

Биология внутренних вод, 2019, № 2-2, стр. 68-74
Плотность и распределение рыб в реке, с ярко выраженной гетерогенностью среды, по результатам гидроакустической съемки
Ю. В. Герасимов 1, Э. С. Борисенко 1, М. И. Базаров 1, И. А. Столбунов 1
1 Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
152742 Ярославская обл., Некоузский р-н, пос. Борок, Россия
Поступила в редакцию 18.08.2017
После доработки 22.03.2018
Принята к публикации 29.05.2018
Аннотация
Эхолотом горизонтального зондирования исследовано пространственное распределение рыб на нижнем участке р. Савалы (приток реки Хопер, Воронежская обл.). В пределах участка сравнивали плотность рыб на отрезках русла с разной степенью меандрированности. Показано, что морфология русла оказывает влияние на пространственное распределение рыб при высоком уровне воды весной. Осенью, при уменьшении плотности рыб, связанного с понижением уровня воды в реке и скатом рыб после завершения нереста, разница в плотности скоплений рыб на участках с разной степенью меандрированности русла уменьшалась, а максимальные плотности наблюдались на наиболее глубоких участках реки.
ВВЕДЕНИЕ
Малые и средние реки играют значительную роль в функционировании рыбного сообщества водоемов, притоками которых являются. Для большинства рыб они служат транспортными магистралями, по которым происходят нерестовые миграции, а затем скат молоди, пополняющей рыбное население водоема или водотока-приемника. Часто малые и средние реки становятся рефугиумами для видов рыб, которые в силу различных причин исчезают в водных объектах. В связи с этим изучение механизмов формирования структуры речных рыбных сообществ имеет важное теоретическое и практическое значение.
Структура речных рыбных сообществ во многом определяется сезонной динамикой факторов, например, зависит от изменчивости гидродинамических и гидрологических показателей (скорости течения, глубины и т.д.). Пространственное распределение рыб сильно зависит и от структурной неоднородности русла, обусловленной наличием поворотов, бороздин, ям, перекатов и т.д., которые могут стать привлекающими факторами для рыб [6, 7, 11]. Поэтому изучение структуры речных рыбных сообществ требует разномасштабного подхода, который отражает иерархию факторов, определяющих распределение гидробионтов в речных системах [12, 15–19]. Оценка численности и пространственного распределения рыб в небольших реках – достаточно сложная методическая задача. Такие реки характеризуются ярко выраженной гетерогенностью среды, включая заросли макрофитов и прибрежных кустарников, обширных отмелей и перекатов, резких перепадов глубин фактически по всей акватории реки и переменной скоростью течения, что затрудняет или делает невозможным определение численности и исследование распределения рыб с использованием сетных орудий лова. Например, коэффициент уловистости закидных неводов в реке в значительной степени зависит от рельефа дна, глубины и скорости течения [8].
Вследствие этого, перспективно использование гидроакустической аппаратуры эффективной в более широком диапазоне изменчивости указанных факторов. Кроме того, она позволяет проводить непрерывное зондирование водной толщи с минимальным возмущением среды, в отличие от методов с использованием сетных орудий лова. Однако из-за малых глубин в небольших водотоках зачастую невозможно применение гидроакустической аппаратуры вертикальной локации. Поэтому для количественной оценки рыбного населения мелководных рек, а также в приповерхностных слоях воды речных акваторий предпочтительнее использование гидроакустических комплексов горизонтального зондирования.
Цель работы – оценить численность и распределение рыб в нижнем течении р. Савалы с использованием гидроакустической аппаратуры.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Река Савала – правый и самый крупный приток р. Хопер, впадающий в 315 км от ее устья [3]. По своим параметрам (длина реки 285 км, площадь бассейна 7720 км2) она относится к средним рекам. Река в устьевой части представляет собой корытообразное или треугольное в поперечном сечении русло с симметричным или асимметричным расположением глубокой части русла относительно берегов. Ширина русла 36–50 м, глубина >4 м, скорость течения ≤0.4 м/с, основной тип зарастания макрофитами – вдольбереговой с проективной площадью зарастания ≤80%. Участок реки, обследованный весной, имел песчаное дно, в устьевой части с примесью гравия, ширина русла участка, обследованного весной, 4–12 м, глубина ≤1 м, скорость течения 0.1 м/с, основной тип зарастания макрофитами – вдольбереговой с проективной площадью зарастания ≤20%. Участок реки, обследованный осенью, имел песчаное дно, ширину русла 16–23 м, глубинy ≤1.5 м, скорость течения 0.1 м/с, основной тип зарастания макрофитами – вдольбереговой с проективной площадью зарастания ≤20%.
Гидроакустические исследования на р. Савале проводили 29–31 мая и 22, 23 сентября 2014 г. Весной обследовали участок от 51°2ʹ45ʺ с.ш., 41°37ʹ30ʺ в.д. вверх по течению до 51°3ʹ11ʺ с.ш., 41°37ʹ25ʺ в.д. и участок устья от 51°2ʹ47ʺ с.ш., 41°40ʹ40ʺ в.д. вверх по течению до 51°2ʹ43ʺ с.ш., 41°39ʹ27ʺ в.д. Осенью уровень воды снизился на 70 см, в результате обследованный весной участок реки обмелел до уровня, когда эксплуатация моторной лодки стала невозможна, и исследования проводили на участке, расположенном ниже по течению – от 51°2ʹ45ʺ с.ш., 41°37ʹ30ʺ в.д. до устья реки 51°2ʹ47ʺ с.ш., 41°40ʹ40ʺ в.д.
Методика отлова рыб. Качественный анализ ихтиофауны исследуемого участка реки проводили с применением мальковой волокуши (длина 9 м, высота 1.5 м, ячея в крыльях и кутке 4 мм) на пригодных для этого местах русла. Использовать другие орудия лова (ставных сетей и закидного невода) оказалось невозможно из-за небольшой ширины и глубины русла. После промеров рыб старших возрастных групп отпускали в реку, раннюю молодь фиксировали 40%-ным формалином и идентифицировали до вида по определителям [1, 2, 5].
Гидроакустическая аппаратура и методика. Исследования проводили с помощью малогабаритного научно-исследовательского комплекса горизонтального зондирования “PanCor” (ООО “Промгидроакустика”, Россия) [3], установленного на лодке. Рабочая частота гидролокатора 455 кГц, ширина характеристики направленности одного луча антенны на уровне –6 дБ – 10°, ширина многолучевого сектора в плоскости веера лучей – 60°, максимальная дистанция регистрации одиночной рыбы – 20 м. Гидроакустические съемки проводили на постоянной скорости (~1–1.5 м/с) лодки с малошумным электромотором. Узость реки не позволяла осуществлять гидроакустическую съемку поперечными галсами, поэтому лодка перемещалась вдоль правого берега реки вверх по течению и вдоль левого – вниз по течению. Акваторию зондировали в горизонтальном режиме в плоскости, перпендикулярной направлению движения [13, 14]. Из-за наличия обширных отмелей, зарастания и извилистости русла реки эхосъемки проводили только в светлое время суток. Достоверность различий плотности скоплений рыб на разных участках анализировали с использованием непараметрического критерия Уилкоксона–Манна–Уитни, поскольку распределение исходных данных в большинстве случаев существенно отличалось от нормального. Связь значений плотности рыб вдоль левого и правого берега исследовали с помощью регрессионного анализа.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
При облове исследуемого участка сетными орудиями лова обнаружено семь видов рыб (табл. 1). Весной в уловах доминировали плотва и язь (~85% общей численности улова). Осенью на фоне снижения численности рыб видовой состав уловов не изменился.
Таблица 1.
Доля видов рыб в уловах малькового невода из р. Савалы
| Вид | % |
|---|---|
| Плотва Rutilus rutilus (L.) | 53.8 |
| Язь Leuciscus idus (L.) | 30.8 |
| Елец L. leuciscus (L.) | 1.3 |
| Елец Данилевского L. danilewskii (Kessler) | 6.4 |
| Уклейка Alburnus alburnus (L.) | 2.6 |
| Голавль L. cephalus (L.) | 1.3 |
| Верховка L. delineatus (Heckel) | 3.8 |
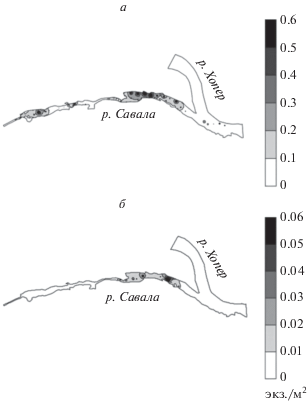
Весной при обследовании русла было выделено два отрезка: верхний – сильно меандрированный (коэффициент извилистости (к.и.) 3.5), и нижний – менее извилистый (к.и. 1.3) (рис. 1). На верхнем отрезке крутые повороты (до 90°) русла реки чередовались с относительно короткими прямыми участками. Нижний отрезок состоял из двух прямых участков, расположенных относительно друг друга под углом 90° (рис. 1). При отсутствии поворотов глубокая часть русла, достигавшая 1/3 ширины русла, меандрировала между берегами, образуя отмели то у левого, то у правого берега.
Рис. 1.
Схема р. Савалы в нижнем течении. 1 – акватория реки, обследованная весной (а – верхний отрезок, б – нижний отрезок); 2 – акватория реки, обследованная осенью (в – верхний отрезок, г – нижний отрезок); 3 – акватория реки, обследованная весной и осенью.
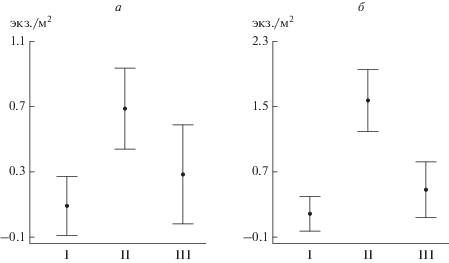
Весной на меандрированном верхнем отрезке плотность рыб была достоверно выше, чем на относительно прямом нижнем (0.67 ± 0.26 и 0.12 ± ± 0.06 экз./м2 соответственно; W = 2564, p < 0.01) (рис. 2). Коэффициент вариации численности рыб был так же значительно выше на верхнем отрезке (214%), чем на нижнем (174%). Сравнение плотности рыб на прямых участках и на поворотах у вогнутых и выпуклых берегов показало, что наибольшие плотности наблюдаются на поворотах у вогнутых берегов как у правого (1.58 ± 1.22 экз./м2), так и у левого (0.69 ± 0.67 экз./м2) (рис. 3). Плотность рыб у выпуклых берегов была 0.49 ± 0.40 экз./м2 (W = 115, p < 0.05) и 0.28 ± 0.17 экз./м2 (W = 198, p < 0.05) соответственно, при отсутствии поворотов – 0.19 ± 0.10 экз./м2 (W = 579, p < 0.05) и 0.09 ± ± 0.04 экз./м2 (W = 708, p < 0.05) соответственно. Самый низкий коэффициент вариации (144%) отмечен на участках у вогнутых берегов с высокой плотностью, на прямых участках и у выпуклых берегов этот показатель был выше (184 и 173% соответственно).
Рис. 2.
Плотность рыб (экз./м2) весной (а) и осенью (б) на верхнем (1) и нижнем (2) отрезках исследованных участков р. Савалы.
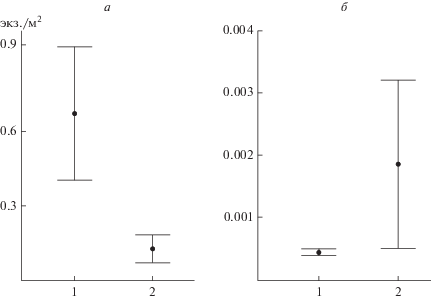
Рис. 3.
Плотность рыб (экз./м2) на разных участках левого (а) и правого (б) берегов р. Савалы весной. I – прямой участок, II – вогнутый берег, III – выпуклый берег.
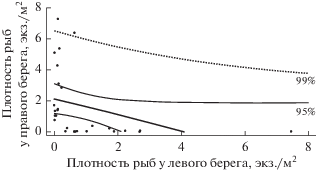
Анализ значений плотности на расположенных друг против друга участках левого и правого берега показал, что после исключения участков с обоюдно низкой плотностью рыб (участки с обоюдно высокой плотностью отсутствовали) между оставшимися обнаружилась обратно пропорциональная зависимость (r = –0.36, F = 3.9; p <0.05) (рис. 4). Средняя плотность рыб на участках с ее высокими значениями превосходила в >20 раз таковую на противоположных участках с более низкими показателями (2.47 ± 0.83 экз./м2 и 0.11 ± ± 0.05 экз./м2 соответственно; W = 725, p < 0.01). Коэффициент вариации на участках с высокими значениями плотности был ниже, чем на противоположных участках с низкими показателями (85 и 130% соответственно).
Осенью на обследованном участке русла выделяли тоже два отрезка. Как и весной, верхний отрезок был более извилистый (к.и. = 1.3), чем нижний (к.и. = 1.1). На верхнем отрезке русло плавно меандрировало, не совершая крутых поворотов (рис. 1). Нижний отрезок был более прямой и переходил в расширенную устьевую часть реки.
Рис. 4.
Связь значений плотности рыб на расположенных друг против друга участках левого и правого берега. Приведены линия регрессии и линии 99 и 95%-го доверительных интервалов (r = –0.41; F = 3.9; p < 0.05).
Осенью на исследованной акватории в основном наблюдались одиночные особи, средняя плотность рыб была значительно ниже, чем весной (0.004 ± 0.003 и 0.494 ± 0.24 экз./м2 соответственно; W = 35251, p < 0.01) (табл. 2), а коэффициент вариации плотности значительно выше (468%, против 184% весной). Осенью, в отличие от весны, отсутствовала достоверная разница между средней плотностью скоплений на поворотах русла реки и на прямых участках (рис. 2а, 2б).
Таблица 2.
Численность рыб в р. Савале в весенний и осенний периоды
| Участок реки | Площадь, м2 | Средняя плотность, экз./м2 | Общая численность, экз. | |||
|---|---|---|---|---|---|---|
| весна | осень | весна | осень | весна | осень | |
| 1 | 24 200 | – | 0.494 | – | 12 000 | – |
| 2 | – | 99 500 | – | 0.004 | – | 400 |
| 3 | 51 100 | 47 000 | 0.476 | 0.023 | 24 300 | 1800 |
Плотность рыб на верхнем отрезке была меньше, чем на нижнем (0.0004 ± 0.0001 и 0.002 ± ± 0.001 экз./м2 соответственно; W = 1218, p < < 0.05) (рис. 2б), а коэффициент вариации выше (471 и 109% соответственно).
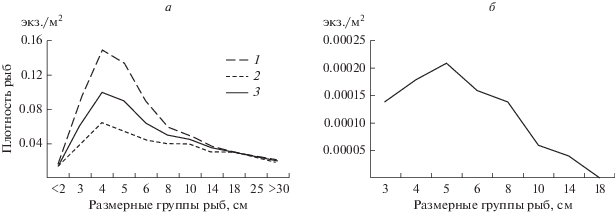
Осеннее снижение плотности произошло во всех размерных группах рыб, зарегистрированных на этом участке реки (рис. 5). На фоне снижения плотности из реки исчезли наиболее крупные особи размером >14 см, составляющие весной >2% общей численности.
Рис. 5.
Размерный состав скоплений рыб в р. Савала весной (а) и осенью (б). 1 – левый берег; 2 – правый берег; 3 – средние значения.
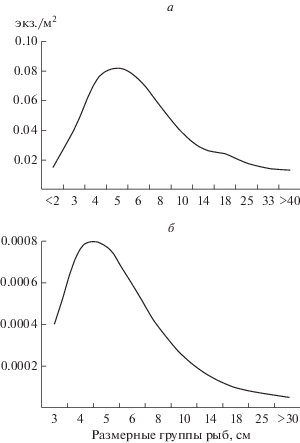
На расширенном устьевом участке реки гидроакустическую съемку проводили весной и осенью. Здесь также наблюдалось значительное снижение плотности рыб осенью (рис. 6а, 6б). Наиболее плотные скопления весной и осенью зарегистрированы в нижней части участка. Как и на других участках реки, осеннее снижение плотности произошло у рыб всех размерных групп (рис. 7а, 7б). На фоне снижения общей плотности здесь почти исчезли наиболее крупные особи (>30 см), которые весной достигали >10% общей численности.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Весной плотность рыб была выше на верхнем отрезке исследованного участка р. Савалы с наиболее высоким коэффициентом извилистости. Разнообразие местообитаний на этом отрезке реки обусловило более высокий коэффициент вариации плотности рыб, чем на менее извилистом нижнем. Это связано с достоверным различием плотностей рыб на прямых участках, поворотах у вогнутых и выпуклых берегов (рис. 3). Наибольшие плотности наблюдаются на поворотах у вогнутых берегов, здесь в петле излучины у вогнутого участка берега образуются плесы – глубокие участки русла реки с медленным течением, где оседают на дно взвешенные частицы, в том числе и кормовые организмы дрифта. На этих участках хорошие кормовые условия сочетаются с пониженными скоростями стокового течения (<15 см/с для рео-лимнофильных рыб) [9, 10], т.е. рыбы “оптимизируют” свое местоположение относительно скорости потока и обилия корма (дрифта).
На обширных песчаных перекатах и плесах, занимающих на прямых участках реки всю ширину русла, происходит размыв реоградиента, что, в свою очередь, приводит к диффузии скоплений рыб [10] и, как следствие, к снижению диапазона колебаний их плотности. Однако в р. Савале даже на прямых участках глубокая часть основного русла, составляющая до ≤1/3 его ширины, меандрирует между берегами, образуя отмели то у левого, то у правого берегов. Подобная асимметричность расположения глубокой части русла относительно берегов определяет наличие обратной связи между значениями плотности рыб на расположенных друг против друга лево- и правобережных участках, т.е. у берега, противоположного участку с высокой плотностью рыб, в большинстве случаев наблюдается скопление с низкой плотностью.
Сезонные изменения сопровождались более чем трехкратным снижением плотности рыб. Изменялся и размерный состав рыб в скоплениях. Осенью исчезали рыбы размерной группы <2 см, что обусловлено ростом сеголетков, которые у большинства карповых и окуневых к этому времени достигают размеров ≥3 см. Исчезновение рыб размерной группы >14 см связано как со скатом половозрелых особей после окончания весеннего нереста, так и с уходом крупных особей при снижении уровня воды вниз по течению реки на более глубокие устьевые участки или в р. Хопер. Глубоководные участки в условиях значительных колебаний уровня воды представляют собой местообитания с более стабильными условиями окружающей среды, чем мелководные [4, 17]. Поэтому в осенний период средняя плотность рыб на верхнем более меандрированном, но мелководном отрезке исследованного участка, была достоверно ниже, чем на менее извилистом, но более глубоководном нижнем. Таким образом, на фоне осеннего снижения уровня воды зависимость плотности рыб от конфигурации русла исчезла, и максимальные плотности отмечены на наиболее глубоких участках реки.
Процесс осеннего снижения плотности рыб связан не только с понижением уровня воды в реке, но и со скатом рыб после завершения нереста. Это подтверждается тем, что даже на наиболее глубоководных приустьевых участках реки осенью наблюдается значительное снижение плотности.
Выводы. Весной, при высоком уровне воды, плотность рыб была выше на верхнем отрезке исследованного участка р. Савалы с наиболее высоким коэффициентом извилистости. Продольная неоднородность распределения рыбы обусловлена зависимостью плотности рыб от конфигурации русла – плотность рыб на прямых участках, поворотах у вогнутых и выпуклых берегов достоверно различалась. Поперечная неоднородность распределения рыб в реке определялась асимметричностью расположения глубокой части русла относительно берегов. Наиболее низкими коэффициентами вариации плотности рыб характеризовались участки у вогнутых берегов с высокой плотностью рыб, на прямых участках и у выпуклых берегов этот показатель был выше. Осенью, при уменьшении плотности рыб, связанном с понижением уровня воды и скатом рыб после завершения нереста, зависимость плотности рыб от конфигурации русла исчезла. Максимальные плотности наблюдались на наиболее глубоких участках реки, а высокими коэффициентами вариации плотности рыб характеризовались мелководные участки с низкими значениями плотности рыб.
Список литературы
Атлас пресноводных рыб России. М.: Наука, 2003. Т. 1. 379 с.
Атлас пресноводных рыб России. М.: Наука, 2003. Т. 2. 253 с.
Дмитриева В.А. Гидрологическая изученность Воронежской области. Каталог водотоков. Воронеж: Издат.-полиграф. центр Воронеж. гос. ун-та, 2008. 225 с.
Ефремов Д.А. Распределение и поведение реофильных видов рыб в реках Восточной Фенноскандии. Автореф. дис. … канд. биол. наук. Петрозаводск, 2012. 24 с.
Коблицкая А.Ф. Определитель молоди пресноводных рыб. М.: Легк. и пищ. пром-сть, 1981. 208 с.
Мартинсен Ю.В. Влияние течений на поведение рыб // Рыб. хоз-во. 1940. № 12. С. 23–27.
Мартинсен Ю.В. Движение рыб в реке // Рыб. хоз-во. 1937. № 2. С. 27–30.
Новожилов Е.П. Влияние коэффициента уловистости орудий рыболовства на точность определения состояния запасов гидробионтов // Изв. Калиниград. гос. тех. ун-та. 2014. № 32. С. 250–254.
Павлов Д.С., Барекян А.Ш., Рипинский И.И. и др. Экологический способ защиты рыб на повороте струей открытого потока. М.: Наука, 1982. 112 с.
Поддубный С.А., Герасимов Ю.В., Новиков Д.А. Структура течений и распределение рыб в речных плесах верхневолжских водохранилищ // Биол. внутр. вод. 2003. № 1. С. 89–97.
Попов П.А. О характере распределения рыб на территории Горного Алтая // Вестн. Томск. гос. ун-та. Сер. Биол. 2013. № 2(22). С. 141–149.
Biggs B.J.F., Nikora V.I., Snelder T.H. Linking scales of variability to lotic ecosystem structure and function // River Res. Appl. 2005. V. 21. P. 283–298.
Borisenko E.S., Gusar A.G., Goncharov S.M. The target strength dependence of some freshwater species on their length-weiht characteristics // Proceedings of the Institute of Acoustics. England: Lowestoft UK., 1989. V. 11. Pt 3. P. 27–34.
Borisenko E.S., Degtev A.I., Mochek A.D., Pavlov D.S. Hydroacoustic characteristics of mass fishes of Ob-Irtish basin // J. Ichthyol. 2006. V. 46. Suppl. 2. P. S227–S234.
Bunn S.E., Arthington A.H. Basic principles and ecological consequences of altered flow regimes for aquatic biodiversity // Environ. Manag. 2002. V. 30. P. 492–507.
Fausch K.D., Torgersen C.E., Baxter C.V., Li H.W. Landscapes to riverscapes: bridging the gap between research and conservation of stream fishes // BioScience. 2002. V. 52. P. 483–498.
Kennard M.J., Olden J.D., Arthington A.H. et al. Multiscale effects of flow regime and habitat and their interaction on fish assemblage structure in eastern Australia // Can. J. Fish. and Aquat. Sci. 2007. V. 64. P. 1346–1359.
Lowe W.H., Likens G.E., Power M.E. Linking scales in stream ecology // BioScience. 2006. V. 56. P. 591–597.
Poff N.L., Allan J.D., Bain M.B. et al. The natural flow regime: a paradigm for riverine conservation and restoration // BioScience. 1997. V. 47. P. 769–784.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод