

Биология внутренних вод, 2019, № 4-2, стр. 55-64
Межгодовые изменения зоопланктона Тайширского водохранилища (Западная Монголия)
А. В. Крылов *
Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
Некоузский р-н, Ярославская обл., пос. Борок, Россия
* E-mail: krylov@ibiw.yaroslavl.ru
Поступила в редакцию 06.07.2017
После доработки 05.04.2018
Принята к публикации 27.11.2018
Аннотация
Показано, что в Тайширском водохранилище, созданном на участке верхнего течения горной р. Дзабхан, увеличиваются электропроводность воды, численность и биомасса зоопланктона за счет ракообразных. Наименьшими количественными показателями отличается зоопланктон литоральной зоны водохранилища, что связано с затоплением участков суши, характеризующихся бедными почвами и наземной растительностью, а также волновым перемешиванием. Выявлено, что при повышении уровня воды и объема водохранилища увеличиваются количественные характеристики зоопланктона прибрежья на всех участках водоема, в русловой зоне – лишь в верхней и средней частях, а в приплотинной, напротив, уменьшаются.
ВВЕДЕНИЕ
Такие квазиприродные системы как водохранилища считаются неотъемлемым элементом ландшафта разных природных зон мира. Это связано с тем, что успешность решения вопросов экономического роста стран напрямую определяется обеспечением энергетическими мощностями, одновременно ведущее значение имеют задачи сохранения качества природных условий. В этом случае часто поддержку получают проекты, направленные на создание гидроэлектростанций. В первое десятилетие ХХI в. на территории Монголии создано два водохранилища, одно из них – Тайширское – самый крупный искусственный водоем страны (Углубленный обзор…, 2011). Для создания водохранилища в верховьях р. Дзабхан, крупнейшего водотока Западной Монголии, построена плотина высотой 50 м, длиной по гребню 190 м, заполнение началось в 2007 г., эксплуатация ГЭС – с ноября 2008 г.
Водохранилища по сравнению с естественными водными объектами обладают рядом специфических черт, в первую очередь, искусственно регулируемым уровенным режимом, который определяется эксплуатацией ГЭС, питанием образующего водотока и общим поверхностным стоком. В аридных условиях Западной Монголии эти факторы имеют большое значение. Для выявления изменений сообществ гидробионтов при разных режимах функционирования водохранилищ с успехом может использоваться зоопланктон, который относится к категории сообществ, играющих ведущую роль в структурно-функциональной организации биоты водоемов, выступает в качестве индикатора изменений условий среды.
Цель работы – изучение межгодовых изменений качественного и количественного состава зоопланктона Тайширского водохранилища.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
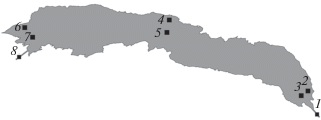
Исследования проводили в августе 2010–2016 гг. на шести станциях – в литоральной зоне и русловой зоне верхнего, среднего и приплотинного участков водохранилища, а также в р. Дзабхан выше границы выклинивания подпора и ниже плотины (рис. 1). На глубинах <1 м пробы собирали ведром, процеживая 100 л воды через газ с размером ячеи 64 мкм, на глубоководных участках – сетью Джеди (размер ячеи 64 мкм, диаметр входного отверстия 12 см), которую протягивали от дна до поверхности (2 подъема). Пробы фиксировали 4%-ным формалином и обрабатывали по стандартной методике (Методика…, 1975). Параллельно с отбором проб измеряли глубину, температуру и электропроводность воды.
Рис. 1.
Местонахождение станций на р. Дзабхан и Тайширском водохранилище: 1 – р. Дзабхан выше водохранилища, 2 – прибрежье в верховье водохранилища, 3 – центр верховья водохранилища, 4 – прибрежье на среднем участке водохранилища, 5 – центр среднего участка водохранилища, 6 – прибрежье в приплотинном участке водохранилища, 7 – центр приплотинного участка водохранилища, 8 – р. Дзабхан ниже водохранилища.
Для статистического анализа данных использовали U-тест Манна–Уитни, Н-критерий Краскела–Уоллиса и коэффициент корреляции Спирмена (p < 0.05).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
В период с 2010 по 2014 гг. средние за май–июль значения уровня воды и объема водохранилища повышались, в 2015 г. – снижались, в 2016 г. зарегистрированы аномально высокие величины, вызванные дождевым паводком (табл. 1). В августе, во время отбора проб, наименьший уровень воды отмечен в 2010 г., в 2011 г. он значительно (на 8.3 м) повысился и до 2015 г. ежегодно возрастал на 0.1–2 м, в 2015 г. снизился на 2.3 м, а в 2016 г. достиг максимально возможной величины, при которой вода переливалась через поверхностный водослив (табл. 1). Аналогично в периоды сборов изменялся и объем водохранилища (табл. 1).
Таблица 1.
Межгодовые изменения уровня воды и объема водохранилища
| Год | Средний уровень, м | Средний объем, млн м3 | ||
|---|---|---|---|---|
| май–июль | август | май–июль | август | |
| 2010 | 1687.5 | 1693.3 | 498.6 | 498.6 |
| 2011 | 1696.7 | 1701.6 | 643.7 | 898.6 |
| 2012 | 1698.1 | 1699.0 | 710.8 | 756.3 |
| 2013 | 1698.2 | 1699.7 | 715.8 | 793.0 |
| 2014 | 1700.2 | 1701.3 | 820.0 | 878.5 |
| 2015 | 1697.8 | 1699.0 | 696.0 | 756.3 |
| 2016 | >1704.0 | >1704.0 | ~1031.2 | ~1031.2 |
Река Дзабхан выше водохранилища (ст. 1). Скорость течения в реке в разные годы колебалась от 0.25 до 0.96 м/с, значительно варьировала температура (от 13.7 до 20.1°С) и электропроводность (от 92 до 152 мСм/см) воды (табл. 2). В составе зоопланктона обнаружено 18 видов беспозвоночных (Rotifera – 14 и Cladocera – 4). В разные годы количество видов варьировало от 2 до 7, наибольшее количество отмечено в 2014 г. (табл. 3). Численность и биомасса организмов возрастали в период до 2014 г., в 2015–2016 гг. сокращались (табл. 4). Основой численности и биомассы были коловратки, наибольшая доля ракообразных отмечена в 2013 г. (рис. 2). По численности в разные годы доминировали Euchlanis dilatata Ehrenberg (2010–2016 гг.), E. meneta Myers (2011), Brachionus calyciflorus Pallas (2010), Trichotria pocillum Muller (2011, 2014), Keratella cochlearis (Gosse) (2016), K. quadrata (Muller) (2012), Kellicottia longispina (Kellicott.) (2016), по биомассе – Euchlanis dilatata (2010–2016), E. meneta (2011), Brachionus calyciflorus (2010), Keratella cochlearis (2016), K. quadrata (2012), Kellicottia longispina (2016), Chydorus sphaericus (O.F. Muller) (2013).
Таблица 2.
Некоторые абиотические характеристики воды р. Дзабхан и Тайширского водохранилища в период исследований
| Показатель | Год | Станции* | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | ||
| Глубина, м | 2010 | 0.3 | 0.5 | – | 0.5 | – | 0.5 | 25.0 | 0.5 |
| 2011 | 0.3 | 0.5 | – | 0.5 | – | 0.5 | 27.0 | 0.5 | |
| 2012 | 0.3 | 0.5 | 2.3 | 0.5 | 6.0 | 0.5 | 27.0 | 0.5 | |
| 2013 | 0.3 | 0.5 | 2.0 | 0.5 | 8.0 | 0.5 | 25.0 | 0.5 | |
| 2014 | 0.3 | 0.5 | 2.3 | 0.5 | 7.0 | 0.5 | 25.0 | 0.5 | |
| 2015 | 0.3 | 0.5 | 2.5 | 0.5 | 8.0 | 0.5 | 25.0 | 0.5 | |
| 2016 | 0.3 | 0.5 | 3.0 | 0.5 | 8.0 | 0.5 | 25.0 | 0.5 | |
| Температура, °С | 2010 | 15.9 | 16.8 | – | 20.4 | – | 19.6 | 17.6 | 16.7 |
| 2011 | 19.0 | 19.2 | – | 21.1 | – | 21.0 | 20.0 | 12.4 | |
| 2012 | 18.6 | 20.1 | 17.8 | 22.4 | 22.9 | 19.8 | 19.7 | 9.8 | |
| 2013 | 20.0 | 21.6 | 20.3 | 21.5 | 20.5 | 19.5 | 19.3 | 13.1 | |
| 2014 | 13.7 | 16.8 | 16.1 | 19.2 | 17.9 | 19.1 | 17.2 | 14.7 | |
| 2015 | 15.2 | 17.1 | 16.5 | 18.8 | 18.6 | 18.1 | 17.5 | 18.0 | |
| 2016 | 16.3 | 18.3 | 18.2 | 18.8 | 18.0 | 17.3 | 17.9 | 17.2 | |
| Электропроводность, мСм/см |
2010 | 110.0 | 110.0 | – | 137.0 | – | 180.0 | 180.0 | 133.0 |
| 2011 | 92.0 | 101.0 | – | 154.0 | – | 194.0 | 194.0 | 114.0 | |
| 2012 | 123.6 | 148.0 | 153.8 | 211.5 | 202.9 | 200.0 | 220.0 | 156.0 | |
| 2013 | 127.0 | 141.0 | 130.0 | 221.0 | 221.0 | 239.0 | 229.0 | 221.0 | |
| 2014 | 152.0 | 165.0 | 155.0 | 265.0 | 215.0 | 235.0 | 233.0 | 238.0 | |
| 2015 | 125.0 | 140.0 | 140.0 | 272.0 | 265.0 | 266.0 | 270.0 | 280.0 | |
| 2016 | 152.0 | 195.0 | 196.0 | 198.0 | 212.0 | 195.0 | 212.0 | 203.0 | |
* Местонахождение станций см. рис. 1; “–” – данные отсутствуют.
Таблица 3.
Количество видов зоопланктона в период исследований р. Дзабхан и Тайширского водохранилища
| Станция | Зоопланктон | 2010 г. | 2011 г. | 2012 г. | 2013 г. | 2014 г. | 2015 г. | 2016 г. |
|---|---|---|---|---|---|---|---|---|
| 1 | Rotifera | 5 | 3 | 3 | 3 | 6 | 2 | 2 |
| Copepoda | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Cladocera | 0 | 0 | 1 | 2 | 1 | 0 | 0 | |
| Всего | 5 | 3 | 4 | 5 | 7 | 2 | 2 | |
| 2 | Rotifera | 10 | 5 | 9 | 9 | 7 | 11 | 6 |
| Copepoda | 0 | 0 | 1 | 2 | 1 | 0 | 4 | |
| Cladocera | 0 | 1 | 1 | 4 | 6 | 0 | 5 | |
| Всего | 10 | 6 | 11 | 15 | 14 | 11 | 15 | |
| 3 | Rotifera | – | – | 3 | 4 | 9 | 6 | 7 |
| Copepoda | – | – | 0 | 0 | 3 | 0 | 4 | |
| Cladocera | – | – | 0 | 1 | 6 | 3 | 3 | |
| Всего | – | – | 3 | 5 | 18 | 9 | 14 | |
| 4 | Rotifera | 6 | 5 | 4 | 6 | 7 | 10 | 6 |
| Copepoda | 0 | 0 | 2 | 1 | 1 | 1 | 3 | |
| Cladocera | 0 | 0 | 2 | 3 | 6 | 4 | 5 | |
| Всего | 6 | 5 | 8 | 10 | 14 | 15 | 14 | |
| 5 | Rotifera | – | – | 9 | 2 | 6 | 5 | 4 |
| Copepoda | – | – | 4 | 5 | 2 | 2 | 5 | |
| Cladocera | – | – | 3 | 3 | 2 | 3 | 3 | |
| Всего | – | – | 16 | 10 | 10 | 10 | 12 | |
| 6 | Rotifera | 8 | 9 | 2 | 5 | 7 | 4 | 8 |
| Copepoda | 3 | 1 | 4 | 4 | 3 | 2 | 2 | |
| Cladocera | 2 | 0 | 2 | 2 | 1 | 2 | 2 | |
| Всего | 13 | 10 | 8 | 11 | 11 | 8 | 12 | |
| 7 | Rotifera | 8 | 5 | 4 | 5 | 9 | 2 | 4 |
| Copepoda | 3 | 4 | 4 | 6 | 3 | 4 | 6 | |
| Cladocera | 4 | 2 | 2 | 3 | 2 | 1 | 5 | |
| Всего | 15 | 11 | 10 | 14 | 14 | 7 | 15 | |
| 8 | Rotifera | 8 | 5 | 7 | 3 | 12 | 9 | 5 |
| Copepoda | 2 | 2 | 2 | 2 | 2 | 0 | 2 | |
| Cladocera | 1 | 0 | 1 | 1 | 3 | 0 | 2 | |
| Всего | 11 | 7 | 10 | 6 | 17 | 9 | 9 |
Таблица 4.
Численность (N) и биомасса (В) зоопланктона р. Дзабхан и Тайширского водохранилища в период исследований
| Год | Станции | |||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |
| N, тыс. экз./м3 | ||||||||
| 2010 | 0.2 | 0.3 | 0.0 | 4.3 | 0.0 | 87.0 | 1.9 | 0.8 |
| 2011 | 0.4 | 1.9 | 0.0 | 3.9 | 0.0 | 0.4 | 28.1 | 1.6 |
| 2012 | 0.6 | 0.9 | 0.2 | 7.0 | 17.4 | 2.5 | 17.5 | 4.1 |
| 2013 | 0.6 | 13.2 | 0.4 | 7.4 | 60.4 | 3.7 | 30.1 | 2.4 |
| 2014 | 1.8 | 10.0 | 64.0 | 3.7 | 23.1 | 7.7 | 129.3 | 2.6 |
| 2015 | 0.5 | 3.5 | 0.8 | 8.7 | 29.5 | 2.4 | 25.5 | 0.7 |
| 2016 | 0.2 | 30.4 | 1.1 | 25.3 | 69.9 | 1.1 | 21.1 | 2.0 |
| B, г/м3 | ||||||||
| 2010 | 0.0006 | 0.001 | – | 0.016 | – | 0.070 | 0.012 | 0.003 |
| 2011 | 0.0007 | 0.003 | – | 0.016 | – | 0.001 | 1.848 | 0.006 |
| 2012 | 0.0007 | 0.003 | 0.0003 | 0.253 | 0.836 | 0.061 | 1.153 | 0.005 |
| 2013 | 0.0015 | 0.051 | 0.0032 | 0.033 | 1.533 | 0.015 | 1.643 | 0.023 |
| 2014 | 0.0022 | 0.711 | 1.7425 | 0.087 | 0.763 | 0.015 | 1.696 | 0.016 |
| 2015 | 0.0013 | 0.007 | 0.0021 | 0.013 | 0.988 | 0.015 | 2.309 | 0.001 |
| 2016 | 0.0001 | 1.028 | 0.0039 | 1.172 | 7.654 | 0.004 | 0.734 | 0.041 |
Рис. 2.
Доля таксономических групп в общей численности (а) и биомассе (б) зоопланктона р.
Дзабхан и Тайширского водохранилища в период исследований. (◻) – Rotifera, ( ) – Copepoda, (◼) – Cladocera.
) – Copepoda, (◼) – Cladocera.

Верхний участок водохранилища. В прибрежье (ст. 2) температура воды по сравнению с незарегулированным водотоком увеличивалась в среднем на 1.6°С, электропроводность – на 16.9 мСм/см (табл. 2). В центре участка (ст. 3) температура была выше, чем в реке в среднем на 1.2°С, но на 1.0°С ниже, чем в прибрежье.
В составе зоопланктона прибрежной зоны за весь период изучения отмечено 44 вида беспозвоночных (Rotifera – 29, Cladocera – 10, Copepoda – 5), в центре, где наблюдений в 2010 и 2011 гг. не проводили, всего обнаружено 32 вида (Rotifera – 20, Cladocera – 7 и Copepoda – 5). В 2012–2014 гг. и в 2016 г. в прибрежье и в центре количество видов увеличивалось, в том числе за счет ракообразных (табл. 3).
Численность и биомасса зоопланктона в прибрежье значительно варьировали, причем, в период с 2010 по 2014 гг. они возрастали, в 2015 г. значительно снизились, а в 2016 г. достигли максимума (табл. 4). Основой численности зоопланктона до 2013 г. были коловратки, но с 2011 г. постепенно увеличивалась доля ракообразных, в 2014 г. достигшая максимума за счет Cladocera, в 2015 г. значительно повысилась доля Rotifera (рис. 2). Среди доминирующих по численности организмов были Euchlanis dilatata (2010–2016 гг.), E. deflexa Gosse (2010), E. meneta (2011), Trichotria pocillum (2011), Brachionus calyciflorus (2010, 2016), B. quadridentatus Ehrenberg (2012, 2016), Lecane luna (Muller) (2012), L. mira Murray (2010), Kellicottia longispina (2014), науплиусы Cyclopoida (2012, 2013), Eucyclops serrulatus (Fischer) (2012), Cyclops abyssorum (G.O. Sars) (2016), Chydorus sphaericus (2013), Daphnia (D.) galeata G.O. Sars, D. (D.) hyalina Leydig., D. (D.) galeata × D. (D.) hyalinа (2014), Alona affinis (Leydig) (2014), Bosmina (B.) longirostris (O.F. Muller) (2016). Основу биомассы в 2010, 2011 и 2015 гг. составляли коловратки, в 2012 г. – веслоногие ракообразные, в 2013, 2014 и 2016 гг. – ветвистоусые (рис. 2). Среди доминантов обнаружены Euchlanis dilatata (2011–2015), E. meneta (2011), Brachionus calyciflorus (2010), Asplanchna girodi Guerne (2010), Cyclops abyssorum (2016), Eucyclops serrulatus (2012), Chydorus sphaericus (2013), Daphnia (2014, 2015).
В русловой части численность и биомасса зоопланктона были меньше, чем в прибрежье, исключение наблюдали лишь в 2014 г., когда в центре зарегистрировали максимальные количественные характеристики (табл. 4). В течение исследованного периода численность и биомасса планктонных беспозвоночных постепенно увеличивались, а в 2014 г. отмечен их резкий скачок. Основой численности были коловратки, только в 2016 г. – ракообразные (рис. 2). Среди доминирующих по численности организмов отмечены Euchlanis dilatata (2012–2015 гг.), E. meneta (2012, 2013), Kellicottia longispina (2014), Brachionus quadridentatus (2015), науплиусы Cyclopoida (2016), Bosmina (B.) longirostris (2016), Daphnia (2014, 2016). Основу биомассы в 2012 и 2015 гг. составляли коловратки, в остальные годы – ветвистоусые ракообразные (рис. 2), доминировали Euchlanis dilatata (2012, 2013, 2015), E. meneta (2012), Brachionus quadridentatus (2015), Alona affinis (2013, 2015), Daphnia (2014, 2016).
Средний участок водохранилища. В литоральной зоне температура воды была больше, чем в прибрежье верховья, в среднем на 1.7°С, электропроводность – на 67.6 мСм/см (табл. 2). На русловом участке температура воды превышала таковую в центре верховья в среднем на 1.8°С, электропроводность – на 68.2 мСм/см.
В составе зоопланктона литоральной зоны обнаружен 31 вид (Rotifera – 19, Cladocera – 9, Copepoda – 3). Наименьшее количество видов, которое представлено только коловратками, отмечено в 2010 г., в 2011–2013 гг. оно возрастало, максимальные величины зарегистрированы в 2014–2016 гг. (табл. 3). В русловой части, где в 2010–2011 гг. исследований не проводили, всего зарегистрировано 24 вида (Rotifera – 13, Cladocera – 4, Copepoda – 7).
Численность зоопланктона в литоральной зоне в течение периода исследований плавно возрастала, лишь в 2014 г. наблюдалось уменьшение плотности организмов, а в 2016 г. – ее резкое повышение (табл. 4). Изменялось также соотношение таксономических групп зоопланктеров: в 2010 г. отмечены только Rotifera, доля которых до 2013 г. сокращалась, в 2014–2015 гг. возросла вновь, в 2016 г. лидирующую позицию заняли Cladocera (рис. 2). Численность зоопланктона здесь значительно превышала таковую литоральной зоны верховья в начале периода изучения – в 2010 г., в 2013, 2014 и 2016 гг. снижалась, при этом чаще всего доля коловраток была меньше, ракообразных – больше. Среди доминирующих по численности организмов отмечены Euchlanis meneta (2013 г.), Brachionus quadridentatus (2010), Filinia longiseta (Ehrenberg) (2012, 2015), Polyarthra vulgaris Carlin (2010), Kellicottia longispina (2014), Keratella cochlearis (2015), Asplanchna brightwelli Gosse (2014), науплиусы Cyclopoida (2012, 2013, 2015), копеподиты Cyclopoida (2013), Bosmina (B.) longirostris (2013, 2016), Daphnia (2012, 2014), Leptodora kindtii (Focke) (2016).
За период изучения в прибрежье отмечены высокие биомассы зоопланктона в 2012 и 2016 гг. (табл. 4). Основу биомассы в 2010 г. составляли коловратки, с 2011 г. преобладали ракообразные, среди которых в 2011–2014 и 2016 гг. превалировали ветвистоусые рачки, в 2015 г. доля всех таксономических групп была примерно одинакова (рис. 2). Биомасса здесь обычно превышала таковую прибрежья верховья (кроме 2013 и 2014 гг.), хотя наблюдался четкий тренд снижения разницы. По биомассе доминировали Brachionus quadridentatus (2010), Asplanchna brightwelli (2014, 2015), науплиусы Cyclopoida (2013, 2015), копеподиты Cyclopoida (2013), Cyclops abyssorum (2016), Bosmina (B.) longirostris (2013), Daphnia (2012, 2014, 2015), Leptodora kindtii (2016).
В центре среднего участка численность и биомасса зоопланктона были выше, чем в прибрежье в 1.2–24.8 и 3.3–78.1 раз соответственно (табл. 4). Максимальная численность зоопланктона зарегистрирована в 2013 и 2016 гг. Доля таксономических групп значительно варьировала, в 2012–2014 гг. увеличивалась доля коловраток, в 2015 г. – веслоногих, в 2106 г. – ветвистоусых ракообразных (рис. 2). Среди доминирующих по численности организмов отмечены Conochilus hippocrepis (Schrank) (2012 г.), Asplanchna brightwelli (2013, 2014), Kellicottia longispina (2014, 2016), Keratella cochlearis (2015, 2016), науплиусы Cyclopoida (2015), Eudiaptomus graciloides Lilljeborg (2015), Acanthodiaptomus denticornis Wierzejski (2012, 2016), Daphnia (2013, 2014, 2016). Биомасса зоопланктона в 2012–2015 гг. варьировала незначительно, однако в 2016 г. наблюдалось ее резкое увеличение (табл. 4). Основу биомассы на протяжении всего периода изучения составляли Cladocera, однако в 2013 и 2014 гг. отмечено повышение доли Rotifera, а в 2015 г. – Copepoda (рис. 2). Доминировали по биомассе Asplanchna brightwelli (2013, 2014), Eudiaptomus graciloides (2015), Acanthodiaptomus denticornis (2012), Daphnia sp. (2012–2016). По сравнению с прибрежьем, а также центром верховья, численность и биомасса в центре среднего участка водоема были значительно выше.
Приплотинный участок водохранилища. В литоральной зоне температура воды была в среднем на 1.3°С ниже, чем в прибрежье средней части, а электропроводность воды в самом начале исследований (2010 г.) была выше, затем ниже таковой в прибрежье средней части, с минимальной разницей в 2016 г. (табл. 2). В русловой зоне по сравнению с прибрежьем различия по температуре воды также, как и в среднем участке водоема, были незначительными – в среднем 0.7°С, малая величина различий отмечена и для электропроводности воды – 4.1 мСм/см.
В составе зоопланктона литоральной зоны обнаружено 35 видов беспозвоночных (20 – Rotifera, 7 – Cladocera, 8 – Copepoda), в пелагиали – 33 вида (Rotifera – 16, Cladocera – 8, Copepoda – 9). В прибрежье наибольшее количество видов зарегистрировано в первый год исследований, в 2011–2012 гг. отмечено их снижение, в последующие два года – повышение, однако в 2015 и 2016 гг. – вновь сокращение (табл. 3). Лишь в первый год изучения количество видов в приплотинном плесе превышало таковое в прибрежном участке средней части водохранилища, однако в дальнейшем оно было меньше, наибольшая разница отмечена по разнообразию Cladocera. В пелагиали изменение количества видов в основном повторяло динамику, обнаруженную в прибрежье, однако было выше в 1.1–1.5 раза, за исключением 2015 г. По сравнению с центром среднего участка водохранилища, кроме 2010 и 2015 гг., число видов зоопланктеров было незначительно больше, причем, в основном за счет Copepoda и Rotifera.
Наибольшая численность зоопланктона в литоральной зоне отмечена в 2010 г., затем, после резкого падения, до 2014 г. наблюдалось увеличение плотности организмов, которая вновь снизилась в 2015 и 2016 гг. (табл. 4). По сравнению с прибрежьем в средней части водохранилища, численность была меньше (за исключением 2010 и 2014 гг.). В период с 2010 до 2012 гг. значительно сокращалась доля коловраток, но в следующие два года их относительное обилие возрастало. В 2015 г. преобладали веслоногие ракообразные, в 2016 г. – коловратки и ветвистоусые рачки (рис. 2). По сравнению с зоопланктоном средней части водоема отмечено увеличение доли Rotifera и Copepoda и сокращение доли Cladocera. Среди доминирующих по численности организмов отмечены Polyarthra vulgaris (2010 г.), Kellicottia longispina (2011, 2014), Conochilus hippocrepis (2014), C. unicornis Rousselet (2011), Keratella cochlearis (2013), науплиусы Cyclopoida (2011–2016), копеподиты Cyclopoida (2015), Chydorus sphaericus (2013), Bosmina (B.) longirostris (2016), Daphnia (2012). Наибольшие биомассы зоопланктона зарегистрированы в 2010 и 2012 гг., наименьшие – в 2011 и 2016 гг. (табл. 4). По сравнению с зоопланктоном прибрежья средней части водохранилища биомасса была меньше (за исключением 2010 и 2015 гг.). Основой биомассы были ракообразные, в 2011 и 2013–2015 гг. – веслоногие, а в 2012 и 2016 гг. – ветвистоусые, только в 2010 г. преобладали коловратки. По сравнению с сообществами в прибрежье средней части водоема в приплотинном плесе сокращались доли Cladocera и Rotifera, и возрастала доля Copepoda (рис. 2). Доминировали по биомассе Polyarthra vulgaris (2010 г.), Asplanchna brightwelli (2011), науплиусы Cyclopoida (2013), копеподиты Cyclopoida (2015), Acanthodiaptomus denticornis (2014), Cyclops strenuus (Fischer) (2011), Eudiaptomus graciloides (2015), Chydorus sphaericus (2013), Bosmina (B.) longirostris (2016) и Daphnia (2012, 2015).
В пелагиали наименьшая численность зоопланктона была в 2010 г., наибольшая – в 2014 г. (табл. 4). В 2010, 2014 и в 2016 гг. основу численности составляли коловратки. Численность зоопланктона в литоральной зоне превышала таковую в пелагиали (за исключением 2010 г.), в 2010 и 2011 гг. была выше доля Copepoda, в 2012–2014 и 2016 гг. – Rotifera, в 2012 и 2015 гг. – Cladocera. По сравнению с центром средней части водохранилища численность была меньше (за исключением 2014 г.), при этом наиболее значимо сокращалась доля ветвистоусых ракообразных (рис. 2). Доминировали по численности Polyarthra vulgaris (2010), Kellicottia longispina (2014, 2016), Conochilus hippocrepis (2012, 2014), C. unicornis Rousselet (2010), Asplanchna brightwelli (2013, 2014), Keratella cochlearis (2016), Filinia longiseta (2016), науплиусы Cyclopoida (2011–2015), копеподиты Calanoida (2015), Eudiaptomus graciloides (2015), Acanthodiaptomus denticornis (2012), Cyclops strenuus (2011, 2012). Минимальная биомасса зафиксирована в 2010 г., в 2011 г. отмечено ее резкое увеличение, повышение с 2012 по 2015 гг., в 2016 г. – понижение (табл. 4). По сравнению с литоральной зоной плеса биомасса здесь была значительно больше (кроме 2010 г.) за счет ветвистоусых ракообразных (рис. 2), по сравнению с центром средней части водоема – она также была выше (кроме 2016 г.). По биомассе доминировали Asplanchna brightwelli (2014 г.), Acanthodiaptomus denticornis (2011, 2012, 2016), Eudiaptomus graciloides (2015), Cyclops strenuus (2010) и Daphnia (2010–2016).
Река Дзабхан ниже водохранилища. По сравнению с участком водотока выше зоны выклинивания подпора в 2010 и 2014–2016 гг. температура воды была выше, в 2011–2013 гг. – ниже, по сравнению с приплотинным глубоководным участком всегда (за исключением 2015 г.) – ниже (табл. 2), что, по-видимому, связано с поступлением воды из придонного слоя плеса. Электропроводность воды в реке ниже плотины в среднем была больше на 66.2 мСм/см, чем до водохранилища, по сравнению с глубоководным участком приплотинной зоны в 2010–2013 и 2016 гг. – меньше в среднем на 41.6 мСм/см, однако в 2014 и 2015 гг. незначительно больше (на 5 и 10 мСм/см соответственно).
В составе зоопланктона обнаружено 29 видов беспозвоночных (Rotifera – 22, Cladocera – 4, Copepoda – 3). По сравнению с участком реки выше водохранилища, количество видов было больше, а по сравнению с приплотинным плесом – меньше (за исключением 2014 и 2015 гг.), когда ниже плотины отмечали больше видов коловраток (табл. 3). Численность зоопланктона варьировала в широких пределах при максимуме в 2012 г. и минимуме в 2015 г. (табл. 4). Плотность зоопланктона здесь всегда была больше, чем в реке выше водохранилища, и меньше, чем в приплотинном плесе. Основой численности были коловратки, лишь в 2016 г. преобладали веслоногие ракообразные (рис. 2). По сравнению с рекой выше влияния подпора водохранилища, ниже плотины снижалась доля коловраток и возрастала доля ракообразных, а по сравнению с приплотинным плесом чаше всего была выше доля коловраток. Среди организмов, доминирующих по численности, зарегистрированы Euchlanis dilatata (2010–2016), E. deflexa (2010), Mytilina ventralis Ehrenberg (2012), Trichotria pocillum (2014), Filinia longiseta (2014–2016), Kellicottia longispina (2015), Keratella quadrata (2015), Chydorus sphaericus (2013, 2016). Минимальная биомасса зоопланктона была в 2015 г., максимальная – в 2016 г. (табл. 4). Основу биомассы в 2010–2013 гг. составляли веслоногие ракообразные, в 2014 и 2016 гг. – ветвистоусые, в 2015 г. – коловратки (рис. 2). По сравнению с рекой выше водохранилища, биомасса была больше (кроме 2015 г.), сокращалась доля коловраток и повышалась доля веслоногих ракообразных. По сравнению с приплотинным плесом биомасса зоопланктона, в реке всегда была меньше, в большинстве случаев доли коловраток и веслоногих ракообразных были больше, доля ветвистоусых ракообразных – меньше. По биомассе доминировали Euchlanis dilatata (2010–2016), Asplanchna brightwelli (2015), Cyclops strenuus (2010), Eudiaptomus graciloides (2015), Acanthodiaptomus denticornis (2014), Chydorus sphaericus (2013, 2016), Daphnia (2014).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Выделяют три этапа формирования биоты водохранилищ – становления, депрессии и относительной стабилизации (Кудерский, 1992; Шаронов, 1966). Этап становления, т.е. изменения состава и количественных характеристик беспозвоночных, развивающихся в условиях проточности, к сообществам, характерным для лимнических систем, как в равнинных реках России (Дзюбан, 1959), прошел в первые 3–4 года (Крылов и др., 2014). По сравнению с равнинными, этот этап в Тайширском водохранилище в меньшей степени зависел от сроков наполнения, что, по-видимому, связано с бедностью затапливаемых почв и растительного покрова. Как и на волжских водохранилищах (Луферова, 1963), этап депрессии в зоопланктоне не был выражен, через 3–4 года начался этап стабилизации, в течение которого дальнейшие изменения зоопланктона могут быть связаны с формированием зарослей водных растений в литоральной зоне, а также особенностями гидрологического режима и метеорологических условий.
Тип формирования зоопланктона Тайширского водохранилища – пойменный, характерный для водохранилищ, создаваемых на реках, почти лишенных зоопланктона (Луферова, 1963). В условиях зарегулирования закономерно увеличились число видов, численность и биомасса зоопланктона за счет ракообразных. Из-за бедности почв и наземной растительности на затопленных участках суши, а также волнового перемешивания большинство количественных показателей зоопланктона в прибрежье было меньше, чем в русловой зоне водохранилища, а по числу видов Rotifera (U = 111.5, р = 0.049), численности Copepoda (U = 94.5, р = 0.013) и Cladocera (U = 102.0, р = = 0.024), общей биомассе (U = 99.0, р = 0.019), биомассе веслоногих (U = 105.0, р = 0.030) и ветвистоусых (U = 83.0, р = 0.005) ракообразных различия были статистически достоверны. На верхнем участке водохранилища значимых различий между показателями зоопланктона литоральной и русловой зон не обнаружено, что, скорее всего, связано с особенностями морфометрии: минимальной шириной, глубиной, а также поступлением вод р. Дзабхан. В центре среднего участка водохранилища достоверно выше были численность Rotifera (U = 1.0, р = 0.016) и Copepoda (U = 3.0, р = = 0.047), общая численность (U = 2.0, р = 0.028) и общая биомасса (U = 3.0, р = 0.047), биомасса веслоногих (U = 3.0, р = 0.047) и ветвистоусых (U = 3.0, р = 0.047) ракообразных; в центре приплотинного участка – количество видов Rotifera (U = 8.5, р = 0.040), численность (U = 6.0, р = 0.018) и биомасса (U = 5.0, р = 0.012) Copepoda (U = 6.0, р = 0.018) и Cladocera (U = 2.0, р = 0.004), а также общая биомасса зоопланктона (U = 5.0, р = 0.012).
По продольному профилю водохранилища достоверно увеличивалась электропроводность воды (в литоральной зоне r = 0.60, в центре r = 0.61), причем, между верхним и средним, верхним и приплотинным участками различия были значимыми (для прибрежных зон Н = 8.73, р = 0.0126, для русловых – Н = 8.74, р = 0.0127). При увеличении электропроводности воды на прибрежных участках повышались число видов Copepoda (r = = 0.46) и Cladocera (r = 0.52), численность (r = = 0.54) и биомасса (r = 0.50) веслоногих ракообразных, на русловых – количество видов Copepoda (r = 0.58), общая численность (r = 0.53) и общая биомасса (r = 0.54), численность (r = 0.79) и биомасса (r = 0.79) веслоногих ракообразных, биомасса ветвистоусых (r = 0.56).
Кроме того, количественные показатели зоопланктона в литоральной зоне (особенно на верхнем и среднем участках) возрастали при увеличении уровня воды (табл. 5). По-видимому, этим определялись и межгодовые различия распределения зоопланктона по продольному профилю водохранилища в литоральной зоне. Так, наибольшее количество видов в пробе, численность и биомассу регистрировали на верхнем и среднем участках и лишь однажды (при снижении уровня воды в 2015 г.) максимальной биомассой отличалось сообщество нижнего участка (табл. 4), для которого получены отрицательные коэффициенты корреляции с уровнем воды в периоды отбора проб (табл. 5).
Таблица 5.
Коэффициенты корреляции Спирмена между уровнем воды (средние значения) в водохранилище и показателями количественного развития зоопланктона его литоральной зоны
| Показатель | Средний уровень воды за май–август | Средний уровень воды за август | ||||||
|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | I | II | III | IV | |
| Количество видов | ||||||||
| Rotifera | – | – | – | – | –0.88 | – | – | – |
| Copepoda | 0.90 | – | – | – | – | – | – | – |
| Cladocera | 0.87 | 0.86 | – | 0.69 | – | – | – | – |
| Весь зоопланктон | 0.89 | – | – | 0.53 | – | – | – | – |
| Численность | ||||||||
| Rotifera | 0.86 | – | – | – | – | – | – | – |
| Copepoda | 0.95 | – | – | 0.44 | – | – | –0.95 | – |
| Cladocera | 0.91 | – | – | 0.71 | – | – | – | 0.48 |
| Весь зоопланктон | 0.86 | – | – | – | – | – | – | – |
| Биомасса | ||||||||
| Rotifera | – | – | – | – | – | – | – | – |
| Copepoda | 0.96 | – | – | 0.52 | – | – | –0.85 | – |
| Cladocera | 0.83 | 0.86 | – | 0.59 | 0.79 | – | –0.76 | – |
| Весь зоопланктон | 0.89 | 0.79 | – | 0.48 | 0.76 | – | –0.90 | – |
В большинстве водохранилищ пойменного типа наиболее высокие показатели количественного развития зоопланктона в русловой зоне наблюдаются на приплотинных участках (Ермолаева, 2008). И в Тайширском водохранилище по биомассе, как правило, преобладал зоопланктон нижнего участка (2012, 2013, 2015 гг.), лишь при максимальном уровне воды и сбросе через поверхностный водослив в 2016 г. – среднего (табл. 4).
По числу видов лидировали сообщества среднего (2012 и 2015 гг.) и нижнего (2013 и 2016 гг.) участков (лишь в 2014 г. – верхнего), по численности – среднего (2012, 2013, 2015 и 2016 гг.) и нижнего (2014 г.).
Достоверных статистических связей количественных показателей зоопланктона русловой зоны водохранилища с уровнем и объемом воды не обнаружено, тем не менее, прослеживается тенденция увеличения средней биомассы зоопланктона водохранилища при увеличении уровня воды, причем, наибольшая величина была в год максимального подъема. Но на разных участках водохранилища направления изменений зоопланктона различались. Так, в верховье и в средней части водохранилища при увеличении уровня воды наблюдалось повышение биомассы, на верхнем участке максимум был при умеренном подъеме в 2014 г., а на среднем участке – при максимальном подъеме в 2016 г. (табл. 4). В приплотинном участке, напротив, увеличение биомассы отмечено при снижении уровня воды, а уменьшение – при максимальном подъеме. Ситуация, наблюдающаяся в приплотинном плесе, описана Монаковым (1958) и впоследствии подтверждена Дзюбаном (1959), как характерная для волжских водохранилищ, в которых при увеличении уровня воды биомасса зоопланктона сокращалась из-за “разбавления” небогатого зоопланктона большими объемами воды. Однако в верховье и средней части Тайширского водохранилища изменения биомассы сообществ определяются, по-видимому, не столько уровнем воды, сколько дальностью проникновения речных вод: при небольших расходах, приводящих к подъему уровня воды в водохранилище на ~2 м, приносимые рекой вещества аккумулируются на верхнем участке, при увеличении расходов – на среднем. Кроме того, повышение уровня воды связано, как правило, с увеличением поверхностного стока, в результате которого соответственно возрастает поступление органических и биогенных веществ с водосбора. Поступающие вещества могут оказывать влияние на развитие гидробионтов на верхнем и среднем участках водохранилища с меньшими глубиной и шириной.
Водохранилище оказывало заметное влияние и на реку ниже плотины. Благодаря его созданию электропроводность воды в р. Дзабхан ниже плотины возросла в среднем в 1.5 раза и различия между ее величинами выше и ниже водохранилища были достоверными (U = 7.0, р = 0.025). В зоопланктоне реки ниже плотины, по сравнению с участком реки выше подпора водохранилища, достоверно выше были общее количество видов (U = 1.5, р = 0.004) и веслоногих ракообразных (U = 3.5, р = 0.008), общие численность (U = 3.0, р = 0.007) и биомасса (U = 5.0, р = 0.015), численность коловраток (U = 7.0, р = 0.029), численность (U = 5.5, р = 0.018) и биомасса (U = 5.5, р = = 0.018) веслоногих ракообразных.
Выводы. Минимальные количественные показатели развития зоопланктона Тайширского водохранилища зарегистрированы в литоральной зоне, что связано с затоплением участков суши, характеризующихся бедными почвами и наземной растительностью, а также ветровым перемешиванием. По продольному профилю водохранилища возрастает электропроводность воды, численность и биомасса зоопланктона за счет ракообразных. Межгодовые изменения количественных характеристик зоопланктона литоральной зоны зависят от уровня воды, что определяет распределение сообществ по продольному профилю водохранилища. При повышении уровня воды и объема водохранилища в русловой зоне верхней и средней частей водохранилища биомасса зоопланктона увеличивается, в нижней – уменьшается.
Список литературы
Дзюбан Н.А. 1959. О формировании зоопланктона водохранилищ // Труды VI совещания по проблемам биологии внутренних вод. Москва: Изд-во АН СССР. С. 597.
Ермолаева Н.И. 2008. Водные экосистемы. Особенности формирования зоопланктона водохранилищ: аналитический обзор // Гос. публич. науч.-техн. б-ка Сиб. отд. Рос. акад. наук; Ин-т вод. и экол. проблем Сиб. отд. Рос. акад. наук. Сер. Экология. Вып. 88. Новосибирск: ГПНТБ СО РАН. 69 с.
Крылов А.В., Солонго Д., Мэндсайхан Б. 2014. Зоопланктон Дургунского и Тайширского водохранилищ (Западная Монголия) // Аридные экосистемы. Т. 20. № 2(59). С. 48.
Кудерский Л.А. 1992. Экологические основы формирования и использования рыбных ресурсов водохранилищ: Автореф. дис. … докт. биол. наук. Москва: Моск. гос. университет. 85 с.
Луферова Л.А. 1963. Формирование зоопланктона Горьковского водохранилища // Тр. Инст. биол. внутр. вод. Вып. 6(9). Москва: Изд-во АН СССР. С. 131.
Методика изучения биогеоценозов внутренних водоемов. 1975. Москва: Наука.
Монаков А.В. 1958. Зоопланктон волжского устьевого участка Рыбинского водохранилища за период 1947–1954 гг. // Тр. биол. ст. “Борок”. Вып. 3. С. 214.
Углублeнный обзор политики и программ в области энергоэффективности: Монголия. 2011. Секретариат Энергетической Хартии // Boulevard de la Woluwe. Т. 56. B-1200. Брюссель. 132 с.
Шаронов И.В. 1966. Формирование ихтиофауны водохранилищ // Экология водных организмов. Москва: Наука, С. 103.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод