

Биология внутренних вод, 2019, № 2-2, стр. 12-19
Побегообразование у горца земноводного Persicaria amphibia (L.) Delarbre с позиций модульной организации
Е. В. Лелекова 1, *, Н. П. Савиных 1
1 Вятский государственный университет
610000 Киров, ул. Московская, 36, Россия
* E-mail: LelekovaEV1980@mail.ru
Поступила в редакцию 07.02.2018
После доработки 12.07.2018
Принята к публикации 01.08.2018
Аннотация
Проведен биоморфологический анализ трех биоморф Persicaria amphibia (L.) Delarbre, отличающихся особенностями побегообразования в разных условиях среды с позиций системного подхода и модульной организации. Побеговые системы всех биоморф формируются по симподиальной длиннопобеговой модели побегообразования и сложены однотипными элементарными модулями. Универсальные модули в различных местообитаниях представлены монокарпическими побегами возобновления, силлептическими, а также побегами с неполным циклом развития. Вариабельность побеговых систем определяется модульной организацией и демонстрирует пластичность габитуса P. amphibia.
ВВЕДЕНИЕ
К настоящему времени достаточно хорошо изучены биоморфологические особенности и индивидуальное развитие многих наземных трав. Водные и прибрежно-водные растения рассматриваются, в основном, в течение последнего столетия [2, 5, 8–10, 12, 25 и др.]. Показано, что многие из них способны существовать в местах с различными условиями увлажнения и очень пластичны: на основе одного генотипа формируются особи с разными габитусами. Поэтому, в отличие от травянистых мезофитов и ксерофитов, прибрежно-водные растения часто встречаются в виде нескольких биоморф. Эта особенность обеспечивает им возможность продолжать развитие в меняющихся условиях, длительно сохраняться на однажды занятых территориях и расселяться в пространстве и времени.
Изучение структурной организации и биологии растений с широким спектром местообитаний и расширение представления о механизмах и способах адаптации организмов – одна из основных задач биоморфологии и экологии в целом. Поэтому объектом исследования выбран горец земноводный Persicaria amphibia (L.) Delarbre. Он произрастает в воде, в условиях прибрежий, по берегам среди травянистой растительности и на суше. Особи водной формы встречаются чаще, именно они цветут и способны обеспечивать семенное воспроизведение ценопопуляций.
Цель работы – изучение и сравнение биоморфологии особей P. amphibia, растущих в воде и на суше, для оценки приспособленности к жизни в разнообразных по степени увлажнения/обводнения и уровня воды водоемах.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
P. amphibia – голарктический вид [4]. Материал для изучения собирали в ходе обследований прибрежных мелководий, водоемов со стоячей и медленно текущей водой, берегов водоемов и водотоков различного типа с прилегающими к ним влажными, обычно затопляемыми лугами, в центральных и южных районах Кировской обл.
Для уточнения деталей структурной организации побеговых систем дополнительно использовали образцы Гербария Института биологии внутренних вод им. И.Д. Папанина РАН и гербарной коллекции кафедры биологии и методики обучения биологии Вятского государственного университета.
Биоморфологический анализ побеговых систем P. amphibiа проводили с учетом представлений о растении как целостном организме [27]. Структуру особей рассматривали с позиций системного подхода [19] и модульной организации [15, 16]. Для этого использовали представления о трех категориях модулей и разнообразии их вариантов. Основной модуль, пространственно-временную структуру, формирующуюся на основе целого универсального модуля или его части и закономерно повторяющуюся в строении зрелых генеративных особей, у этого вида сложно охарактеризовать в виду ранней морфологической дезинтеграции и трудностей изъятия целостного организма, особенно из донного грунта. Универсальный модуль – одноосный побег, образованный в результате деятельности одной апикальной меристемы путем последовательного формирования элементарных модулей. Элементарный модуль – участок побега с узлом, листом, пазушной почкой (или ее производным) и нижележащим междоузлием.
Основные методы исследования: разработанный И.Г. Серебряковым [19, 21] на базе сравнительно-морфологического анализа способ описания конкретных структур; ритмологический [18, 20]; биоморфологический [23].
Особенности побеговых систем оценивали по аналогии с таковыми у трав мезофитов: строение монокарпических побегов, степень их структурно-функциональной зональности (по [3] и [30]), развитие их (по [20] и [22]), тип и механизм морфологической дезинтеграции (по [24]).
Габитус растения характеризовали в соответствии с предложенным ранее алгоритмом описания жизненных форм водных и прибрежно-водных растений [16]: с учетом числа плодоношений, длительности жизни, степени вегетативной подвижности и воздействия особей на среду обитания, длительности жизни надземных осей и листьев, типа подземных органов у многолетников и малолетников вегетативного происхождения, типа побега по длине междоузлий и положению в пространстве. Жизненная форма оценена также с учетом подходов Раункиера [28].
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
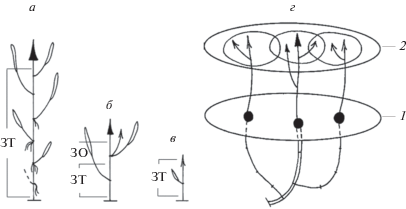
Водная форма P. amphibiа встречается в медленно текущих или стоячих водах: реках, старицах, озерах, прудах. Побеговая система образуется в результате симподиального нарастания и ветвления исходного побега (рис. 1). Она состоит из удлиненных моно- и дициклических монокарпических побегов возобновления (МПВ), монокарпических силлептических побегов двух типов (МСП-1 и МСП-2) и побегов с неполным циклом развития (ПНЦ), не переходящих в фазу бутонизации, цветения и плодоношения. Все вышеперечисленные побеги отличаются по периоду покоя почек, из которых образуются.
Рис. 1.
Монокарпические побеги возобновления (МПВ) и силлептические побеги (МСП) P. amphibiа. 1 – соцветие; 2 – вегетативный побег; 3 – почка в пазухе чешуевидного листа; 4 – погруженный лист; 5 – плавающий лист; 6 – придаточные корни; 7 – многолетние структуры; 8 – однолетние структуры; 9 – терминальное соцветие МПВ n-го порядка; 10 – МСП-2 n + 1-го порядка; 11 – вегетативный силлептический побег n + 2-го порядка; 12 – МСП-1 n + 1-го порядка; 13 – МПВ n + 1-го порядка в стадии геофильного побега; 14 – геофильный участок МПВ n-го порядка; ЗВ – зона возобновления; СЗТ-1 – средняя зона торможения-1; СЗТ-2 – средняя зона торможения-2; ЗО – зона обогащения; ГС – главное соцветие.
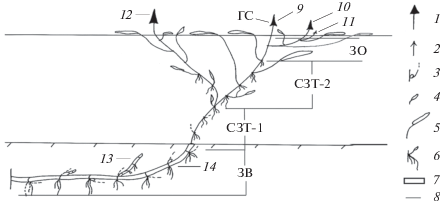
МПВ формируются из почек регулярного возобновления на геофильном участке такого же побега предыдущего порядка ветвления. Фаза почки продолжается от 10 мес до ≥1 года. Анализ побеговых систем и их развития показал, что в первый год у одного МПВ трогаются в рост, в основном одна–три почки на геофильном участке, остальные остаются в покое.
Фаза геофильного побега длится 11–12 мес. В это время из почки возобновления формируется геофильный участок до ≥50 см длиной и ~0.6 см толщиной. На верхушке располагается небольшая промежуточная почка, покрытая чешуевидными листьями. Эта часть МПВ сложена однотипными элементарными модулями с длинным междоузлием и чешуевидным листом, в пазухе которого находится почка. В виде такого геофильного побега с промежуточной почкой МПВ зимует (рис. 2).
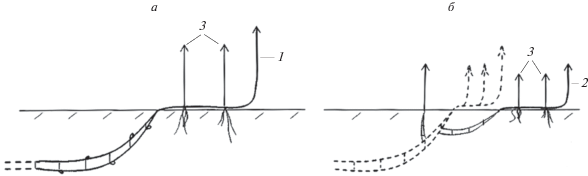
Рис. 2.
Развитие монокарпического побега возобновления P. amphibiа: а – конец вегетационного периода; б – середина следующего вегетационного периода. 1 – МПВ n-го порядка; 2 – МПВ n + 1-го порядка в стадии геофильного побега; 3 – МПВ n + 1-го порядка; 4 – ПНЦ n+1-го порядка.
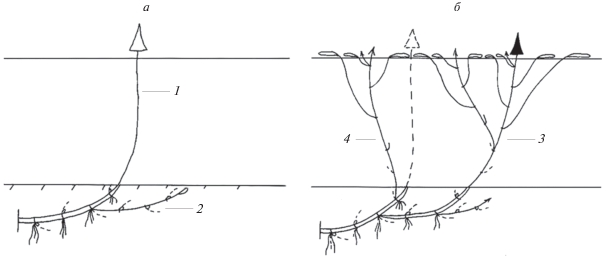
Этот участок в будущем обеспечивает возобновление растения. При рассеянном и разновременном образовании побегов он принимает на себя функции зоны возобновления. Одновременно у него сохраняются функции поглощения и запаса питательных веществ, закрепления в почве и вегетативного расселения.
Фаза вегетативного ассимилирующего побега непродолжительна. С наступлением весны геофильный побег выходит на поверхность субстрата, развиваются метамеры внутриводного участка МПВ. Главная функция этой части – вынос побега на поверхность воды. Его размер определяется глубиной водоема и регулируется длиной междоузлий: от 3–6 до 45 см. Все элементарные модули имеют длинные междоузлия, но различаются по наличию/отсутствию неветвящихся темно-бурых или красноватых стеблеродных придаточных корней (за счет них растение укореняется при полегании под тяжестью побеговых систем и/или снижения уровня воды), развитию почек и их производных, строению листьев. МПВ P. amphibiа свойственна гетерофиллия. Базальные метамеры внутриводного участка у сформированного побега имеют почти разрушенные стеблеобъемлющие чешуевидные листья 0.5–2.0 см длиной. Пазушные почки в побеги, как правило, не реализуются. Это средняя зона торможения (СЗТ-1). Выше по стеблю расположены метамеры с черешковыми листьями с небольшими тонкими листовыми пластинками, которые уже в середине вегетационного периода полностью разрушаются. Такие метамеры включаются в СЗТ-2, которая дополнительно принимает на себя функцию зоны обогащения: из пазушных почек иногда образуются боковые побеги. Они подобны по строению МПВ, но не имеют геофильного участка и возникают из почек обогащения без периода покоя. Это – МСП-1 (рис. 1). Четыре–шесть самых ближних к поверхности воды метамеров содержат длинночерешковые листья с плавающими блестящими продолговатыми листовыми пластинками, иногда они приподнимаются над поверхностью воды.
Рис. 3.
Фрагмент внутриводного участка МПВ P. amphibiа. 1 – апикальный участок МПВ n-го порядка; 2 – соцветие МПВ n-го порядка; 3 – МСП-2 n+1-го порядка; 4 – почка, реализующаяся в МСП-2 n+2-го порядка.
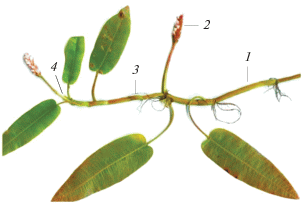
Моноподиальное нарастание МПВ завершается образованием терминального соцветия. Одновременно с фазой бутонизации, цветения и плодоношения его из самой верхней почки под соцветием (зона обогащения) образуется МСП-2 – небольшой побег, по строению аналогичный апикальному участку МПВ с листьями срединной формации (рис. 1, рис. 3). Пазушная почка самого верхнего метамера этого побега также развивается в МСП-2 с меньшим числом метамеров. В результате формируется система из МСП-2 двух–четырех порядков ветвления. Не все МСП-2 успевают сформировать соцветие, функционируя как ПНЦ. Последовательное образование побегов разных порядков у P. amphibiа – один из вариантов итеративного ветвления, свойственного многим водным и прибрежно-водным травам [5, 11, 17 и др.].
В конце осени надводные и внутриводные участки большинства МПВ полностью отмирают. Часть из них основаниями полегают на дно и укореняются. Пазушные почки остаются живыми и могут тронуться в рост следующей весной. При условии укоренения и успешной перезимовки участки МПВ с системами из МСП-2 служат неспециализированными диаспорами для вегетативного размножения, как показано в опытах М.В. Маркова [12].
Далее наступает фаза вторичной деятельности, когда геофильный участок становится резидом недолго живущего гипогеогенного корневища (рис. 4). Морфологическая дезинтеграция особей – ранняя, полная.
Рис. 4.
Фрагмент симподиально нарастающего гипогеогенного корневища P. amphibiа. 1 – геофильные участки МПВ с верхушечными почками, зимующие в толще грунта; 2 – отмершие и разрушенные метамеры анизотропных побегов; n, n + 1, n + 2 – геофильные участки побегов последовательных порядков ветвления.
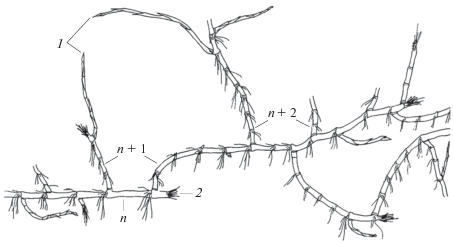
Ветвление МПВ P. amphibiа растянуто во времени и значительно повышается за счет разновременной реализации пазушных почек резидов. В первый год развития реализуются одна–три почки геофильного участка исходного МПВ. На следующий год дополнительно возможно формирование одного–трех МПВ или ПНЦ. Из почек этой зоны, находящихся в состоянии покоя более двух лет, также формируются типичные МПВ. Так, в третий и четвертый годы на основе резида исходного МПВ развиваются до шести–семи побегов следующего порядка (МПВ и ПНЦ). В результате разновременного образования МПВ и повторного освоения занятой территории формируется центр закрепления, подобный парциальному кусту многолетних наземных трав. Эту структуру условно можно считать основным модулем в побеговой системе P. amphibiа. Такое побегообразование представляет собой один из резервов сохранения растения и способ поддержания ценопопуляций в виде неоднократного освоения занятой территории; способствует закреплению парциальных структур и целостных особей в грунте, обеспечивая произрастание растения в водотоках.
Подобное часто встречается у водных трав: ежеголовника [1], кубышки желтой [25] и др. По-видимому, удлинение фазы почки у гидрофитов – следствие конкуренции за пространство водной поверхности. В качестве компенсации задержки развития МПВ в фазе почки у них формируются МСП-2 и ПНЦ у поверхности воды: на образование их требуется меньше пластических веществ при одинаковом результате – обеспечении успешной репродукции.
Таким образом, основa побегового тела P. amphibiа – анизотропные моно- или дициклические МПВ, МСП-1, МСП-2 и ПНЦ с разными по числу метамеров вегетативными участками. Побеги всех этих типов – универсальные модули (рис. 1, рис. 5а–5в).
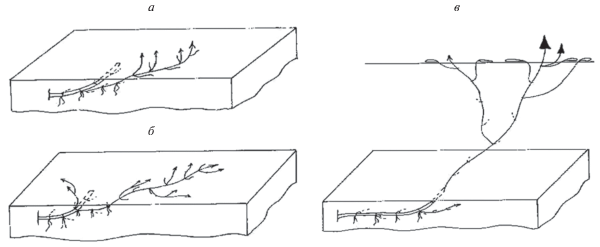
Рис. 5.
Типы силлептических побегов и центры воздействия на среду отдельной особи P. amphibiа: а – МСП-1; б – МСП-2 (a и б могут быть ПНЦ); в – силлептический ПНЦ; г – центры воздействия на среду (1 – уровень поверхности грунта, 2 – поверхность воды). ЗТ – зона торможения (пазушные почки не реализуются либо крайне редко), ЗО – зона обогащения.
На поверхности донного грунта можно различить явно выраженные центры воздействия на среду особей P. amphibiа: растение типичное явнополицентрическое (рис. 5г, позиция 1). Однако, на поверхности воды в результате развития МСП-2 с длинночерешковыми листьями, эти центры перекрываются, и особь принимает вид ацентрической: иногда поверхность водоема сплошь покрыта листьями.
На основании вышесказанного, биоморфа водных растений P. amphibiа определена как поликарпический явнополицентрический на поверхности субстрата и ацентрический на поверхности воды вегетативно подвижный длиннопобеговый летне-зеленый травянистый малолетник вегетативного происхождения с ранней полной специализированной и неспециализированной морфологической дезинтеграцией и анизотропными системами МПВ из МСП-1, МСП-2 и ПНЦ; гидрогеофит. Поддержание ценопопуляций, в основном, вегетативное.
Наземная форма P. amphibiа встречается нечасто на песчаных и глинистых берегах, в посевах, на пойменных лугах, по обочинам дорог и обсыхающих днищах временных водоемов после спуска воды. Авторами рассмотрены два варианта развития побеговых систем у наземной биоморфы P. amphibiа: у особей, произрастающих на лугу и песчаном берегу водоема. В обоих случаях они формируются в результате симподиального нарастания.
На лугах побеговые системы растений сложены, в основном, стелющимися и приподнимающимися ПНЦ двух типов: с геофильным участком и без него (рис. 6). Первые – анизотропные, полегающие основанием и укореняющиеся. По-видимому, это незацветающие аналоги МПВ водной формы.
Рис. 6.
Побеговые системы наземной биоморфы P. amphibiа на основе исходного побега с геофильным участком: а – 1-й год развития; б – 2-й год развития. 1 – ПНЦ n-го порядка; 2 –ПНЦ n + 1-го порядка; 3 – силлептические ПНЦ.
Геофильные участки этих побегов сложены однотипными элементарными модулями: с длинным междоузлием, чешуевидным листом и почкой с одним периодом покоя. Как и у некоторых прибрежно-водных трав [17], они служат для расселения, закрепления особей в почве и ветвления. Элементарные модули надземного участка также однотипны: с длинным междоузлием, почти сидячими листом срединной формации и почкой, либо вегетативным побегом следующего порядка – силлептическим ортотропным ПНЦ с меньшим числом метамеров, укореняющимся при полегании на субстрат (рис. 6). По своему строению они подобны МСП-1, но не ветвятся в апикальной части и не переходят к цветению. Таким образом, в течение вегетационного периода на основе исходного побега формируется система из ПНЦ до трех порядков ветвления. Укореняющиеся вегетативные участки могут заноситься песком во время половодья, перезимовывать, пассивно погружаясь в грунт или среди травяной ветоши, и выполнять функции зоны возобновления, обеспечивая возобновление растения. Поддержание ценопопуляций – только вегетативное.
По-видимому, в стратегии специализированного патиента [14] P. amphibiа способна существовать в течение длительного времени в луговых ценозах по берегам рек. Биоморфа растения в этих условиях – поликарпик; ацентрический вегетативно-подвижный длиннопобеговый летнезеленый травянистый малолетник/однолетник вегетативного происхождения с ранней полной специализированной (за счет партикуляции в области геофильных участков) и неспециализированной (за счет партикуляции в области надземных участков) морфологической дезинтеграцией в виде побеговых систем двух–трех порядков ветвления из анизотропных и ортотропных ПНЦ; геофит, протогемикриптофит.
На песках вблизи уреза воды побеговые системы растений отличаются значительными размерами. Здесь выделена крупная заросль P. amphibia площадью ~2 м2, растущая в направлении от берега к воде. Других растений рядом не обнаружено. Возможно, сформировавшиеся в прошлом году участки наземных побеговых систем были занесены песком во время весеннего половодья. По выходу побега из почки геофильного участка на поверхность субстрата в результате рассеянного ветвления формируется надземная побеговая система трех и более порядков ветвления из плагиотропных укореняющихся ПНЦ. Поэтому на песках растение существует как и летне-зеленый малолетник/однолетник вегетативного происхождения, но в виде побеговых систем из плагиотропных ползучих на поверхности ПНЦ; геофит, протогемикриптофит. В отличие от растений в составе луговых ценозов, у этих особей более значительны вегетативное разрастание и размножение. Укоренившиеся верхушки побегов всех порядков ветвления после ранней морфологической дезинтеграции функционируют как неспециализированные диаспоры и обеспечивают самоподдержание ценопопуляций при отсутствии семенного воспроизведения, особенно после засыпания их песком на берегу, и пассивное расселение при переносе во время половодья. За счет развития побегов из почек на геофильных участках возможно перемещение растения в водоем.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В зависимости от условий среды P. amphibiа представлена разными жизненными формами (рис. 7). Их анализ с учетом строения побегов универсального модуля, типа ветвления и его порядков, длительности жизни геофильных частей, способности к образованию диаспор, способа нарастания при формировании побеговых систем, морфологической дезинтеграции и типа биоморфы по способу воздействия на среду показал следующее.
Рис. 7.
Биоморфы P. amphibiа в различных местообитаниях: а – на лугах; б – на песчаных прибрежьях; в – в водоемах.
Во всех условиях у побегов, на основе которых формируются разветвленные побеговые системы, имеются геофильные участки, входящие в состав недолгоживущих гипогеогенных корневищ. Морфологическая дезинтеграция особей ранняя полная. Вегетативное размножение происходит за счет специализированной дезинтеграции при разрушении геофильных участков побегов и за счет неспециализированной – при распаде надгрунтовых участков с образованием неспециализированных диаспор. P. amphibiа – малолетник вегетативного происхождения. Такая биоморфа в условиях водной среды определена для многих водных и прибрежно-водных растений [6, 7, 9, 11, 13, 17, 29 и др.].
Все побеговые системы формируются по симподиальной длиннопобеговой модели побегообразования. Способ нарастания при этом бази-, мезо- и акросимподиальный. Основа побеговых систем – монокарпические побеги возобновления. МСП и ПНЦ выполняют, в основном, функции разрастания особей и увеличения ассимилирующей поверхности. В зависимости от условий среды биоморфы P. amphibiа, как и многих растений [26], сложены разными вариантами универсального модуля. Так, у водной формы преобладают МПВ; на лугах – МПВ с неполным циклом развития и типичными силлептическими ПНЦ (ортотропными и анизотропными). На песчаных пляжах формируются более разветвленные побеговые системы такого же происхождения, но из плагиотропных укореняющихся побегов. При изменении условий местообитания возможен переход одной жизненной формы P. amphibiа в другую. Так, луговые биоморфы с ортотропными побегами при переселении на песчаные отмели могут стать плагиотропными стелющимися растениями. При продвижении в воду побегов, образующихся из геофильных участков у наземных форм, возможно возникновение биоморфы с анизотропными системами МПВ, формирующимися в этих условиях. Такая вариабельность побегообразования цветковых растений может быть связана с их модульной организацией, способностью возврата к стартовому зачатку – почке и формированию из нее только тех универсальных модулей, которые обеспечивают существование особи в конкретных условиях среды.
Выводы. P. amphibiа существует в природе в виде трех биоморф. Все они поликарпические, явнополицентрические/ацентрические, вегетативно подвижные длиннопобеговые летне-зеленые травянистые малолетники вегетативного происхождения с ранней полной специализированной и неспециализированной морфологической дезинтеграцией; гидрогеофиты – в воде, геофиты и протогемикриптофиты – на суше. Побеговые системы растений сложены однотипными элементарными и универсальными модулями. В воде они формируются на основе анизотропных МПВ и состоят из МСП-1, МСП-2 и ПНЦ. На лугах за счет ветвления и нарастания ПНЦ с геофильным участком развивается побеговая система из анизотропных и ортотропных ПНЦ; на песчаных прибрежьях – из плагиотропных ползучих ПНЦ с большей степенью ветвления. Вариабельность побеговых систем определяется модульной организацией. P. amphibiа очень лабильна и способна сохранять в генотипе весь набор элементарных и универсальных модулей с возможностью реализации лишь тех, которые соответствуют условиям среды.
Список литературы
Беляков Е.А. Биология некоторых представителей рода Sparganium L. (сем. Typhaceae): Автореф. дис. … канд. биол. наук. Сыктывкар, 2016. 21 с.
Бобров Ю.А. Жизненные формы водных трав Северо-Востока Европейской России // Arctic Environ. Res. 2017. Т. 17. № 2. С. 104–112.
Борисова И.В., Попова Т.А. Разнообразие функционально-зональной структуры побегов многолетних трав // Бот. журн. 1990. Т. 75. № 10. С. 1420–1425.
Губанов И.А., Новиков В.С., Тихомиров В.Н. Определитель высших растений средней полосы европейской части СССР. М.: Просвещение, 1981. 288 с.
Даровских Е.А. Типы монокарпических побегов Potamogeton lucens L. // Тр. VII Междунар. конф. по морфологии растений, посвященной памяти И.Г. и Т.И. Серебряковых. М.: Мос. пед. гос. ун-т, 2004. С. 76–77.
Жмылев П.Ю., Гололобова М.А. Разнообразие жизненных форм водных сосудистых растений европейской части России // Вестн. Рос. ун-та дружбы народов. Сер. Экология и безопасность жизнедеятельности. 2009. № 4. С. 5–15.
Жмылев П.Ю., Леднев С.А. Биоморфологическое разнообразие водных сосудистых растений ЗБС // Тр. Звенигород. биол. ст. им. С.Н. Скадовского. М.: МГУ, 2011. Т. 5. С. 80–85.
Жмылев П.Ю., Леднев С.А., Щербаков А.В. Биоморфология водных растений: проблемы и подходы к классификации жизненных форм // Леонид Васильевич Кудряшов Ad Memoriam: Сб. статей. М.: Макс ПРЕСС, 2012. С. 101–128.
Лапиров А.Г. Рдест гребенчатый // Биологическая флора Московской области. М.: Изд-во МГУ, 1995. Вып. 11. С. 37–55.
Лебедева О.А., Лапиров А.Г. Формирование побеговой системы, цветорасположение и модульная организация Batrachium trichophyllum (Chaix) Bosch // Матер. VI Всерос. шк.-конф. по водным макрофитам “Гидроботаника-2005”. Рыбинск: Дом печати, 2006. С. 294–298.
Лелекова Е.В. Биоморфология водных и прибрежно-водных семенных растений северо-востока Европейской России: Автореф. дис. … канд. биол. наук. Пермь, 2006. 19 с.
Марков М.В., Ключникова Н.М., Федорин А.К. Разнообразие жизненных форм и систем репродукции в роде Polygonum S.L. в аспекте вторичного перехода к водному образу жизни // Преподаватель XXI век. 2010. Т. 2. № 1. С. 207–215.
Петухова Д.Ю. Биоморфология столонно-розеточных гидрофитов: Автореф. дис. … канд. биол. наук. Сыктывкар, 2008. 19 с.
Работнов Т.А. Изучение ценотических популяций в целях выяснения стратегии жизни видов растений // Бюл. МОИП. Отд. биол. 1975. Т. 80. Вып. 2. С. 5–17.
Савиных Н.П. Модули у растений // Тез. докл. II Междунар. конф. по анатомии и морфологии растений. СПб.: Изд-во Ботанического института им. В.Л. Комарова РАН, 2002. С. 95–96.
Савиных Н.П. О жизненных формах водных растений // Гидроботаника: методология, методы: Матер. школы по гидроботанике. Рыбинск: Дом печати, 2003. С. 39–48.
Савиных Н.П., Шабалкина С.В., Лелекова Е.В. Биоморфологические адаптации гелофитов // Сиб. экол. журн. 2015. Т. 22. № 5. С. 671–681.
Серебряков И.Г. Структура и ритм в жизни цветковых растений. Ч. 2 // Бюл. МОИП. Отд. биол. 1949. Т. 54. Вып. 2. С. 47–62.
Серебряков И.Г. Морфология вегетативных органов высших растений. М.: Сов. наука, 1952. 390 с.
Серебряков И.Г. О методах изучения ритмики сезонного развития растений в геоботанических стационарах // Докл. совещ. по стационарным геобот. исследованиям. Л., 1954. С. 145–159.
Серебряков И.Г. Жизненные формы высших растений и их изучение // Полевая геоботаника. М.; Л.: Наука, 1964. Т. 3. С. 148–208.
Серебрякова Т.И., Воронин Н.С., Еленевский А.Г. и др. Ботаника с основами фитоценологии. Анатомия и морфология растений. М.: Академкнига, 2006. 543 с.
Современные подходы к описанию структуры растений. Киров: ООО “Лобань”, 2008. 355 с.
Смирнова О.В., Заугольнова Л.Б., Торопова Н.А., Фаликов Л.Д. Критерии выделения возрастных состояний и особенности хода онтогенеза у растений различных биоморф // Ценопопуляции растений: Основные понятия и структура. М.: Наука, 1976. С. 14–43.
Чернова А.М. Сезонная динамика продуктивности кубышки желтой (Nuphar lutea, Nymphaeaceae) в условиях малых рек верхнего Поволжья: Автореф. дис. … канд. биол. наук. Борок, 2013. 23 с.
Шабалкина С.В., Савиных Н.П. О поливариантности развития растений в местах с переменным обводнением // Биоморфологические исследования на современном этапе: Матер. конф. с международным участием “Современные проблемы биоморфологии”. Владивосток: Мор. гос. ун-т, 2017. С. 219–221.
Шафранова Л.М., Гатцук Л.Е. Растение как пространственно временная метамерная (модульная) система // Успехи экологической морфологии и ее влияние на смежные науки: Межвузовский сб. науч. трудов. М., 1994. С. 6–7.
Raunkiaer C. The life form of plants and statistical plant geography. Oxford: Clarendon Press, 1934. 632 p.
Santamaria L., Garcia A.I.L. Latitudinal variation in tuber production in an aquatic pseudo-annual plant, Potamogeton pectinatus // Aquat. Bot. 2004. V. 79. P. 51–64.
Troll W. Die Infloreszenzen. Jena: Fischer Verlag, 1964. Bd 1. 615 s.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод