

Биология внутренних вод, 2019, № 4-1, стр. 68-74
Пространственная неоднородность зообентоса и зооперифитона в озерах-старицах (Западная Сибирь)
Т. А. Шарапова 1, *, Е. С. Бабушкин 2, 3
1 Институт проблем освоения Севера ФИЦ ТюмНЦ Сибирское отделение Российской академии наук
625026 Тюмень, ул. Малыгина, 86, Россия
2 Омский государственный педагогический университет
644043 Омск, наб. им. Тухачевского, 14, Россия
3 Сургутский государственный университет
628403 Сургут, пр. Ленина, 1, Россия
* E-mail: tshartum@mail.ru
Поступила в редакцию 14.05.2018
После доработки 16.10.2018
Принята к публикации 27.11.2018
Аннотация
Приведены качественные и количественные характеристики зообентоса и зооперифитона двух модельных озер-стариц бассейна Средней Оби. В результате обследования трех участков в каждой из стариц (кутового, наименее связанного с рекой, срединного и у протоки) выявлена специфическая продольная структура распределения таксономического богатства и обилия контурных беспозвоночных. Показано, что наиболее благоприятные условия обитания зообентоса складываются в кутовой части, для зооперифитона – в середине и у впадения протоки. Для понимания общих закономерностей функционирования водных экосистем, полученные результаты обсуждены в свете известных теоретических концепций ‒ экотона, краевого эффекта и озерного континуума.
ВВЕДЕНИЕ
Озера-старицы – типичный элемент поймы крупных и средних рек Западной Сибири [23, 25]. Своим генезисом они обязаны рекам и долго сохраняют связь с ними [14, 15, 17, 18, 25]. Она обычно осуществляется через одну протоку, вторая быстро заиливается и исчезает. Поступление воды из реки через протоку происходит в половодье, на противоположном конце озера ‒ участок с интенсивным зарастанием макрофитами, сопровождающимся процессами разложения растений и илонакоплением, здесь формируются условия водоема прудового типа. Показано, что поступление воды в период паводка из реки или протоки в старицу вызывает изменение химического состава стариц [7]. Многие черты биоты стариц зависят от воздействия паводковых вод [22]. Исследование брюхоногих моллюсков на трех участках в продольном направлении показало значительные различия в богатстве и составе моллюсков [3]. Старицы, находясь в непосредственной связи с рекой и испытывая воздействие половодий и паводков, тем не менее бóльшую часть времени существуют и функционируют как лентические экосистемы. В ходе сукцессии водных экосистем [14, 15] они в результате дальнейшего обособления превращаются в пойменные водоемы, со временем все меньше связанные с рекой. Следовательно, можно попытаться рассмотреть старицы в качестве переходной зоны (экотона) [21] между различными сообществами ‒ лотическими и лентическими, ‒ которая, однако, может иметь значительную протяженность. Нередко в таких зонах на границе сообществ наблюдается так называемый краевой эффект ‒ тенденция к увеличению разнообразия и плотности организмов [21], как, например, показано для устьевых областей равнинных водохранилищ [11] и водоемов речных долин [12]. Речное происхождение стариц и их характерная вытянутая форма позволяют предположить для них метамерное распределение беспозвоночных, присущее речным системам [8]. Однако в литературе имеются указания на формирование в старицах радиальной структуры в биотопическом и биономическом смысле [31]. При этом формируется нарушенная радиальная структура, поскольку оси-радиусы в старицах неравноценны в отличие от округлых пойменных и материковых озер [22].
Таким образом, изучение пространственной неоднородности распределения гидробионтов в старицах представляет интерес в свете проверки концепций экотона, краевого эффекта и озерного континуума, что важно для понимания общих закономерностей функционирования природных экосистем. Неравномерность развития биоты – характерная черта биосферы, связанная с влиянием различных абиотических и биотических факторов [24]. Неравномерность распределения гидробионтов Западной Сибири ранее изучали в основном на примере малых, средних и крупных рек [1, 2, 9, 16, 19, 27, 28, 30, 32–34].
Цель работы – выявить изменения таксономического состава, качественного и количественного развития, а также состава доминирующего комплекса контурных беспозвоночных в продольном измерении озер-стариц на примере двух модельных водоемов бассейна среднего течения р. Обь (Западная Сибирь).
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Гидробиологическая изученность водоемов Юганского заповедника, расположенного в бассейне Средней Оби, недостаточна, в основном она касается зообентоса и малакофауны рек и озер [4, 5, 10, 29, 34]. Данные по зообентосу и зооперифитону озер-стариц бассейна р. Большой Юган в литературе отсутствуют.
Исследование зообентоса и зооперифитона проводили на двух старицах в пойме рек Большой Юган (старица Школьный Урий) и Негусъях (безымянная старица) (рис. 1). В каждом озере одновременно отбирали по продольному профилю по три пробы на трех станциях: ст. 1 – кутовый участок, схожий с прудовым, мелководный (0.5 м в старице р. Негусъях и до 1.7 м в старице Школьный Урий), зарастающий макрофитами; ст. 2 – середина старицы (глубины 1.0‒1.7 м); ст. 3 – у протоки (глубины соответственно 1.4‒1.8 и 1.3‒2.0), соединяющей старицу с рекой. Длина протоки, связывающей р. Большой Юган с ее старицей, 140 м, длина протоки, соединяющей р. Негусъях с ее старицей, 65 м. Расстояние от протоки до ст. 3 в старице р. Большой Юган было 200 м, в старице р. Негусъях ‒ 15 м, от ст. 3 до ст. 2 – соответственно 970 и 350 м, от ст. 2 до ст. 1 – 1500 и 350 м.
Рис. 1.
Схема расположения водоемов и станций отбора проб (1–3). Выноски (I, II) – укрупненная схема месторасположения озер-стариц.
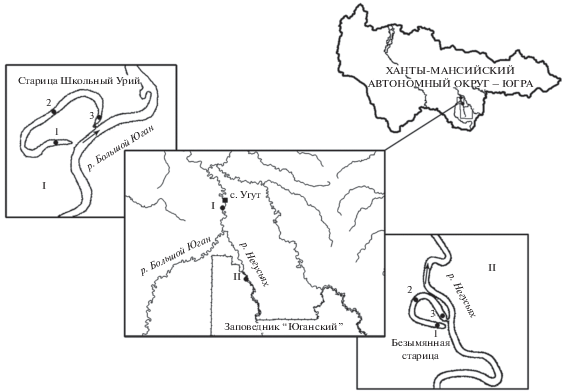
Площадь старицы р. Большой Юган достигала 0.37 км2, максимальная глубина ‒ 2.1 м, в период проведения исследований (август 2012 г.) прозрачность по диску Секки была 0.7 м, температура воды +20°С. Площадь старицы р. Негусъях ‒ 0.05 км2, максимальная глубина ‒ 1.8 м, в период проведения исследований (июль 2012 г.) прозрачность по диску Секки была 0.78 м, температура воды +28.5°С. Грунты в обеих старицах представлены серыми илами с детритом.
Пробы зообентоса отбирали по общепринятым методам [20, 26] дночерпателем Петерсена (площадь захвата 0.025 м2), по две выемки. Пробы зооперифитона собирали с затопленной ивы, учитывая площадь субстратов [29], с глубин 0.3‒0.5 м от поверхности воды. Все пробы фиксировали 4%-ным формальдегидом. Только до рода определяли олигохет, нематод, турбеллярий, остракод, водных клещей, тардиград и личинок мокрецов, a остальные группы – до вида. При статистической обработке данных вычисляли коэффициент сходства Серенсена.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Зообентос. В составе беспозвоночных бентоса озер найдено 26 так называемых НОТ (низших определяемых таксонов) [6]), включая олигохет сем. Tubificidae, нематод, пиявок, молодых особей моллюсков сем. Lymnaeidae, ракушковых рачков (Ostracoda), личинок вислокрылок (Megaloptera) Sialis sordida Klingstedt, личинок поденок (Ephemeroptera) сем. Baetidae, личинок двукрылых Diptera. Двукрылые представлены семействами мокрецов Ceratopogonidae, Chaoboridae (Chaoborus flavicans Meigen) и Chironomidae (17 видов). В старице р. Большой Юган найдено 17 таксонов, в старице р. Негусъях – 19. Коэффициент сходства Серенсена зообентоса этих стариц – 0.61. Распределение таксонов по станциям имело сходный характер, минимальное количество таксонов выявлено на ст. 2 (в старице р. Большой Юган 41% общего количества, в старице р. Негусъях 26%), максимальное – на ст. 1 (соответственно 82 и 68%) (табл. 1). Количественное развитие зообентоса этих двух стариц значительно различалось. В старице р. Большой Юган средняя численность зообентоса на разных станциях колебалась в пределах 3253–12 533 экз./м2, биомасса – 6.94–11.20 г/м2. В старице р. Негусъях отмечены более низкие значения численности (167‒5587 экз./м2) и биомассы (0.19‒9.94 г/м2). В зообентосе водоемов Западной Сибири наибольшее значение имеют три группы беспозвоночных: олигохеты, моллюски и личинки хирономид [13, 16, 28, 34]. В изученных нами водоемах моллюски имели низкую численность и значительной роли не играли, невысокая численность отмечена для олигохет (табл. 1). Основу численности и биомассы создавали личинки хирономид (доминировали виды родов Chironomus и Camptochironomus) и личинки сем. Chaoboridae (Chaoborus flavicans).
Таблица 1.
Показатели развития зообентоса на станциях (1–3) озер-стариц рек Большой Юган (Школьный Урий) и Негусъях (безымянная)
| Показатели | р. Б. Юган | р. Негусъях | ||||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 1 | 2 | 3 | |
| Число НОТ | 14 | 7 | 13 | 13 | 5 | 6 |
| Средняя численность, экз./м2 | 12 533 | 3253 | 10 947 | 5587 | 167 | 427 |
| в т.ч. Oligochaeta | 1107 | 40 | 1547 | 633 | 7 | 0 |
| Chaoborus | 233 | 2540 | 133 | 0 | 133 | 387 |
| Chironomidae | 10 713 | 647 | 9100 | 4860 | 13 | 13 |
| Chironomus + Camptochironomus | 7927 | 507 | 7133 | 1960 | 0 | 7 |
| Ceratopogonidae | 420 | 27 | 147 | 13 | 0 | 0 |
| Средняя биомасса зообентоса, г/м2 | 10.05 | 6.94 | 11.20 | 9.94 | 0.19 | 0.62 |
| в т.ч. Oligochaeta | 2.27 | 0.03 | 0.76 | 0.24 | 0.01 | 0 |
| Chironomidae | 6.35 | 2.44 | 9.71 | 9.35 | 0.02 | 0.02 |
| Chironomus +Camptochironomus | 4.39 | 1.57 | 8.07 | 4.72 | 0 | 0.01 |
| Chaoborus | 0.48 | 4.35 | 0.28 | 0 | 0.16 | 0.58 |
В зообентосе старицы р. Большой Юган высокие показатели численности и биомассы отмечены на ст. 1 и 3. На этих участках зарегистрированы близкие значения численности и биомассы основных групп (табл. 1). Доминировали личинки хирономид (85‒87% численности), из них наибольший вклад в количественные показатели был у детритофагов родов Chironomus и Camptochironomus (63‒65% численности и 63‒87% биомассы). Численность олигохет (сем. Tubificidae) имела близкие значения, но биомасса на ст. 1 в 3 раза превышала таковую ст. 3. У крупных личинок хирономид доминирующего комплекса Chironomus + Camptochironomus также отмечены сходные значения численности, но биомасса на ст. 3 была выше в 1.8 раза. Зообентос на ст. 2 значительно отличался от такового ст. 1 и ст. 3, здесь зарегистрированы минимальные значения численности и биомассы всех групп (табл. 1), кроме личинок хаоборид, плотность и биомасса которых была максимальна, и они доминировали как по численности (78%), так и по биомассе (62%). Для зообентоса старицы р. Негусъях максимальные значения численности и биомассы отмечены также на ст. 1, личинки хирономид доминировали по численности (87%) и биомассе (94%), среди них наибольшую роль играл комплекс Chironomus + Camptochironomus. Низкие значения развития отмечены на ст. 2 и ст. 3 (табл. 1), где преобладали личинки Chaoborus flavicans (79‒90% численности и 84‒94% биомассы).
Зооперифитон. В составе зооперифитона озер найдено 42 таксона, включая гидр, олигохет семейств Naididae и Tubificidae, нематод, турбеллярий, пиявок (3 вида), молодых особей брюхоногих моллюсков, мшанок (2 вида), ветвистоусых рачков (4 вида), тихоходок, водных клещей, коллембол, личинок водных жуков, поденок (3 вида), стрекоз, двукрылых семейств Ceratopogonidae и Chironomidae (14 видов). Количество таксонов в зооперифитоне обеих стариц очень близко – 28 в старице р. Большой Юган и 29 в старице р. Негусъях. Коэффициент сходства Серенсена – 0.53. Распределение таксонов зооперифитона по станциям носит сходный характер. Минимальное количество таксонов отмечено на ст. 1 ‒ в старице р. Большой Юган 39% общего количества, в старице р. Негусъях – 62%, на ст. 2 количество таксонов достигало 82 и 69%, на ст. 3 – 68 и 76% соответственно.
Показатели количественного развития зооперифитона стариц резко изменялись по станциям. В старице р. Большой Юган максимальная численность отмечена на ст. 1, в 2 раза ниже она была на ст. 3, минимальная – на ст. 2 (табл. 2). На ст. 1 по численности доминировали олигохеты сем. Naididae (59%) и личинки хирономид (38%), на ст. 2 и ст. 3 – личинки хирономид (66‒74%). Основу биомассы на всех станциях создавали личинки хирономид (70‒91%), преимущественно рода Glyptotendipes (50‒80% биомассы хирономид). Высокие значения биомассы зарегистрированы на ст. 3 и ст. 2, низкие – на ст. 1.
Таблица 2.
Показатели развития зооперифитона на станциях (1–3) озер-стариц рек Б. Юган и Негусъях
| Показатели | р. Б. Юган | р. Негусъях | ||||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 1 | 2 | 3 | |
| Число НОТ | 11 | 23 | 19 | 18 | 20 | 22 |
| Средняя численность, экз./м2 | 203 155 | 73 880 | 107 880 | 271 467 | 63 640 | 157 055 |
| в т.ч. Hydra | 0 | 0 | 1072 | 25 276 | 2978 | 5607 |
| Chydorus sphaericus + Bosmina sp. | 0 | 0 | 0 | 173 698 | 11 643 | 20 617 |
| Oligochaeta | 119 847 | 16 924 | 27 135 | 15 888 | 3862 | 26 687 |
| Nematoda | 6145 | 1003 | 6490 | 4143 | 1056 | 18 160 |
| Ephemeroptera | 118 | 149 | 808 | 0 | 87 | 223 |
| Chironomidae | 76 925 | 54 903 | 71 531 | 49 175 | 41 030 | 78 452 |
| Glyptotendipes | 11 967 | 17 568 | 15 569 | 1531 | 6521 | 8058 |
| Средняя биомасса, г/м2 | 20.37 | 26.50 | 32.67 | 12.07 | 125.93 | 38.48 |
| в т.ч. Hydra | 0 | 0 | 0.14 | 3.40 | 0.28 | 0.56 |
| Bryozoa | 0 | 0.02 | 0 | 0.1 | 104.57 | 4.64 |
| Chironomidae | 14.30 | 24.06 | 29.67 | 5.49 | 20.24 | 26.24 |
| Glyptotendipes | 7.23 | 19.29 | 22.40 | 1.56 | 12.66 | 18.00 |
В старице р. Негусъях выявлено такое же, как и в старице р. Б. Юган, распределение численности ‒ максимальная на ст. 1, в 1.7 раза меньше на ст. 3 и минимальная ‒ на ст. 2 (табл. 2). Состав доминирующего комплекса отличался. На ст. 1 в массе развивались мелкие ветвистоусые рачки – комплекс Chydorus sphaericus + Bosmina sp. (64% общей численности), на ст. 2 и ст. 3 их плотность снижалась в 8‒15 раз. Следует отметить высокую положительную корреляцию (0.94) между численностью мелких кладоцер и гидр, для которых, очевидно, эти ветвистоусые рачки служат кормом. На ст. 2 и ст. 3 по численности преобладали личинки хирономид (50–64%), но плотность Glyptotendipes, по сравнению со старицей р. Большой Юган, была ниже на всех станциях (табл. 2). В обеих старицах по биомассе из хирономид наибольшее значение имели личинки рода Glyptotendipes (28–69%). Минимальная биомасса отмечена на ст. 1, где доминировали личинки хирономид (45%) и гидры (28%), на ст. 3 биомасса зооперифитона выше в 3 раза, преобладают личинки хирономид (68%). На ст. 2 основу биомассы (83%) создавали крупные колонии мшанки Hyalinella punctata (Hancock).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Несмотря на различия в таксономическом составе беспозвоночных озер, отраженном в невысоком индексе сходства Серенсена, а также в количественном развитии бентосных и перифитонных беспозвоночных, в обеих старицах отмечается сходство в особенностях развития и структуре на исследуемых участках как зообентоса, так и зооперифитона. Минимальные значения индекса сходства Серенсена отмечены между ст. 1 и 2 как для зообентоса (старица р. Большой Юган 0.47 и р. Негусъях 0.22), так и для зооперифитона (соответственно 0.58 и 0.68), наибольшие – между ст. 2 и ст. 3 (для зообентоса 0.70 и 0.36, для зооперифитона 0.71 и 0.76).
Для зообентоса обеих стариц характерны высокие показатели НОТ и численности на ст. 1, где в массе развивались детритофаги ‒ олигохеты сем. Tubificidae и хирономиды родов Chironomus и Camptochironomus (в старице р. Большой Юган Chironomus solitus Linevich и Camptochironomus tentans (Fabricius), в старице р. Негусъях Chironomus nigrocaudatus Erbaeva и Camptochironomus pallidivittatus (Malloch), которые преобладали и по биомассе) (табл. 1). В середине озер (ст. 2) доминировали по численности и биомассе виды рода Chaoborus. На этом участке отмечены минимальные показатели развития зообентоса, как в отношении разнообразия НОТ, так и количественных характеристик. Наибольшие различия между зообентосом озер выявлены на участке у протоки (ст. 3). Так, в старице р. Большой Юган ст. 3 по количественным показателям схожа со ст. 1, а в старице р. Негусъях ‒ со ст. 2.
Зооперифитон обеих стариц также развит неравномерно. В отличие от зообентоса, наибольшее число НОТ выявлено на ст. 2 и 3, наименьшее – на ст. 1. Наиболее высокие показатели численности зооперифитона в обоих озерах наблюдаются на ст. 1, где в старице р. Большой Юган доминируют олигохеты сем. Naididae, в старице р. Негусъях – мелкие кладоцеры. В обеих старицах отмечены минимальные значения плотности на середине озера (ст. 2), что связано со снижением на этой станции численности молодых личинок хирономид длиной <2 мм, доминирующих на всех станциях. В обеих старицах максимальная плотность личинок поденок отмечена на ст. 3 – у протоки. Наименьшие значения численности и биомассы личинок хирономид Glyptotendipes glaucus (Meigen), играющих большую роль в создании биомассы, отмечены на ст. 1 (кутовый участок), на ст. 2 и ст. 3 численность и биомасса этих личинок значительно выше. Следует отметить изменение средней массы этих личинок по станциям: в старице р. Большой Юган на ст. 1 средняя индивидуальная масса личинки была 0.60, на ст. 2 – 1.10, на ст. 3 – 1.43 мг, в старице р. Негусъях, соответственно, 1.00, 1.94 и 2.23 мг. Минимальные показатели биомассы зооперифитона на ст. 1 в обеих старицах определяются мелкими размерами и невысокой плотностью личинок хирономид Glyptotendipes. Небольшие колонии мшанок Hyalinella punctata в старице р. Большой Юган найдены только в середине озера (ст. 2) и большой роли в создании биомассы не играют, в старице р. Негусъях эта мшанка встречалась на всех станциях, но самые крупные колонии найдены на ст. 2, где создают основу биомассы (83%).
Рассматривая развитие беспозвоночных в озерах-старицах, можно отметить, что на ст. 1 зоо-бентос достигает максимального развития как в отношении таксономического богатства, так и количественного развития, доминируют личинки хирономид. Также здесь выявлены высокие значения численности олигохет сем. Tubificidae и личинок мокрецов сем. Ceratopogonidae. Для зооперифитона этого участка отмечены максимальные значения численности, в одном озере доминировали олигохеты сем. Naididae, в другом – мелкие ветвистоусые рачки, но биомасса и таксономическое богатство зооперифитона были минимальны. Следовательно, кутовые участки обеих стариц наиболее отличны от соседних, что, по-видимому, обусловлено их обособлением, ‒ они реже испытывают влияние паводков и половодий, в них складываются специфические условия, характерные для лентических экосистем. Особенности качественного и количественного развития зообентоса и зооперифитона кутовых участков стариц свидетельствует о замедленном водообмене, накоплении органики, естественном эвтрофировании – поступательных процессах, характерных для лентических экосистем, способствующих превращение водоемов в наземные экосистемы [14, 15, 21, 31].
В середине озера выявлены низкие показатели численности зообентоса и зооперифитона ‒ результат снижения плотности олигохет, нематод и хирономид. В зообентосе этого участка основу численности создают хищные личинки Chaoborus, в зооперифитоне хирономиды молодых личиночных стадий. В зооперифитоне середины озера максимальных величин достигает биомасса мшанок. На участках озер в середине (ст. 2) и у протоки (ст. 3) наибольшее таксономическое богатство и биомасса выявлены в зооперифитоне. Возле протоки в зооперифитоне зарегистрированы высокие показатели численности личинок поденок и хирономид рода Glyptotendipes.
Таким образом, в исследованных старицах существует пространственная неоднородность распределения качественных и количественных характеристик зообентоса и зооперифитона вдоль водоема от кутового участка до протоки, соединяющей старицу с рекой. Предположение о метамерном строении таких водоемов и метамерном распределении гидробионтов нашими данными не подтверждается. Очевидна также неприменимость концепций лотических экосистем (речного континуума, четырехмерной природы, динамики пятен) для описания стариц и распределения в них гидробионтов [8]. Выявленная продольная структура распределения зообентоса и зооперифитона очевидно отражает градиент перехода от лотических к лентическим экосистемам, при этом в переходной зоне (ст. 2) формируются наименее благоприятные условия. Сравнивая таксономическое богатство и обилие зообентоса и зооперифитона изученных стариц с аналогичными показателями рек Большой Юган и Негусъях, полученными нами ранее [34], можно предположить существование краевого эффекта. Действительно, средние численность и биомасса зообентоса и особенно зооперифитона в старицах в несколько раз выше. Однако для проверки такого предположения пока не хватает исследований в лентических экосистемах, следующих звеньях сукцессии водных экосистем – пойменных водоемах и материковых озерах. Следовательно, делать выводы о существовании краевого эффекта преждевременно. Возможно, выявленная нами продольная структура – часть нарушенной радиальной структуры от берегов к центру. Для проверки последнего предположения следует планировать станции отбора проб на нескольких разрезах от берега к берегу через центр, отражающих гипотетическую нарушенную радиальную структуру. Также в рамках концепции озерного континуума для проверки гипотезы формирования в старицах более сложной пространственной структуры лентических экосистем, по сравнению с лотическими, интересно изучить распределение гидробионтов различных экологических группировок (планктона, бентоса, перифитона) вдоль основных градиентов – радиального, вертикального и временного. Важно понять какие процессы в таких экотонных экосистемах преобладают во временном измерении – циклические, как в реках (половодье‒межень) и озерах (стагнация и перемешивание), или поступательные однонаправленные, как в озерах (эвтрофикация). Вероятно, в разных зонах стариц либо в старицах разного возраста преобладают различные процессы.
Выводы. Зообентос и зооперифитон на трех исследуемых участках озер-стариц отличаются качественным составом, количественными показателями развития и доминирующими комплексами. В кутовых участках стариц складываются благоприятные условия для развития зообентоса с высокими качественными и количественными показателями и доминированием детритофагов. Наибольшие показатели таксономического богатства и биомассы зооперифитона отмечены в центральных участках стариц и у проток, соединяющих их с реками, при доминировании фильтраторов. Постепенно обособляясь от основного потока, старые речные русла существуют и функционируют отлично от лотических экосистем, непосредственное влияние которых они, тем не менее, периодически испытывают. Подтверждением отличий служит формирование специфической структуры распределения гидробионтов в старицах. По-видимому, эта структура динамична и сильно зависит от колебаний условий среды. Решение затронутых в работе проблем имеет фундаментальное и прикладное значение. Существует много вопросов для дальнейших исследований устройства и функционирования как озер-стариц, так и основной сукцессионной последовательности водных экосистем в ряду: речное русло–придаточные–пойменные водоемы–непойменные или материковые озера.
Список литературы
Абдуллина Г.Х. Развитие зоопланктона р. Туры // Вестник экологии, лесоведения и ландшафтоведения. 2007. № 7. С. 131–137.
Абдуллина Г.Х., Алексюк В.А. К изучению зоопланктона водоемов и водотоков бассейна р. Таз // Человек и Север: антропология, археология, экология: Матер. Всерос. конф. Тюмень: Институт проблем освоения Севера СО РАН, 2009. С. 213–217.
Азявчикова Т.В., Клещенко Е.П. Популяционная структура брюхоногих моллюсков старицы реки Сож // Альманах современной науки и образования. 2013. № 11(78). С. 13–14.
Андреев Н.И., Андреева С.И., Бабушкин Е.С. Фауна и распределение двустворчатых моллюсков (Bivalvia) в бассейне р. Малый Юган (Среднее Приобье) // Ruthenica: Рус. малакол. журн. 2016. Т. 26. № 3–4. С. 191–201.
Бабушкин Е.С. К изучению динамики макрозообентоса рек бассейна реки Большой Юган (Среднее Приобье) // Динамика окружающей среды и глобальные изменения климата. 2015. Т. 6. № 2(12). С. 44–54.
Баканов А.И. Использование характеристик разнообразия зообентоса в мониторинге состояния пресноводных экосистем // Мониторинг биоразнообразия. М.: Ин-т проблем экологии и эволюции РАН, 1997. С. 278–282.
Баянов Н.Г., Кривдина Т.В. Межсезонная динамика гидролого-гидрохимических показателей реки Керженец и ее стариц // Изв. РАН. Сер. геогр. 2013. № 2. С. 52–67.
Беклемишев В.Н. Биоценозы реки и речной долины в составе живого покрова Земли // Тр. Всесоюз. гидробиол. об-ва. 1956. Т. 7. С. 77‒97.
Бурмакин Е.В. Кормовые ресурсы Гыданского залива и близлежащих водоемов // Тр. НИИ полярн. земледелия, животноводства и промысл. хоз-ва. Серия “Промысл. хоз.”. 1941. Вып. 15. С. 159–177.
Винарский М.В., Андреев Н.И., Бабушкин Е.С., Каримов А.В. Материалы по фауне пресноводных легочных моллюсков Юганского заповедника и сопредельных территорий // Динамика окружающей среды и глобальные изменения климата. 2015. Т. 6. № 2(12). С. 34–43.
Гидроэкология устьевых областей притоков равнинного водохранилища. Ярославль: Филигрань, 2015. 466 с.
Ермохин М.В. Проблемы и перспективы исследования краевых структур биоценозов рек и водоемов речных долин // Актуальные вопросы изучения микро-, мейозообентоса и фауны зарослей пресноводных водоемов. Нижний Новгород: Вектор ТиС, 2007. С. 101–129.
Залозный Н.А. Роль олигохет и пиявок в экосистемах водоемов Западной Сибири // Биологические ресурсы внутренних водоемов Сибири и Дальнего Востока. М.: Наука, 1984. С. 124–142.
Иоганзен Б.Г. Принцип единства биотопа биоценоза // Вопр. географии Сибири. Томск, 1966. Сб. 6. С. 142–154.
Иоганзен Б.Г., Глазырина Е.И., Залозный Н.А. и др. Сукцессия водных экосистем в бассейне Средней Оби // Сукцессии животного населения в биоценозах поймы реки Оби. Новосибирск: Наука, 1981. С. 78–99.
Иоффе Ц.И. Донная фауна Обь-Иртышского бассейна и ее рыбохозяйственное значение // Изв. Всесоюз. НИИ озер. и реч. рыб. хоз-ва. 1947. Т. 25. Вып. 1. С. 113–160.
Китаев С.П. Основы лимнологии для гидробиологов и ихтиологов. Петрозаводск: Карельск. науч. центр РАН, 2007. 395 с.
Константинов А.С. Общая гидробиология. М.: Высш. шк., 1986. 472 с.
Кузикова В.Б., Бусленко Н.М. Донная фауна реки Сыни и ее роль в питании рыб // Сб. науч. тр. Гос. НИИ озер. и реч. рыб. хоз-ва. 1989. № 305. С. 81.
Методы изучения биогеоценозов внутренних водоемов. М.: Наука, 1975. С. 158–170.
Одум Ю. Экология. Т. 2. М.: Мир, 1986. 376 с. (Odum E.P. Basic Ecology. Philadelphia: Saunders College Publ., 1983. 613 p.).
Папченков В.Г. Растительный покров водоемов и водотоков Среднего Поволжья. Ярославль: ЦМП МУБ и НТ, 2001. 200 с.
Природные условия и естественные ресурсы СССР. Западная Сибирь. М.: Изд-во АН СССР, 1963. 488 с.
Протасов А.А. Жизнь в гидросфере. Очерки по общей гидробиологии. Киев: Академпериодика, 2011. 704 с.
Ресурсы поверхностных вод СССР. Алтай и Западная Сибирь. Средняя Обь. Т. 15. Вып. 2. Л.: Гидрометеоиздат, 1972. 415 с.
Руководство по методам гидробиологического анализа поверхностных вод и донных отложений. Л.: Гидрометеоиздат, 1983. С. 39–48.
Степанов Л.Н. Разнообразие зообентоса водоемов и водотоков бассейнов рек Сетная и Нгояха (полуостров Ямал, Ямало-Ненецкий Автономный округ) // Фауна Урала и Сибири. 2016. № 1. С. 90–104.
Степанова В.Б. Макрозообентос Нижней Оби // Вестник экологии, лесоведения и ландшафтоведения. 2009. № 9. С. 155–162.
Шарапова Т.А. Зооперифитон внутренних водоемов Западной Сибири. Новосибирск: Наука, 2007. 167 с.
Palatov D.M., Chertoprud M.V. The rheophilic fauna and invertebrate communities of the tundra zone: a case study of the southern Yamal // Inland Water Biol. 2012. V. 5. № 1. P. 19–28. https://doi.org/10.1134/S1995082912010117
Protasov A.A. River and lake continua: an attempt at analysis and synthesis // Inland Water Biol. 2008. V. 1. № 2. P. 105–113. https://doi.org/10.1134/S1995082908020016
Sharapova T.A. Basic Characteristics of Zooperiphyton in Large Rivers of Western Siberia (Tyumen Oblast) // Inland Water Biol. 2012. V. 5. № 4. P. 350–356. https://doi.org/10.1134/S199508291204013X
Sharapova T.A. Spatial Structure of Zooperiphyton from the Ob River // Contemporary Problems of Ecology. 2009. V. 1. P. 72–76.
Sharapova T.A., Babushkin E.S. Comparison of Zoobenthos and Zooperiphyton of Large and Medium Rivers // Contemporary Problems of Ecology. 2013. V. 6. № 6. P. 622–626.
Townsend C. The patch dynamics concept of stream community ecology // J. N.Y. Benthol. Soc. 1989. V. 8. № 1. P. 36–50.
Vannote R.L., Minchall G.W., Cummins K.W. et al. The river continuum concept // Can. J. Fish. Aquat. Sci. 1980. V. 37. № 1. P. 130–137.
Ward J.V. The four-dimensional nature of lotic ecosystems // J. N.Y. Benthol. Soc. 1989. V. 8. № 1. P. 2–8.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод