

Биология внутренних вод, 2019, № 2-2, стр. 42-50
Факторы, определяющие организацию перифитонного сообщества Bacillariophyta
Н. А. Исакова 1, *, Е. И. Вейсберг 1
1 Ильменский государственный заповедник
456317 Челябинская обл., Миасс, Россия
* E-mail: isakova_70@mail.ru
Поступила в редакцию 16.10.2016
После доработки 31.10.2017
Принята к публикации 07.12.2017
Аннотация
В предгорном оз. Большое Миассово (Южный Урал) исследованы состав и структура перифитонного сообщества Bacillariophyta на четырех широко распространенных видах погруженных гидрофитов – Myriophyllum sibirica Kom., Potamogeton lucens L., P. perfoliatus L. и Stratiotes aloides L. Установлено, что основные факторы, определяющие его организацию, – характер береговой линии и изменения физико-химических параметров озерной воды, вызванные процессами естественного эвтрофирования и дистрофирования.
ВВЕДЕНИЕ
Перифитонные сообщества Bacillariophyta на погруженных гидрофитах считаются одним из основных компонентов водных экосистем [2, 3, 7, 21]. Они развиваются в литоральной зоне – экотоне. На формирование сообществ экотонов, принадлежащих к различным уровням иерархии (biome, landscape, population, plant ecotone), воздействуют факторы от глобальных и ландшафтных (климат, макро- и мезорельеф, почвенный покров и др.) до локальных (микроклимат, микрорельеф, межвидовые взаимоотношения и др.) [10, 17, 18]. Диатомовые водоросли как планктонные, так и литоральные, благодаря хорошей сохранности в донных отложениях и индикационным свойствам, используются при палеолимнологических реконструкциях климата. Воздействие локальных факторов на формирование диатомовых комплексов донных отложений при палеореконструкциях учесть затруднительно. Недостаток данной информации может привести к искажению полученных выводов о тенденциях климатических изменений, поэтому актуально предварительное исследование факторов, влияющих на состав и структуру современных сообществ [11], в том числе и перифитонных Bacillariophyta.
Результаты исследований по данной тематике многочисленны и часто противоречивы. Большинство авторов придерживается мнения, что различия в структуре фитоперифитонных комплексов на макрофитах обусловлены преимущественно биотическими факторами, например микроклиматом, который создается физиологическими и микротопографическими особенностями водных растений [21, 29], но не менее важны и внешние физико-химические факторы среды [19, 27, 28]. С точки зрения других авторов, воздействие на них оказывает комбинация биотических и абиотических факторов [16, 30]. В некоторых работах анализируются данные из разнотипных по генезису и трофическому статусу озер или из различных экологических групп макрофитов, или учитываются все систематические отделы водорослей перифитона. В результате обрабатывается массив разнородных данных, позволяющий оценить воздействие факторов в комплексе. При этом ведущие факторы, которые необходимо учитывать при флористических и гидробиологических исследованиях, остаются скрытыми.
Цель работы – выявить латентные факторы, влияющие на структуру перифитонного сообщества Bacillariophyta, и оценить его как систему организованных группировок с помощью разведочных статистических методов.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Оз. Большое Миассово, расположенное в центральной части Ильменского государственного заповедника, имеет тектоническое происхождение по линиям меридиональных разломов, что определяет удлиненно-овальную конфигурацию его котловины (рис. 1). Длина озера 8.0 км, средняя ширина 1.5 км, площадь водного зеркала 11.4 км2, максимальная и средняя глубина 25.0 и 11.2 м соответственно [8]. Литоральная зона развита слабо: площадь дна до 5-метровой изобаты достигает 27.8%. Коэффициент изрезанности береговой линии 2.9 (максимальная величина для озер заповедника). В оз. Большое Миассово выделяют 10 заливов глубиной 4.0–8.0 м. Прозрачность воды большую часть безледного периода колеблется в пределах 3.0–5.0 м. Наименьшие ее значения отмечены весной, после схода льда и осенью, начиная с конца августа [15]. Озеро относится к водоемам с прямой стратификацией летом и обратной – зимой. В летний период температура придонных слоев воды 5–6°С, поверхностных – 25–26°С. В заливах стратификация не наблюдается, а гомотермия устанавливается к началу июля [1, 15].
Рис. 1.
Схема расположения пробных площадей в литоральной зоне оз. Большое Миассово. Здесь и в табл. 1 и 2: 1 – кордон, 2 – научно-производственная база, 3 – залив Зимник, 4 – залив Латочка, 5 – залив Штанная курья, 6 – залив Няшевская курья, 7 – залив Узкая курья, 8 – залив Липовая курья. ИГЗ – Ильменский государственный заповедник.
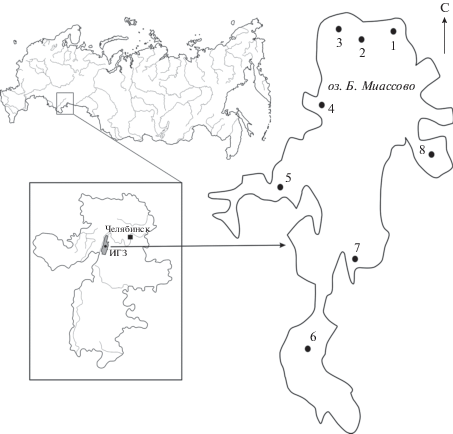
В литоральной зоне озера у открытых участков берега и в заливах, именуемых курьями, 17–19 августа 2015 г. было обследовано восемь пробных площадей (ПП) (рис. 1).
Кордон (ПП 1) – открытый участок у северного каменисто-песчаного берега, окружен невысоким земляным валом. Грунт каменисто-песчаный, местами заиленный. Гидрофитная растительность фрагментарная. В составе сообществ доминируют Potamogeton lucens L., Stratiotes aloides L. и Chara aspera Deth. ex Willd.
Научно-производственная база (ПП 2) – открытый участок у северного берега с выходами камней и фрагментами каменисто-песчаных пляжей, окружен невысоким земляным валом. Грунт песчаный, местами заиленный. У берега разреженная гидрофитная растительность представлена Potamogeton perfoliatus L., P. lucens, Myriophullum sibiricum Kom., Chara aspera, реже – Batrachium circinatum (Sibth.) Spach; на глубине 1.2 м развиты сомкнутые сообщества с доминированием Myriophullum sibiricum и Stratiotes aloides.
Залив Зимник (ПП 3) окаймлен высоким береговым валом с выходами камней, имеет незамкнутые очертания и сообщается с центральной частью озера. На каменисто-песчаном грунте у берега растительность представлена немногочисленными экземплярами Potamogeton perfoliatus, P. lucens, Myriophullum sibiricum, Chara aspera, реже – Batrachium circinatum, Persicaria amphibia (L.) S.F. Gray; глубже, на илистом грунте, развиты мозаичные сообщества с доминированием Myriophullum sibiricum, Stratiotes aloides и Potamogeton lucens, встречаются Elodea canadensis Michx., Batrachium circinatum и Nitellopsis obtusa (Desv. in Lois.) Gr.
Залив Латочка (ПП 4) – небольшой залив, имеющий незамкнутые очертания. Береговая линия включает фрагменты тростниково-осоковой сплавины. Грунт илистый. Среди водной растительности преобладает Nuphar lutea (L.) Smith, изредка встречается Nymphaea candida J. et C. Presl. В их плотных зарослях произрастают погруженные гидрофиты Potamogeton compressus L., P. lucens, P. perfoliatus, Myriophullum sibiricum, Fontinalis antipyretica Hedw., Chara tomentosa L. и Nitellopsis obtusa.
Залив Штанная курья (ПП 5) имеет вытянутую в западном направлении форму. Береговая линия сильно изрезана, с характерной комбинацией каменистых и заболоченных участков. Грунт илистый. Водная растительность хорошо развита, доминируют Nuphar lutea и Myriophullum sibiricum. Встречаются фрагменты сообществ рдестов и харовых водорослей.
Залив Няшевская курья (ПП 6) – большой мелководный залив, имеет мешковидную форму и замкнутые очертания. Дно илистое. Разнообразные сообщества гелофитов и гидрофитов полностью занимают его площадь.
Залив Узкая курья (ПП 7) имеет высокие берега, понижающиеся в южном направлении. Грунт илисто-песчаный, илистый. Заболоченная часть береговой линии окаймлена полосой Nuphar lutea, на остальной части отмечены фрагментарные сообщества рдестов и урути сибирской с участием гелофитов.
Залив Липовая курья (ПП 8) – большой блюдцеобразный залив, имеет замкнутые очертания. Мелководный участок береговой линии с прибрежной тростниково-осоковой сплавиной. Грунт илистый, торфяно-илистый. Со сплавиной граничит полоса плотных зарослей N. lutea, переходящая в сообщества Myriophullum sibiricum и Stratiotes aloides с участием Batrachium circinatum, Elodea canadensis и Chara spp.
Гидрохимические пробы отбирали согласно общепринятым международным стандартам и обрабатывали в Южно-Уральском центре коллективного пользования по исследованию минерального сырья (аттестат аккредитации № ААС.А.00300). Определение азота и фосфора проводилось фотометрическими методами: аммонийной формы азота $\left( {{\text{NH}}_{4}^{ + }} \right)$ – с реактивом Несслера, нитритной формы азота $\left( {{\text{NO}}_{2}^{ - }} \right)$ – с реактивом Грисса, нитратной формы азота $\left( {{\text{NO}}_{3}^{ - }} \right)$ – с салицилатом натрия C7H5NaO3. Для определения содержания общего фосфора пробы нефильтрованной воды предварительно окисляли путем кипячения с пироксодисульфатом аммония (NH4)2S2O8. Растворимые фосфаты (PO43–) определяли реактивом, содержащим молибден, сурьму и аскорбиновую кислоту. Оптическую плотность проб замеряли на фотометре фотоэлектрическом “КФК-3”. Растворенный кислород измеряли термооксиметром “АНИОН-4140”.
На ПП c глубины 1.0–1.5 м отбирали распространенные по акватории водоема погруженные гидрофиты Myriophullum sibiricum, Potamogeton lucens, P. perfoliatus и Stratiotes aloides. Анализировали вегетативную часть растений, выбранных в 5-кратной повторности случайным образом и произрастающих недалеко друг от друга. Всего 27 проб. Фитоперифтон отделяли от растений энергичным встряхиванием в холодной водопроводной воде. Суспензию сливали в емкость бо́льшего объема [18]. По окончании отмывки фитоперифитона объем воды фиксировали, а гидрофитов взвешивали на аналитических весах. Далее полученную суспензию фильтровали через полиамидное сито (размер ячеек 14 мкм). Фильтрат объемом 2 мл переносили в стеклянную пробирку, добавляли раствор, содержащий гипохлорит-ион, выдерживали до полного обесцвечивания, многократно отмывали дистиллированной водой, а затем разводили в 20 мл воды. Подготовку проб к микроскопированию проводили по стандартной методике [4]. Створки диатомей заключали в среду Эльяшева. Их подсчитывали при окуляре ×10 и объективе ×100 (микроскоп Микмед-6) до 100 клеток в препарате [7]. Далее пересчитывали количество клеток на общий объем воды, а затем на 1 г гидрофита. При идентификации видов диатомовых водорослей использовали определители [5, 6, 20, 22–26, 31].
При статистической обработке материала применяли кластерный анализ (cluster analysis) и анализ соответствий (correspondence analysis). Для измерения сходства между 27 описаниями, содержащими информацию о флористическом составе бентосных диатомей (планктонных и планктонно-бентосных Bacillariophyta исключали из описаний как “случайные”) и о их абсолютной численности, использовали индекс Морисита (соответствует индексу Чекановского–Съеренсена ICS форма f). Этот показатель не связан с определенными типами распределения видов по обилию, удобен при сравнении выборок разного объема, взятых случайно из искусственного сообщества, имеет высокую стабильность оценок (индекс Чекановского–Съеренсена ICS в формах a и b повышается с увеличением объема сравниваемых выборок) и учитывает вероятности встреч видов в сравниваемых сообществах [13]. Кластеризацию проводили методом Варда, применимым для нечетких множеств, а затем альтернативным методом k-средних. Далее матрицу полученных данных трансформировали в частотную матрицу в форме таблицы сопряженности, которая, как правило, служит “входом” для анализа соответствий.
Структурированность группировок после ранжирования их видовых списков по убыванию численности оценивали по кривым доминирования [9]. Предварительно значения абсолютной численности были трансформированы по шкале корней квадратных [13].
Установление достоверной связи между погруженными гидрофитами и структурой перифитонных группировок Bacillariophyta в цели данной работы не входило.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
По гидрофизическим и гидрохимическим показателям оз. Большое Миассово характеризуется как типичный мезотрофный водоем с признаками олиготрофии [15]. В крупных заливах, например Липовая курья, Штанная курья, Няшевская курья, и у северного берега по некоторым показателям (содержанию ${\text{NH}}_{4}^{ + },$ ${\text{PO}}_{4}^{{3 - }},$ ${\text{NO}}_{3}^{ - }$ и др.) (табл. 1) отмечены мезоэвтрофные и даже эвтрофные условия [12].
Таблица 1.
Физико-химические параметры воды оз. Большое Миассово на исследуемых пробных площадях
| Физико-химические параметры | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
|---|---|---|---|---|---|---|---|---|
| ${\text{NH}}_{4}^{ + }$, мг N/л | 0.110 | 0.112 | 0.122 | 0.112 | 0.128 | 0.138 | 0.122 | 0.231 |
| ${\text{NO}}_{2}^{ - }$, мг N/л | <0.003 | <0.003 | <0.003 | <0.003 | 0.007 | <0.003 | 0.003 | <0.003 |
| ${\text{NO}}_{3}^{ - }$, мг N/л | 0.38 | 0.20 | 0.17 | 0.17 | 0.20 | 0.20 | 0.20 | 0.15 |
| ${\text{PO}}_{4}^{{3 - }}$, мг P/л | 0.011 | 0.011 | 0.011 | 0.011 | <0.010 | <0.010 | <0.010 | <0.010 |
| Pобщ, мг P/л | 0.082 | 0.076 | 0.074 | 0.069 | 0.136 | 0.094 | 0.058 | 0.060 |
| N/P | 2.1 | 3.8 | 1.8 | 5.6 | 3.3 | 1.6 | 7.4 | 1.1 |
| О2, % насыщения | 219.0 | 116.0 | 169.4 | 208.1 | 153.6 | 190.7 | 180.0 | 182.0 |
| Цветность, град. Cr-Co | 29.0 | 28.0 | 29.0 | 30.5 | 43.5 | 33.0 | 27.5 | 29.0 |
Для выяснения степени однородности данного перифитонного сообщества методом кластеризации была построена иерархическая структура. В ней 27 описаний агрегировались (расстояние объединения 0.6) в три кластера, или группировки: описания 15–17, 19–23 образовали I кластер, 5, 6, 8, 13, 14, 25, 26 – II кластер, 1–4, 7, 9–12, 18, 24, 27 – III кластер (рис. 2).
Рис. 2.
Дендрограмма объединения флористических описаний Bacillariophyta (индекс Морисита, метод Варда).
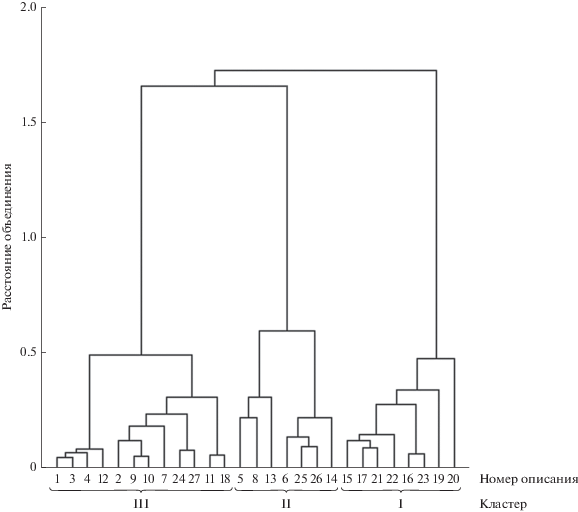
Затем альтернативным способом (методом k-средних с заданным числом k = 3 и k = 4) определяли наиболее устойчивые описания, формирующие центры кластеров. В первом случае (k = 3) описания 17, 21 (из I кластера) и 25, 26 (из II кластера) сгруппировались с описаниями III кластера. Во втором случае (k = 4) описания 6, 14, 25, 26 (из II кластера) и 1, 3 (из III кластера) сформировали IV кластер. Альтернативный способ кластеризации позволил установить, что при заданных значениях k = 3 и k = 4 есть описания, сохраняющие свои позиции в кластерах (устойчивые), и есть такие, которые их меняют (неустойчивые), т.е. переходят из одного кластера в другой. В состав IV кластера входят только неустойчивые описания. При дальнейшем анализе описания, изменяющие положение в кластерах, не учитывали, из них четыре описания (6, 14, 21, 26) соответствовали ПП 6 (залив Няшевская курья). Описания, не изменяющие своего положения при кластеризации, образовали центры I кластера (15, 16, 19, 20, 22, 23), II (5, 8, 13), III (2, 7, 9–12, 18, 24, 27).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В составе устойчивых группировок было выявлено 65 видов диатомовых водорослей, относящихся к 33 родам и 20 семействам, организованным по системе Раунда [31]. Сем. Cymbellaceae включало пять родов, Fragilariaceae и Achnanthidiaceae содержали по четыре рода, Gomphonemataceae, Catenulaceae и Rhopalodiaceae – по два рода, остальные 14 семейств – по одному роду. По количеству видов наиболее представительны были три рода: Navicula (семь видов), Nitzschia и Gomphonema (по пять видов). Остальные роды содержали один–три вида. Из таблицы сопряженности (табл. 2) видно, что в формировании устойчивых группировок принимали участие диатомовые водоросли, отмеченные как на разных видах гидрофитов, так и в различных местообитаниях.
Таблица 2.
Абсолютная частота встречаемости диатомовых водорослей в устойчивых группировках на погруженных гидрофитах в различных местообитаниях
| Кластер | Вид гидрофитов | Местообитаниe | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| M. sibirica | P. perfoliatus | P. lucens | S. aloides | 1 | 2 | 3 | 4 | 5 | 7 | 8 | |
| I | 0 | 39 | 103 | 0 | 0 | 0 | 0 | 23 | 0 | 47 | 47 |
| II | 54 | 22 | 0 | 0 | 0 | 0 | 0 | 47 | 0 | 0 | 29 |
| III | 41 | 88 | 25 | 36 | 18 | 39 | 47 | 0 | 0 | 19 | 21 |
Исходя из этого, можно предположить, что на структуру перифитонного сообщества Bacillariophyta оказывали влияние “скрытые” градиенты среды, для выявления которых был использован анализ соответствий (рис. 3).
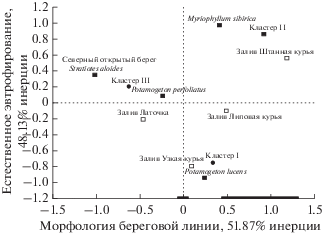
Интерпретацию осей проводили на основе вкладов, которые каждая точка вносила в инерцию рассматриваемой оси. По отношению к горизонтальной оси это были следующие точки, расположенные в порядке убывания значения вклада: залив Штанная курья (1.1856), кордон (‒1.0115), НПБ (–1.0115), залив Зимник (–1.0115), залив Липовая курья (0.4961), залив Латочка (‒0.4609). При этом первая и пятая точки имели положительные координаты по оси, а вторая, третья, четвертая и шестая – отрицательные. Координаты точек кордона, НПБ и залива Зимник, представляющих открытые участки северного берега, на графике совпадали. Для “положительных” точек характерны замкнутые очертания береговой линии и слабый уклон дна, для “отрицательных” – открытые берега с более резким уклоном. Следовательно, полюса горизонтальной оси можно было определить как “открытый плес” – “заливы”, а руководящий латентный фактор – как “морфология береговой линии”.
Аналогично, учитывая значения вкладов, знак координат и экологические условия соответствующих точек (залив Узкая курья (–0.7903), залив Штанная курья (0.5680), кордон (0.3524), НПБ (0.3524), залив Зимник (0.3524)), противоположные полюса вертикальной оси можно интерпретировать как “мезотрофия”–“эвтрофия”, а скрытый градиент среды – как “естественное эвтрофирование”. Вдоль этой оси хорошо выстраиваются точки Potamogeton lucens (–0.9389) и Myriophyllum sibirica (0.9813) у отрицательного и положительного полюсов соответственно, точка Stratiotes aloides (0.3524) занимает промежуточное положение. Так как горизонтальная ось объясняет максимальную долю инерции (~52%), то в составе группировок должны наблюдаться наибольшие различия по фактору “морфология береговой линии”. Вертикальную ось (“Естественное эвтрофирование”) можно считать производной горизонтальной оси, поскольку гидрохимические, гидрофизические, гидробиологические и другие процессы зависят от морфометрических особенностей озерной котловины.
Таким образом, были выделены три группировки диатомовых водорослей, которые по основному градиенту среды (морфологии береговой линии) распределились следующим образом: группировка III (кластер III) более характерна для открытой береговой линии, группировка I (кластер I) в основном приурочена к мезотрофным водам заливов, группировка II (кластер II) предпочитает более эвтрофные условия в заливах. Они насчитывают 47, 39 и 49 видов соответственно. Сравниваемые группировки содержали 32–36 общих таксонов и обладали значительным видовым сходством по индексу Чекановского–Съеренсена ICS = 0.70–0.82 (без учета обилия видов). В группировке I были встречены (по убыванию численности) Achnanthidium minutissimum (Kützing) Czarnecki, Cocconeis placentula Ehrenberg, Encyonopsis microcephala (Grunow) Krammer, Amphipleura pellucida (Kützing) Kützing, Epithemia sorex Kützing, Amphora pediculus (Kützing) Grunow, Staurosira construens Ehrenberg, Pseudostaurosira brevistriata (Grunow) Williams et Round, Planothidium lanceolatum (Brébisson ex Kützing) Lange-Bertalot, Aneumastus minor Lange-Bertalot, Cavinula scutelloides (W. Smith) Lange-Bertalot, Amphora ovalis (Kützing) Kützing, Cymbella cymbiformis C. Agardh, Diploneis parma Cleve и др., в группировке II – Staurosira construens, Achnanhidium minutissimum, Cocconeis placentula, Planothidium lanceolatum, Pseudostaurosira brevistriata, Staurosira binodis, Epithemia adnata (Kützing) Brébisson, E. sorex, Halamphora thumensis (A. Mayer) Levkov, Cymatopleura solea (Brébisson) W. Smith и др.; в группировке III – Cocconeis placentula, Achnanhidium minutissimum, Epithemia adnata, E. sorex, Amphora pediculus, Encyonopsis microcephala, Encyonema silesiacum (Bleisch) D.G. Mann, Staurosira construens, Pseudostaurosira brevistriata, Caloneis silicula (Ehrenberg) Cleve, Eunotia pectinalis (Kützing) Rabenhorst, Cavinula cocconeiformis (Gregory) D.G. Mann et Stickle, Reimeria sinuata (Gregory) Kociolek et Stoermer и др. Значения индекса Морисита Iδ, вычисленные с учетом абсолютной численности видов, варьировали от 0.29 до 0.96. Следовательно, основные различия наблюдались не в видовом составе группировок, а в количественном преобладании видов, т.е. в структуре доминирования.
Для оценки структурированности группировок после ранжирования их видовых списков по убыванию численности были получены кривые доминирования [9]. Они описываются экспоненциальными уравнениями:
где а – константа, определяет наклон кривой и имеет экологический смысл (чем резче выражено доминирование, тем круче наклон кривой, тем больше значение а). Константу a можно считать вполне объективным критерием выраженности доминирования в сообществе и показателем сложности его структуры. Показатель доминирования (ПД) удобнее использовать в виде обратной величины – коэффициента структурированности (КС): КС = 1/ПД × 100 (табл. 3). Чем больше величина КС, тем сложнее структура группировки [14].Таблица 3.
Структура доминирования перифитонных группировок Bacillariophyta в оз. Большое Миассово
| Кластер | Доминант | Субдоминант | Показатели доминирования | |
|---|---|---|---|---|
| a | КС | |||
| I | Achnanthidium minutissimum | C. placentula | 42.6–137.2 | 0.73–2.35 |
| II | Staurosira construens f. venter | A. minutissimum, S. construens, C. placentula, Pseudostaurosira brevistriata |
79.5–123.5 | 0.81–1.26 |
| III | Cocconeis placentula | A. minutissimum | 33.9–102.4 | 0.98–2.95 |
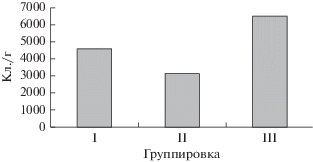
В доминантный комплекс группировок вошли фактически одни и те же виды (табл. 3), которые могли играть роль доминантов или субдоминантов. Вероятно, на эти различия в структуре доминирования мог оказывать влияние другой градиент среды – естественное эвтрофирование (рис. 3), вдоль которого группировки располагались следующим образом: I < II < III. С повышением трофности отмечался рост значений КС между I и II группировками (табл. 3), что подтверждается данными исследований о структуре зоопланктона [14]. Однако ожидаемое увеличение значений КС у группировки III не наблюдалось. Ее слабую структурированность и высокое число субдоминантов можно объяснить физико-химическими особенностями залива Штанная курья, к которому она приурочена. Этот мелководный залив с торфяно-илистым дном расположен в понижении рельефа и окружен болотными сообществами. Низкое содержание доступных для водорослей фосфатов на фоне повышенного общего фосфора, преобладание аммонийных форм азота над нитратными, высокая цветность, более низкое содержание кислорода (табл. 1), низкие общая численность диатомей (рис. 4) и видовое богатство в II группировке свидетельствовали о дистрофных условиях в заливе. В условиях дефицита доступных форм питательных веществ сформировались группировки, сходные по структурированности с сообществами олиготрофных вод. Кроме того, при дистрофировании из-за низкой прозрачности и высокой цветности воды возник дефицит другого ресурса – света, что также привело к повышению межвидовой конкуренции.
Выводы. Один из ведущих факторов, играющих роль в организации перифитонного сообщества Bacillariophyta оз. Большое Миассово, – морфология береговой зоны, а именно степень замкнутости берегов и уклон дна. На показатели структурированности сообщества влияют изменения физико-химических параметров озерной воды, вызванные процессами естественного эвтрофирования и дистрофирования. Исследованное перифитонное сообщество Bacillariophyta, развивающееся на погруженных гидрофитах, слабо структурировано. Доминирующий комплекс выявленных группировок схож по видовому составу и отличается лишь количественным преобладанием в нем видов. Эти изменения отражают приспособление сообщества диатомей к локальным условиям биотопа.
Список литературы
Андреева М.А. Озера Среднего и Южного Урала. Челябинск: Южно-Уральск. кн. изд-во, 1973. 269 с.
Бегун А.А. Биоиндикация состояния морской среды по диатомовым водорослям эпифитона макрофитов (залив Петра Великого, Японское море) // Изв. Тихоокеан. н.-исслед. рыб. центр. 2012. Т. 169. С. 1–17.
Бегун А.А., Рябушко Л.И., Звягинцев А.Ю. Влияние поверхности субстрата и времени его экспозиции в море на количественное развитие диатомовых водорослей перифитона // Изв. ТИНРО. 2010. Т. 163. С. 240–263.
Диатомовые водоросли СССР (ископаемые и современные). Л.: Наука, 1974. Т. 1. 403 с.
Диатомовые водоросли СССР (ископаемые и современные). Л.: Наука, 1988. Т. 2. Вып. 1. 116 с.
Диатомовые водоросли СССР (ископаемые и современные). СПб.: Наука, 1992. Т. 2. Вып. 2. 125 с.
Комулайнен С.Ф. Методические рекомендации по изучению фитоперифитона в малых реках. Петрозаводск: Карельск. науч. центр РАН, 2003. 43 с.
Ландшафтный фактор в формировании гидрологии озер Южного Урала. Л.: Наука, 1978. 248 с.
Методика изучения биогеоценозов внутренних водоемов. М.: Наука, 1975. 240 с.
Неронов В.В. Развитие концепции экотонов и их роль в сохранении биологического разнообразия // Успехи соврем. биологии. 2001. Т. 121. № 4. С. 323–336.
Общие закономерности возникновения и развития озер. Методы изучения истории озер. Серия: История озер СССР. Л.: Наука, 1986. 254 с.
Оксиюк О.П., Давыдов О.А. Методологические принципы оценки состояния водных объектов по микрофитобентосу // Гидробиол. журн. 2006. Т. 42. № 2. С. 98–112.
Песенко Ю.А. Принципы и методы количественного анализа в фаунистических исследованиях. М.: Наука, 1982. 282 с.
Рогозин А.Г. Особенности структурной организации зоопланктонного сообщества в озерах разного трофического статуса. Видовые популяции // Экология. 2000. № 6. С. 438–443.
Экология озера Большое Миассово. Миасс: Ильменск. гос. заповедник Уральск. отд. РАН, 2000. 318 с.
Cattaneo A., Galanti G., Gentinetta S., Romo S. Epiphytic algae and macroinvertebrates on submerged and floating-leaved macrophytes in an Italian lake // Freshwater Biol. 1998. V. 39. № 4. P. 725–740.
Gosz J.R. Landscape boundaries consequence for biotic diversity landscape flows // Ecological studies. N.Y.: Springer-Verlag, 1992. P. 1–55.
Gross E.M., Feldbaum C., Graf A. Epiphyte biomass and elemental composition on submersed macrophytes in shallow eutrophic lakes // Hydrobiologia. 2003. V. 506. № 1. P. 559–565.
Hutchinson G.E. A Treatise on Limnology // Limnological Botany. N.Y.: Wiley and Sons, 1975. V. 3. P. 1–660.
Khursevich G.K., Kociolek J.P. A preliminary worldwide inventory of the extinct freshwater fossil diatoms from the orders Thalassiosirales, Stephanodiscales, Paraliales, Aulacoseirales, Melosirales, Coscinodiscales and Biddulphiales // Nova Hedwigia. 2012. Beih. 141. P. 315–364.
Kiss M.K., Lakatos G., Borics G., Gidó Z., Deák C. Littoral macrophyte-periphyton complexes in two Hungarian shallow waters // Hydrobiologia. 2003. V. 506. P. 541–548.
Krammer K., Lange-Bertalot H. Bacillariophyceae. Teil 1: Naviculaceae // Süsswasserflora von Mitelleuropa. Jena: Gustav Fischer Verlag, 1986. Bd 2. 876 s.
Krammer K., Lange-Bertalot H. Bacillariophyceae. Teil 2: Bacillariaceae, Epithemiaceae, Surirellaceae // Süsswasserflora von Mitteleuropa. Stuttgart; N.Y.: Gustav Fischer Verlag, 1988. Bd 2. 596 s.
Krammer K., Lange-Bertalot H. Bacillariophyceae. Teil 3: Centrales, Fragilariaceae, Eunotiaceae // Süsswasserflora von Mitteleuropa. Stuttgart; Jena: Gustav Fischer Verlag, 1991. Bd 2. 576 s.
Krammer K., Lange-Bertalot H. Bacillariophyceae. Teil 4: Achnanthaceae, Kritische Ergänzungen zu Navicula (Lineolatae) and Gomphonema // Süsswasserflora von Mitteleuropa. Stuttgart; Jena: Gustav Fischer Verlag, 1991. Bd 2. 436 s.
Lange-Bertalot H. Diatoms of Europe. Navicula sensu stricto, 10 genera separated from Navicula sensu lato Frustulia. . Rugell: A.R.G. Gantner Verlag K.G., 2001. V. 2. 526 p.
Laugaste R., Reunanen M. The composition and density of epiphyton on some macrophyte species in the partly meromictic Lake Verevi // Hydrobiologia. 2005. V. 547. P. 137–150.
Liboriussen L., Jeppesen E. Structure, biomass, production and depth distribution of periphyton on artificial substratum in shallow lakes with contrasting nutrient concentrations // Freshwater Biol. 2006. V. 51. № 1. P. 95–109.
Messyasz B., Kuczynska-Kippen N. Periphytic algal communities: a comparison of Typha angustifolia L. and Chara tomentosa L. beds in three shallow lakes (West Poland) // Poland J. Ecol. 2006. № 54. P. 15–27. https://doi.org/10.1111/j.1365-2427.2005.01481.x
Pip E., Robinson G.G.C. A comparison of algal periphyton composition on eleven species of submerged macrophytes // Hydrobiol. Bul. 1981. № 18. P. 109–118.
Round F.E., Crawford R.M., Mann D.G. The Diatoms. Biology and Morphology of the Genera. Cambridge: Cambridge Univ. Press, 1990. 747 p.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод