечены на остатках нитчатых водорослей.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Морфология P. leve из оз. Маныч-Гудило по форме клеток, отсутствию орнаментации створок и строению прижгутиковой зоны совпадает с первоописанием вида [22]. По мнению М. Фауст с соавт. [22], эти признаки отличают P. leve oт других видов рода Prorocentrum. Клетки из озера крупнее клеток из Карибского и Эгейского морей, но входят в размерный диапазон, указанный для культуры P. leve из Бискайского залива (табл. 1). В отличие от клеток из морского прибрежья, экземпляры из озера имели большее количество пор на больших пластинках и в их краевой зоне (табл. 1). Как и в культурах P. leve из Бискайского залива, в исследованных экземплярах на больших пластинках было отмечено два типа пор: большие и маленькие. Диаметр больших пор совпадал с приведенным в первоописании вида (табл. 1). Кроме того, диаметр углублений, в которых располагались поры, заметно превышал таковой для клеток из Бискайского залива (табл. 1). Имеющиеся морфологические различия, возможно, связаны с процессом приспособления организма к необычной для него среде, с высокой в равной степени хлоридной и сульфатной соленостью.
Таблица 1.
Сравнительная морфологическая характеристика Prorocentrum leve
| Признак | Карибское море, прибрежье Белиза [22] | Северная часть Эгейского моря [18] | Бискайский залив [19] |
оз. Маныч-Гудило (данные авторов) |
|---|---|---|---|---|
| Форма клетки | Овальная | Овальная | Овальная | Широкоовальная |
| Длина клетки, мкм | 40−44 | 38−49 | 37−51 (45 ± 4) | 44−50 (48 ± 2) |
| Ширина клетки, мкм | 37−40 | 33−37 | 30−42 (36 ± 3) | 40−45 (43 ± 1) |
| Отношение длины к ширине | − | 1.1–1.3 | 1.3* | 1.1–1.2 |
| Количество пор: | ||||
| на пластинке | 221−238 | − | 211−222 | 203−310 |
| краевых | 99−130 | − | 105 | 134−164 |
| Диаметр углублений, мкм | − | − | 0.27−0.59 | 0.45−0.93 |
| Диаметр пор, мкм: | ||||
| больших | 0.13−0.19 | – | 0.12−0.19 | 0.12−0.21 |
| маленьких | – | – | 0.05−0.10 | 0.06−0.11 |
| Биотоп | Плавающий детрит | Макрофиты, водная толща, осадки | Макрофиты | Водная толща, макрофиты |
До настоящего времени P. leve отмечен исключительно в морях тропической [22], субтропической (северная часть Средиземного моря [18, 27, 29]) и умеренной (Атлантика, у побережья Пиренейского п-ва в Бискайском заливе [19]) зон. Это первая находка P. leve в континентальном водоеме, который к тому же характеризуется высокой минерализацией. Помимо этого вида в озере зарегистрированы представители других родов морских динофлагеллят – Akashiwo Hansen & Moestrup, 2000, Heterocapsa Stein, 1883 и Gyrodinium Kofoid & Swezy, 1921. Виды этих родов встречались в озере и ранее при минерализации водоема ≤30 г/л [7, 16]. В современных условиях, при содержании в воде солей ≥50 г/л, среди динофитовых водорослей обнаружены морские виды Glenodinium pilula (Ostenfeld) Schiller, Scrippsiella acuminata (Ehrenberg) Kretschmann et al., Prorocentrum scutellum Schröder, Heterocapsa rotundata (Lohmann) Hansen [5, 6].
Динофлагеллята Prorocentrum leve ранее не была отмечена в водоемах России. Однако еще в 30-х годах прошлого столетия из заливов Северо-Восточного Каспия с соленостью вод 31–53‰ И.А. Киселевым [1] была описана динофлагеллята Еxuviaella caspica Kisselew. Позже этот вид указывали для восточной части залива Сиваш Азовского моря, водоемов Украины и озер равнинной Грузии [8]. В середине 1970-х гг. род Exuviaella Cienkowski, 1881 был упразднен, а вид сведен в синонимы к Prorocentrum lima (Ehrenberg) Stein [20]. А.Ф. Крахмальный [3] посчитал этот вид самостоятельным таксоном и предложил новую номенклатурную комбинацию – Prorocentrum caspica (Kisselew) Krachmalny, но в список современных свободноживущих динофлагеллят P. caspica не был включен [23]. В действительности форма и размеры Exuviaella caspica совпадают с таковыми у Prorocentrum leve. Однако широкоовальная форма и пиреноид в центре клетки имеются не только у P. leve, но еще у нескольких видов бентосных динофлагеллят, в том числе у P. lima, характеризующегося широкой вариабельностью [25]. Описание орнаментации клетки Exuviaella caspica как “гладкой или пунктированной” [1, c. 112] позволяет предположить, что в Каспийском море обитают два морфологически сходных вида бентосных Prorocentrum. Необходимо провести верификацию вида из типового местонахождения, описанного как Exuviaella caspica, с применением современных методов исследования.
Трудно судить, как и когда Prorocentrum leve попал в оз. Маныч-Гудило. Возможно, что это автохтонный вид с широкой толерантностью к колебаниям солености, которые вид успешно переживал в виде цист. В предыдущих исследованиях планктона P. leve не отмечен [4–7, 10, 16], вероятно, потому что ведет преимущественно эпифитный образ жизни [18, 22], а исследований микроводорослей в обрастаниях здесь не проводили. Впервые вид обнаружен нами во время сильного волнения в озере, в результате которого эпифитные организмы могли быть вынесены в толщу воды. Остаются невыясненными вопросы, какой биотоп в данном водоеме предпочитает Prorocentrum leve, каковы его сезонная динамика, распространение в озере, а также присуще ли местной популяции способность продуцировать токсин.
Выводы. Впервые во внутреннем водоеме России, гипергалинном оз. Маныч-Гудило, зарегистрирована динофитовая морская водоросль Prorocentrum leve. Морфологические характеристики вида в целом соответствуют первоописанию, но имеется ряд структурных особенностей. Массового развития в озере P. leve не достигал, отмечен только в северо-западной части водоема в вегетативном состоянии в июле и октябре в планктоне и в переходном состоянии к цистам в феврале на водорослевых матах. Находка этого вида в оз. Маныч-Гудило может служить косвенным доказательством фрагментации ареалов водорослей морского происхождения в результате регрессии палеобассейнов и существенно расширяет знания о составе альгофлоры внутриконтинентальных водоемов России, а также об экологии и распространении этого морского вида.
Список литературы
Киселев И.А. Фитопланктон Северо-восточного Каспия с его заливами Комсомолец (Мертвый Култук) и Кайдак по материалам экспедиций Академии паук СССР в 1934 и 1935 гг. // Тр. по комплексному изучению Каспийского моря. 1940. Вып. 3. С. 103–125.
Крахмальный А.Ф. Новые таксономические и номенклатурные комбинации в Dinophyta // Альгология. 1993. Т. 3. № 4. С. 87–88.
Крахмальный А.Ф. Динофитовые водоросли Украины. Киев: Альтерпресс, 2011. 444 с.
Круглова В.М. Пролетарское водохранилище. Ростов-на-Дону: Ростов. гос. ун-т, 1972. 180 с.
Лужняк О.Л. Обнаружение морских динофитовых водорослей в озере Маныч-Гудило // Естественные и инвазийные процессы формирования биоразнообразия водных и наземных экосистем: Тез. междунар. конф. Ростов-на-Дону: Юж. науч. центр РАН, 2007. С. 196–197.
Лужняк О.Л., Глущенко Г.Ю. Сезонные изменения фитопланктона водоемов Кумо-Манычской впадины в 2010 г. // Актуальные проблемы обеспечения продовольственной безопасности юга России: Матер. междунар. конф. Ростов-на-Дону: Юж. науч. центр РАН, 2011. С. 73–75.
Маныч-Чограй: история и современность. Ростов-на-Дону: Эверест, 2005. 152 с.
Матвиенко О.М., Литвиненко Р.М. Визначник прiсноводних водоростей Украiнськоi РСР. III. Ч. 2. Пiрофiтовi водоростi – Pyrrophytа. Киев: Наук. думка, 1977. 387 с.
Матишов Д.Г., Орлова Т.А., Гаргопа Ю.М., Павельская Е.В. Многолетняя изменчивость гидрохимического режима водной системы Маныч-Чограй // Вод. ресурсы. 2007. Т. 34. № 5. С. 560–564.
Михайловская З.Н. Динамика фитопланктона осолоненных манычских водоемов // Науч. конф., посвящ. 80-летию Ростов. ун-та: Тез. докл. Вып. 1. Ростов-на-Дону: Росиздат, 1949. С. 128–129.
Набоженко М.В., Булышева Н.И., Шохин И.В. Современное состояние макрозообентоса водоемов Кумо-Манычской депрессии в летний период // Современное состояние и технологии мониторинга аридных и семиаридных экосистем Юга России. Ростов-на-Дону: Юж. науч. центр РАН, 2010. С. 113–118.
Прошкина-Лавренко А.И., Макарова И.В. Водоросли планктона Каспийского моря. Л.: Наука, 1968. 291 с.
Радченко И.Г., Капков В.И., Федоров В.Д. Практическое руководство по сбору и анализу проб морского фитопланктона: Учебно-методическое пособие для студентов биол. специальностей университетов. М.: Мордвинцев, 2010. 60 с.
Румянцев В.А., Драбкова В.Г., Измайлова А.В. Озера европейской части России. СПб.: Лема, 2015. 392 с.
Свиточ А.А., Янина Т.А., Новикова Н.Г. и др. Плейстоцен Маныча (вопросы строения и развития). М.: Географ. факультет МГУ, 2010. 136 с.
Фуштей Т.В. Весенний фитопланктон озера Маныч-Гудило // Тр. заповедника Ростовский. Вып. 2. Ростов-на-Дону: Северо-Кавказ. науч. центр высшей школы, 2002. С. 81–88.
Хаусман К., Хюльсман Н., Радек Р. Протистология. М.: Товарищество науч. изданий КМК, 2010. 495 с.
Aligazaki K., Nikolaidis G., Katikou P. et al. Potentially toxic epiphytic Prorocentrum (Dinophyceae) species in Greek coastal waters // Harmful Algae. 2009. V. 8. № 2. P. 299–311.
David H., Laza-Martinez A., Garcia-Etxebarria K., Riobó P. Characterization of Prorocentrum elegans and Prorocentrum levis (Dinophyceae) from the southeastern Bay of Biscay by morphology and molecular phylogeny // J. Phycol. 2014. V. 50. № 4. P. 718–726.
Dodge J.D. The Prorocentrales (Dinophyceae). II. Revision of the taxonomy within the genus Prorocentrum // Bot. J. Linn. Soc. 1975. V. 71. № 2. P. 103–125.
Ehrenberg C.G. Dritter Beitrag zur Erkenntniss grosser Organisation in der Richtung des kleinsten Raumes // Abhandlungen der Königlichen Akademie der Wissenschaften zu Berlin. Berlin: Druckerei der Königlichen Akademie der Wissenschaften, 1835. P. 145–336.
Faust M.A., Vandersea M.W., Kibler S.R. et al. Prorocentrum levis, a new benthic species (Dinophyceae) from a mangrove island, Twin Cays, Belize // J. Phycol. 2008. V. 44. P. 232–240.
Gómez F. A checklist and classification of living Dinoflagellates (Dinoflagellata, Alveolata) // CICIMAR Oceánides. 2012. V. 27. № 1. P. 65–140.
Hammer U.T. Saline Lake Ecosystems of the World. Monographiae biologicae. V. 59. Dordrecht: Junk Publ., 1986. 619 p.
Hoppenrath M., Chomérat N., Horiguchi T. et al. Taxonomy and phylogeny of the benthic Prorocentrum species (Dinophyceae) – A proposal and review // Harmful Algae. 2013. V. 27. P. 1–28.
Hoppenrath M., Murray S.A., Chomérat N., Horiguchi T. Marine benthic dinoflagellates – unveiling their worldwide biodiversity // Kleine Senckenberg-Reihe. 2014. Bd 54. 276 p.
Ingarao C., Lanciani1 G., Iorio D.Di. et al. Prorocentrum levis and Coolia monotis in Abruzzo coastal waters (W. Adriatic) // Harmful algae news. 2010. № 41. P. 12–14.
Kameneva P.A., Efimova K.V., Rybin V.G., Orlova T.Yu. Detection of dinophysistoxin-1 in clonal culture of marine dinoflagellate Prorocentrum foraminosum (Faust M.A., 1993) from the Sea of Japan // Toxins. 2015. V. 7. № 10. P. 3947–3959.
Pistocchi R., Guerrini F., Pezzolesi L. et al. Toxin Levels and Profiles in Microalgae from the North-Western Adriatic Sea – 15 Years of Studies on Cultured Species // Mar. Drugs. 2012. V. 10. P. 140–162.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод