

Биология внутренних вод, 2019, № 4-1, стр. 25-32
Таксономическая структура и экология фитопланктона малых лесных озер в зоне техногенеза сульфидного месторождения (Южный Урал)
Л. В. Снитько 1, *, В. П. Снитько 1
1 Ильменский государственный заповедник
456317 Челябинская обл., Миасс, Россия
* E-mail: lvs223@yandex.ru
Поступила в редакцию 23.04.2018
После доработки 30.10.2018
Принята к публикации 24.12.2018
Аннотация
Проведена систематизация многолетних данных (2009–2017 гг.) по таксономической структуре фитопланктона малых лесных озер Южного Урала на территории сульфидного медно-цинкового месторождения и в зоне воздействия медеплавильного комбината. Выявлена закономерность изменения видового богатства фитопланктона в градиенте увеличения ацидности, концентрации тяжелых металлов, содержания сульфат-иона – значительное снижение таксономического разнообразия фитопланктона. Видовое богатство золотистых водорослей увеличивается в олигоацидных условиях лесных озер буферной зоны техногенеза. Проанализированы связи структурных показателей фитопланктона с абиотическими факторами.
ВВЕДЕНИЕ
На Южном Урале распространены пресные гидрокарбонатные нейтрально-слабощелочные природные воды [10, 18], фитопланктон которых характеризуется высоким разнообразием [15, 16]. В зонах геохимических аномалий (горнорудных сульфидных медно-цинковых месторождениях) формируются воды другого типа: кислые сульфатные, с вовлечением в миграционные циклы металлов халькофильного ряда – Cu, Zn, Pb, Сd и др. [18]. Карабашский горно-обогатительный и медеплавильный комбинат, расположенный на месторождении с 1908 г., загрязняет озера водами, образованными окислением сульфидов в отвалах добычи и обогащения руд, и к природной аномалии добавляется техногенная. Зона воздействия расширяется за счет воздушного переноса загрязнений. При прохождении дождей через шлейф газопылевых выбросов комбината на сопредельной территории выпадают кислотные осадки с высоким содержанием металлов [18]. Техногенная миграция микроэлементов в кислых рудничных водах, поступающих в водоемы, обуславливает формирование в них специфичных гидрохимических условий, оказывающих влияние на гидробионты. Для выявления трансформации фитопланктона в условиях техногенеза необходим количественный и качественный анализ сообществ.
Цель работы – изучение изменения таксономической структуры пресноводного фитопланктона в условиях комплексного техногенного воздействия и ацидификации.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Материалом работы послужили пробы фитопланктона из семи малых водоемов (20–21 из каждого), расположенных в районе сульфидного месторождения (рис. 1). Пробы отбирали автоматическим батометром объемом 1 л в центре исследуемых водоемов: в период открытой воды (с мая по октябрь) 2009–2010, 2012–2014 гг. раз в два месяца или чаще в зависимости от температурного режима в подледный период 2015–2017 гг. (февраль, март–апрель) по две–три пробы в каждом водоеме. Фитопланктон осаждали на мембранные фильтры “Владипор” с диаметром пор 1 мкм, а также концентрировали с помощью сачка оригинальной конструкции через полиамидное сито с порами 14 мкм. Живой материал просматривали под световым микроскопом Nikon Eclipse E600 с объективами Nikon 60×/0.80, 40×/0.65 Ph2, 10×/0.25 Ph1 и для дальнейшего определения фиксировали раствором Люголя с добавлением ледяной уксусной кислоты. Учет численности фитопланктона проводили под световым микроскопом в счетной камере объемом 0.01 мл [11], идентификацию диатомовых и золотистых водорослей – с помощью СЭМ Tescan Vega 3Sbu (× (2–20) × 103). Номенклатура и размерные характеристики водорослей и цианопрокариот представлены согласно сводкам [2, 3, 8, 9, 12, 14, 20, 21, 25–27, 29–31, 33–36]. Параллельно отбирали пробы воды для гидрохимического анализа, выполненного в Южно-Уральском центре коллективного пользования по исследованию минерального сырья (аттестат аккредитации № РОСС RU.001.514536).
Рис. 1.
Схема района исследования. Территория воздействия техногенеза сульфидного месторождения: 1 – промышленные объекты Карабашского медеплавильного комбината (отвалы руды, шахты, плавильные печи), 2 – территория, подверженная техногенным поверхностным стокам, 3–7 – исследованные водоемы, 8 – верхний створ Аргазинского водохранилища при впадении рек Миасс и Сак-Элга. Стрелки указывают направление стоков комбината.
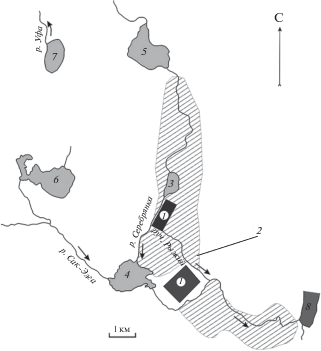
Исследованные водоемы (рис. 1, табл. 1), расположенные на небольшой территории (55°18′53″–55°31′51″ с.ш., 60°07′28″–60°13′20″ в.д.) на высоте 302–472 м над уровнем моря, относятся к Обскому и Волго-Камскому речным бассейнам, водосбор полностью или частично покрыт лесом, проточно-сточные озера и пруды имеют плотины. Согласно классификации поверхностных природно-техногенных вод Карабашской геосистемы [1], Карабашский и Богородский пруды относятся к импактной зоне, оз. Серебры – к импакт-буферной, озера Барахтан и Уфимское – к буферной, озера Сириккуль и Карматкуль, находящиеся на территории Ильменского заповедника в 15 км к югу от комбината, – к фоновой зоне. Зоны выделены в соответствии с градиентом комплексного техногенного загрязнения: увеличения ацидификации (рН воды), суммарной концентрации металлов (Cu, Zn, Pb, Сd, Fe, Mn), сульфат-иона и уменьшения гидрокарбонат-иона. Озера фоновой зоны характеризуются нейтрально-щелочным уровнем рН 7.0–9.4, содержанием металлов в поверхностном слое воды ≤0.05 мг/дм3 и значительным преобладанием гидрокарбонатов над сульфатами. Величины рН в буферной, импакт-буферной зоне и импактной зонах воздействия составляют 6.1–7.2, 5.2–7.1 и 5.0–7.2, содержание металлов – 0.3–2.0, 0.3–3.0 и 1.0–15.0 мг/дм3 соответственно (табл. 1). Водоемы импактной зоны в наибольшей степени подвержены ацидификации (закислению) и загрязнению металлами, привносимыми поверхностными водотоками из-под отвалов руды и атмосферными выбросами.
Таблица 1.
Морфометрические и гидрохимические характеристики исследованных водоемов, относящихся к различным зонам
| Показатель | Импактная | Импакт-буферная | Буферная | Фоновая | |||
|---|---|---|---|---|---|---|---|
| Кара-башский (3) | Бого-родский (4) | Серебры (5) | Барахтан (6) | Уфимское (7) | Карматкуль | Сириккуль | |
| Пруды | Озера | ||||||
| Площадь зеркала, км2 | 0.56 | 1.24 | 1.65 | 1.85 | 0.90 | 0.92 | 0.86 |
| Глубина, м (средняя/макс.) | 1.2/3.0 | 2.0/5.5 | 3.0/6.0 | 1.1/4.0 | 1.3/3.5 | 2.0/4.0 | 2.0/5.0 |
| Объем, млн м3 | 0.67 | 2.48 | 4.95 | 2.04 | 1.17 | 1.84 | 1.72 |
| Площадь водосбора, км2 | 2.66 | 3.15 | 6.20 | 6.30 | 4.90 | 4.20 | 2.86 |
| рН | 5.5–7.2 | 5.0–7.0 | 5.2–7.1 | 6.1–7.2 | 6.2–6.9 | 7.0–9.1 | 7.4–9.4 |
| Цветность, град | 41–47 | 50–90 | 9–25 | 56–59 | 36–55 | 28–62 | 32–46 |
| Электропроводность, мкСм/см | 403–520 | 102–416 | 153–164 | 119–134 | 86–315 | 235–309 | 198–336 |
| Гидрокарбонаты, мг/дм3 | 35.0–54.9 | 31.7–57.9 | 18.3–37.8 | 19.5–29.9 | 21.3–61.0 | 86.0–134 | 73.2–164 |
| Сульфаты, мг/дм3 | 136–167 | 91.8–238 | 53.0–57.0 | 38.1–38.7 | 31–184 | 18.5–25.4 | 18.1–30.5 |
| Робщ, мг/дм3 | 0.036–0.321 | 0.029–0.136 | 0.057–0.110 | 0.043–0.050 | 0.051–0.145 | 0.060–0.133 | 0.080–0.170 |
| Металлы Cu, Zn, Pb, Cd, Fe, Mn, мг/дм3 | 1.0–15.0 | 0.3–3.0 | 0.3–2.0 | <0.05 | |||
По минерализации водоемы пресные и ультрапресные, по цветности – олиго-мезогумозные. Низкая (до 0.2 м) прозрачность воды обусловлена высоким содержанием минеральной взвеси и меняется в зависимости от атмосферных осадков и перемешивания. По микроэлементному составу, определяемому загрязнением металлами, водоемы фоновой зоны относятся к умеренно загрязненным, озера буферной – к сильнозагрязненным, водоемы импактной зоны – к техногенным вне природной классификации. Водоемы импактной и буферной зоны влияния комбината подвержены ацидификации, но поверхностные слои в период открытой воды преимущественно олигоацидные, озера фоновой зоны – нейтрально-щелочные (табл. 1). По уровню трофии, определяемой содержанием общего фосфора и азота [13], водоемы эвтрофные, а озера фоновой зоны в летний период достигают уровня гипертрофии.
Вследствие попадания техногенных закисленных рассолов (рН 2.8–3.5, электропроводность до 956 мкСм/см) из поверхностного и подземного стока, гидрохимический режим водоемов неустойчив [10, 18]. Содержание металлов в зоне влияния комбината может в сотни раз превосходить концентрации, допустимые рыбохозяйственными нормативами. Стоки комбината текут в верхний створ Аргазинского водохранилища (рис. 1), оказывая негативное влияние на его водные сообщества [17].
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
B составе планктонной альгофлоры семи исследованных озер выявлено 356 таксонов водорослей рангом ниже рода, относящихся к 10 отделам (табл. 2). Основу фитопланктона (74%) составляют широко распространенные виды. По отношению к солености воды их бóльшая часть принадлежит к олигогалобам и индифферентам, а Cyclotella meneghiniana Kützing и Fragilaria crotonensis Kitton – галофилы. По отношению к рН среды большинство видов относится к индифферентам, встречаются ацидофильные виды из родов Closterium и Ulothrix и алкалифилы (Aulacoseira). Наибольшим видовым богатством характеризуется фитопланктон озер фоновой зоны: 290 таксонов рангом ниже рода из 10 отделов. В зоне влияния комбината число таксонов всех рангов существенно снижается (табл. 2).
Таблица 2.
Количество видов, разновидностей и форм фитопланктона водоемов в различных зонах воздействия техногенеза Карабашского сульфидного месторождения
| Таксон | Импактная | Импакт-буферная | Буферная | Фоновая | Всего |
|---|---|---|---|---|---|
| Cyanoprokaryota | |||||
| Croococcophyceae | 2 | 2 | 6 | 25 | 25 |
| Aphanothece | 1 | 0 | 2 | 2 | 2 |
| Rhabdoderma | 1 | 0 | 1 | 1 | 1 |
| Aphanocapsa | 0 | 1 | 2 | 5 | 5 |
| Merismopedia | 0 | 0 | 0 | 1 | 1 |
| Coelosphaerium | 0 | 0 | 0 | 1 | 1 |
| Snowella | 0 | 0 | 0 | 3 | 3 |
| Woronichinia | 0 | 0 | 0 | 3 | 3 |
| Gomphosphaeria | 0 | 0 | 0 | 2 | 2 |
| Мicrocystis | 0 | 0 | 0 | 5 | 5 |
| Chroococcus | 0 | 1 | 1 | 2 | 2 |
| Hormogoniophyceae | |||||
| Oscillatoriales | 4 | 4 | 8 | 16 | 21 |
| Romeria | 0 | 0 | 0 | 1 | 1 |
| Pseudanabaena | 0 | 0 | 0 | 1 | 1 |
| Spirulina | 0 | 0 | 1 | 1 | 1 |
| Limnothrix | 1 | 1 | 1 | 2 | 2 |
| Planktolyngbya | 1 | 1 | 1 | 1 | 1 |
| Planktothrix | 0 | 0 | 0 | 1 | 1 |
| Tychonema | 0 | 0 | 1 | 1 | 1 |
| Phormidium | 0 | 0 | 1 | 1 | 2 |
| Lyngbya | 0 | 0 | 0 | 2 | 2 |
| Oscillatoria | 2 | 2 | 3 | 6 | 9 |
| Hormogoniophyceae Nostocales | 0 | 3 | 0 | 10 | 11 |
| Anabaena | 0 | 2 | 0 | 6 | 7 |
| Aphanizomenon | 0 | 1 | 0 | 2 | 2 |
| Tolypothrix | 0 | 0 | 0 | 2 | 2 |
| Всего | 6 | 9 | 14 | 51 | 57 |
| Euglenophyta | |||||
| Всего | 2 | 10 | 8 | 27 | 35 |
| Dinophyta | |||||
| Всего | 0 | 5 | 4 | 11 | 13 |
| Cryptophyta | |||||
| Всего | 1 | 1 | 0 | 2 | 3 |
| Chrysophyta | |||||
| Chrysophyceae | 7 | 0 | 10 | 9 | 12 |
| Uroglena | 0 | 0 | 0 | 1 | 1 |
| Uroglenopsis | 0 | 0 | 0 | 1 | 1 |
| Dinobryon | 4 | 0 | 6 | 7 | 7 |
| Epipyxis | 2 | 0 | 2 | 0 | 2 |
| Kephyrion | 1 | 0 | 0 | 0 | 1 |
| Pseudokephyrion | 0 | 0 | 1 | 0 | 1 |
| Chrysosphaerella | 0 | 0 | 1 | 0 | 1 |
| Synurophyceae | 4 | 0 | 7 | 4 | 7 |
| Mallomonas | 2 | 0 | 4 | 2 | 4 |
| Synura | 2 | 0 | 2 | 2 | 2 |
| Всего | 11 | 0 | 17 | 13 | 19 |
| Bacillariophyta | |||||
| Coscinodiscophyceae | 2 | 9 | 8 | 11 | 11 |
| Fragilariophyceae | 5 | 6 | 6 | 11 | 11 |
| Bacillariophyceae | 9 | 7 | 41 | 53 | 77 |
| Всего | 16 | 22 | 55 | 75 | 99 |
| Xanthophyta | |||||
| Всего | 0 | 3 | 1 | 4 | 4 |
| Raphidophyta | |||||
| Всего | 0 | 0 | 0 | 4 | 4 |
| Chlorophyta | |||||
| Ulotrichales | 4 | 4 | 3 | 5 | 11 |
| Chlamidomonadales | 3 | 3 | 3 | 4 | 4 |
| Volvocales | 0 | 0 | 0 | 3 | 3 |
| Chlorococcales | 1 | 1 | 2 | 4 | 5 |
| Sphaerophorales | 9 | 25 | 17 | 50 | 51 |
| Oedogoniales | 0 | 0 | 0 | 1 | 1 |
| Chlorellales | 2 | 7 | 7 | 8 | 8 |
| Всего | 19 | 40 | 32 | 75 | 83 |
| Streptophyta | |||||
| Zygnematales | 0 | 0 | 0 | 3 | 3 |
| Desmidiales | 8 | 18 | 15 | 25 | 36 |
| Всего | 8 | 18 | 15 | 28 | 39 |
| Весь фитопланктон | 63 | 108 | 146 | 290 | 356 |
В составе фитопланктона наиболее разнообразно представлены зеленые (Chlorophyta, Streptophyta) и диатомовые водоросли (табл. 2). Максимальным числом таксонов характеризовались порядки Sphaeropleales с преобладанием родов Scenedesmus и Pediastrum (по 7 таксонов) и Desmidiales, включаюший роды Staurastrum (12), Cosmarium (9) и Closterium (9).
В закисляемых водоемах не выявлено высокого обилия ацидофильных десмидиевых водорослей, за исключением часто встречавшегося в небольшом количестве Closterium aciculare T. West. Зигнемовые водоросли родов Mougeotia и Zygnema отмечены преимущественно в озерах фоновой и буферной зон. Во всех водоемах присутствовали зеленые водоросли Botryococcus braunii Kützing, Dictyosphaerium pulchellum H.C. Wood, Koliella longiseta (Vischer) Hindák, Scenedesmus acuminatus (Lagerheim) Chodat, формы Pediastrum boryanum (Turpin) Meneghini. Наибольшей встречаемостью характеризовались P. boryanum var. boryanum (62%), P. boryanum var. cornutum (Raciborski) Sulek (56%), P. duplex Meyen var. duplex (49%) и Dictyosphaerium pulchellum (48%). В зоне влияния комбината и буферной зоне виды рода Pediastrum и жгутиковые формы Chlamidomonadales обнаружены не только в период открытой воды, но и подо льдом. В наиболее загрязняемом Карабашском пруду выявлены зеленые и стрептофитовые водоросли с деформированными хроматофорами.
Во всех водоемах вегетировали диатомовые водоросли Synedra acus Kützing, разновидности S. ulna (Nitzsch) Ehrenberg, Tabellaria flocculosa (Roth) Kützing. Наибольшей встречаемостью в пяти озерах буферной и фоновой зон характеризовались Asterionella formosa Hassal (57%), Aulacoseira italica var. italica (Kützing) Simonsen (52%) и Fragilaria crotonensis Kitton (45%), отсутствовавшие в самых загрязняемых Карабашском и Богородском прудах. Из представителей Coscinodiscophyceae в зоне влияния комбината обнаружены Cyclotella meneghiniana Kützing и единичные пустые створки Aulacoseira granulatа (Ehreberg) Simonsen. Обычный планктонный вид озер фоновой зоны Handmannia comta (Grunov) Kociolek et Khursevich (Cyclotella radiosa Grunov) отсутствовал в импактной зоне и был отмечен единично в отдельных озерах буферной зоны. В зоне влияния комбината зарегистрированы клетки диатомей с морфологическими отклонениями в виде искривленных створок.
Видовое разнообразие Euglenophyta в импактной зоне также существенно снижалось по сравнению с фоновой, причем в буферной зоне найдены виды, отсутствующие в фоновой. Наиболее представительны были род Trachelomonas (22), не обнаруженный в импактной зоне, и род Phacus (8), который вместе с Euglena единично встречался в этой зоне.
Видовое богатство золотистых в меньшей степени менялось при ацидификации и росте концентрации тяжелых металлов, при этом в импакт-буферной зоне (оз. Серебры) эти водоросли не обнаружены (табл. 2). В шести водоемах отмечены Dinobryon sociale var. americanum (Brunnthaler) Bachmann, в пяти – D. bavaricum Imhof var. bavaricum, D. divergens var. angulatum (Seligo) Brunnthaler и Synura petersenii Korschikov. В озерах буферной зоны число семейств Chrysophyta, а также число видов рода Mallomonas было выше, чем в фоновой зоне.
Максимальное варьирование видового разнообразия выявлено для цианопрокариот Chroococcales, Oscillatoriales, Nostocales: сильное снижение в наиболее загрязняемых водоемах и значительное увеличение количества видов синезеленых в нейтральных гидрокарбонатных озерах фоновой зоны. Во всех озерах отмечена Oscillatoria tenuis Agardh еx Gomont, в шести – Planktolyngbya limnetica (Lemmermann) Komárkova-Legnerova et Cronberg и Limnothrix planctonica (Woloszki) Meffert. Виды Мicrocystis и большинство родов Chroococcales встречались только в фоновой зоне. В пределах одной зоны видовой состав цианопрокариот был сходным (табл. 2).
При низких рН и увеличении концентрации металлов происходило уменьшение общего видового богатства фитопланктона (рис. 2а). Доминирующие комплексы изменялись в градиенте ацидности и концентрации металлов. Для водоемов зоны влияния Карабашского комбината характерно преобладание золотистых водорослей как в подледный, так и в период открытой воды. К доминирующим относились виды Dinobryon sertularia Ehrenberg var. sertularia, D. sociale var. americanum (до 120 тыс. кл./дм3 в Богородском пруду), к субдоминантам – Synura petersenii. Они развивались при концентрации меди в поверхностном слое ~0.02 мг/дм3, цинка >0.20 мг/дм3, что согласуется с результатами, полученными для некоторых видов в лабораторных экспериментах [3]. В периоды с увеличенным поверхностным стоком в доминирующем комплексе отсутствовали золотистые и преобладали цианопрокариоты Planktolyngbya limnetica, Limnothrix planctonica, а также диатомовые Achnanthes minutissima Kützing var. minutissima. В оз. Серебры круглогодично доминировали зеленые (Sphaerophorales) и диатомовые водоросли, летом периодически отмечались слабые вспышки Aphanizomenon (Cyanoprokaryota). В озерах буферной зоны развивался диатомово-хризофитовый комплекс (Dinobryon divergens Imnof var. divergens, Mallomonas punctifera Korschikov, Rhizosolenia longiseta Zacharias, Tabellaria fenestrata (Lyngbye) Kützing, T. flocculosa (Roth) Kützing) с участием крупных видов Desmidiales (Streptophyta).
Рис. 2.
Видовое разнообразие фитопланктона (а) и состав доминирующих комплексов (б) в водоемах импактного воздействия Карабашского месторождения: 1 – Cyanophyta, 2 – Euglenophyta, 3 – Dinophyta, 4 – Cryptophyta, 5 – Chrysophyta, 6 – Bacillariophyta, 7 – Chlorophyta, 8 – Streptophyta. По оси ординат: а – число таксонов рангом ниже рода, б – доля водорослей разных отделов; по оси абсцисс: I – фоновая зона, II – буферная, III – импакт-буферная, IV – импактная.
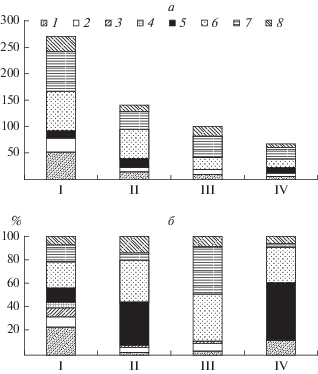
Озера фоновой зоны, в меньшей степени подверженные ацидификации и загрязняющиеся только воздушным путем, характеризовались более сложной структурой доминирующего комплекса. В его состав входили представители Bacillariophyta, Chrysophyta, Dinophyta, Chlorophyta и Streptophyta, в отдельные годы летом доминировали Euglenophyta, Raphidophyta и Cryptophyta (рис. 2б), ежегодно отмечено развитие Cyanoprokaryota до степени визуального “цветения” воды.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Данные по таксономической структуре фитопланктона малых озер Южного Урала в зоне влияния Карабашского горно-обогатительного и медеплавильного комбината продемонстрировали существенное снижение видового богатства водорослей по техногенному градиенту и градиенту рН среды.
Уменьшение количества видов цианопрокариот с ростом загрязнений наблюдается в водоемах различных регионов [19, 23]. Наиболее устойчивыми к техногенному воздействию оказались виды Planktolyngbya limnetica и Limnothrix planctonica, доминирующие в закисленных водах зоны влияния комбината. Известно, что ацидификация нарушает цикл азота за счет подавления процесса нитрификации, протекающего в щелочной среде [37]. Потребление иона аммония приводит в конечном итоге к активизации фермента уреазы и гибели азотфиксаторов [4, 5]. При периодическом поступлении нейтральных вод рН в озерах колеблется от кислого до нейтрального, что компенсирует негативное для развития цианопрокариот увеличение кислотности и создает условия, отличные от таковых в ацидных олиготрофных водах [6].
Известно, что зеленые водоросли доминируют в кислых озерах [7, 24]. Это отмечено и в оз. Серебры при значительном снижении рН из-за кислых осадков из дымовых шлейфов и поровых вод донных отложений с рН 3.0–3.5, дефиците нитрит- и нитрат-ионов (общий азот 0.102–0.328 мг/дм3). В лабораторных условиях при дефиците азота ионы аммония не оказывали ингибирующего действия на хлорококковые водоросли из-за снижения интенсивности гидролиза мочевины [4]. При добавлении мочевины увеличивалась численность зеленых водорослей, а цианопрокариот снижалась. У некоторых представителей Chlorophyta обнаружена высокая активность глутаматдегидрогеназы, ослабляющей влияние аммиака [4]. Dictyosphaerium pulchellum вместе и с Pediastrum sp. ежегодно превалировали в оз. Серебры в период открытой воды. Толерантность D. pulchellum к низким рН и азотному лимитированию [28] дает ему конкурентные преимущества. Виды Pediastrum boryanum и P. duplex в массе (до 200 тыс. кл./дм3) развивались в поверхностном слое озера до поздней осени.
Увеличение разнообразия десмидиевых водорослей отмечено в исследуемом ряду водоемов: с 8 видов в загрязняемых закисляемых до 25 в фоновых нейтральных (табл. 2). Во всех исследованных водоемах (загрязненных и фоновых) не наблюдали увеличения разнообразия и обилия десмидиевых и зигнемовых водорослей, которые (за исключением Closterium aciculare) встречались единично. На развитие стрептофитовых водорослей-ацидофилов в большей степени может влиять наличие минеральной взвеси, чем рН среды. Снижение разнообразия и обилия различных групп водорослей под влиянием минеральной пыли и повышенных концентраций металлов в средах показано также для других лесных регионов [19, 22].
Несмотря на загрязнение ионами металлов, вода в озерах буферной зоны и зоны влияния комбината остается слабоминерализованной пресной и ультрапресной с электропроводностью от 86 до 416 мкСм/см (табл. 1), обычной для обитания золотистых водорослей. Периодическое снижение рН в водоемах импактной и буферной зоны приводит к увеличению ферментативной активности клеток водорослей. Это способствует их обеспечению минеральным питанием при нарушениях обмена или дефиците доступных форм фосфора [3, 38] и относится к числу адаптивных приспособлений к закислению.
На содержание доступных форм фосфора в воде влияет присутствие катионов металлов-комплексообразователей, которые осаждают анионы биогенных веществ. В условиях слабой биодоступности биогенов источником питания для водорослей становятся бактерии, в водоемах импактной зоны – в основном железобактерии, окисляющие двухвалентное железо до трехвалентного [18] и освобождающие при этом энергию для усвоения углерода. Миксотрофия золотистых водорослей также дает преимущество в этих условиях. Возрастание роли миксотрофных флагеллят (к ним относятся золотистые водоросли) при увеличении продукции бактериопланктона отмечено и в водохранилищах Волжского каскада [7, 32]. В отличие от миксотрофных золотистых, автотрофные зеленые водоросли ограничены условиями прозрачности и доступности биогенных веществ, поэтому увеличения их таксономического разнообразия в исследованных ацидных озерах не происходит (остается низким).
Наибольшее количество золотистых водорослей отмечают в диапазоне рН 5.5–6.5 [39], а наибольшее их разнообразие в водоемах Севера России – при рН 5.5–7.5 [3]. Такие условия типичны для поверхностных слоев исследованных водоемов. В озерах фоновой зоны с рН 8.5–9.4 изменяется карбонатное равновесие и золотистые не способны использовать углерод из-за нарушения проницаемости клеточной оболочки [38]. Поэтому даже при низком уровне загрязнения металлами наблюдается уменьшение видового богатства и отсутствие доминирования этих водорослей.
Выводы. Таксономическое разнообразие фитопланктона малых озер Южного Урала под воздействием техногенных выбросов уменьшается от 290 таксонов рангом ниже рода в фоновой зоне до 62 в импактной зоне. Наиболее низкое видовое богатство отмечено у цианопрокариот из порядков Chroococcales, Oscillatoriales и Nostocales. Снижение числа видов выявлено для зеленых водорослей Chlorophyta (Sphaeropleales, Chlorellales) и Streptophyta. В буферной зоне техногенеза в сульфатных кислых водах отмечено увеличение таксономического богатства и обилия представителей Chrysophyceae и Synurophyceae (Chrysophyta), Sphaerophorales и Chlorellales (Chlorophyta). Периодическая ацидификация и миксотрофные условия дают конкурентные преимущества для развития золотистых водорослей в водоемах импактной и буферной зоны техногенеза.
Список литературы
Аминов П.Г. Биогеохимия тяжелых металлов при горнопромышленном техногенезе (на примере Карабашской геотехнической системы, Южный Урал): Автореф. дис. … канд. г.-м. наук. Новосибирск, 2010. 17 с.
Ветрова З.И. Эвгленофитовые водоросли // Флора континентальных водоемов Украинской ССР. Киев: Наук. думка, 1993. Вып. 1. Ч. 2. 260 с.
Волошко Л.Н. Золотистые водоросли водоемов Севера России. СПб.: Реноме, 2017. 380 с.
Клоченко П.Д., Грубинко В.В., Гуменюк Г.Б., Арсан О.М. Особенности ассимиляции аммонийного азота зелеными и синезелеными водорослями // Гидробиол. журн. 2002. Т. 38. № 2. С. 88–93.
Клоченко П.Д., Сакевич А.И., Усенко О.М., Шевченко Т.Ф. Изменение структуры фитопланктона под воздействием мочевины // Гидробиол. журн. 2000. Т. 36. № 6. С. 62–74.
Корнева Л.Г. Альгофлора планктона водохранилищ Волжского бассейна // Ботан. журн. 2008. Т. 93. № 11. С. 1673–1690.
Корнева Л.Г. Таксономический состав, экология Chlorophyta и Streptophyta в слабоминерализованных мелководных лесных озерах // Альгология. 2012. № 3. С. 259–274.
Косинская Е.К. Конъюгаты, или сцеплянки. Десмидиевые водоросли // Флора споровых растений СССР. М.; Л.: Наука, 1960. Т. 5. Вып. 1. 706 с.
Крахмальный А.Ф. Динофитовые водоросли Украины (иллюстрированный определитель). Киев: Альтерпрес, 2011. 444 с.
Масленникова А.В., Удачин В.Н., Дерягин В.В. Палеоэкология и геохимия озерной седиментации голоцена Урала. Екатеринбург: Ильменск. гос. заповедник УрО РАН, 2014. 136 с.
Методические рекомендации по сбору и обработке материалов при гидробиологических исследованиях на пресноводных водоемах. Фитопланктон и его продукция. Л.: ГосНИИ озер. и реч. рыб. хоз-ва, 1984. 31 с.
Мошкова Н.А., Голлербах М.М. Зеленые водоросли. Класс Улотриксовые. Порядок Улотриксовые // Определитель пресноводных водорослей СССР. Л.: Наука, 1986. Вып. 10. Ч. 1. 360 с.
Оксиюк О.П., Жукинский В.П., Брагинский Л.П. и др. Комплексная экологическая классификация качества поверхностных вод суши // Гидробиол. журн. 1993. Т. 29. № 4. С. 62–76.
Рундина Л.А. Зигнемовые водоросли России (Chlorophyta: Zygnematophyceae, Zygnematales). СПб.: Наука, 1998. 351 с.
Снитько Л.В. Экология и сукцессии фитопланктона озер Южного Урала. Миасс: Ильменск. гос. заповедник УрО РАН, 2009. 376 с.
Снитько Л.В., Снитько В.П. Индикаторная роль фитопланктона в оценке многолетних изменений качества воды озер Большое Миассово и Тургояк (Южный Урал) // Вод. ресурсы. 2014. Т. 41. № 2. С. 216–224.
Снитько Л.В., Снитько В.П. Многолетние изменения фитопланктона Аргазинского водохранилища (Южный Урал) // Вод. ресурсы. 2015. Т. 42. № 2. С. 232–240.
Удачин В.Н., Аминов П.Г., Филиппова К.А. Геохимия горнопромышленного техногенеза Южного Урала. Екатеринбург: УрО РАН, 2014. 251 с.
Хайбуллина Л.С., Суханова Н.В., Кабиров Р.Р. Флора и синтаксономия почвенных водорослей и цианобактерий урбанизированных территорий. Уфа: Гилем, 2011. 216 с.
Царенко П.М. Краткий определитель хлорококковых водорослей Украинской ССР. Киïв: Наук. думка, 1990. 208 с.
Царенко П.М. Номенклатурно-таксономические изменения в системе “зеленых водорослей” // Альгология. 2005. Т. 15. № 4. С. 459–467.
Шилова Н.А. Влияние тяжелых металлов на представителей пресноводного фито- и зоопланктона в условиях засоления: Дис. … канд. биол. наук. Саратов, 2014. 133 с.
Шкундина Ф.Б., Асадуллина Г.Р. Оценка экологического состояния рек на территории города на основании классификации по показателям развития планктонных водорослей, цианобактерий и биотестирования // Вода: химия и экология. 2013. № 2. С. 26–30.
Almer B., Dickson W., Ekstrum C., Hurnstrum E. Effects of acidification on Swedish lakes // Ambio. 1974. № 1. P. 30–36.
Coesel Peter F.M., Meesters K.J. Desmids of the Lowlands. Mesotaeniaceae and Desmidaceae of the European Lowlands. Zeist: KNNV Publ., 2007. 351 p.
Ettl H. Chlorophyta. Phytomonadina // Susswasserflora von Mitteleuropa. Jena: VEB Gustav Fischer Verlag, 1983. Bd 9. 807 p.
Ettl H., Gartner G. Chlorophyta II. Tetrasporales, Chlorococcales, Gloedendrales // Susswasserflora von Mitteleuropa. Jena: VEB Gustav Fischer Verlag, 1983. Bd 10. 436 p.
Irfanullah H.M., Moss B. Ecology of Dictyosphaerium pulchellum Wood (Chlorophyta, Chlorococcales) in a shallow, acid, forest lake // Aquat. Ecol. 2006. V. 40. № 1. P. 1–12.
Komárek J., Anagnostidis K. Modern approach to the classification system of cyanophytes. 4. Nostocales // Archiv für Hydrobiol. 1989. V. 82. P. 247–345.
Komárek J., Anagnostidis K. Cyanoprocaryota. 1. Teil. Chroococcales // Süsswasserflora Mitteleuropa. 19/1. Jena: Fischer Verlag, 1998. 548 p.
Komárek J., Anagnostidis K. Cyanoprocaryota. 2. Teil. Oscillatoriales // Süsswasserflora Mitteleuropa. 19/2. München. 2005. 759 p.
Korneva L.G., Solovyeva V.V. Golden algae (Chrysophyta) in plankton of the Volga River reservoirs: Taxonomic structure, dynamics of diversity, and abundance. Inland Water Biol. 2017. V. 10. № 2. P. 168–175. https://doi.org/10/7868/S0320965217020073
Krammer K., Lange-Bertalot H., Bacillariophyceae 1–4 Teil // Süsswasserflora von Mitteleuropa. Stuttgart: Gustav Fischer Verlag, 1991. 876 pp., 596, 576, 434 pp.
Kristiansen J. Biogeography of silica-scaled chrysophytes // Nova Hedwigia. Beih. 2001. V. 122. P. 23–39.
Kristiansen J., Preising H.R. Chrysophyte and Haptophyte Algae. 2. Teil: Synurophyceae // Süsswasserflora von Miteleuropa. Heidelberg: Spektrum Akad. Verlag, 2007. 252 p.
Lenzenweger R. Desmidiaceenflora von Ősterreich T. 1–3 // Bibliotheca Phycologica. Bd 101–104. Berlin; Stuttgart. 1996. 1997. 1998. 162 pp., 216, 218 pp.
Rudd J.W. M., Kelly C.A., Schindler D.W., Turner M.A. Distribution of the nitrogen cycle in acidified lakes // Science. 1988. V. 240. P. 1515–1517.
Siver P.A. The distribution of chrysophytes along environmental gradients: their use as biological indicators // Chrysophyte algae (ecology, phylogeny and development). N.Y.: Cambridge Univ. Press, 1995. P. 232–269.
Siver P.A., Hamer J.S. Multivariate statistical analysis of the factors controlling the distribution of scaled chrysophytes // Limnol., Oceanogr. 1989. V. 34. P. 368–381.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод