

Биология внутренних вод, 2019, № 4-2, стр. 90-97
Модельная оценка биомассы фитопланктона оз. Иссык-Куль по данным дистанционного зондирования
А. И. Абакумов a, *, С. Я. Пак a, М. А. Морозов a, А. К. Тыныбеков b
a Институт автоматики и процессов управления, Дальневосточное отделение Российской академии наук
Владивосток, Россия
b Кыргызский государственный университет им. Арабаева
Бишкек, Кыргызстан
* E-mail: packsa@iacp.dvo.ru
Поступила в редакцию 03.04.2018
После доработки 24.05.2018
Принята к публикации 01.08.2018
Аннотация
Выполнена модельная оценка биомассы фитопланктона фотического слоя центральной части оз. Иссык-Куль. Метод основан на данных дистанционного зондирования для численного решения математической модели функционирования фитопланктона в неподвижном столбе воды. Для верификации параметров модели использованы натурные пробы, собранные в пелагиали озера (разрез Тамга–Григорьевка) в августе 1999 г.
ВВЕДЕНИЕ
Сохранение жизнеспособности водных экосистем составляет одну из главных задач экологического мониторинга на протяжении второй половины прошлого и начала текущего столетия. Основой трофической пирамиды большинства водных экосистем служит фитопланктон, создающий в процессе фотосинтеза питательный ресурс для всей фауны водоема. В условиях антропогенных угроз растительные организмы способны активировать механизмы самоочищения, благодаря процессам фотосинтеза и гетеротрофного усвоения растворенных органических соединений. Микроводоросли, обладая высокой фотосинтетической активностью, утилизируют около трети атмосферного и примерно половину растворенного в воде углекислого газа (Hoppenrath et al., 2009).
При исследовании водных экосистем возникает необходимость оценивать продукционный потенциал как отдельных участков, так и больших акваторий. Последнее стало возможным благодаря методам дистанционного зондирования, включающим использование спутниковых спектрорадиометров (Чурилова и др., 2016; Aleksanin et al., 2012) и позволяющим получать данные о распределении фитопланктона на основе диагностики основного растительного пигмента Хл а. Принципиальное ограничение дистанционных методов – для обработки спутниковым сигналом доступен только поверхностный слой. Традиционные же контактные методы получения данных для толщи воды (Маторин и др., 2002; Kulchin et al., 2008) носят преимущественно локальный характер. Пути преодоления факторов, лимитирующих использование природного мониторинга, лежат в разработке математических моделей функционирования фитопланктона (Abakumov, Izrailsky, 2012; Abakumov et al., 2015; Allen, Polimene, 2011; Grangere et al., 2009; Nikolaou, Hartmann et al., 2016; Platt et al., 1991). Посредством конструирования и верификации моделей на основе дистанционных и натурных данных становится возможной оценка биопотенциала заданного географического объекта (Menshutkin et al., 2014).
Цель работы – использование методов математического моделирования в совокупности с натурными измерениями и дистанционным наблюдением для оценки биомассы фитопланктона центральной части оз. Иссык-Куль.
Озеро Иссык-Куль – объект научного и практического интереса (Кулумбаева, 1975; Маторин и др., 2002; Тыныбеков, 2016), представляет собой крупный незамерзающий слабосоленый (общая минерализация воды ~5.97‰) глубоководный (максимальная глубина 668 м, средняя глубина 278.4 м) водоем площадью 6200 км2, который занимает 23 место по величине среди озер мира (Атлас …, 1987). Для озера характерны цикличные изменения уровня воды с периодом, кратным десятилетиям. Это приводит к значительным затруднениям в оценке водного баланса водоема. Расхождения краткосрочных показателей запасов воды в озере со среднемноголетними могут доходить до 67% (Черкасов, 1977). На протяжении многих лет озеро привлекает огромное число туристов и, несмотря на постоянно действующий комплекс природоохранных мероприятий, подвергается антропогенному воздействию.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Предпринята попытка применить для оценки биомассы фитопланктона в толще воды оз. Иссык-Куль вертикальную модель, построенную на концепции функции приспособленности (Пак, Абакумов, 2014; Полуэктов, Пых и др., 1980; Фрисман, Жданова, 2009) и верифицированную по натурным пробам. В качестве начальных условий использованы данные дистанционного зондирования. Результаты, полученные нами ранее (Пак, Абакумов, 2014; Абакумов, Пак, 2016), основаны на разработке и использовании модели, которая имитирует динамику биомассы фитопланктона в неподвижном вертикальном столбе воды. Допущения, принятые на этапе построения модели, подробно изложены в этих работах. В отличие от большинства систем подобного рода данная модель основана не на балансовых соотношениях, а на оптимизации функции приспособленности, а также на гипотезе об обратной связи биомассы фитопланктона с концентрацией биогенных элементов. Поскольку модель стационарная, в уравнениях учитываются только изменения, зависящие от распределения водорослей по глубине.
Общий вид модели следующий:
(1)
$\left. \begin{gathered} \frac{{dy}}{{dx}} = \left[ {\frac{{d{\mu }}}{{dx}} - e\left( y \right)} \right]y, \hfill \\ \frac{{dz}}{{dx}} = \nu p\left( {y,{{y}_{0}}} \right)z, \hfill \\ \frac{{dI}}{{dx}} = - k\left( {y,z} \right)I, \hfill \\ \end{gathered} \right\},$При моделировании удельной скорости роста использован подход, предполагающий мультипликативную зависимость от основных благоприятствующих факторов: обеспеченности минеральным питанием, ФАР, температуры. Последняя играет регулирующую роль в репродукции фитопланктона и ее учет необходим при построении функции приспособленности в уравнениях (1):
Влияние концентрации биогенных веществ на скорость роста фитопланктона является позитивным и традиционно описывается кривой Моно (Monod, 1949):
где z0 – константа полунасыщения.Аналогичным образом принято описывать зависимость скорости продуцирования от освещенности (Динамические модели ..., 2001–2019):
Здесь I0 – константа полунасыщения по освещенности.Нами выбран именно такой способ представления светового влияния, хотя он не единственно возможный.
При моделировании температурного влияния на скорость роста фитопланктона была применена концепция интервала толерантности, согласно которой соответствующий коэффициент представлен в экспоненциальной форме:
Перейдем ко второму уравнению системы (1). Если придерживаться избранной нами гипотезы об обратной связи фитопланктона с ресурсом биогенов z, то функцию p можно выбрать в виде зависимости Михаэлисa–Ментен:
где y0 – константа полунасыщения по фитопланктону.Третье уравнение системы имитирует ослабление освещенности с глубиной, обусловленное поглощением водной толщей, затенением фитопланктоном и взвешенным органическим веществом. Соответствующее распределение описывается функцией:
где k0 – коэффициент общей мутности воды, k1 – коэффициент затенения фитопланктоном, k2 – коэффициент затенения минеральными веществами.Модель базируется на трактовке воспроизводства фитопланктона, при которой минеральное питание заменено понятием “пищевой ресурс”. Более подробно тактика построения системы (1) изложена в работе (Абакумов, Пак, 2016).
Информацией, использованной в качестве начальных условий для решения системы уравнений (1), послужили данные спутниковых наблюдений для района с координатами 42°–43° с.ш., 77°–78° в.д., полученные в июле и августе 2002 г. Спутник, оснащенный спектрорадиометром Aqua, с помощью которого получены данные, размещенные на сайте NASA, выведен на околоземную орбиту в мае 2002 г. (рис. 1). Данные предоставлены Центром коллективного пользования регионального спутникового мониторинга окружающей среды Института автоматики и процессов управления ДВО РАН (Владивосток). Программное обеспечение, которым располагает Центр, позволило сформировать числовые проекции данных по концентрации Хл а в поверхностном водном слое, освещенности водной поверхности и температуре, построенных с помощью спутниковой обработки.

В нашем распоряжении были материалы, полученные при проведении экспедиционных работ на оз. Иссык-Куль (Маторин и др., 2002; Тыныбеков, Маторин, 2009) в августе 1999 г. в пелагиали разреза Тамга–Григорьевка (рис. 2). Данные по вертикальному распределению биомассы фитопланктона, освещенности и температуры воды использованы для уточнения параметров, входящих в уравнения системы (1) в качестве постоянных величин. Согласно работе Тыныбекова (2016), вертикальное распределение температуры в толще озера в течение нескольких лет демонстрирует высокую степень единообразия, что дает основание предположить малую изменчивость абиотических характеристик и считать их среднемноголетнее распределение в столбе воды квазипостоянным. Таким образом, верификацию модели (1) на основе подспутниковых измерений 1999 г., как и последующее ее решение по исходной спутниковой информации 2002 г., можно считать вполне правомочной, учитывая факт, что все имеющиеся данные относятся к одному и тому же вегетационному периоду.
Рис. 2.
Вертикальное распределение температуры (а), подводной освещенности (б) и биомассы фитопланктона (в) в пелагиали оз. Иссык-Куль.
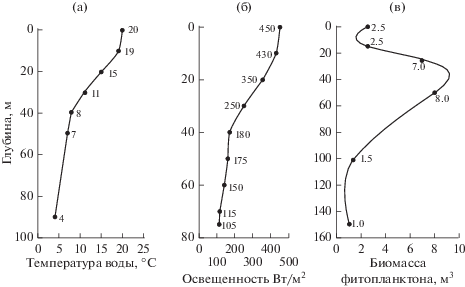
Для анализа распределения биогенных соединений (нитритов, нитратов и фосфатов) по глубине использованы данные, собранные в июле 1998 г. (табл. 1).
Таблица 1.
Распределение биогенных элементов (мг/дм3) по глубине оз. Иссык-Куль (соединения NO2, NO3, PO4)
| Глубина, м | 0.5 | 10 | 25 | 50 | 100 | 200 | 300 | 400 | 500 | 650 |
|---|---|---|---|---|---|---|---|---|---|---|
| Концентрация | 0.10 | 0.14 | 0.11 | 0.10 | 0.11 | 0.13 | 0.14 | 0.15 | 0.20 | 0.24 |
Построенные кривые вертикального распределения биомассы фитопланктона, температуры воды, освещенности и концентраций биогенов переведены в единицы измерения, которые соответствуют числовым проекциям спутниковых наблюдений, а затем аппроксимированы методом сплайн-интерполяции с целью последующей минимизации функционала невязки.
Минимизируется по параметрам µ0, z0, I0, θopt, τ среднеквадратичное отклонение левой и правой части первого уравнения системы (1) при подстановке в него интерполированных на основе экспериментальных наблюдений данных
(2)
${{\left[ {\mathop \smallint \limits_0^{75} {{{\left( {\frac{{dy}}{{dx}} - \left[ {\frac{{d{\mu }}}{{dx}} - e\left( y \right)} \right]y} \right)}}^{2}}dt} \right]}^{{1/2}}}.$Пределы интегрирования выбраны в соответствии с теми горизонтами, на которых известны опорные значения всех построенных сплайнов. В частности, сплайн-функция, аппроксимирующая вертикальное распределение освещенности, ограничена глубиной 75 м, которая и определяет верхний предел интегрирования. В качестве начального приближения выбран вектор b0 = (µ0, z0, I0, θopt, τ) с компонентами, представленными в табл. 2. Их значения частично взяты из открытых источников (Ryabov et al., 2010; Tanaka, Mano, 2012), работ (Абакумов, Пак, 2016; Abakumov, Izrailsky, 2012), а также составлены на основе анализа натурных данных, представленных в работе (Маторин и др., 2002). В результате минимизации пространственной нормы (2) вектор значений (µ0, z0, I0, θopt, τ) соответственно равняется (2.0, 0.55, 39.5052, 9.5, 11.5).
Таблица 2.
Численные значения основных параметров модели
| Параметр | Обозначения | Численные значения |
|---|---|---|
| Максимальная скорость роста фитопланктона | μ0 | 0.5 |
| Константа полунасыщения, г/м3: | ||
| для хлорофилла для минеральных веществ |
y0 z0 |
3.1 4.0 |
| Максимальная удельная скорость роста концентрации минеральных веществ, сут–1 | v | 0.03 |
| Удельная скорость элиминации фитопланктона, сут–1 | e0 | 0.023 |
| Константа полунасыщения для освещенности, Eм–2 сут–1 | I0 | 60 |
| Удельная скорость затухания освещенности, м–1 | k0 | 0.045 |
| в том числе, г–1 м2: | ||
| за счет хлорофилла за счет минеральных веществ |
k1 k2 |
0.018 0.010 |
| Оптимальная температура, °С | θopt | 8.0 |
| Интервал толерантности по температуре, °С | τ | 10.0 |
Полученные значения использованы в качестве параметров первого уравнения модели (1). Параметры прочих уравнений взяты из вышеназванных литературных источников (Ryabov et al., 2010; Tanaka, Mano, 2012), а не посредством минимизации интегральной нормы наподобие (2) по следующим соображениям. Распределение биомассы фитопланктона с максимумом в подповерхностном слое и снижением в толще воды ниже термоклина (рис. 2в) соответствует описанному в работах (Кукси, 1964; Свирежев, Логофет, 1978). Вертикальные профили распределения биогенных элементов (табл. 1) и освещенности (рис. 2б), как следует из работ (Маторин и др., 2002; Тыныбеков, Маторин, 2009), носят локальный характер. В открытых источниках нам не удалось найти описания распределения минеральных веществ по глубине и вертикального хода освещенности. Поэтому при численной реализации модели (1) не стали прибегать к верификации параметров по имеющимся в распоряжении данным, а воспользовались опубликованными в тематической литературе (табл. 2).
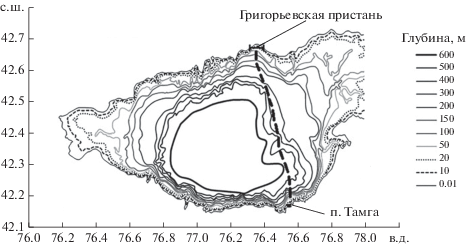
Выбранный географический квадрат включал разрез Тамга–Григорьевка, где получены контактные пробы (рис. 2). На основе топографических материалов была составлена карта глубин озера (рис. 3), построенная путем оцифровки растрового изображения топографической карты масштаба 1 : 500 000. Географическую привязку и оцифровку осуществляли при помощи программы Vextractor (www.vextrasoft.com) в полуавтоматическом режиме. Исходное растровое изображение озера преобразовывали в векторный вид (векторизовано), после чего вручную был проведен отбор и маркировка изобат с одновременной правкой и коррекцией дефектов, неизбежно возникающих в процессе автоматической векторизации. Построенные изолинии сохранены в текстовом формате, пригодном для импорта в программу Surfer 14, а затем преобразованы в регулярную сетку методом триангуляции с линейной интерполяцией. Узлы сетки задавали таким образом, чтобы совместить значения глубин с координатами точек, в которых получены данные дистанционного зондирования по концентрации Хл а, поверхностной температуре воды и освещенности верхнего слоя.
Рис. 3.
Карта глубин, включающая разрез Тамга-Григорьевка оз. Иссык-Куль. Пунктиром показан приблизительный маршрут промеров.
Биомассу фитопланктона можно оценивать по концентрации Хл а (Динамические модели в биологии), содержание которого в единице биомассы микроводорослей колеблется в некоторых пределах (Сиделев, Бабаназарова, 2008). Данные о концентрации пигмента на разных глубинах в средней части озера (Маторин и др., 2002) позволяют предположить, что в данном случае хлорофилльная квота варьирует от 0.01 до 0.06. Считая, что концентрация Хл а, зарегистрированная спутниковым спектрорадиометром, соответствует случайной величине, равномерно распределенной в промежутке от 0.01 до 0.06, получили биомассу фитопланктона на единицу поверхностной площади. Глубина фотического слоя, ограниченного поступлением 1% падающей радиации, принята от x1 = 0 до дна xb, но ≤200 м. Концентрация биогенных веществ в поверхностном слое выбрана по материалам работы (Тыныбеков, Маторин, 2009). Распределение температуры по глубине аппроксимировано кусочно-линейной функцией вида (3), имитирующей изменение температуры в пелагиали и на глубине (рис. 1а):
(3)
$\theta \left( x \right) = \left\{ {\begin{array}{*{20}{c}} {{{\theta }_{1}} + \left( {{{\theta }_{2}} - {{\theta }_{1}}} \right)\frac{{x - {{x}_{1}}}}{{{{x}_{2}} - {{x}_{1}}}},\,\,\,\,{{x}_{1}} \leqslant x \leqslant {{x}_{2}}} \\ {{{\theta }_{2}} + \left( {{{\theta }_{b}} - {{\theta }_{2}}} \right)\frac{{x - {{x}_{2}}}}{{{{x}_{b}} - {{x}_{2}}}},\,\,\,\,{{x}_{2}} \leqslant x \leqslant {{x}_{b}}} \end{array}} \right..$Построение распределения биомассы фитопланктона в заданном географическом квадрате состояло в численном решении задачи Коши для вертикальной модели (1) в каждой информативной точке регулярной сетки с шагом 0.0033 градуса или 11.88 угловых секунд между ними по долготе (условно по горизонтали на рис. 1) и 0.002448 по широте (условно по вертикали). Под информативностью подразумевается наличие в текущем узле корректной спутниковой информации, получению которой может препятствовать облачность. В последнем случае точка исключалась из расчетов. Интегрирование проводили согласно вышеописанной карте глубин. В точках, где глубина достигает 300–500 м, ограничились 200-метровой границей, которая, согласно предварительным расчетам, характеризует максимальную толщину фотического слоя. Значения биомассы в точках, где данные не удалось зафиксировать из-за неблагоприятных условий распространения спутникового сигнала, получены путем автоматической триангуляции по тем опорным точкам, где корректные данные зарегистрированы успешно.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
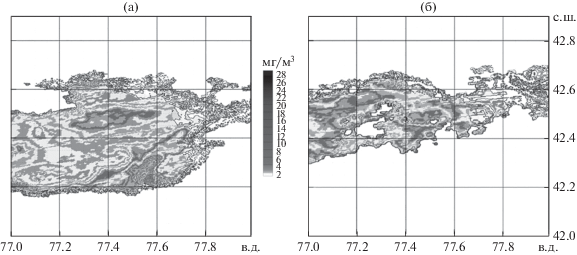
Описанным методом удалось сформировать распределение биомассы фитопланктона в толще воды исследуемого района оз. Иссык-Куль в июле, августе 2002 г. (рис. 4). Максимальные величины интегральной биомассы фитопланктона в этот период достигают 24–28 мг/м3, что несколько расходится с данными, приведенными в (Атлас …, 1987; Кулумбаева, 1975), но вполне согласуется с опубликованными в (Маторин и др., 2002). Это можно объяснить как изменениями методов сбора данных, так и снижением биологической продуктивности озера.
Рис. 4.
Распределение биомассы фитопланктона в водной толще оз. Иссык-Куль 30 июля (а) и 26 августа (б) 2002 г.
Аналитическое описание внешних функций модели (1), способы верификации основных параметров зависят от объема исходных данных, которыми располагает исследователь, и качества их описания. В частности, в работе Пак, Абакумова, (2014) моделирование температурного режима, его сезонных изменений и прохождения холодных течений было выполнено на основе биогеографического описания заданного объекта. В данном случае имелся доступ к данным, распределенным по глубине и полученным непосредственно в экспедиционных условиях. Это позволило уточнить значения основных параметров, но при этом сузить область, доступную для спутниковой обработки, во избежание больших погрешностей аппроксимации.
Предложенный метод исследования позволяет рассчитать оценочные биотические характеристики больших по площади водных объектов с заданным промежутком времени. Так, данные по биомассе фитопланктона получены нами не только для поверхностного слоя, но и для толщи воды. С решением такой проблемы неразрывно связаны алгоритмы уточнения спутниковых измерений (Левин и др., 2010) и данных, полученных контактными способами (Маторин, Рубин, 2012). Визуализация результатов численного решения вертикальной модели дает представление о пространственном распределении удельных концентраций совокупной биомассы фитопланктона, что, в свою очередь, делает возможным анализ формирования ее объема в зависимости от внешних факторов, включая климат, антропогенное воздействие, донный ландшафт и прочее. Доступность спутниковой информации позволяет сформировать массивы данных таким образом, чтобы можно было сконструировать динамический режим изменения показателей состояния фитопланктона. Динамические модели – инструмент для выявления истинных причин расхождения результатов исследований, проводимых в разное время. Для этих целей могут быть также использованы статистические методы.
Выводы. Модельные расчеты подтверждают экспериментальные выводы о том, что максимум биомассы фитопланктона в оз. Иссык-Куль приурочен не к поверхности, а к глубинам 25–50 м. Сопоставление вертикального распределения фитопланктона с температурной кривой свидетельствует о всплеске биомассы в подповерхностном слое, где в летний сезон образуется термоклин. Это обусловлено интенсивным перемешиванием нижних, относительно насыщенных биогенами слоев с поверхностными. Кроме того, уровень ФАР, проникающий на данные горизонты, достаточно высок, что в совокупности с активным притоком биогенов обеспечивает высокую скорость роста водорослей. Переменный уровень воды, характерный исключительно для оз. Иссык-Куль (Атлас…, 1987; Черкасов, 1977), усиливает хаотичность влияния внешних факторов, чем обусловливает случайный характер распределения биомассы. Распределение максимальных и минимальных значений интегральной биомассы в столбе воды не поддается выраженному районированию, что объяснимо с точки зрения гидрологии озер, где скорость и направленность перемещения водных масс, влекущее за собой перераспределение температуры и биогенного притока, значительно уступает морям (Михайлов и др., 2007).
Список литературы
Абакумов А.И., Пак С.Я. 2016. Модельные методы оценки содержания фитопланктона и расчет первичной продукции в Японском море по спутниковым данным // Вестник Дальневосточного отделения Российской академии наук. № 4. С. 78.
Атлас Киргизской ССР. Озеро Иссык-Куль. 1987. Москва: Главное управление геодезии и картографии при СМ CССР. Т. 1.
Динамические модели в биологии. 2001–2019. Кафедра биофизики МГУ. http://www.dmb.biophys.msu.ru.
Кукси М.С. 1964. Вертикальное распределение фитопланктона в Новосибирском водохранилище // Водоросли и грибы Западной Сибири (Труды центрального Сибирского ботанического сада). Новосибирск: РИО СО АН СССР, 1964. Ч. 1. Вып. 8. С. 204.
Кулумбаева А.А. 1975. Первые данные о численности и биомассе фитопланктона пелагиали озера Иссык-Куль // Изв. Акад. наук Киргизской ССР. № 2. С. 71.
Левин В.А., Алексанин А.И., Алексанина М.Г. и др. 2010. Разработка технологий спутникового мониторинга окружающей среды по данным метеорологических спутников // Открытое образование. № 5. С. 41.
Маторин Д.Н., Антал Т.К., Шаршенова А.А. и др. 2002. Изучение природного фитопланктона oзера Иссык-Куль, полученная с использованием погружного флуориметра // Вестник Моск. унив. Сер. 16: Биология. № 1. С. 43.
Маторин Д.Н., Рубин А.Б. 2012. Флуоресценция хлорофилла высших растений и водорослей. Ижевск: Институт компьютерных исследований.
Михайлов В.Н., Добровольский А.Д., Добролюбов С.А. 2007. Гидрология: Учебник для вузов. Москва: Высш. школа.
Пак С.Я., Абакумов А.И. 2014. Модельный способ восстановления состояния фитопланктона в вертикальном столбе воды по спутниковым данным о поверхностном слое // Информатика и системы управления. № 3(41). С. 23.
Полуэктов Р.А., Пых Ю.А., Швытов И.А. 1980. Динамические модели экологических систем. Ленинград: Гидрометеоиздат.
Свирежев Ю.М., Логофет Д.О. 1978. Устойчивость биологических сообществ. М.: Главная редакция физико-математической литературы изд-ва “Наука”.
Сиделев С.И., Бабаназарова О.В. 2008. Анализ связей пигментных и структурных характеристик фитопланктона высокоэвтрофного озера // Журн. Сибирского федерального унив. Сер. Биол. Т. 1. № 2. С. 162.
Тыныбеков А.К. 2016. Математическая модель состояния фитопланктона озера Иссык-Куль // Бишкек: Межд. унив. Ала-Тоо. № 1. С. 369.
Тыныбеков А.К., Маторин Д.Н. 2009. Состояние фитопланктона озера Иссык-Куль. Бишкек: Изд-во КРСУ.
Фрисман Е.Я., Жданова О.Л. 2009. Эволюционный переход к сложным режимам динамики численности двухвозрастной популяции // Генетика. Т. 45. № 9. С. 1277.
Черкасов Л.Г. 1977. О водном балансе озера Иссык-Куль // Труды Среднеазиатского регионального научно-исследовательского гидрометеорологического института. Вып. 50(131). С. 81.
Чурилова Т.Я., Суслин В.В., Кривенко О.В. и др. 2016. Спектральный подход к оценке скорости фотосинтеза фитопланктона в Черном море по спутниковой информации: методологические аспекты развития региональной модели // Журн. Сиб. Федер. унив. Сер. Биол. Т. 9. № 4. С. 367.
Abakumov A.I., Izrailsky Yu.G. 2012. Environment influence on the phytoplankton distribution in a basin // Math. Biol. Bioinf. V. 7. № 1. P. 274.
Abakumov A.I., Izrailsky Yu.G., Park S. 2015. Functioning of the phytoplankton in seas and estimates of primary production for aquatic ecosystems // Dev. Environ. Modell. V. 27. P. 339.
Aleksanin A.I., Kim V., Orlova T.Y., Stonik I.V., Shevchenko O.G. 2012. Phytoplankton of the Peter the Great Bay and its remote sensing problem // Oceanology. V. 52. № 2. P. 219.
Allen J.I., Polimene L. 2011. Linking physiology to ecology: towards a new generation of plankton models // J. Plankton Res. V. 33. № 7. P. 989.
Grangere K., Lefebvre S., Menesguen A., Jouenne F. 2009. On the interest of using field primary production data to calibrate phytoplankton rate processes in ecosystem models // Estuarine, Coastal and Shelf Science. V. 81. P. 169.
Hoppenrath M., Elbrachter M., Drebes G. 2009. Marine phytoplankton: selected microphytoplankton species from the North Sea around Helgoland and Sylt. Stuttgart: Schweizerbart.
Kulchin Yu.N., Voznesenskii S.S., Gamayunov E.L. et al. 2008. Fiber optic fluorometer with dipping module // Meas. Tech. V. 51. № 1. P. 40.
Menshutkin V.V., Rukhovets L.A., Filatov N.N. 2014. Ecosystem modeling of freshwater lakes (review): 2. Models of freshwater lake’s ecosystem // Water Resources. V. 41. № 1. P. 32. https://doi.org/10.1134/S0097807814010084
Monod J. 1949. The growth of bacterial cultures // Annu. Rev. Microbiol. V. 111. № 2. P. 371.
Nikolaou A., Hartmann P., Sciandra A. et al. 2016. Dynamic coupling of photoacclimation and photoinhibition in a model of microalgae growth // J. Theor Biol. V. 390. P. 61.
Platt T., Caverhill C., Sathyendranath S. 1991. Basin-scale estimates of oceanic primary production by remote sensing: The North Atlantic // J. Geophys. Res. V. 96. № 8. P. 15 147. https://doi.org/10.1029/91JC01118
Ryabov A.B., Rudolf L., Blasius B. 2010. Vertical distribution and composition of phytoplankton under the influence of an upper mixed layer // J. Theor. Biol. V. 263. P. 120.
Tanaka Y., Mano H. 2012. Functional traits of herbivores and food chain efficiency in a simple aquatic community model // Ecol. Modell. V. 237/238. P. 88.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод