

Биология внутренних вод, 2019, № 4-2, стр. 44-54
Изменения структуры макрозообентоса Рыбинского водохранилища в условиях повышения температуры
С. Н. Перова *
Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
Некоузский р-н, Ярославская обл., пос. Борок, Россия
* E-mail: perova@ibiw.yaroslavl.ru
Поступила в редакцию 26.01.2017
После доработки 24.07.2017
Принята к публикации 29.05.2018
Аннотация
Анализ результатов многолетних наблюдений выявил изменения видового состава и значительный рост количественного обилия макрозообентоса глубоководной зоны Рыбинского водохранилища в экстремально теплые годы начала ХХI в. После аномально жаркого лета 2010 г. видовое богатство и разнообразие снизились до минимальных значений. Наблюдавшийся в последующие годы пик численности и биомассы личинок хирономид рода Chironomus и полисапробных видов олигохет, – по-видимому, результат увеличения темпов эвтрофирования.
ВВЕДЕНИЕ
С начала XXI в. все актуальнее становится проблема потепления климата и его последствий для наземных и водных экосистем. Рыбинское водохранилище – крупнейшее и наиболее изученное в Волжском каскаде. По результатам многолетних наблюдений на его побережье с 1976 г. зарегистрирован устойчивый рост температуры воздуха – главного индикатора изменения климата (Литвино, Законнова, 2014) . По данным Росгидромета, в 2009 г. все сезоны были “незначительно теплее нормы”, и ни один не относился к экстремальным, не только в среднем по России, но и по каждому из рассматриваемых регионов (Доклад ..., 2010). Поэтому 2009 г., с близкими к среднемноголетним температурам воздуха и воды в районе Рыбинского водохранилища, можно считать “фоновым” при сравнении с последующими годами изучения. Следующий, 2010 г. сложился из экстремально холодной зимы, рекордно жаркого лета и экстремально теплой осени. По данным Ярославского центра по гидрометеорологии и мониторингу окружающей среды, 2010 г. по многим показателям (продолжительность летней жары, атмосферной и почвенной засухи) характеризовался как аномальный за >100-летний ряд наблюдений (Климатические ..., 2006–2010). В последующие годы тенденция повышения средней температуры воздуха продолжалась: в ряд самых теплых лет за период наблюдений вошли 2011–2014 гг. (Доклад ..., 2011, 2012, 2013, 2014, 2015). По всем данным мировых климатических центров, 2015 г. оказался самым теплым в глобальном масштабе за все время наблюдений, а предыдущий максимум повышения средней температуры наблюдался годом ранее (Доклад ..., 2015, 2016).
Важная часть экосистемы водохранилища – донные макробеспозвоночные, участвующие в трансформации органического вещества в системе толща воды–донные отложения, что обусловливает их значительную роль в процессах самоочищения вод. По изменениям таксономического состава и обилия макрозообентоса можно судить о состоянии всей экосистемы.
Цель работы – охарактеризовать изменения структуры макрозообентоса глубоководной зоны Рыбинского водохранилища в условиях экстремально высоких температур.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Материалом для работы послужили результаты мониторинга макрозообентоса на стандартных станциях, расположенных в глубоководной зоне Главного и Волжского плесов Рыбинского водохранилища (рис. 1). Макрозообентос собирали в сентябре–октябре 2009–2015 гг. с интервалом в ~2 нед. В эти годы измеряли температуру в поверхностном и придонном слое воды в течение вегетационного сезона с мая по октябрь, также с интервалом в ~2 нед. Пробы грунта отбирали модифицированным дночерпателем Экмана-Берджа (ДАК-250) с площадью захвата 1/40 м2 и ДАК-100 с площадью захвата 1/100 м2, по 2 подъема на каждой станции. Сбор, разборку, камеральную и статистическую обработку собранного материала проводили по стандартной методике (Методика ..., 1975). Всего собрано 115 проб. Для оценки состояния сообществ макрозообентоса использовали: численность (N), биомассу (B), частоту встречаемости (Р), количество видов (S) и индекс видового разнообразия Шеннона–Уивера, рассчитанный по численности (HN).
Рис. 1.
Схема расположения гидробиологических станций в Рыбинском водохранилище. Плесы водохранилища: I – Волжский, II – Моложский, III – Шекснинский, IV – Главный; станции: 1 – Коприно, 2 – Молога, 3 – Наволок, 4 – Измайлово, 5 – Средний Двор, 6 – Брейтово; 1 – русла затопленных рек, 2 – границы плесов.
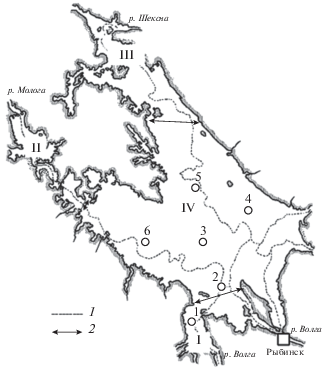
Большинство стандартных станций было расположено в русловых участках глубоководной зоны (рис. 1) и относилось к биотопу серых (глинистых) илов. Они находились на участках бывших русел рек Волга (ст. 1) и Молога (ст. 6), в устье р. Молога (ст. 2) с интенсивным илонакоплением, а также в северной части Главного плеса у затопленного с. Средний Двор (ст. 5) (рис. 1). Глубина на этих станциях достигала 12–16 м. На ст. 3, расположенной в центральной части Главного плеса (у бывшего с. Наволок) глубина была 5–8 м, мозаичный грунт состоял из торфянистого ила, почвы, песка и ракушечника. На ст. 4 (бывшая пойма р. Шексны в восточной части Главного плеса, напротив с. Измайлово) глубина не превышала 5–8 м, донные отложения представлены гравием и песком, иногда встречались остатки размытых торфянистых почв.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
По результатам наблюдений осенью 2009–2015 гг. в глубоководной зоне Рыбинского водохранилища выявлено 90 видов донных макробеспозвоночных: лидировали моллюски (27 видов), хирономиды (24) и олигохеты (24). Наибольшее число видов (62) зарегистрировано на ст. 2. Удельное видовое богатство макрозообентоса было значительно выше на серых илах русловых участков (ст. 1, 2, 6), чем на участках с другими типами донных отложений (табл. 1).
Таблица 1.
Видовое богатство и разнообразие макрозообентоса в глубоководной зоне Рыбинского водохранилища (станции 1–6)
| Год | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| 2009 | $\frac{{2.38--2.40}}{{9--11}}$ | $\frac{{2.66--3.16}}{{15--17}}$ | $\frac{{1.62--1.93}}{{4--8}}$ | $\frac{{0.00--1.79}}{{1--4}}$ | $\frac{{0.81--1.60}}{{2--4}}$ | $\frac{{2.38--2.49}}{{12--19}}$ |
| 2010 | $\frac{{0.91--1.01}}{{4--6}}$ | $\frac{{2.43--2.49}}{{9--11}}$ | $\frac{{0.00--0.79}}{{1--2}}$ | $\frac{{0.00--0.54}}{{1--2}}$ | $\frac{{0.00--1.00}}{{0--2}}$ | $\frac{{1.84--2.72}}{{6--12}}$ |
| 2011 | $\frac{{1.00--2.23}}{{5--10}}$ | $\frac{{2.45--2.63}}{{6--12}}$ | $\frac{{2.16--2.86}}{{5--9}}$ | $\frac{{1.79--2.55}}{{7--8}}$ | $\frac{{0.65--1.46}}{{2--3}}$ | $\frac{{2.63--3.35}}{{15--16}}$ |
| 2012 | $\frac{{2.11--2.96}}{{14--17}}$ | $\frac{{2.40--2.60}}{{9--23}}$ | $\frac{{2.54--2.70}}{9}$ | $\frac{{1.50--1.58}}{3}$ | $\frac{{2.28--2.37}}{{9--12}}$ | $\frac{{2.14--2.22}}{{15--17}}$ |
| 2013 | $\frac{{2.24--2.71}}{{24}}$ | $\frac{{2.81--2.85}}{{21--23}}$ | $\frac{{2.35--2.37}}{{7--18}}$ | $\frac{{2.16--2.76}}{{5--10}}$ | $\frac{{2.37--2.49}}{{6--8}}$ | $\frac{{1.78--2.68}}{{18--19}}$ |
| 2014 | $\frac{{2.18--2.44}}{{10}}$ | $\frac{{2.03--2.38}}{{10--14}}$ | – | – | – | – |
| 2015 | $\frac{{2.10--2.16}}{{12--16}}$ | $\frac{{2.53--2.56}}{{17--20}}$ | $\frac{{1.43--2.13}}{{5--12}}$ | $\frac{{2.58--2.91}}{{8--9}}$ | $\frac{{2.25--2.31}}{{8--9}}$ | $\frac{{2.11--2.64}}{{14--21}}$ |
Примечание. Над чертой – индекс видового разнообразия Шеннона–Уивера HN, бит/экз., под чертой – число видов в пробе. “–” – здесь и далее данные отсутствуют; названия станций см. рис. 1.
Индекс видового разнообразия Шеннона–Уивера колебался в широких пределах, наиболее высокие значения отмечены на биотопе серых илов. Осенью 2010 г. число видов макрозообентоса на всех станциях снизилось в ~2 раза по сравнению с 2009 г. (табл. 1). На большинстве станций значительно сократилось количество видов моллюсков, из состава сообществ исчезли ракообразные и пиявки (из группы “прочие”) (рис. 2). Не встречен каспийский вселенец – пиявка Archaeobdella esmonti Grimm, впервые обнаруженная в Рыбинском водохранилище в 2009 г. В течение вегетационного сезона 2009 г. она имела высокую частоту встречаемости, численность и биомассу (Perova, 2012). Осенью 2010 г. A. esmonti не зарегистрирована, в 2011 г. отмечена в единственном экземпляре на ст. 4, в 2012–2015 гг. ее находили на тех же станциях, что и в 2009 г., но изредка и единично. Моллюски-дрейссениды, регулярно встречавшиеся на стандартных станциях водохранилища в течение вегетационного сезона 2009 г., после аномально жаркого лета 2010 г. стали отмечаться значительно реже, а их численность и биомасса, по сравнению с 2009 г., снизились в ~10–20 раз. В течение всего вегетационного периода 2012 г. и с весны до августа 2013 г. дрейссенид на стандартных станциях Рыбинского водохранилища не регистрировали. Начиная с августа 2013 г., их обнаруживали редко и в небольшом количестве, в 2014 и 2015 гг. они стали встречаться чаще, их обилие увеличилось. Однако максимальные значения численности и биомассы дрейссенид, зарегистрированные в июле 2014 г. на ст. Брейтово (300 экз./м2 и 109.4 г/м2 соответственно) были существенно ниже, чем в 2009 г. (Perova, 2012) .
Рис. 2.
Таксономическая структура макрозообентоса Рыбинского водохранилища: а – ст. 1, б – ст. 2, в – ст. 3, г – ст. 4, д – ст. 5, е – ст. 6. 1 – хирономиды, 2 – олигохеты, 3 – моллюски, 4 – прочие.
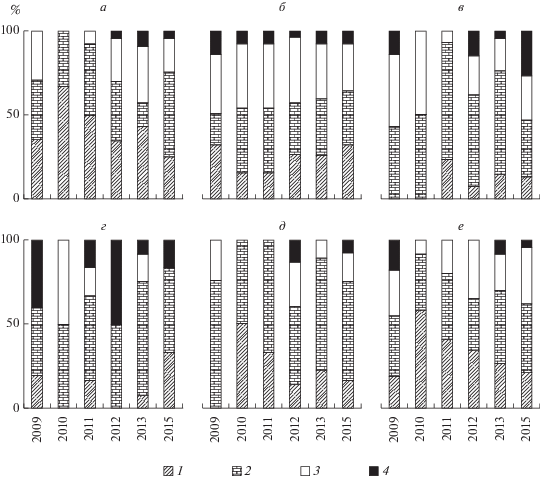
После аномально жаркого лета 2010 г., погодные условия которого негативно повлияли на макрозообентос водохранилища и вызвали значительное снижение его видового богатства и изменения структуры, сообщества донного населения постепенно восстанавливались. В последующие годы наблюдался рост видового богатства, его максимум на большинстве станций отмечен осенью 2012 и 2013 гг. (табл. 1).
Количественное обилие макрозообентоса на участках, занятых серыми илами, во все периоды наблюдений было намного выше, чем на других грунтах (табл. 2). В 2011–2015 гг. на серых илах отмечен существенный рост численности и биомассы, значения которых увеличились в десятки раз по сравнению 2009 г. Максимальные средние величины биомассы зарегистрированы осенью 2015 г. на ст. 2 и 6 (табл. 2). По структуре численности и биомассы макрозообентоса исследованные участки существенно различались (рис. 3 и 4). На ст. 3 и 4 (с мозаичным типом донных отложений) по численности и биомассе лидировали олигохеты и хирономиды, при этом, в отдельные годы (2009, 2012, 2015) преобладала группа “прочие”, в которую входили пиявки и бокоплав Gmelinoides fasciatus (Stebbing) (рис. 3в, 4в, 4г, 4е). На участках, занятых серыми илами, по численности, в основном, преобладали олигохеты, их доля значительно увеличилась за период с 2009 по 2015 гг. (рис. 3). Основу биомассы, в большинстве случаев, составляли личинки хирономид, среди которых доминировали представители рода Chironomus (рис. 4а, 4б, 4д, 4е).
Таблица 2.
Численность и биомасса макрозообентоса в глубоководной зоне Рыбинского водохранилища (станции 1–6)
| Год | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| 2009 | $\frac{{0.67 \pm 0.27}}{{10.74 \pm 2.92}}$ | $\frac{{1.96 \pm 0.30}}{{17.94 \pm 0.89}}$ | $\frac{{0.26 \pm 0.04}}{{0.63 \pm 0.41}}$ | $\frac{{0.20 \pm 0.08}}{{0.27 \pm 0.04}}$ | $\frac{{0.70 \pm 0.46}}{{1.58 \pm 0.79}}$ | $\frac{{2.47 \pm 1.27}}{{18.54 \pm 12.22}}$ |
| 2010 | $\frac{{2.27 \pm 0.17}}{{27.14 \pm 5.82}}$ | $\frac{{0.82 \pm 0.34}}{{4.49 \pm 2.73}}$ | $\frac{{0.13 \pm 0.03}}{{0.24 \pm 0.07}}$ | $\frac{{0.10 \pm 0.06}}{{0.31 \pm 0.27}}$ | $\frac{{0.02 \pm 0.02}}{{0.59 \pm 0.59}}$ | $\frac{{1.89 \pm 0.87}}{{14.19 \pm 4.68}}$ |
| 2011 | $\frac{{2.52 \pm 0.60}}{{48.42 \pm 5.76}}$ | $\frac{{2.79 \pm 1.17}}{{39.57 \pm 6.42}}$ | $\frac{{0.27 \pm 0.11}}{{0.95 \pm 0.67}}$ | $\frac{{0.48 \pm 0.12}}{{0.73 \pm 0.08}}$ | $\frac{{0.18 \pm 0.60}}{{1.91 \pm 0.84}}$ | $\frac{{7.97 \pm 3.05}}{{63.22 \pm 17.44}}$ |
| 2012 | $\frac{{6.17 \pm 2.55}}{{21.81 \pm 8.83}}$ | $\frac{{38.25 \pm 0.31}}{{82.09 \pm 0.18}}$ | $\frac{{0.49 \pm 0.05}}{{1.15 \pm 0.32}}$ | $\frac{{0.07 \pm 0.01}}{{0.18 \pm 0.01}}$ | $\frac{{2.77 \pm 1.35}}{{10.15 \pm 5.44}}$ | $\frac{{39.37 \pm 8.77}}{{108.33 \pm 10.07}}$ |
| 2013 | $\frac{{24.78 \pm 7.12}}{{77.97 \pm 21.84}}$ | $\frac{{39.62 \pm 5.80}}{{57.76 \pm 6.22}}$ | $\frac{{1.39 \pm 1.03}}{{3.07 \pm 2.72}}$ | $\frac{{0.43 \pm 0.27}}{{0.33 \pm 0.16}}$ | $\frac{{1.15 \pm 0.71}}{{8.60 \pm 2.58}}$ | $\frac{{32.75 \pm 9.85}}{{106.74 \pm 5.67}}$ |
| 2014 | $\frac{{1.88 \pm 0.62}}{{14.73 \pm 3.82}}$ | $\frac{{11.87 \pm 0.03}}{{31.20 \pm 11.84}}$ | – | – | – | – |
| 2015 | $\frac{{11.53 \pm 2.33}}{{28.40 \pm 2.76}}$ | $\frac{{44.69 \pm 21.17}}{{150.73 \pm 54.45}}$ | $\frac{{1.34 \pm 1.20}}{{1.90 \pm 1.48}}$ | $\frac{{0.37 \pm 0.05}}{{0.63 \pm 0.02}}$ | $\frac{{2.81 \pm 1.85}}{{18.88 \pm 15.58}}$ | $\frac{{36.38 \pm 6.76}}{{132.29 \pm 40.03}}$ |
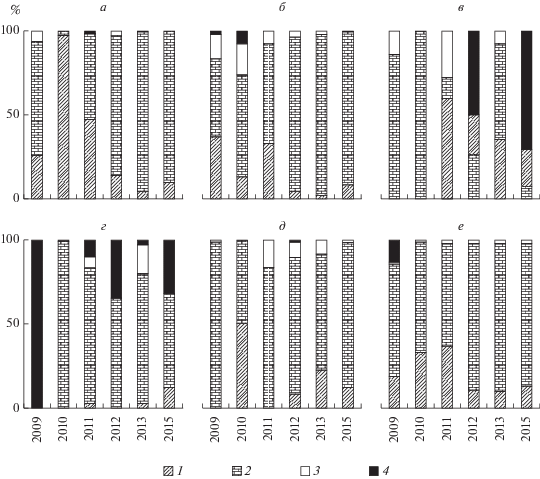

Следует отметить, что многолетняя тенденция к потеплению и вызванные этим изменения условий среды, повлияли на соотношение видов олигохет в составе макрозообентоса Рыбинского водохранилища. Так, до конца 1980-х гг. в Рыбинском водохранилище основой биомассы олигохет был β-мезосапроб Tubifex newaensis (Michaelsen), отличающийся крупными размерами особей (Архипова, 2016; Поддубная, 1988). В 1990-е гг. численность вида снизилась в 5–6 раз, а с 2010 г. он встречался редко и единично, что привело к снижению биомассы олигохет и, соответственно, общей биомассы макрозообентоса, основную долю которой уже представляли личинки хирономид. С конца 1980-х гг. начала увеличиваться численность других олигохет-тубифицид, среди которых доминировали полисапробные виды Limnodrilus hoffmeisteri Claparéde и Tubifex tubifex (Müller). Наиболее значительный рост обилия этих олигохет и их доли в общей численности макрозообентоса наблюдался на участках, покрытых серыми илами, в 2012–2015 гг. (рис. 3а, 3б, 3д, 3е).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Температура воды – один из наиболее значимых экологических факторов, существенно влияющий на гидробиологический режим водоемов. В Рыбинском водохранилище с 1976 г. прослеживается рост температуры воды, тенденция к потеплению усилилась после 2000 г. (табл. 3). В 2001–2010 гг. средняя температура воды за май–октябрь была 15°С, что на 0.9°С выше среднемноголетних значений (Литвинов, Законнова, 2014). В дальнейшем эта тенденция сохранилась: по нашим данным в 2009–2015 гг. средняя температура воды за май–октябрь достигала 16°С (табл. 3, табл. 4).
Таблица 3.
Средняя за десятилетний период температура воды (T) в глубоководной зоне Рыбинского водохранилища (по: Литвинов, Законнова, 2014)
| Период (май–октябрь) |
T, °С |
|---|---|
| 1951–1960 | 13.6 |
| 1961–1970 | 13.8 |
| 1971–1980 | 13.3 |
| 1981–1990 | 13.8 |
| 1991–2000 | 14.1 |
| 2001–2010 | 15.0 |
| 2009–2015* | 16.3 |
Таблица 4.
Среднегодовая температура воды (°C) в поверхностном и придонном слоях глубоководной зоны Рыбинского водохранилища
| Год (мая–октябрь) |
Поверхностный слой |
Придонный слой |
|---|---|---|
| 2009 | 16.9 | 15.2 |
| 2010 | 16.1 | 14.0 |
| 2011 | 18.2 | 17.2 |
| 2012 | 16.8 | 15.8 |
| 2013 | 17.5 | 15.3 |
| 2014 | 18.7 | 15.7 |
| 2015 | 15.2 | 15.3 |
Как сказано выше, 2010 г. отличался от других лет наблюдений аномальными погодными условиями (Климатические ..., 2006–2010). Летом 2010 г. зарегистрированы экстремально высокие значения поверхностной (27.9°С) и придонной (25.8°С) температуры воды (табл. 5), которые в Рыбинском водохранилище отмечались впервые за все годы наблюдений. Несмотря на это, средняя за май–октябрь температура придонного слоя воды в 2010 г. была ниже, чем в предыдущем 2009 г., и во все последующие годы наблюдений (табл. 4). Это объясняется тем, что сезонный ход температуры воды характеризовался существенной разницей ее значений в весенние, летние и осенние месяцы 2010 г. (табл. 5).
Таблица 5.
Температура воды (°С) у поверхности и дна в глубоководной зоне Рыбинского водохранилища в 2010 г. (станции 1–6)
| Дата | 1 | 2 | 3 | 4 | 5 | 6 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| П | Д | П | Д | П | Д | П | Д | П | Д | П | Д | |
| 04.V | 9.0 | 8.2 | 7.9 | 4.7 | 3.4 | 3.4 | 2.7 | 2.7 | 3.2 | 3.2 | 5.4 | 5.3 |
| 15.VI | 17.8 | 17.0 | 17.1 | 16.8 | 16.2 | 15.6 | 15.8 | 15.2 | 15.8 | 15.2 | 16.0 | 15.6 |
| 28.VI | 20.8 | 16.5 | 20.5 | 16.6 | 19.2 | 18.9 | 19.2 | 18.9 | 21.2 | 16.4 | 21.6 | 16.2 |
| 14.VII | 26.7 | 18.7 | 26.9 | 18.6 | 26.9 | 19.8 | 27.1 | 18.8 | 27.9 | 17.3 | 27.9 | 17.5 |
| 10.VIII | 25.7 | 21.5 | 25.6 | 24.0 | 26.0 | 23.3 | 26.5 | 25.8 | 26.4 | 19.4 | 26.0 | 19.4 |
| 24.VIII | 18.6 | 18.4 | 18.6 | 18.5 | 18.0 | 17.9 | 19.1 | 18.8 | 19.8 | 18.9 | 19.2 | 18.0 |
| 07.IX | 14.2 | 13.8 | 12.7 | 12.4 | 13.0 | 12.9 | 13.1 | 13.1 | 13.5 | 13.3 | 14.5 | 12.7 |
| 05.X | 9.9 | 9.5 | 9.5 | 8.8 | 9.7 | 9.6 | 10.3 | 9.7 | 10.3 | 9.6 | 10.0 | 9.2 |
| 19.X | 6.5 | 6.5 | 4.8 | 4.9 | 5.4 | 5.4 | 5.5 | 5.5 | 5.8 | 5.8 | – | – |
Примечание. П – поверхность, Д – придонный слой. Названия станций см. рис. 1.
Аномально высокие температуры воздуха и воды влияют на многие гидробиологические характеристики водохранилищ. Увеличение температуры воды удлиняет вегетационный период, увеличивает доступность питательных веществ, уменьшает количество хищников и приводит к неконтролируемому росту водорослей (Cazzolla Gatti, 2016; Mosello, Lami, 2011; Park et al., 2013). При “цветении” воды часто происходит резкое снижение концентрации растворенного кислорода, что негативно сказывается на жизнедеятельности гидробионтов, вплоть до их гибели (Adrian et al., 2009). Известно, что в неглубоких озерах и водохранилищах умеренного пояса потепление часто приводит к росту скорости процессов эвтрофирования (Копылов и др., 2012; Alimov, 2010; Cazzolla Gatti, 2016; Downing, 2014). Усиление темпов эвтрофикации вод, обусловленное не только антропогенным воздействием, но и глобальными факторами, в том числе, изменением климата в сторону потепления, приводит к росту количественных показателей донных биоценозов и повышению трофического статуса природных олиготрофных водоемов (Кашулин и др., 2012). На начальной стадии процесса эвтрофикации может происходить увеличение видового богатства (Лазарева, Соколова, 2013; Heino et al., 2009; Van Looy et al., 2016). Установлено, что при дальнейшем загрязнении или эвтрофировании водоемов структура сообществ гидробионтов упрощается: снижается разнообразие, уменьшается количество видов хищных животных и фильтраторов, доминирование переходит к эврибионтным видам (Adrian et al., 2009; Cazzolla Gatti, 2016; Hershkovitz et al., 2015; Park et al., 2013). Кроме того, вследствие влияния потепления климата на экосистемы происходит вытеснение узкоспециализированных видов видами эврибионтами (Downing, 2014), а также вселение чужеродных видов (Kernan, 2015; Mosello, Lami, 2011).
Жарким летом 2010 г. в водохранилищах Волги отмечено катастрофическое ухудшение качества вод, вызванное аномальным развитием цианобактерий (Копылов и др., 2012). В Рыбинском водохранилище температура воды у поверхности в июле и августе 2010 г. достигала 27–28°С (табл. 5), что было выше нормы (≤19–22°С), одновременно с этим наблюдалось увеличение видового богатства и обилия зоопланктона (Лазарева, Соколова, 2013). При этом, значения температуры в придонном слое были на ~1–10°С ниже, чем у поверхности воды (табл. 5). Максимальная температура придонной воды (25.8°С) зарегистрирована в начале августа 2010 г. (табл. 5). По-видимому, негативное влияние на зообентос оказало не столько повышение температуры воды, как вызванные этим изменения других факторов среды, в первую очередь, снижение до критических значений (<5 мг/л) содержания растворенного О2. Кроме того, в анаэробных условиях в результате микробиологической деструкции органического вещества грунтов могут выделяться ядовитые для беспозвоночных и рыб соединения (Дзюбан, 2006).
По наблюдениям на водоемах-охладителях Украины, незначительный подогрев ведет к увеличению числа видов и биомассы макрозообентоса, тогда как сильный вызывает уменьшение числа видов и существенное снижение численности и биомассы, причем, сильное угнетение макрозообентоса наступает уже при летней придонной температуре >25°С (Пидгайко и др., 1970; Протасов и др., 1991; Протасов, Силаева, 2012; Протасов и др. 2017). Влияние аномально высокой температуры воды на макрозообентос Рыбинского водохранилища в полной мере проявилось уже осенью 2010 г., когда видовое богатство и разнообразие снизились до минимальных значений: из состава сообществ исчезли хищники (пиявки), значительно уменьшилось количество моллюсков-фильтраторов (пизидиид и дрейссенид). На серых илах доминировали полисапробы (личинки мотыля), достигавшие ~70–99% общей биомассы (рис. 4). Наблюдаемые изменения свидетельствует о неустойчивом состоянии бентосных зооценозов, испытывающих влияние неблагоприятных факторов. Аналогичные процессы происходят в водоемах-охладителях из-за длительного повышения летней температуры воды >30°С (Пидгайко и др., 1970; Протасов и др., 1991, 2017; Яныгина, 2011). По-видимому, жарким летом 2010 г. в глубоководной зоне Рыбинского водохранилища условия для развития большинства групп макрозообентоса, за исключением личинок рода Chironomus, были неблагоприятны из-за высокой придонной температуры. А.С. Константиновым (1958) установлено, что скорость роста личинок хирономид в лабораторных условиях значительно увеличивается с повышением температуры, причем 25°С – оптимум для их развития. Дальнейшее повышение температуры ведет к угнетению роста и развития личинок хирономид. Олигохеты (кроме устойчивого к высокой температуре Limnodrilus hoffmeisteri) (Каратаев, 1990; Каратаев, Каратаева, 1987)) и моллюски, в отличие от личинок хирономид, более чувствительны к длительному (>2 нед) воздействию высокой температуры придонной воды, поэтому они первыми выпадают из состава донного населения (Пидгайко, 1971; Протасов и др., 1991). Кроме непосредственного воздействия на беспозвоночных, повышение температуры вследствие аномально жаркой погоды, наблюдавшейся в июле и августе 2010 г., привело к снижению содержания растворенного кислорода в придонных слоях воды (Копылов, 2012), что так же негативно повлияло на донное население. Многолетние наблюдения на глубоководных участках Рыбинского водохранилища показали, что из-за повышенного прогрева (>20°С) глубоких слоев воды с 2007 г. стал чаще встречаться летний (июль–август) придонный дефицит растворенного О2. В 2011–2015 гг. регулярно наблюдалось его низкое содержание у дна (<5 мг/л) длительностью от двух недель до трех месяцев (Лазарева и др., 2018). Одним из последствий повышения температуры воды и обусловленного этим дефицита растворенного О2 было значительное снижение обилия чувствительных к недостатку содержания кислорода видов β-мезосапробов. Так, в 2010–2011 гг. на стандартных станциях почти не встречались моллюски-дрейссениды, редко и единично отмечена олигохета Tubifex newaensis. Личинки Chironomus plumosus (L.) имеют преимущество перед другими видами донного населения, поскольку за счет высокой концентрации в их гемолимфе дыхательного пигмента гемоглобина могут обитать в среде, бедной кислородом, а также и в анаэробных условиях благодаря сложным приспособительным механизмам, в частности за счет повышения содержания в теле гликогена (Мотыль ..., 1983).
В северных водоемах эвтрофирование не всегда приводит к деградации водных экосистем, особенно при умеренном проявлении таких процессов (Яковлев, 2005). На всех этапах сукцессии (от олиготрофного до эвтрофного статуса) водная экосистема способна адаптироваться, изменяя свою структурно-функциональную организацию и перестраивая механизмы утилизации органического вещества (Яковлев, 2005). В Рыбинском водохранилище в последующие годы после аномально жаркого лета 2010 г. на всех станциях наблюдалось увеличение видового богатства зообентоса и индекса видового разнообразия Шеннона–Уивера, максимальные значения этих показателей зарегистрированы в 2012–2013 гг. (табл. 1). На большинстве исследованных участков увеличилось число видов олигохет (в основном полисапробов) и их доля в общей численности макрозообентоса (рис. 2, рис. 3). Это свидетельствует об избыточном содержании органического вещества в грунтах, а также интенсивных процессах самоочищения с активным участием олигохет. Сравнительный анализ характеристик макрозообентоса на шести стандартных станциях Рыбинского водохранилища осенью 2009 и 2013–2015 гг. показал, что на серых илах (ст. 1, 2, 5, 6) произошел существенный рост численности и биомассы (табл. 2). На ст. 3, 4, где отсутствует интенсивное илонакопление, показатели макрозообентоса изменились незначительно. Ст. 1, 2, 6 всегда отличались от остальных участков высоким обилием макрозообентоса (Поддубная, 1988; Щербина, 2000; Perova, 2012). По данным за 1994 г., средняя за сезон биомасса макрозообентоса на ст. 1, 2, 6 была соответственно 24.98, 33.01, 18.91 г/м2; отмечалось, что значения биомассы возросли в 3–13 раз по сравнению с 1955 г. (Щербина, 2000). В осенний сезон 2011–2015 гг. на этих станциях произошел еще более существенный рост биомассы макрозообентоса, ее средние значения увеличились в ~3–7 раз по сравнению с отмеченными в 1994 г. (табл. 2). Наибольшие значения биомассы зарегистрированы в сентябре 2015 г. на ст. 2 (205 г/м2) и ст. 6 (173 г/м2), основу биомассы (73–76%) составляли личинки мотыля (в основном Chironomus plumosus). Известно (Мотыль ..., 1983), что повышение трофии внутренних водоемов способствует процветанию и расселению этого вида с широкой экологической валентностью. Следует отметить, что такое высокое обилие макрозообентоса на стандартных станциях выявлено впервые за весь период наблюдений, причем стремительный рост биомассы произошел за относительно короткий временной промежуток – с 2011 г. по 2015 г., после экстремально теплых лет. В Рыбинском водохранилище такие высокие значения биомассы макрозообентоса отмечали лишь однажды – весной 1993 г. на приплотинных участках, расположенных на бывшем русле р. Шексны (143–200 г/м2) (Щербина, 2000).
Значительный рост численности и биомассы макрозообентоса в 2011–2015 гг. свидетельствует об изменении трофического статуса Рыбинского водохранилища. В первые годы после образования водохранилища его воды характеризовались как “чистые” олиго-мезотрофного уровня продуктивности при отсутствии сильного антропогенного воздействия (Рыбинское ..., 1972). Рост средних температур воздуха и воды привел к повышению интенсивности процессов фотосинтеза, увеличению первичной продукции и накоплению органического вещества, способствующему бурному развитию микроорганизмов (Романенко, 1985) и повлиял на всю экосистему водохранилища.
В структуре макрозообентоса Рыбинского водохранилища преобладают трофические группировки детритофагов-глотателей (грунтоядных олигохет) и фитодетритофагов-фильтраторов + собирателей (личинок хирономид), которые питаются органическими частицами в верхнем слое донных отложений (Монаков, 1998). Соответственно, количественное обилие донного населения в значительной степени зависит от содержания органического вещества в грунтах, одним из показателей которого служит концентрация осадочных пигментов (суммы хлорофилла а с феопигментами) (Сигарева и др., 2013). Осадочные пигменты могут быть индикатором связи осадкообразования и первичной продукции (Сигарева и др., 2013). С помощью корреляционного анализа и метода главных компонент выявлена достоверная положительная связь между биомассой макрозообентоса и концентрацией осадочных пигментов в донных отложениях (Тимофеева и др., 2018). Установлено, что в аномально жаркие годы, каким был 2010 г., на глубоководных участках, где обычно происходит седиментация основной части органического вещества и формируются илы, содержание осадочных пигментов увеличивается. Более высокое содержание пигментов в следующий за аномально жарким годом дает основание говорить об эффекте последействия на состояние донной подсистемы и экосистемы в целом (Тимофеева, Сигарева, 2012). Так, в 2011 г. по концентрации хлорофилла с дериватами в донных отложениях состояние донной подсистемы Рыбинского водохранилища оценивалось как высокоэвтрофное (Сигарева и др., 2012). Средние величины содержания пигментов в грунтах Рыбинского водохранилища, как и в водной толще, в большинстве случаев соответствуют мезо- и эвтрофным водоемам (Тимофеева, Сигарева, 2012). На станциях, расположенных на участках накопления илов, среднегодовые концентрации пигментов в 2011–2014 гг. были выше, чем в прежние годы (Сигарева и др., 2016). В исследуемый период высокие концентрации пигментов, как и высокая численность бактериобентоса (Kopylov et al., 2016), создали хорошие трофические условия для массового развития фитодетритофагов-фильтраторов + собирателей – личинок хирономид р. Chironomus и детритофагов-глотателей – олигохет. В результате доминирования этих групп на участках, занятых серыми илами, наблюдался интенсивный рост обилия макрозообентоса в 2011–2015 гг. В соответствии с классификацией “шкалы трофности” для озер и водохранилищ (Китаев, 2007) по биомассе зообентоса осенью 2015 г. ст. 4 относилась к α-олиготрофному классу (<1.25 г/м2), ст. 3 – к β-олиготрофному (1.25–2.5), ст. 5 – к α-евтрофному (10–20 г/м2), ст. 1 – к β-евтрофному (20–40 г/м2), ст. 2 и 6, где биомасса достигала >40 г/м2 – к α-политрофному, тогда как в 2009 г. самые продуктивные станции, расположенные на серых илах, характеризовались как α-евтрофные (табл. 2). Таким образом, по показателям макрозообентоса отмечено увеличение темпов эвтрофирования Рыбинского водохранилища в экстремально теплые годы начала ХХI в.
Выводы. Изменения структуры макрозообентоса Рыбинского водохранилища, отмеченные в начале ХХI в., совпали с годами наиболее интенсивного потепления. Повышение температуры воды аномально жарким летом 2010 г. и вызванные этим неблагоприятные условия среды привели к значительному снижению видового богатства и разнообразия, угнетению почти всех представителей макрозообентоса, кроме личинок хирономид рода Chironomus, составлявших 70–99% общей численности и биомассы. В последующие годы наблюдалось восстановление сообществ донного населения, отмечен существенный рост видового богатства на всех исследованных участках. При этом, в структуре макрозообентоса уменьшилась роль видов-индикаторов β-мезосапробных условий (моллюсков-дрейссенид). Значительно возросла численность олигохет, среди которых произошла смена доминирующих видов – в сообществах донного населения преобладали полисапробы Limnodrilus hoffmeisteri и Tubifex tubifex, тогда как β-мезосапроб Tubifex newaensis, доминировавший в конце ХХ в., стал редким видом. Интенсивный рост численности и биомассы мотыля и полисапробных видов олигохет, зарегистрированный в глубоководной зоне Рыбинского водохранилища осенью 2011–2015 г., свидетельствует об увеличении содержания органического вещества в донных отложениях и изменении трофического статуса Рыбинского водохранилища.
Список литературы
Архипова Н.Р. 2016. Структурная организация сообщества олигохет (Annelida: Oligochaeta) на продуктивных серых илах Рыбинского водохранилища // Тр. ИБВВ РАН. Вып. 74(77). Ярославль: Филигрань, С. 100–112.
Дзюбан А.Н. 2006. Микробиологическая характеристика донных отложений Верхней Волги в условиях антропогенного воздействия // Биология внутр. вод. № 1. С. 16–23.
Доклад об особенностях климата на территории Российской Федерации за 2009 г. [Электронный ресурс]. М.: Росгидромет, 2010. http://www.meteo.ru, file.pdf (обращение 4 апреля 2012 г.).
Доклад об особенностях климата на территории Российской Федерации за 2010 г. [Электронный ресурс]. М.: Росгидромет, 2011. http://www.meteo.ru, file.pdf (обращение 4 апреля 2012 г.).
Доклад об особенностях климата на территории Российской Федерации за 2011 г. [Электронный ресурс]. М.: Росгидромет, 2012. http://www.meteo.ru, file.pdf (обращение 04. 04.2012 г.).
Доклад об особенностях климата на территории Российской Федерации за 2012 год. М.: Росгидромет, 2013. 86 с.
Доклад об особенностях климата на территории Российской Федерации за 2013 год. М.: Росгидромет, 2014. 109 с.
Доклад об особенностях климата на территории Российской Федерации за 2014 год. М.: Росгидромет, 2015. 107 с.
Доклад об особенностях климата на территории Российской Федерации за 2015 год. М.: Росгидромет, 2016. 68 с.
Каратаев А.Ю. 1990. Воздействие подогрева на пресноводные экосистемы // Вестн. Белорус. ун-та. Минск, 132 с. Деп. в ВИНИТИ 07.05.1990, № 2440–90.
Каратаев А. Ю., Каратаева И. В. 1987. Влияние сброса теплых вод на макрозообентос литорали водоема-охладителя // Вестн. Белорус. ун-та. Сер. 2. № 1. С. 46–49.
Кашулин Н.А., Денисов Д.Б., Валькова С.А. и др. 2012. Современные тенденции изменений пресноводных экосистем Евро-Арктического региона // Тр. Кольск. науч. центра РАН. № 2. С. 7–54.
Китаев С.П. 2007. Основы лимнологии для гидробиологов и ихтиологов // Петрозаводск: Карельск. науч. центр РАН, 395 с.
Климатические рекорды прошедшего пятилетия на территории Ярославской области (Электронный ресурс) // Сайт ГУ “Ярославский ЦГМС”. Режим доступа: http://www.yacgms.ru – 2006–2010. Дата обращения: 28.04.2012.
Константинов А.С. 1958. Биология хирономид и их разведение // Тр. Саратов. отд. ВНИОРХ. Т. 5. Саратов: Изд-во Коммунист, 363 с.
Копылов А.И., Лазарева В.И., Минеева Н.М. и др. 2012. Влияние аномально высокой температуры воды на развитие планктонного сообщества водохранилищ Средней Волги летом 2010 г. // Докл. РАН. Т. 442. № 1. С. 133–135.
Лазарева В.И., Соколова Е.А. 2013. Динамика и фенология зоопланктона крупного равнинного водохранилища: отклик на изменение климата // Успехи соврем. биологии. Т. 133. № 6. С. 564–574.
Лазарева В.И., Степанова И.Э., Цветков А.И. и др. 2018. Кислородный режим водохранилищ Волги и Камы в период потепления климата: последствия для зоопланктона и зообентоса // Тр. Ин-та биологии внутр. вод РАН. № 81(84). С. 47–84.
Литвинов А.С., Законнова А.В. 2014. Экологические условия в Рыбинском водохранилище при потеплении климата // Географ. вестн. Физическая география и геоморфология. № 2(29). С. 41–45.
Методика изучения биогеоценозов внутренних водоемов. М.: Наука, 1975. 240 с.
Монаков А.В. 1998. Питание пресноводных беспозвоночных. М.: Ин-т проблем экологии и эволюции РАН, 322 с.
Мотыль Chironomus plumosus L. (Diptera, Chironomidae). Cистематика, морфология, экология, продукция. М.: Наука, 1983. 312 с.
Пидгайко М.Л. 1971. Зоомакробентос Кураховского водохранилища и влияние на него подогрева // Гидрохимия и гидробиология водоемов-охладителей тепловых электростанций СССР. Киев: Наук. думка, С. 207–216.
Пидгайко М.Л., Гринь В.Г., Поливанная М.Ф. и др. 1970. Итоги изучения гидробиологического режима пресных водоемов-охладителей юга УССР // Гидробиол. журн. Т. 6. № 2. С. 36–44.
Поддубная Т.Л. 1988. Многолетняя динамика структуры и продуктивность донных сообществ Рыбинского водохранилища // Структура и функционирование пресноводных экосистем. Л.: Наука, Вып. 55(58). С. 112–140.
Протасов А.А., Сергеева О.А., Кошелева С.И. и др. 1991. Гидробиология водоемов-охладителей тепловых и атомных электростанций Украины. Киев: Наук. думка, 192 с.
Протасов А.А., Силаева А.А. 2012. Контурные группировки гидробионтов в техно-экосистемах ТЭС и АЭС. Киев: Ин-т гидробиологии НАН Украины, 274 с.
Протасов А.А., Силаева А.А., Новоселова Т.Н. и др. 2017. Техноэкосистема АЭС: 18 лет гидробиологических наблюдений // Журн. Сиб. фед. ун-та. Серия: Биология. Т. 10. № 4. P. 459–484.
Романенко В.И. 1985. Микробиологические процессы продукции и деструкции органического вещества во внутренних водоемах. Л.: Наука, 295 с.
Рыбинское водохранилище и его жизнь. Л.: Наука, 1972. 364 с.
Сигарева Л.Е., Тимофеева Н.А., Законнов В.В. 2012. Трофическое состояние донной подсистемы в Волжских водохранилищах // Бассейн Волги в ХХI-м веке: структура и функционирование экосистем водохранилищ: Матер. докл. Всерос. конф. Борок, 2012. Ижевск: Издатель Пермяков С.А. С. 261–264.
Сигарева Л.Е., Законнов В.В., Тимофеева Н.А., Касьянова В.В. 2013. Осадочные пигменты и скорость илонакопления как показатели трофического состояния Рыбинского водохранилища // Вод. ресурсы. Т. 40. № 1. С. 62–69.
Сигарева Л.Е., Пырина И.Л., Тимофеева Н.А. 2016. Межгодовая динамика хлорофилла в планктоне и донных отложениях Рыбинского водохранилища // Тр. Ин-та биологии внутр. вод РАН. № 76(79). С. 119–130.
Техно-экосистема АЭС. Гидробиология, абиотические факторы, экологические оценки. Киев: Ин-т гидробиологии НАН Украины, 2011. 234 с.
Тимофеева Н.А., Сигарева Л.Е. 2012. Влияние аномальной жары на содержание растительных пигментов в донных отложениях водохранилища // Бассейн Волги в ХХI-м веке: структура и функционирование экосистем водохранилищ: Матер. докл. Всерос. конф. Борок, С. 304–306.
Тимофеева Н.А., Перова С.Н., Сигарева Л.Е. 2018. Распределение осадочных пигментов и макрозообентоса в глубоководной зоне Рыбинского водохранилища // Сиб. экол. журн. № 6. С. 766–775.
Щербина Г.Х. 2000. Макрозообентос // Современная экологическая ситуация в Рыбинском и Горьковском водохранилищах: состояние биологических сообществ и перспективы рыборазведения. Ярославль: Изд-во Ярослав. гос. техн. ун-та, С. 216–231.
Яковлев В.А. 2005. Пресноводный зообентос Северной Фенноскандии (разнообразие, структура и антропогенная динамика). Апатиты: Изд-во Кольск. науч. центра PАН, Ч. 1. 161 с. Ч. 2. 145 с.
Яныгина Л.В. 2011. Экология сообществ донных беспозвоночных в водоемах-охладителях Сибири // Вод. ресурсы. Т. 38. № 5. С. 618–630.
Alimov A.F. 2010. Biological Diversity and the Community Structure of Organisms // Inland Water Biol. V. 3. № 3. P. 207–213. https://doi.org/10.1134/S1995082910030016
Adrian R., O’Reilly C.M., Zagarese H. et al. 2009. Lakes as sentinels of climate change // Limnol., Oceanogr. V. 54. № 6 (part 2). P. 2283–2297.
Cazzolla Gatti R. 2016. Freshwater biodiversity: a review of local and global threats // Int. J. Environ. Stud. V. 73. № 6. P. 887–904.
De Knijf G., Flenker U., Vanappelghem C. et al. 2011. The status of two boreo-alpine species, Somatochlora alpestris and S. arctica, in Romania and their vulnerability to the impact of climate change (Odonata: Corduliidae) // Int. J. Odonatol. V. 14. № 2. P. 111–126.
Downing J.A. 2014. Productivity of Freshwater Ecosystems and Climate Change // Global Environmental Change. Series Handbook of Global Environmental Pollution. V. 1. P. 221–229.
Heino J., Virkkala R., Toivonen H. 2009. Climate change and freshwater biodiversity: detected patterns, future trends and adaptations in northern regions // Biol. Rev. V. 84. P. 39–54.
Hershkovitz Y., Dahm V., Lorenz A.W., Hering D. 2015. A multi-trait approach for the identification and protection of European freshwater species that are potentially vulnerable to the impacts of climate change // Ecol. Indicators. V. 50. P. 150–160.
Kernan M. 2015. Climate change and the impact of invasive species on aquatic ecosystems // Aquat. Ecosyst. Health & Manag. V. 18. № 3. P. 321–333.
Kopylov A.I., Kosolapov D.B., Zabotkina E.A., Rumyantseva E.V. 2016. Viruses in Bottom Sediments of a Mesotrophic Reservoir (Rybinsk Reservoir, Upper Volga) // Inland Water Biol. V. 9. № 3. P. 251–257. https://doi.org/10.1134/S199508291603010X
Mosello R., Lami A. 2011. Climate Change and Related Effects on Water Quality: Examples from Lake Maggiore (Italy) // Global bioethics. V. 24. № 1–4. P. 95–98.
Park H.K., Cho K.H., Won D.H. et al. 2013. Ecosystem responses to climate change in a large on-river reservoir, Lake Paldang, Korea // Climat. Change. V. 120. № 1. P. 477–489.
Perova S.N. 2012. Taxonomic composition and abundance of macrozoobenthos in the Rybinsk Reservoir at the beginning of the 21st century // Inland Water Biol. V. 5. № 2. P. 199–207. https://doi.org/10.1134/S1995082912020125
Van Looy K., Floury M., Ferréol M. et al. 2016. Long-term changes in temperate stream invertebrate communities reveal a synchronous trophic amplification at the turn of the millennium // Science of the Total Environ. V. 565. P. 481–488.
Yasarer L.M.W., Sturm B.S.M. 2016. Potential impacts of climate change on reservoir services and management approaches // Lake and Reservoir Management. V. 32. № 1. P. 13–26.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод