

Биология внутренних вод, 2019, № 3, стр. 3-10
Моделирование биотического потока энергии в планктонном сообществе экосистемы озерного типа при участии микробиальных организмов
В. В. Бульон *
Зоологический институт Российской академии наук
199034 Санкт-Петербург, Университетская наб., 1, Россия
* E-mail: vboulion@mail.ru
Поступила в редакцию 24.04.2018
После доработки 18.12.2018
Принята к публикации 06.02.2019
Аннотация
Для экосистемы озерного типа построена масс-балансовая модель биотического потока энергии в планктонном сообществе с участием микробиальных организмов. Входные абиотические параметры модели: географическая широта, средняя глубина, цветность воды и содержание в воде общего фосфора. Модель сконструирована на базе опубликованных данных для Рыбинского водохранилища и озер Монголии. Анализ модели показал, что при подключении микробиальной “петли” к “традиционной” линейной трофической цепи биомасса и продукция метазойного планктона и планктоноядных рыб снижаются почти вдвое. Предполагается, что величина конечной продукции экосистемы (продукции рыб) тесно связана со степенью выраженности микробиальной “петли” на протяжении вегетационного сезона.
ВВЕДЕНИЕ
Микроорганизмы размером <100 мкм создают на базе линейной пищевой цепи так называемую микробиальную “петлю” или микробиальную пищевую сеть, исследование значения которой в биотическом потоке энергии в водных экосистемах – одно из приоритетных направлений современной гидробиологии. В микробиальную сеть входят фотосинтезирующие и гетеротрофные микроорганизмы: пико- и нанофитопланктон, гетеротрофные бактерии, флагеллаты и инфузории [19, 21, 24, 26].
Фитопланктон прижизненно или, чаще всего, в результате лизиса, фотолиза и разрушения клеток водорослей зоопланктоном теряет часть синтезированных органических веществ в растворенной форме. Потребителями автохтонного и аллохтонного РОВ служат гетеротрофные бактерии. Главная функция бактерий заключается в трансформации РОВ во взвешенное органическое вещество (ВОВ), доступное бактериофагам, а функция бактериофагов – в регенерации биогенных элементов [12–14, 16, 28].
За фитопланктон конкурируют представители нехищного мета- и протозойного планктона. Гетеротрофные жгутиконосцы и инфузории – основные потребители бактериопланктона и, одновременно, жертвы коловраток, кладоцер и копепод. Гетеротрофные флагеллаты – объекты охоты не только для метазойного планктона, но и цилиат [18, 22, 23, 25, 27, 29]. Такая многоканальность потока энергии ведет к снижению эффективности еe переноса от первичных продуцентов к многоклеточному зоопланктону и, в конечном счете, к планктоноядным рыбам.
Цель работы – создать масс-балансовую модель биотического потока энергии в экосистеме озерного типа, соединив в ней “классическую” пищевую цепь с микробиальной “петлeй”.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Для исследования скорости биотического потока энергии в пищевой сети экосистем озерного типа и влияния на нее факторов среды автором ранее была разработана масс-балансовая модель. В новую модель в формате Excel (рис. 1) были встроены ключевые группы гидробионтов – фитопланктон, бактериопланктон, гетеротрофные флагеллаты, инфузории, нехищный и хищный метазоопланктон, планктоноядные рыбы. Для верификации и калибровки модели было выбрано Рыбинское водохранилище – водоем, наиболее детально исследованный в отношении продукционных характеристик одноклеточных и многоклеточных организмов с разным типом питания [6–8, 17].
Рис. 1.
Модель биотического потока энергии при участии микробиальной пищевой сети в экосистемах озерного типа (на примере Рыбинского водохранилища). Входные параметры: Lat, Dmean, TP, Pt, DAN. Объяснение см. в тексте.
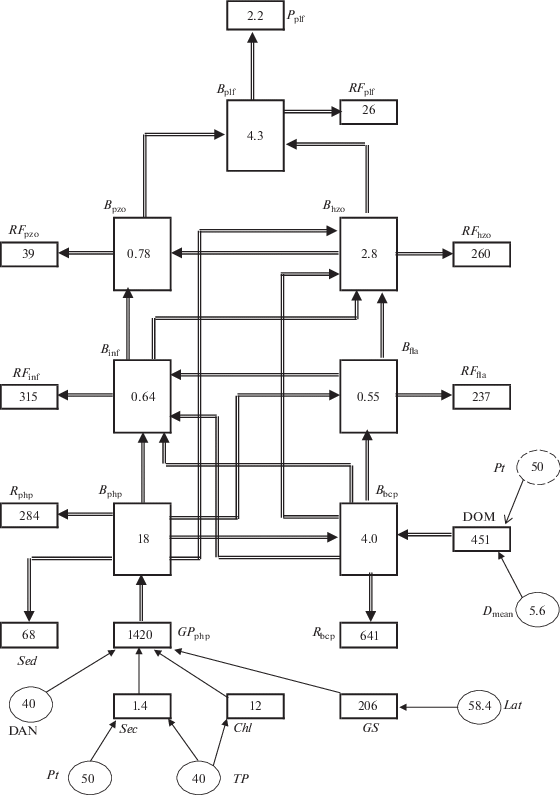
В качестве входных параметров модели использованы следующие независимые переменные:
1) биотические факторы – скорость оборота биомассы гидробионтов (V), суточное ассимиляционное число Хл. a (DAN) и эффективность роста (E) (табл. 1), коэффициенты распределения продукции гидробионтов между потребителями (DC) (табл. 2);
Таблица 1.
| Трофическое звено | V, сут-1 | E | ||
|---|---|---|---|---|
| Фитопланктон | Vphp | 0.30 | Ephp | 0.8 |
| Бактериопланктон, ассимилирующий | ||||
| автохтонное РОВ | Vbcp | 0.40 | Ebcp[aut] | 0.44 |
| аллохтонное РОВ | Vbcp | 0.40 | Ebcp[all] | 0.22 |
| Флагеллаты | Vfla | 0.7 | Efla | 0.25 |
| Инфузории | Vinf | 0.8 | Einf | 0.25 |
| Нехищный зоопланктон | Vhzo | 0.08 | Ehzo | 0.15 |
| Хищный зоопланктон | Vpzo | 0.06 | Epzo | 0.20 |
| Планктоноядные рыбы | Vplf | 0.0025 | Eplf | 0.08 |
Таблица 2.
| Трофические цепи | DC | |
|---|---|---|
| Фитопланктон: | ||
| нехищный зоопланктон | DCphp_hzo | 0.3 |
| инфузории | DCphp_inf | 0.6 |
| флагеллаты | 1 – DCphp_hzo – DCphp_inf | 0.1 |
| Бактериопланктон: | ||
| флагеллаты | DCbcp_fla | 0.8 |
| инфузории | DCbcp_inf | 0.15 |
| нехищный зоопланктон | 1 – DCbcp_fla – DCbcp_inf | 0.05 |
| Флагеллаты: | ||
| инфузории | DCfla_inf | 0.5 |
| нехищный зоопланктон | 1 – DCfla_inf | 0.5 |
| Инфузории: | ||
| нехищный зоопланктон | DCinf_hzo | 0.8 |
| хищный зоопланктон | 1 – DCinf_hzo | 0.2 |
| Нехищный зоопланктон: | ||
| хищный зоопланктон | DChzo_pzo | 0.6 |
| планктоноядные рыбы | 1 – DChzo_pzo | 0.4 |
2) абиотические факторы – географическая широта (Lat, град с.ш.); цветность воды (Pt, град); содержание в воде общего фосфора (TP, мкг/л); средняя глубины водоема (Dmean, м).
Уравнения, используемые в модельном алгоритме:
длительность вегетационного сезона [15]
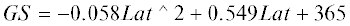
концентрация Хл. а, мкг/л [1]
прозрачность воды по диску Секки в среднем за вегетационный сезон, м [20]
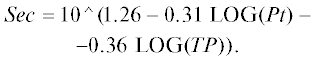
Модельный алгоритм
Фитопланктон:
1) биомасса фитопланктона, ккал/м2
2) валовая продукция фитопланктона, ккал/(м2 · год)
3) биомасса фитопланктона, осевшая на дно, ккал/(м2 · год)
линейная скорость седиментации фитопланктона принята за 0.1 м/сут [4];
4) потери энергии фитопланктоном при дыхании, ккал/(м2 · год)
Бактериопланктон:
1) биомасса бактериопланктона, ккал/м2
2) продукция фитопланктона, ассимилированная бактериопланктоном, ккал/(м2 · год)
принято, что 20% чистой продукции фитопланктона включается в бактериальную продукцию;
3) аллохтонное РОВ, ассимилированное бактериопланктоном, ккал/(м2 · год)
4) потери энергии бактериопланктоном при дыхании, ккал/(м2 · год)
Флагеллаты:
1) биомасса флагеллат, ккал/м2
2) продукция фитопланктона, потребленная флагеллатами, ккал/(м2 · год)
3) продукция бактериопланктона, потребленная флагеллатами, ккал/(м2 · год)
4) потери энергии флагеллатами при дыхании и с неусвоенной пищей, ккал/(м2 · год)
Инфузории:
1) биомасса инфузорий, ккал/м2
2) продукция фитопланктона, потребленная инфузориями, ккал/(м2 · год)
3) продукция бактериопланктона, потребленная инфузориями, ккал/(м2 · год)
4) продукция флагеллат, потребленная инфузориями, ккал/(м2 · год)
5) потери энергии инфузориями при дыхании и с неусвоенной пищей, ккал/(м2 · год)
Нехищный метазойный планктон:
1) биомасса нехищного метазойного планктона, ккал/м2
2) продукция фитопланктона, потребленная нехищным метазойным планктоном, ккал/(м2 · год)
3) продукция бактериопланктона, потребленная нехищным метазойным планктоном, ккал/(м2 · год)
4) продукция флагеллат, потребленная нехищным метазойным планктоном, ккал/(м2 · год)
5) продукция инфузорий, потребленная нехищным метазойным планктоном, ккал/(м2 · год)
6) потери энергии нехищным метазойным планктоном при дыхании и с неусвоенной пищей, ккал/(м2 · год)
Хищный метазойный планктон:
1) биомасса хищного метазойного планктона, ккал/м2
2) продукция нехищного метазойного планктона, потребленная хищным метазойным планктоном, ккал/(м2 · год)
3) продукция инфузорий, потребленная нехищным метазойным планктоном, ккал/(м2 · год)
4) потери энергии хищным метазойным планктоном при дыхании и с неусвоенной пищей, ккал/(м2 · год)
Планктоноядные рыбы:
1) биомасса планктоноядных рыб, ккал/м2
2) продукция нехищного метазойного планктона, потребленная планктоноядными рыбами, ккал/(м2 · год)
3) продукция хищного метазойного планктона, потребленная планктоноядными рыбами, ккал/(м2 · год)
4) потери энергии планктоноядными рыбами при дыхании и с неусвоенной пищей, ккал/(м2 · год)
Биотические параметры V, DAN, E и DC, заданные как x-переменные, выведены на связанные с моделью панели, аналогичные табл. 1 и табл. 2. Результаты анализа модели даются в табличной форме и включают в себя значения энергетических потоков, текущих по трофическим цепям по типу “жертва–хищник” (табл. 3), и биотический баланс водоема, т.е. биомассу и продукцию ключевых групп гидробионтов (табл. 4).
Таблица 3.
Распределение биотического потока энергии между компонентами трофической сети в экосистеме Рыбинского водохранилища (результаты моделирования)
| Трофические цепи | Поток, ккал/(м2 ∙ год) | Трофические цепи | Поток, ккал/(м2 ∙ год) |
|---|---|---|---|
| Фитопланктон: | Инфузории: | ||
| бактериопланктон | 516 | нехищный зоопланктон | 80–93 |
| нехищный зоопланктон | 166 | хищный зоопланктон | 20–24 |
| инфузории | 331 | Флагеллаты: | |
| флагеллаты | 55 | нехищный зоопланктон | 32–61 |
| седименты | 68 | инфузории | 32–61 |
| Бактериопланктон: | Нехищный зоопланктон: | ||
| инфузории | 38–82 | хищный зоопланктон | 26–31 |
| флагеллаты | 203–436 | планктоноядные рыбы | 17–21 |
| нехищный зоопланктон | 13–27 | Хищный зоопланктон: | |
| Аллохтонное РОВ: бактериопланктон | 120–1440 | планктоноядные рыбы | 9.2–11 |
Таблица 4.
Результаты моделирования при наличии и отсутствии протозойного планктона в Рыбинском водохранилище
| Трофическое звено | B, ккал/м2 | P, ккал/(м2 ∙ год) | V, год–1 | P/Pphp, % |
|---|---|---|---|---|
| Наличие протозойного планктона | ||||
| Фитопланктон | 18 | 1136 | 62 | 100 |
| Бактериопланктон | 3.1–6.6 | 254–544 | 82 | 22–48 |
| Флагеллаты | 0.45–0.85 | 65–123 | 144 | 5.7–11 |
| Инфузории | 0.61–0.72 | 100–119 | 165 | 8.8–10 |
| Нехищный зоопланктон | 2.6–3.2 | 44–52 | 16 | 3.8–4.6 |
| Хищный зоопланктон | 0.75–0.89 | 9.2–11 | 12 | 0.81–1.0 |
| Планктоноядные рыбы | 4.1–5.0 | 2.1–2.6 | 0.52 | 0.19–0.23 |
| Отсутствие протозойного планктона | ||||
| Фитопланктон | 18 | 1136 | 62 | 100 |
| Бактериопланктон | 3.1–6.6 | 254–544 | 82 | 22–48 |
| Нехищный зоопланктон | 7.3–10 | 121–164 | 16 | 11–14 |
| Хищный зоопланктон | 1.2–1.6 | 14–20 | 12 | 1.3–1.7 |
| Планктоноядные рыбы | 10–13 | 5.0–6.8 | 0.52 | 0.44–0.60 |
Примечание. B – биомасса; P – продукция; V – скорость оборота биомассы (P/B); P/Pphp – отношение продукции гидробионтов к продукции фитопланктона. Абиотические параметры, как в табл. 3.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Для Рыбинского водохранилища Lat = 58.4° с.ш., TP ~ 40 мкг/л, Pt ~50°, DAN ~ 40 мг С/мг хлорофилла a за сутки [5, 9]. На основании этих входных факторов модель прогнозирует GS = 206 сут, Chl = 12 мкг/л и Sec = 1.4 м, а с учетом последних – чистую продукцию фитопланктона Pphp = = 1420 ккал/(м2 ∙ год). Значения спрогнозированных параметров практически совпадают с данными натурных наблюдений [8, 9].
Полевые исследования на Рыбинском водохранилище [8] показали, что суммарная биомасса планктона (~30 ккал/м2 в среднем за вегетационный сезон) складывается из фитопланктона (68%), бактериопланктона (16%), простейших (5%) и многоклеточного зоопланктона (11%). Близкие к этим вклады компонентов планктона в суммарную биомассу получены для Рыбинского водохранилища с помощью модели: 59–70, 12–22, 5–6 и 12–14% соответственно.
Анализ модели показал, что средняя за вегетационный период биомасса простейших (1.1–1.6 ккал/м2) в 2.7–3 раза ниже таковой метазоопланктона (3.3–4.1 ккал/м2). Напротив, суммарная продукция протозоа (138–181 ккал/(м2 ∙ год)) выше суммарной продукции метазойного планктона (27–32 ккал/(м2 ∙ год)) в 4.9–5.7 раза. Следует подчеркнуть, что продукция протозойного и метазойного планктона рассчитывалась как сумма продукций хищников и их жертв за вычетом потребленной хищниками продукции жертв.
Модель прогнозирует, что продукция фитопланктона распределяется между потребителями следующим образом: ~45% используется бактериопланктоном в форме экстрацеллюлярной продукции, ~15% – нехищным метазойным планктоном, ~29% – инфузориями, ~5% – гетеротрофными флагеллатами, оставшаяся часть продукции фитопланктона (~6%) оседает на дно (табл. 3). Подавляющую часть продукции бактерий (~80%) потребляют гетеротрофные флагеллаты, ~15% – инфузории и ~5% – нехищный метазойный планктон.
Нехищный метазойный планктон выедает ~80% продукции инфузорий, оставшуюся часть – хищный метазойный планктон. Продукция гетеротрофных флагеллат распределяется почти поровну между нехищным метазойным планктоном и инфузориями. Хищный метазойный планктон потребляет ~60% продукции нехищного метазойного планктона, оставшуюся часть – планктоноядные рыбы. Вся продукция хищного метазойного планктона входит в рацион планктоноядных рыб.
Из модели следует также, что половину рациона хищного метазойного планктона представляет собой продукция нехищного метазойного планктона, другую половину – продукция цилиат. Рацион нехищного метазоа состоит на 48–57% из фитопланктона, 11–17% рациона составляют гетеротрофные флагеллаты, 27–28% – цилиаты, и только 4–8% – бактериопланктон. Бактериопланктон удовлетворяет 80–90% рациона гетеротрофных флагеллат, фитопланктон – 10–20%. Рацион инфузорий складывается на 70–82% из фитопланктона, на 10–17% из бактериопланктона и на 8–13% из флагеллат. Ассимиляционная активность бактериопланктона обеспечивается на 25–80% продукцией фитопланктона (еe экстрацеллюлярной фракцией) и на 20–75% аллохтонным РОВ, содержание которого по данным модели варьирует от 23 до 70 г С/м2, что соответствует данным Б.А. Скопинцева и А.Г. Бакулиной [11].
Согласно прогнозу, дыхание всего планктона в Рыбинском водохранилище изменяется от 1070 до 2250 ккал/(м2 ∙ год). Дыхание бактериопланктона в сумме с дыханием простейших достигает 60–80% дыхания планктона в целом. Отношение продукции фитопланктон к суммарному дыханию планктона варьирует в границах 0.5–1.0
Модель микробиальной пищевой сети можно без труда преобразовать в “традиционную” линейную трофическую цепь. Для этого надо лишь переформатировать значения DC (табл. 2) таким образом, чтобы перекрыть поток энергии от фитопланктона и бактериопланктона к протозойному планктону и перенаправить его к метазойному планктону. В результате такой операции получаем (табл. 4), что биомасса и продукция нехищного метазойного планктона увеличивается в среднем в 2.9 раза, планктоноядных рыб – 2.5 раза, хищного метазойного планктона – в 1.6 раза.
По данным гидроакустических съемок [3, 10], ихтиомасса в Рыбинском водохранилище колеблется на протяжении многих лет от 4.5 до 14 г/м2. Принимая, что 1 г сырой массы эквивалентен 1 ккал, находим неплохое соответствие с показаниями модели: ~4.3 ккал/м2 – при активном функционировании микробиальной “петли” и ~11 ккал/м2 – при еe редукции (табл. 4). Умножив зарегистрированные при натурных наблюдениях значения ихтиомассы на скорость ее оборота (Vplf ~0.5 год–1), получаем, что продукция планктоноядных рыб варьирует от 2.3 до 7 ккал/(м2 ∙ год), что хорошо согласуется с модельными данными – 2.2–5.5 ккал/(м2 ∙ год) (табл. 4).
В “классической” линейной цепи продукция планктоноядных рыб составляет ~0.48% продукции фитопланктона. При подключении к ней микробиальной “петли” отношение продукции рыб к продукции фитопланктона снижается до 0.2%.
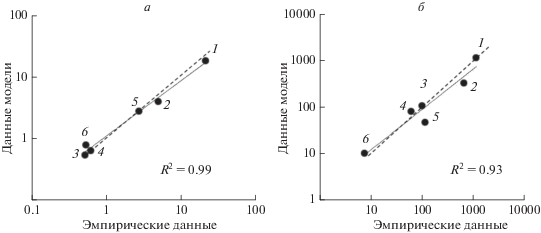
Как видно из рис. 2, значения биомассы и продукции ключевых групп планктона для Рыбинского водохранилища, спрогнозированные моделью, хорошо согласуются с показаниями натурных наблюдений [8]. Следует подчеркнуть, что сходство эмпирических и прогнозируемых значений B и P достигается лишь вследствие уменьшения в модельном алгоритме обычно принятых коэффициентов эффективности роста для флагеллат, цилиат (в 1.3 раза) и хищного метазойного планктона (в 1.6 раза). Таким образом, выполненный в данной работе анализ микробиальной трофической сети в водоемах озерного типа (на примере Рыбинского водохранилища) свидетельствует о целесообразности взаимопроверки надежности эмпирических и модельных данных.
Рис. 2.
Сравнение эмпирических и модельных данных по биомассе (а) и продукции (б) фитопланктона (1), бактериопланктона (2), гетеротрофных флагеллат (3), инфузорий (4), нехищного (5) и хищного (6) метазойного планктона в Рыбинском водохранилище. Сплошная линия – линия регрессии, пунктирная – линия симметрии (x = y).
Выводы. Проведен первый опыт моделирования микробиальной трофической сети, в котором использованы данные по структуре и функционированию планктонного сообщества в Рыбинском водохранилище и ряде озер Монголии [6, 8]. Модель построена на балансовом принципе, т.е. при строгом соблюдении закона сохранения энергии. Показано, что микробиальная пищевая сеть и линейная пастбищная цепь внутри экосистемы контактируют в “точке” выедания бактерий и простейших метазойным планктоном. В формировании структуры трофической сети, биомассы и продукции компонентов планктона доминирующую роль играют пищевые ресурсы, т.е. контроль “снизу”. К основным ресурсам, необходимым для развития фитопланктона, относятся биогенные элементы, для роста бактериопланктона – автохтонное и аллохтонноe РОВ. Представленная модель учитывает эти абиотические факторы. По мере накопления знаний о функционировании экосистем и уточнения деталей модель может стать инструментом для реконструкции элементов пищевой сети и экспертной оценки трофического состояния экосистем.
ФИНАНСИРОВАНИЕ РАБОТЫ
Работа выполнена при поддержке гос. задания № АААА-А17-117021310121-0 “Исследования динамики трофических связей, биологического разнообразия и закономерностей круговорота биогенных элементов в экосистемах континентальных водоемов, вызванной антропогенным воздействием и колебаниями климата”.
Список литературы
Бульон В.В. Новый взгляд на парадигму фосфорного контроля в лимнологии // Успехи соврем. биол. 2016. Т. 136. № 3. С. 311–318.
Бульон В.В. Автохтонное и аллохтонное органическое вещество в трофической цепи озерных экосистем // Тр. Зоол. ин-та РАН. 2017. Т. 321. № 2. С. 115–128.
Герасимов Ю.В. Популяционная динамика рыб Рыбинского водохранилища за период его существования: роль естественных и антропогенных факторов // Тр. Всерос. НИИ озер и реч. рыб. хоз-ва. 2015. Т. 156. С. 67–90.
Даценко Ю.С., Пуклаков В.В., Эдельштейн К.К. Анализ влияния абиотических факторов на развитие фитопланктона в малопроточном стратифицированном водохранилище // Тр. Карельск. науч. центра РАН. 2017. № 10. С. 73–85.
Дебольский В.К., Григорьева И.Л., Комиссаров А.Б. и др. Современная гидрохимическая характеристика реки Волга и ее водохранилищ // Вода: химия и экология. 2010. № 11. С. 2–12.
Копылов А.И., Косолапов Д.Б., Романенко А.В. и др. Микробная “петля” в планктонных сообществах озер разного трофического статуса // Журн. общ. биологии. 2007. Т. 68. № 5. С. 350–360.
Копылов А.И., Лазарева В.И., Пырина И.Л. и др. Микробная петля в планктонной трофической сети крупного равнинного водохранилища // Успехи соврем. биол. 2010. Т. 130. № 6. С. 544–556.
Копылов A.И., Косолапов Д.Б. Микробная “петля” в планктонных сообществах морских и пресноводных экосистем. Ижевск: КнигоГрад, 2011. 332 с.
Минеева Н.М. Первичная продукция в водохранилищах Волги. Ярославль: Принтхаус, 2009. 279 с.
Проект ОДУ в Рыбинском водохранилище на 2014 год // www.yarregion.ru.
Скопинцев Б.А., Бакулина А.Г. Органическое вещество в водах Рыбинского водохранилища // Тр. Ин-та биологии внутр. вод АН СССР. 1966. Вып. 13(16). С. 3–32.
Baines S.B., Pace M.L. The production of dissolved organic matter by phytoplankton and its importance to bacteria: Patterns across marine and freshwater systems // Limnol., Oceanogr. 1991. V. 36. № 6. P. 1078–1090.
Boulion V.V. Structure and function of a microbial “loop” in lake ecosystem plankton // Biologiya Vnutrennikh Vod. 2002. № 2. P. 5–14.
Bratbak G. Carbon flow in an experimental microbial ecosystem // Mar. Ecol. Prog. Ser. 1987. V. 36. P. 267–276.
Häkanson L., Boulion V.V. The lake foodweb – modelling predation and abiotic/biotic interactions. Leiden: Backhuys Publ., 2002. 344 p.
Jahnke R.A., Craven D.B. Quantifying the role of heterotrophic bacteria in the carbon cycle // Limnol., Oceanogr. 1995. V. 40. № 2. P. 436–441.
Kopylov A.I., Maslennikova T.S., Zabotkina E.A. et al. Structural and functional characteristics of planktonic microbial communities of the Rybinsk Reservoir in the impact area of a large industrial center// Inland Water Biol. 2012. V. 5. № 1. P. 36–45.
Nagata T., Takai K., Kawabata K. et al. The trophic transfer via a picoplankon – flagellate – copepods food chain during a picocyanobacterial bloom in Lake Biwa // Arch. Hydrobiol. 1996. V. 137. № 2. P. 145–160.
Nardini E., Kisand V., Letteri T. Microbial Biodiversity and molecular approach. Ispra: JRS, 2010. 49 p.
Nürnberg G.K., Shaw H. Productivity of clear and humic lakes: nutrients, phytoplankton, bacteria // Hydrobiologia. 1998. V. 382. № 1–3. P. 97–112.
Pace M.L. Heterotrophic microbial processes // The trophic cascade in lakes. Cambridge: Cambridge Univ. Press, 1993. P. 252–277.
Pace M.L., McManus G.B., Findlay S.E.G. Plankton community structure the fate of bacterial production in a temperate lake // Limnol., Oceanogr. 1990. V. 35. № 4. P. 795–808.
Pace M.L., Vaque D. The importance of Daphnia in determining mortality rates of protozoans and rotifers in lakes // Limnol., Oceanogr. 1994. V. 39. № 5. P. 985–996.
Pomeroy L.R., Williams P.J., Azam F., Hobbie J.E. The microbial loop // Oceanography. 2007. V. 20. № 2. P. 28–33.
Porter K.G., Paerl H.W., Hodson R.E. et al. Microbial interactions in lake food webs // Complex interactions in lake ecosystems. N.Y.: Springer Verlag, 1988. P. 209–227.
Stockner J.G., Porter K.C. Microbial food web in freshwater planktonic ecosystems // Complex interactions in lake ecosystems. N.Y.: Springer Verlag, 1988. P. 69–83.
Stockner J.G., Shortreed K.S. Algal picoplankton production and contribution to food webs in oligotrophic British Columbia lakes // Hydrobiologia. 1989. V. 173. № 2. P. 151–166.
Stone L., Weisburd R.S.J. Positive feedback in aquatic ecosystems // Trends Ecol. Evol. 1992. V. 7. № 8. P. 263–267.
Vezina A.F., Pace M.L. An inverse model analysis of planktonic food webs in experimental lakes // Can. J. Fish Aquat. Sci. 1994. V. 51. № 9. P. 2034–2044.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод