

Биология внутренних вод, 2019, № 2-2, стр. 3-11
Разнообразие и особенности пространственной структуры водорослевых сообществ водоемов и водотоков устья р. Лены
В. А. Габышев 1, П. М. Царенко 2, А. П. Иванова 1, *
1 Институт биологических проблем криолитозоны Сибирского отделения Российской академии наук
677980 Якутск, пр. Ленина, д. 41, Россия
2 Институт ботаники им. Н.Г. Холодного Национальной академии наук Украины
01601 Киев, МСП-1, ул. Терещенковская, д. 2, Украина
* E-mail: a.p.ivanova@rambler.ru
Поступила в редакцию 24.01.2018
После доработки 06.09.2018
Принята к публикации 27.11.2018
Аннотация
Впервые обобщены сведения об альгофлоре водоемов и водотоков устья р. Лены. В результате ревизии полученных данных в соответствии с современными таксономическими сводками выявлен видовой список водорослей (646 видов). Установлено, что таксономическая структура водорослей характеризует исследованные водоемы как арктические ненарушенные. Проведенный анализ показал, что крупная транзитная р. Лена, протекающая в меридиональном направлении, способствует переносу бореальной флоры в арктический регион. В зоне смешения морских и пресных вод выявлены признаки краевого эффекта. Установлено влияние морской флоры на все исследованные водные объекты низовья р. Лены (занос морских и солоновато-водных видов водорослей).
ВВЕДЕНИЕ
В устьевой части р. Лена образует дельту, площадью 30 тыс. км2, третью среди крупнейших речных дельт мира и первую в России по площади. Низовье р. Лены и прибрежные участки моря Лаптевых находятся под охраной Усть-Ленского заповедника и крупнейшего российского биосферного резервата Лена-Дельта площадью >60 тыс. км2. Водоемы региона расположены за Полярным кругом, севернее 71° с.ш., в зоне сплошного залегания вечномерзлых грунтов, где в отсутствие дренажа распространены эфемерные водоемы: ручьи, мочажины. Регион богат болотами и озерами, характерными для арктической тундры. В низовье р. Лены, как и в низовьях других больших арктических рек, образуются крупные области смешения пресных и соленых вод, где могут формироваться так называемые маргинальные зоны. В последнее время отмечено повышение активности промышленного освоения региона. В 2017 г. компанией Роснефть начата сейсморазведка на шельфе моря Лаптевых, следовательно, усилилась антропогенная нагрузка. В этих условиях возрастает актуальность поддержания биоразнообразия водных экосистем региона, особенно в маргинальной зоне, где весьма существенна роль экотонов, доказанная рядом исследователей [44, 47].
Данные о водорослях водоемов устьевой части р. Лены разрознены. Депонированная рукопись И.И. Васильевой и П.А. Ремигайло содержит наиболее полный видовой список, сравнительный флористический анализ водорослей и сведения о сапробиологическом состоянии водоемов региона [5]. Некоторые данные о видовом составе водорослей региона приводятся в отчетах Тиксинского территориального управления по гидрометеорологии и мониторингу окружающей среды [27, 28], а также в работах по водорослям моря Лаптевых [13, 15, 18, 26, 34, 35, 41]. Имеющаяся информация касается, главным образом, планктона и в незначительной степени выжимок из мха. Несмотря на длительную историю альгологических исследований, генерализация и анализ накопленных сведений отсутствуют.
Цель работы – обобщение и таксономическая ревизия всех имеющихся альгологических данных, изучение особенностей таксономического состава и пространственной структуры водорослевых сообществ водоемов и водотоков устьевой части р. Лены.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Материалом послужили пробы фитопланктона, собранные в сентябре 2009 г. в р. Лене (район о. Тит-Ары); в августе-сентябре 2014 г. ‒ в бухте Тикси и заливе Неелово моря Лаптевых, а также в озерах побережья бухты Тикси, залива Неелово и в протоке дельты Оленёкская.
Пробы фитопланктона отбирали с помощью планктонной сети Апштейна (газ SEFAR NITEX, размер ячеи 30 мкм) в литорали и пелагиали водоемов с поверхностного горизонта воды (0–0.3 м). Всего было собрано 100 планктонных проб, их обработку проводили в лаборатории флористики и геоботаники Института биологических проблем криолитозоны (ИБПК) СО РАН. Для идентификации диатомей было изготовлено 70 постоянных препаратов путем прокаливания створок и помещения их в синтетическую смолу BIO MOUNT. Микроскопирование препаратов выполняли с помощью микроскопов “Laboval” и “Olympus BH-2” по общепринятым методикам. Использованы ранее неопубликованные данные из внутренних отчетов ИБПК СО РАН по материалам сборов на участках дельты в районе урочищ Хохочу, Чай-Тумус и Буор-Хая, в протоке Гусинка и в устье р. Тыллах в сентябре 1994 г. Также приведены фондовые материалы Усть-Ленского заповедника в форме видовых списков [23, 24], полученных при обработке сборов, выполненных в бухте Тикси и в р. Лене (район о. Столб) в 1980–1990 гг. Были использованы системы для таксономической ревизии [37], для отдела Dinophyta [22], для отдела Chrysophyta [9]. Таксономическое положение отдельных видов, а также принадлежность водорослей к морской флоре, уточняли на интернет-портале Algabase.org. Эколого-географические характеристики водорослей взяты из работы С.С. Баринова и др. [2]. Для оценки флористического сходства использовали коэффициент Серенсена [48].
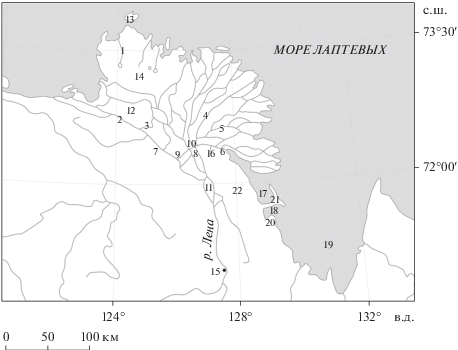
Исследованная акватория условно была разделена на пять участков в соответствии с их положением на местности и особенностями гидрологии и морфометрии водных объектов (рис. 1). Преддельтовый участок включал отрезок р. Лены от пос. Чекуровка до о. Столб. Дельтовый участок объединял основные протоки (Оленёкская, Большая Трофимовская, Сардахская, Арангастах-Юряге, Быковская, Гусинка), а также устье р. Тыллах и р. Лену возле урочища Чай-Тумус. Взморье представляли прибрежные участки моря Лаптевых (залив Неелово, бухта Тикси, губа Буор-Хая) и опресненная часть, примыкающая к авандельте. В отдельную группу были объединены девять озер ледникового, водно-эрозионного, эрозионно-термокарстового и речного происхождения, расположенные по берегам бухты Тикси, залива Неелово и Оленёкской протоки. Пробы из выжимок мхов, тундроболот, горных ручьев, мочажин берегов бухты Тикси, мыса Косистый, п-ова Быковский, отрогов Хараулахского хребта, островов Тит-Ары, Харданг-Сисе, Дунай, Арга-Муора-Сисэ были объединены в группу “заболоченные участки”.
Рис. 1.
Карта-схема района работ. Протоки дельты р. Лены: 1 – Арангастах-Юряге, 2 – Оленёкская, 3 – Гусинка, 4 – Большая Трофимовская, 5 – Сардахская, 6 – Быковская. Урочища: 7 – Чай-Тумус, 8 – Хохочу, 9 – Буор-Хая. Острова: 10 – Столб, 11 – Тит-Ары, 12 – Харданг-Сисе, 13 – Дунай, 14 – Арга-Муора-Сисэ. Прочие географические объекты: 15 – пос. Чекуровка, 16 – устье р. Тыллах, 17 – залив Неелово, 18 – бухта Тикси, 19 – губа Буор-Хая, 20 – мыс Косистый, 21 – п-ов Быковский, 22 – Хараулахский хребет.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
В результате обобщения собственных и литературных данных в водоемах дельты р. Лены выявлено 646 видов водорослей (698 таксонов, рангом ниже рода, включая номенклатурный тип вида) из 11 отделов, 18 классов, 42 порядков, 95 семейств, 234 родов (табл. 1). Вид Bicosoeca borealis Hilliard, обнаруженный единично в оз. Ладаннах, расположенном на берегу залива Неелово, был исключен из списка, так как, по современным данным, он относится к царству Stramenopilae. Наиболее богат по числу видов был отдел диатомовых (273 вида), за ним следовали зеленые (116), стрептофитовые (95) и цианопрокариоты (81); меньшую роль играли представители желто-зеленых (22), золотистых и динофитовых (по 19) и эвгленовых (13). Виды из отделов Eustigmatophyta (4), Cryptophyta (3) и Rhodophyta (1) были встречены единично.
Таблица 1.
Систематический состав альгофлоры водоемов низовья р. Лены
| Отдел | Класс | Порядок | Семейство | Род | Вид | Вид + + разновидность | Доля видов, % общего количества | Новые виды для флоры Якутии |
|---|---|---|---|---|---|---|---|---|
| Cyanoprokaryota | 1 | 6 | 17 | 33 | 81 | 83 | 12.5 | 3 |
| Euglenophyta | 1 | 2 | 2 | 6 | 13 | 13 | 2.0 | – |
| Chrysophyta | 2 | 2 | 5 | 11 | 19 | 21 | 2.9 | 5 |
| Xanthophyta | 1 | 3 | 7 | 10 | 22 | 22 | 3.4 | – |
| Eustigmatophyta | 1 | 1 | 2 | 3 | 4 | 4 | 0.6 | – |
| Bacillariophyta | 3 | 8 | 22 | 71 | 273 | 296 | 42.3 | 12 |
| Dinophyta | 1 | 4 | 8 | 10 | 19 | 20 | 2.9 | 2 |
| Cryptophyta | 1 | 1 | 1 | 1 | 3 | 3 | 0.5 | – |
| Rhodopyta | 1 | 1 | 1 | 1 | 1 | 1 | 0.2 | – |
| Chlorophyta | 4 | 11 | 23 | 70 | 116 | 124 | 18.0 | 10 |
| Streptophyta | 2 | 3 | 7 | 18 | 95 | 111 | 14.7 | 3 |
| Всего | 18 | 42 | 95 | 234 | 646 | 698 | 100.0 | 35 |
На уровне классов по видовому богатству выделялись Bacillariophyceae (36.5% видового состава), Zygnematophyceae (14.6%), Cyanophyceae и Chlorophyceae (по 12.5%); на уровне порядков – Raphales (31.4%), Desmidiales (13.8%) и Sphaeropleales (9.3%). К наиболее крупным семействам по числу видов относились Naviculaceae (12.1% общего числа видов) и Desmidiaceae (10.5%). К десяти родам из головной части списка принадлежали Cosmarium (27 видов), Closterium и Navicula (по 19), Eunotia, Pinnularia и Nitzschia (по 18), Gomphonema (15), Surirella (13), Cymbella (11), Staurastrum (10). Одно- и двувидовые роды составляли 70.9% общего числа родов и включали 32.8% видового состава. Пропорции флоры (относительное число семейств, принятое за 1 : среднее число родов в семействе : среднее число видов в семействе : среднее число внутривидовых таксонов в семействе, включая номенклатурный тип вида) – 1 : 2.5 : 6.8 : 7.3. Родовая насыщенность видами – 2.8. Отношение числа видов и разновидностей к числу таксонов видового ранга (вариабельность вида) – 1.1.
Выявлено было 35 новых для альгофлоры Якутии видов и разновидностей из шести отделов [6, 13, 15, 18, 26–28, 34, 35, 41]. Среди последних наиболее богаты видами диатомовые и зеленые водоросли, в меньшей степени – золотистые, стрептофитовые, цианопрокариоты и динофитовые (см. табл. 1).
Во флоре исследованного региона было найдено 47 морских видов, включая диатомовые (35 видов), цианопрокариоты (2), динофитовые (9) и эвгленовые (1). Часть морских видов – новые для водоемов (водных объектов) Якутии представители диатомовых водорослей (7 видов) и цианопрокариоты (2). Наибольшее число морских видов было встречено во флоре взморья (35). Другие водные объекты низовья р. Лены в разной степени испытывают влияние солоновато-водной флоры. Так, на преддельтовом участке было отмечено семь морских видов, в дельте и озерах – по четыре и на заболоченных участках – два вида. Очевидно, морские виды водорослей легко могут быть занесены в дельту реки из взморья в результате приливов и ветрового нагона. Небольшое число таких видов во флоре дельтового участка может свидетельствовать о ее недостаточной изученности.
Выделенные участки исследованного региона флористически существенно различаются. Коэффициент сходства видового состава для большинства участков низкий и варьирует в пределах 0.17–0.29. Сходство альгофлоры пар участков преддельтовый–дельта (0.35) и дельта–взморье (0.31) несколько выше из-за их смежного расположения. Наиболее высокое сходство альгофлоры получено для преддельтового участка и взморья (0.57), что оказалось неожиданным, учитывая различие условий, в которых развиваются водоросли преддельтового участка р. Лены и прибрежья моря Лаптевых. Для уточнения этих данных требуется более длительное изучение альгофлоры региона. Уровень видового разнообразия также значительно различается по участкам. Наиболее богаты в видовом отношении флоры взморья и преддельтового участка (рис. 2), флоры заболоченных участков и озер уступают им в 1.5–2 раза, наименее разнообразна флора дельты р. Лены.
Рис. 2.
Общее число видов (на правой оси ординат) и доля ведущих отделов водорослей, % (левая ось ординат) во флоре различных участков низовья р. Лены. I – озера, II – преддельтовый участок, III – дельта, IV – взморье (прибрежье моря Лаптевых), V – заболоченные участки. Отделы водорослей: 1 – Cyanoprokaryota, 2 – Euglenophyta, 3 – Chrysophyta, 4 – Xanthophyta, 5 – Bacillariophyta, 6 – Dinophyta, 7 – Chlorophyta, 8 – Streptophyta, 9 – другие. Сплошная кривая линия ‒ общее число видов.
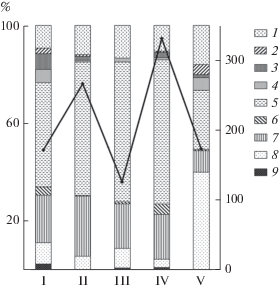
Различия между участками отмечены и в структуре флоры. На уровне отделов флористически наиболее обособлены заболоченные участки (рис. 2). Стрептофитовые водоросли здесь – самая многочисленная по числу видов группа. Доли диатомей и зеленых водорослей во флоре значительны, но существенно меньше, чем на других участках. Большое значение имеют цианопрокариоты и желто-зеленые водоросли, а эвгленовые представлены наиболее полно, по сравнению с другими участками.
В структуре флоры озер, наряду с характерным для всей устьевой области реки доминированием диатомовых и зеленых и существенном вкладе цианопрокариот и стрептофитовых водорослей, отмечается повышенное разнообразие золотистых, желто-зеленых и динофитовых (см. рис. 2). Только в озерах встречались представители отдела Eustigmatophyta.
Преддельтовый участок, дельта и взморье сходны между собой по структуре флоры, что выражается в доминировании диатомей и значительном вкладе зеленых водорослей и цианопрокариот (рис. 2). Особенности флоры взморья заключаются в относительно высоком разнообразии золотистых. Здесь динофитовые водоросли достигают максимального разнообразия среди исследованных водоемов, а обнаруженные криптофитовые водоросли более нигде в низовье р. Лены не встречались. В исследованных водоемах найден единственный представитель красных водорослей, который был отмечен в дельте р. Лены.
На уровне классов во флоре большинства участков лидировал Bacillariophyceae, вторую и третью позицию занимали Chlorophyceae и Cyanophyceae. Видами во флоре заболоченных участков наиболее разнообразно был представлен класс Zygnematophyceae.
На уровне порядков флора заболоченных участков также обособлена, наиболее разнообразны в ней представители Desmidiales, на втором месте по числу видов – Raphales. На других участках список ведущих порядков возглавляют шовные диатомеи.
Ведущее семейство для большинства участков – Naviculaceae. Лишь во флоре заболоченных участков на первом и втором местах находятся Desmidiaceae и Closteriaceae. Это наблюдается и на уровне родов, во флоре болот среди 11 ведущих родов 6 – представители десмидиевых и клостериевых (Cosmarium, Closterium, Euastrum, Staurastrum, Cosmoastrum и Staurodesmus) (табл. 2). Роды из сем. Desmidiaceae (Cosmarium и Staurastrum) входят в число ведущих еще только на одном участке – в озерах. Родовой спектр озер отличается от остальных участков исследованной акватории наличием представителей эвгленовых и золотистых водорослей (Kephyrion и Trachelomonas).
Таблица 2.
Ведущие по числу видов роды альгофлор различных участков низовья р. Лены
| Род | Озера | Преддельтовый участок | Дельта | Взморье | Заболоченные участки |
|---|---|---|---|---|---|
| Aulacoseira | 6 | 8 | 6 | 8 | 4 |
| Characiopsis | – | – | – | – | 4 |
| Closterium | – | 8 | 6 | 6 | 13 |
| Cosmarium | 7 | – | – | – | 21 |
| Cosmoastrum | – | – | – | – | 5 |
| Cymbella | – | 7 | 3 | 5 | – |
| Desmodesmus | – | – | 3 | 6 | – |
| Diatoma | – | 5 | – | 6 | – |
| Dictyosphaerium | 3 | – | 3 | – | – |
| Diploneis | – | – | – | 5 | – |
| Dolichospermum | – | 5 | 4 | 8 | – |
| Euastrum | – | – | – | – | 9 |
| Eunotia | 6 | 7 | – | – | 8 |
| Fragilaria | – | – | 3 | – | – |
| Gomphonema | 3 | 8 | 3 | 13 | – |
| Gyrosigma | – | – | 3 | – | – |
| Kephyrion | 3 | – | – | – | – |
| Melosira | – | – | 4 | – | – |
| Monoraphidium | – | 5 | – | 5 | – |
| Navicula | 6 | 8 | 6 | 14 | – |
| Nitzschia | 6 | 8 | 3 | 16 | – |
| Nostoc | – | – | – | – | 5 |
| Pinnularia | 8 | 5 | – | 13 | 4 |
| Placoneis | 3 | – | – | – | – |
| Protoperidinium | – | – | – | 5 | – |
| Spirogyra | – | – | 3 | – | – |
| Staurastrum | 3 | – | – | – | 6 |
| Staurodesmus | – | – | – | – | 4 |
| Surirella | – | – | 3 | 7 | – |
| Thalassiosira | – | – | – | 7 | – |
| Trachelomonas | 3 | – | – | – | – |
| Ulnaria | – | 6 | 7 | 5 | – |
Диатомовые преобладают в списке ведущих родов на всех исследованных участках, кроме болот (табл. 2). Это, главным образом, роды порядка Raphales. На преддельтовом участке из шовных диатомей по числу видов лидируют роды Navicula, Nitzschia и Gomphonema, в дельте – Navicula, во взморье – Nitzschia, Navicula, Pinnularia и Gomphonema, в озерах – Pinnularia, Nitzschia, Aulocoseira, Navicula и Eunotia. Роды бесшовных диатомей входят в число ведущих только на трех участках ‒ дельте (Ulnaria и Fragilaria), преддельтовом и взморье (Ulnaria и Diatoma). В число ведущих родов диатомовых из класса Coscinodiscophyceae входят Aulacoseira, Melosira и Thalassiosira. Они входят в головную часть спектра на всех участках, но наиболее широко представлены видами во флоре взморья, дельты и преддельтового участка.
Отмечено 15 родов, которые вошли в число ведущих только на одном из выделенных участков (табл. 2). Пять из них относится к флоре заболоченных участков: представители десмидиевых (Cosmoastrum, Euastrum, Staurodesmus), цианопрокариот (Nostoc) и желто-зеленых (Characiopsis). Четыре таких рода отмечены во флоре дельты р. Лены: диатомовые и стрептофитовые (Melosira, Fragilaria, Gyrosigma и Spirogyra). Три рода характерны для озерной флоры: из диатомей ‒ Placoneis, золотистых ‒ Kephyrion и эвгленовых ‒ Trachelomonas. Во взморье также отмечено три рода (Thalassiosira, Diploneis и Protoperidinium). На преддельтовом участке таких родов не обнаружено.
В эколого-географической структуре альгофлоры исследованных участков были выявлены различия. На большинстве участков количество алкалифилов и алкалибионтов (видов, предпочитающих слабощелочную реакцию среды) превышает таковое ацидофилов (табл. 3). Только для болот характерна большая доля водорослей-ацидофилов. Индифференты составляют наибольшую часть видов-индикаторов pH воды, как по участкам, так и для флоры в целом (118 видов и разновидностей, или 40% водорослей-индикаторов).
Таблица 3.
Количество видов и разновидностей водорослей основных экологических и фитогеографических групп на различных участках низовья р. Лены
| Группа водорослей | Озера | Преддельтовый участок | Дельта | Взморье | Заболоченные участки |
|---|---|---|---|---|---|
| Ацидофилы | 10 | 21 | 9 | 19 | 20 |
| Индифференты к pH | 40 | 73 | 39 | 86 | 28 |
| Алкалифилы | 25 | 64 | 33 | 82 | 13 |
| Алкалибионты | 4 | 10 | 5 | 9 | 2 |
| Галофобы | 10 | 12 | 5 | 12 | 17 |
| Индифференты к солености | 78 | 160 | 73 | 184 | 56 |
| Галофилы | 10 | 29 | 17 | 35 | 5 |
| Эвригалинные | 2 | 3 | 2 | 8 | 2 |
| Мезогалобы | 1 | 4 | 2 | 12 | 6 |
| Планктонные | 40 | 66 | 35 | 80 | 26 |
| Бентосные | 40 | 88 | 41 | 118 | 33 |
| Планктонно-бентосные | 43 | 94 | 40 | 90 | 42 |
| Эпибионты | – | – | – | – | 2 |
| Почвенные | – | – | – | 1 | 1 |
| Космополиты | 96 | 190 | 92 | 210 | 80 |
| Бореальные | 9 | 22 | 8 | 34 | 4 |
| Арктоальпийские | 4 | 17 | 5 | 25 | 10 |
На всех участках во флоре преобладают индифференты по отношению к солености воды, составляющие от трети до половины видового списка (табл. 3). Большая доля во флоре видов, адаптированных к повышенной солености воды, характерна для дельты, взморья и преддельтового участка. В озерах галофобы и галофилы равны по числу видов, на заболоченных участках число галофобов наиболее высоко.
Во флоре взморья отмечено максимальное число эвригалинных видов (8) и мезогалобов (12), что соответствует условиям обитания в маргинальной зоне, расположенной на границе смешения пресных и соленых вод.
Количество планктонных видов водорослей меньше, чем представителей бентоса, на всех участках кроме озер (табл. 3). Значительную часть везде составляют гетеротопные виды, обитающие как в планктоне, так и в бентосе.
В планктонной флоре преобладают космополиты, достигая на различных участках от 43 до 71% видового состава (табл. 3). Наибольшее сходство с бореальной флорой имеет флора озер – бореальные виды преобладают над аркто-альпийскими. На преддельтовом участке, дельте и взморье флора умеренного пояса также богаче североальпийской. Аркто-альпийский элемент сильнее проявляется во флоре болот (см. табл. 3).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В результате проведенной работы более чем в 2 раза пополнен флористический список водорослей водоемов низовья р. Лены, ранее насчитывавший 302 вида и разновидности из 8 отделов [5]. Соотношение филумов во флоре исследованного региона характерно для водоемов севера европейской части России [11], Сибири [17, 19, 36] и североамериканского сектора Арктики [39, 42, 45]. Флористические пропорции близки к рассчитанным исследователями для альгофлор Большеземельской тундры [12], Полярного Урала [29] и других арктических и горных регионов [16, 31]. Преобладание в таксономическом спектре водорослей водоемов низовья р. Лены одно- и двувидовых родов и семейств – зональная специфика альгофлоры северных водоемов [12]. Состав альгофлоры исследованного региона характерен для северных ненарушенных водоемов.
Флора взморья наиболее богата в видовом отношении среди всех выделенных нами участков низовья р. Лены. Водоросли развиваются здесь на границе пресных и соленых вод, в так называемой маргинальной зоне, для которой часто отмечается увеличение видового богатства, по сравнению с соседними местообитаниями [25]. Бедный видовой состав водорослей дельты – несомненно результат недостаточной изученности ее флоры, учитывая значительную площадь последней и потенциальное разнообразие экотопов в условиях разветвленной сети проток.
Ведущее положение сем. Naviculaceae, отмеченное нами для флор большинства участков исследованного региона, характерно также для флоры Полярного Урала [33], Карелии [21] и Большеземельской тундры [12].
Наибольший вклад во флору представителей цианопрокариот выявлен для заболоченных участков, где по числу видов выделяется род Nostoc. Известно, что представители этого рода отличаются видовым разнообразием в водоемах различных секторов Арктики, например о. Шпицберген [14] и Полярного Урала [12].
Высокое разнообразие Chrysophyta в водоемах российского сектора Арктики подтверждается результатами исследований с применением методов электронной микроскопии. Их использование позволило выявить в водоемах п-ова Таймыр 23 вида чешуйчатых золотистых [40], в низовье р. Енисей и в термокарстовых озерах арктической части его бассейна 67 [1] и 47 видов соответственно [4], в водоемах Полярного Урала и Большеземельской тундры 46 [8] и 75 видов [10], соответственно. В водоемах исследованного нами региона среди золотистых водорослей наиболее полно представлен род Dinobryon, включающий семь видов. Хризофитовых с чешуйчатым панцирем отмечено немного – по одному виду из родов Mallomonas и Synura, что объясняется недостаточной изученностью их флоры. Золотистые водоросли не входили в головную часть спектра родов ни на одном из участков низовья р. Лены. Известно, что условия среды обитания в большинстве водоемов исследованного региона близки к экологическому оптимуму для золотистых водорослей [7], который определен по pH (5.5–6.5) и температуре воды (10–20°C) в работах [43, 46]. Соответственно, при дальнейших исследованиях с применением СЭМ следует ожидать значительного пополнения видового состава золотистых на всех участках низовья р. Лены. На заболоченных участках их видовое богатство может быть ограничено низкими значениями рН, но и здесь возможны находки хризофитовых водорослей, особенно из групп ацидофильных и ацидобионтных видов.
Сем. Desmidiaceae занимает лидирующее положение в структуре флор арктических и горных районов с суровым климатом [20, 37]. Известна также приуроченность десмидиевых к заболоченным местообитаниям [11]. Поэтому закономерно высокое видовое разнообразие представителей этого семейства, выявленное нами в озерах, и особенно в болотах исследованного региона. Однако во флорах преддельтового участка, дельты и взморья, виды сем. Desmidiaceae представлены единично. К особенностям флоры этих участков, представляющих собой единую речную систему, относится преобладание бореальных видов над северо-альпийскими. Очевидно, что источником флоры умеренного пояса служит сток фитопланктона из верхних участков Лены – крупной транзитной реки, протекающей в меридиональном направлении и пересекающей различные природно-климатические зоны, что способствует продвижению на Север представителей бореальной флоры.
Роды Navicula и Aulacoseira, которые входят в головную часть спектра на преддельтовом участке р. Лены, в ее дельте и принимающем речные воды взморье, входят в число ведущих родов и в устьях других крупных арктических рек – Енисея и Оби [30, 32]. Род Stephanodiscus в низовье р. Лены представлен только тремя видами. В устьях Енисея и Оби количество видов этого рода увеличилось после зарегулирования стока [3, 30], что связано с прогрессирующим процессом эвтрофирования.
Отношения числа видов-галофобов к галофилам, а также числа алкалифилов и алкалибионтов к ацидофилам соответствуют условиям среды, характерным для исследованного региона [7].
Небольшая доля истинно планктонных видов в альгофлоре низовья р. Лены, вероятно, обусловлена ее обогащением представителями бентоса и обрастаний за счет ветро-волнового перемешивания, которое типично как для взморья, дельты и преддельтового участков, так и для небольших водоемов, в условиях заполярной тундры.
Выводы. Альгофлора низовья р. Лены характеризуется богатым видовым составом, а водоемы исследованного региона сохраняют признаки ненарушенных арктических местообитаний. Река Лена, протекающая в меридиональном направлении, способствует переносу альгофлоры умеренного пояса в арктический регион. Это проявляется в особенностях таксономической структуры флоры и преобладании числа бореальных видов на участках низовья р. Лены, представляющих единую речную систему: преддельтовый участок, дельта и взморье. Ярко выраженный краевой эффект характерен для флоры взморья, что выражено в увеличенном видовом разнообразии. Морская флора оказывает влияние на видовой состав водорослей водоемов исследованного региона, которое происходит в результате заноса морских и солоновато-водных видов водорослей.
ФИНАНСИРОВАНИЕ РАБОТЫ
Работа выполнена в рамках государственного задания Института биологических проблем криолитозоны СО РАН на 2017–2020 гг. по теме “Фундаментальные и прикладные аспекты изучения разнообразия растительного мира Северной и Центральной Якутии”, регистрационный номер АААА-А17-117020110056-0.
Список литературы
Балонов И.М., Кузьмина А.Е. Золотистые водоросли // Гидрохимические и гидробиологические исследования водоемов Хантайского водохранилища. Новосибирск: Наука, 1986. С. 59–70.
Баринова С.С., Медведева Л.А., Анисимова О.В. Биоразнообразие водорослей-индикаторов окружающей среды. Тель-Авив: Студия Пилиес, 2006. 498 с.
Бессудова А.Ю., Сороковикова Л.М., Фирсова А.Д., Томберг И.В. Современное состояние вод нижнего участка реки Енисей // География и природные ресурсы. 2014. № 3. С. 93–99.
Бессудова А.Ю., Сороковикова Л.М., Букин Ю.С. и др. Гидрохимия и альгофлора термокарстовых озер арктической части бассейна нижнего Енисея // Синергия наук. 2017. № 16. С. 1175–1199.
Васильева И.И., Ремигайло П.А. Флора пресноводных водорослей Ленской дельты. Якутск, 1986. 24 с. Деп. в ВИНИТИ. № 2414-В86.
Васильева И.И., Ремигайло П.А., Габышев В.А. и др. Водоросли // Разнообразие растительного мира Якутии. Новосибирск: Изд-во СО РАН, 2005. С. 150–272.
Венглинский Д.Л., Лабутина Т.М., Огай Р.И. и др. Особенности экологии гидробионтов нижней Лены. Якутск: Изд-во Якутск. филиала СО АН СССР, 1987. 184 с.
Волошко Л.Н. Золотистые водоросли // Биоразнообразие экосистем Полярного Урала. Сыктывкар: Ин-т биологии Коми науч. центр УрО РАН, 2007. С. 57–69.
Волошко Л.Н. Современная система золотистых водорослей (Chrysophyta) // Ботан. журн. 2008. Т. 963. № 8. С. 1250–1264.
Волошко Л.Н., Гецен М.В., Гаврилова О.В. Разнообразие золотистых водорослей Большеземельской тундры // Морфология, систематика, онтогенез, экология и биогеография диатомовых водорослей: Тез. докл. IX школы диатомологов России и стран СНГ. Борок: Ин-т биологии внутр. вод РАН, 2005. С. 30.
Гецен М.В. Водоросли в экосистемах Крайнего Севера. Л.: Наука, 1985. 165 с.
Гецен М.В., Стенина А.С., Патова Е.Н. Альгофлора Большеземельской тундры в условиях антропогенного воздействия. Екатеринбург: Наука, 1994. 145 с.
Гогорев Р.М. Некоторые особенности горизонтального распределения фитопланктона в море Лаптевых (август–сентябрь 1993 г.) // Научные результаты экспедиции ЛАПЭКС-93. СПб.: Гидрометеоиздат, 1994. С. 337–352.
Давыдов Д.А. Cyanoprokaryota Шпицбергена, состояние изученности флоры // Ботан. журн. 2010. Т. 95. № 2. С. 169–176.
Дружкова Е.И., Макаревич П.Р. Исследования фитоплакнтона моря Лаптевых: история и современность // Тр. Кольск. науч. центра РАН. 2013. Вып. 1(14). С. 71–79.
Ермолаев В.И., Левадная Г.Д., Сафонова Т.А. Альгофлора водоемов окрестностей Таймырского стационара // Биогеоценозы Таймырской тундры и их продуктивность. Л.: Наука, 1971. С. 116–129.
Ермолаев В.И., Ремигайло П.А., Габышев В.А. Водоросли планктона водоемов бассейна озера Таймыр // Сиб. экол. журн. 2003. Т. 10. № 4. С. 381–392.
Зернова В.В., Нютик Е.М., Шевченко В.П. Вертикальные потоки микроводорослей в северной части моря Лаптевых (по данным годичной постановки седиментационной ловушки) // Океанология. 2000. Т. 40. № 6. С. 850–858.
Комаренко Л.Е. Планктон бассейна реки Яны. М.: Наука, 1968. 151 с.
Комулайнен С.Ф. Экология фитоперифитона малых рек Восточной Скандинавии. Петрозаводск: Карельск. науч. центр РАН, 2004. 182 с.
Комулайнен С.Ф., Чекрыжева Т.А., Вислянская И.Г. Альгофлора озер и рек Карелии. Таксономический состав и экология. Петрозаводск: Карельск. науч. центр РАН, 2006. 81 с.
Крахмальный А.Ф. Динофитовые водоросли Украины (иллюстрированный определитель). Киев: Альтпресс, 2011. 444 с.
Летописи природы Государственного природного заповедника “Усть-Ленский”. Гл. 7. Фитопланктон. Книги 8. Тикси: Фонды ФГБУ ГПЗ “Усть-Ленский”, 1997. С. 90–114.
Летописи природы Государственного природного заповедника “Усть-Ленский”. Гл. 7. Фитопланктон. Книги 9. Тикси: Фонды ФГБУ ГПЗ “Усть-Ленский”, 1997. С. 95–105.
Одум Ю. Основы экологии. М.: Мир, 1975. 740 с. (Odum Eu.P. Fundamentals of ecology. Philadelphia: Saunders, 1971. 555 р.)
Околодков Ю.Б. Водоросли льдов моря Лаптевых // Новости системат. низш. раст. 1992. Т. 28. С. 29–34.
Павлюкова Т.И. Фитопланктон. Ежегодник качества поверхностных вод по гидробиологическим показателям на территории деятельности Тиксинского УГМС за 1992 г. Тикси: Тиксинское ТУГМС, 1993. С. 7–9, 23–31.
Павлюкова Т.И. Фитопланктон. Обзор состояния загрязнения морской среды в зоне деятельности Тиксинского УГМС по гидробиологическим показателям за 1992 г. Тикси: Тиксинское ТУГМС, 1993. С. 19–23, 56–68.
Патова Е.Н., Демина И.В. Водоросли других отделов // Биоразнообразие экосистем Полярного Урала. Сыктывкар: Ин-т биологии Коми науч. центра УрО РАН, 2007. С. 69–90.
Приймаченко А.Д., Шевелева Н.Г., Покатилова Т.Н. и др. Продукционно-гидробиологические исследования Енисея. Новосибирск: Наука, 1993. 197 с.
Сафонова Т.А. Водоросли горных водотоков юга Западной Сибири. Разнообразие и таксономическая структура // Сиб. экол. журн. 1997. № 1. С. 91–96.
Семенова Л.А. Многолетние исследования фитопланктона Нижней Оби // Вестн. экологии, лесоведения и ландшафтоведения. 2009. № 9. С. 163–173.
Стенина А.С. Диатомовые водоросли // Биоразнообразие экосистем Полярного Урала. Сыктывкар: Ин-т биологии Коми науч. центра УрО РАН, 2007. С. 41–56.
Суханова И.Н., Флинт М.В., Георгиева Е.Ю. и др. Структура сообществ фитопланктона в восточной части моря Лаптевых // Океанология. 2017. Вып. 57. № 1. С. 86–102.
Тимофеев С.Ф. Пелагическая экосистема моря Лаптевых // Биология и океанография Карского и Баренцова морей (по трассе Севморпути). Апатиты: Изд-во Кольск. науч. центра РАН, 1998. С. 75–88.
Цибульский В.Р., Валеева Э.И., Арефьев С.П. и др. Природная среда Ямала. Тюмень: Изд-во Ин-та проблем освоения Севера СО РАН, 1995. Т. 1. 168 с.
Algae of Ukraine: Diversity, Nomenclature, Taxonomy, Ecology and Geography // Wasser and Eviatar Nevo: V. 4: Charophyta. Ruggell: A.R.G. Gantner Verlag, 2014. 703 p.
Coesel P.F.M. Biogeography of desmids // Hydrobiology. 1996. V. 336. P. 41–43.
Hilliard D.K. Notes on the phytoplankton of Karluk Lake, Kodiak Island, Alaska // Can. Field–Natur. 1959. V. 73. P. 135–143.
Kristiansen J., Düwel L., Wegeberg S. Silica-scaled chrysophytes from the Taymyr Peninsula, Northern Siberia // Nova Hedwigia. 1997. V. 65. № 1. P. 337–351.
Okolodkov Yu.B. A checklist of dinoflagellates recorded from Russian Arctic seas // Sarsia. 1998. V. 83. P. 267–292.
Prescott G.M. Ecology of Alaskan freshwater algae. Introduction: general considerations // Trans. Amer. Microscop. Soc. 1963. V. 82. № 1. P. 83–98.
Sandgren C.D. The ecology of chrysophyte flagellates. Their growth and perennation strategies as freshwater phytoplankton // Growth and reproductive strategies of freshwater phytoplankton. USA; N.Y.: Cambridge Univ. Press, 1988. P. 9–104.
Schilthuizen M. Ecotone: Speciation-prone // Trends Ecol. Evol. 2000. V. 15. № 4. P. 130–131.
Sheath R.G., Munawar M. Phytoplankton composition of a small subarctic lake in the Northwest Territories, Canada // Phycologia. 1974. V. 13. № 2. P. 149–161.
Siver P.A., Hammer J.S. Multivariate statistical analysis of the factors controlling the distribution of scaled chrysophytes // Limnol., Oceanogr. 1989. V. 34. P. 368–381.
Smith T.B., Wayne R.K., Girman D.J., Bruford M.W. A role for ecotones in generating rainforest biodiversity // Science. 1997. V. 276. № 5320. P. 1855–1857.
Sörensen T. A method of establishing groups of equal amplitude in plant sociology based on similarity of species content // Kongel. Danke Vid. bernes Selskab. Biol. Krifter. 1948. Bd 5. № 4. P. 1–34.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод