

Биология внутренних вод, 2019, № 4-2, стр. 27-35
Признаки естественного эвтрофирования мелководного оз. Неро по осадочным пигментам
Л. Е. Сигарева a, *, Н. А. Тимофеева a, В. В. Законнов a, А. Г. Русанов b, Н. В. Игнатьева b, Ш. Р. Поздняков b
a Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
Некоузский р-н, Ярославская обл., пос. Борок, Россия
b Институт озероведения Российской академии наук
Санкт-Петербург, Россия
* E-mail: sigareva@ibiw.yaroslavl.ru
Поступила в редакцию 25.07.2018
После доработки 14.09.2018
Принята к публикации 27.11.2018
Аннотация
Проанализированы данные спектрофотометрического определения растительных пигментов в донных отложениях оз. Неро по материалам съемки 2017 г. Установлена низкая вариабельность (СV = 27–31%) содержания хлорофилла а и феопигментов по площади дна и по вертикали в кернах сапропеля. Значения концентраций пигментов в донных отложениях озера относятся к гипертрофной категории. Высокопродуктивный статус озера по содержанию пигментов (>300 мкг/г сухого грунта) сохраняется для периода формирования 80-сантиметрового слоя сапропеля (~800 лет). Выявлены тренды увеличения концентрации растительных пигментов от нижних слоев кернов к верхним при достоверной связи с вкладом органического вещества и общего азота, что свидетельствует о естественном эвтрофировании озера.
ВВЕДЕНИЕ
Потенциальные запасы сапропеля в РФ оценивают в 250 млрд м3 (Борисов и др., 2014). Интерес к изучению мелководных сапропелевых озер многогранен, поскольку их экосистемы находятся на конечном этапе олиготрофно-эвтрофной сукцессии и могут служить объектами для теоретической экологии, в том числе для решения вопросов о механизмах и типах эвтрофирования – естественного и антропогенного (Мальцев и др., 2014; Хендерсон-Селлерс, Маркленд, 1990; Gebrehiwot et al., 2017; Sanchez et al., 2017; Søndergaard et al., 2003; Stankevica et al., 2014; Strakhovenko et al., 2014; Yermolaeva et al., 2016). Практическое значение таких озер обусловлено возможностями использования сапропеля в сельском хозяйстве из-за высокой концентрации азота, а также в медицине для получения природных антибиотиков – гидрокортизона, преднизолона и др. (Борисов и др., 2014; Платонов и др., 2014; Kurzo et al., 2017).
Озеро Неро (57°10″ с.ш., 39°26″ в.д.) – уникальный водоем, один из наиболее продуктивных в Европе. Трофическое состояние озера по содержанию растительных пигментов в планктоне и донных отложениях – гипертрофное (Состояние…, 2008). Озеро подвергается комплексному воздействию природных и антропогенных факторов. В настоящее время интерес к оз. Неро возрос в связи с планированием его интенсивного использования в разных целях – от рекреации до добычи сапропеля. Однако имеющаяся база научных данных для решения практических задач недостаточна. Один из главных вопросов, от которого зависит характер водоохранных мероприятий, – эвтрофирование озера. Изучение эвтрофирования водоемов до сих пор представляет определенные трудности, поскольку имеются разногласия по этой проблеме как методические, так и методологические (Даценко, 2007; Bul’on, 1998; Datsenko, 1998; Eutrophication…, 1982). Есть мнение, что эвтрофирование должно рассматриваться только как антропогенное при полном отрицании естественного (Bul’on, 1998). Один из адекватных подходов к изучению эвтрофирования – анализ пространственно-временной динамики пигментных характеристик донных отложений (Guilizzoni et al., 1983; Сигарева, 2012; Sigareva et al., 2013; Sigareva, 2014).
Цель работы – выявить признаки естественного эвтрофирования оз. Неро по многолетним трендам характеристик продуктивности экосистемы: содержанию в донных отложениях растительных пигментов, органического вещества и биогенных элементов.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Станции наблюдений располагались по продольному разрезу озера от устья р. Сара до истока р. Векса, а также по периметру прибрежных участков, включающих места впадения ручьев и стоков г. Ростов, в зарослях макрофитов и на открытой акватории (рис. 1). Пробы донных отложений отбирали из верхнего 10-сантиметрового слоя дночерпателем Экмана–Берджа на ст. 1–16. Керны длиной 40 и 45 см (ст. 2, 10, 12) отбирали гравитационной трубкой из полихлорвинила, 70 и 80 см (ст. 4, 5, 7, 15) – 1-метровым батометром из органического стекла с помощью аквалангиста. Короткие керны делили на слои толщиной 5 см, длинные – 10 см. Образцы донных отложений хранили в холодильнике до анализа.
Гранулометрический состав отложений определяли с применением электромагнитной просеивающей установки “Analysette-3” фирмы Alfred Fritsch & CO (Германия) методом мокрого рассева. Отдельные размерные фракции сапропеля смывали с сит и высушивали на водяной бане. Выделяли фракции с диаметрами частиц >1, 0.5–1.0, 0.2–0.5, 0.1–0.2, 0.05–0.1, 0.01–0.05 и <0.01 мм, на основе которых рассчитывали средний диаметр. Объемную массу грунта, естественную влажность, ОВ определяли традиционными методами (Аринушкина, 1970; Буторин и др., 1975), общий азот – в соответствии с ГОСТ 26107-84 (ГОСТ…, 1984), общий фосфор – по методу Мета в модификации Мартыновой и Шмидеберг (1983), содержание Хл и Ф – спектрофотометрическим методом в 90%-ном ацетоновом экстракте (Сигарева, 2012; Lorenzen, 1967). Соотношение каротиноидов и Хл оценивали, исходя из D экстрактов в максимумах поглощения этих пигментов (480 и 665 нм) без поправки (D480/D665) и с поправкой (D480/1.7D665к) на дериваты Хл.
Вычисляли средние значения показателей и их ошибки, коэффициенты вариации и корреляции; достоверность различий между рядами параметров определяли по критерию Стьюдента при p = 0.05.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Основа изучения эвтрофирования озера – оценка современных продукционных показателей трофического статуса по материалам пространственного распределения по ложу водоема. Источниками растительных пигментов в донных отложениях оз. Неро служат в основном фитопланктон, макрофиты и эпифитон, а на участках с глубинами в пределах фотосинтезирующей зоны (менее трех прозрачностей) – микрофитобентос. Бентосные формы водорослей развиваются на сапропеле и остатках отмерших макрофитов. Только близ устья р. Сара (ст. 1) сапропель содержал включения из мелкого песка. Гранулометрический состав, высокая влажность и низкая воздушно-сухая объемная масса отложений соответствуют водно-физическим свойствам сапропеля (табл. 1). Уровень содержания органического вещества (11–40%), общего азота (0.5–1.4%) и общего фосфора (0.2–1.9 мг/г сухого грунта) характеризует сапропель как биологически активный. Содержание Хл в сумме с Ф (Хл + Ф) в поверхностном слое сапропеля изменяется по станциям от 42 до 529 мкг/г сухого грунта. Большая часть пигментов (78–99%) состоит из дериватов Хл (табл. 2). Минимальная концентрация Хл + Ф отмечена в р. Сара (ст. 1), недалеко от места впадения в озеро, максимальная – на зарастающей макрофитами ст. 10а. Среднее значение Хл + Ф для всех станций озера (313 ± 20 мкг/г сухого грунта, 1.09 ± 0.07 мг/г ОВ) близко к уровню величин, полученных в предшествующий период (2000, 2002–2004 гг.) (Состояние…, 2008) и значительно выше, чем в других водоемах верхневолжского бассейна (Сигарева, 2012). Концентрация осадочных пигментов на речной ст. 1 относится к мезотрофной категории, все другие озерные участки – типично гипертрофные (Möller, Scharf, 1986). Такие данные согласуются с высокими среднегодовыми концентрациями Хл в планктоне (33–97 мкг/л) (Состояние…, 2008).
Таблица 1.
Характеристика верхнего (0–10 см) слоя донных отложений на станциях оз. Неро в июне 2017 г.
| Номер cтанции | Глубина, м |
Прозрач- ность, м |
Средний диаметр частиц, мм |
Влаж- ность, % |
Воздушно-сухая объемная масса, г/см3 | ОВ, % | Nобщ, % | Робщ, мг/г сухого грунта |
|---|---|---|---|---|---|---|---|---|
| 1 | 2.6 | 1.1 | 0.21 | 56.4 | 0.60 | 11.4 | 0.5 | 1.91 |
| 2 | 1.5 | 0.6 | 0.18 | 86.9 | 0.14 | 25.3 | 0.9 | 0.54 |
| 3 | 2.0 | 0.3 | 0.16 | 91.6 | 0.09 | 31.6 | 1.19 | 1.06 |
| 4 | 4.0 | 0.3 | 0.14 | 91.0 | 0.10 | 29.5 | 1.15 | 1.81 |
| 5 | 1.65 | 0.3 | 0.14 | 90.3 | 0.10 | 27.3 | 1.16 | 1.0 |
| 6 | 1.4 | 0.2 | 0.09 | 90.9 | 0.10 | 30.2 | 1.17 | 0.58 |
| 6а* | 0.4 | 0.3 | 0.16 | 92.1 | 0.08 | 37.9 | 1.31 | 0.84 |
| 7 | 1.5 | 0.3 | 0.08 | 90.8 | 0.10 | 28.8 | 1.05 | 1.0 |
| 8 | 1.6 | 0.3 | 0.12 | 88.0 | 0.13 | 29.6 | 1.05 | 0.58 |
| 9 | 1.3 | 0.3 | 0.14 | 86.4 | 0.15 | 23.5 | 0.85 | 0.65 |
| 9а* | 0.5 | 0.5 | 0.20 | 88.0 | 0.13 | 11.4 | 1.11 | 0.56 |
| 10 | 1.5 | 0.5 | 0.13 | 88.7 | 0.12 | 27.6 | 1.01 | 0.58 |
| 10а* | 1.0 | 0.5 | 0.13 | 93.5 | 0.07 | 34.6 | 1.32 | 1.2 |
| 11 | 1.7 | 1.5 | 0.23 | 91.3 | 0.09 | 37.5 | 1.31 | 0.24 |
| 12* | 1.3 | 0.5 | 0.19 | 92.3 | 0.08 | 38.7 | 1.36 | 0.3 |
| 12а* | 1.2 | 0.4 | 0.23 | 91.1 | 0.09 | 32.5 | 1.13 | 0.62 |
| 12б* | 1.2 | 0.45 | 0.21 | 93.2 | 0.07 | 39.5 | 1.3 | 0.92 |
| 13 | 1.8 | 0.6 | 0.14 | 89.2 | 0.12 | 25.4 | 0.95 | 0.7 |
| 14 | 1.5 | 0.3 | 0.14 | 89.8 | 0.11 | 28 | 0.96 | 0.64 |
| 15 | 2.0 | 0.3 | 0.10 | 90.6 | 0.10 | 27.5 | 1.1 | 0.7 |
| 16 | 1.7 | 0.35 | 0.13 | 89.0 | 0.12 | 25.0 | 0.97 | 0.35 |
Примечание. * Станции с зарослями макрофитов: 6а – рдест плавающий (Potamogeton natans L.), рдест пронзеннолистный (P. perfoliatus L.), тростник южный (Phragmites australis (Cav.) Trin. ex Steud.), 9а – кубышка желтая (Nuphar lutea (L.) Smith), 10а – рогоз узколистный (Typha angustifolia L.), тростник, 12, 12а – камыш озерный (Scirpus lacustris L.), 12б – тростник, рогоз узколистный. На ст. 1 – сапропель с тонким песком, на остальных – сапропель или сапропель с остатками растений. Здесь и в табл. 4 Nобщ, Робщ – концентрация общего азота и фосфора, соответственно.
Таблица 2.
Среднее содержание растительных пигментов в донных отложениях на станциях, сгруппированных по разным признакам, в оз. Неро (июнь 2017 г., слой 0–10 см)
| Варианты* | ОВ, % | Хл + Ф | Доля Ф в (Хл + Ф), % | D480/D665 | D480/1.7D665к | ||
|---|---|---|---|---|---|---|---|
| мкг/г сухого грунта | мг/г ОВ | мг/(м2 · мм) | |||||
| I | 11.4 | 42.4 | 0.37 | 25.5 | 78.1 | 2.40 | 1.63 |
| II | 28.2 ± 0.9 | 277.1 ± 16.2 | 0.98 ± 0.04 | 30.0 ± 0.7 | 89.9 ± 1.1 | 3.9 ± 0.1 | 2.5 ± 0.1 |
| III | 32.4 ± 4.3 | 397.1 ± 41.8 | 1.34 ± 0.19 | 33.3 ± 1.3 | 85.6 ± 1.5 | 3.4 ± 0.1 | 2.2 ± 0.1 |
| IV | 31.0 ± 2.8 | 347.8 ± 34.6 | 1.18 ± 0.13 | 32.4 ± 1.0 | 87.1 ± 1.2 | 3.5 ± 0.1 | 2.2 ± 0.1 |
| V | 27.8 ± 0.8 | 278.3 ± 17.8 | 1.00 ± 0.05 | 29.5 ± 0.8 | 90.2 ± 1.5 | 4.0 ± 0.1 | 2.5 ± 0.1 |
| VI | 29.4 ± 1.4 | 313.1 ± 20.6 | 1.09 ± 0.07 | 31.0 ± 0.7 | 88.7 ± 1.0 | 3.8 ± 0.1 | 2.4 ± 0.1 |
Примечание. * I – р. Сара (ст. 1), II – открытая часть без зарослей (ст. 2–11, 13–16), III – с зарослями макрофитов (ст. 6а, 9а, 10а, 12, 12а, 12б), IV – глубины менее трехкратной прозрачности (ст. 2, 6а, 9а, 10, 10а, 11, 12, 12а, 12б, 13), V – глубины более трехкратной прозрачности (ст. 3–9, 14–16), VI – все станции озера.
Зарастающие макрофитами участки выделяются более высоким содержанием растительных пигментов в сапропеле (от 250 до 529 мкг/г сухого грунта), чем участки без макрофитов (от 190 до 397) (табл. 2). Средняя для открытых станций концентрация Хл + Ф в сапропеле – 277 ± 16, для зарастающих участков в ~1.5 раза больше – 397 ± ± 42 мкг/г сухого грунта. Следовательно, высшая водная растительность обогащает сапропель растительными пигментами и поэтому влияет на трофические показатели дна. Средняя концентрация пигментов Хл + Ф на станциях с вероятным развитием микрофитобентоса (348 ± 35 мкг/г) была достоверно больше (p < 0.05), чем на остальных станциях (278 ± 18 мкг/г).
Показатели деградации пигментного фонда (процентная доля Ф в Хл + Ф, соотношение каротиноидов и Хл, оцененное по индексам D480/D665 и D480/1.7D665к, свидетельствуют о том, что растительные пигменты в донных отложениях находятся в сильно деградированном виде (табл. 2). Высокая степень деградации фотосинтетических пигментов – итог утилизации и трансформации первичной продукции органического вещества в экосистеме.
В 2017 г. среднее содержание Хл + Ф в верхнем слое донных отложений (0–10 см) было сопоставимым с таковым в предыдущие годы наблюдений при наиболее часто встречаемых значениях в пределах 200–400 мкг/г сухого грунта. Концентрации Хл + Ф в расчете на 1 м2 и толщину слоя 1 мм грунта при естественной влажности в июне 2017 г. достигали 31.0 ± 0.72, в августе 2017 г. – 32.7 ± ± 3.6 мг/(м2 · мм), что также входит в диапазон значений, полученных ранее (28.4–58.2 мг/(м2 · мм)) (Состояние…, 2008). Сходство средних концентраций во все годы наблюдений свидетельствует о сохранении высокого уровня продуктивности первичного звена в озере.
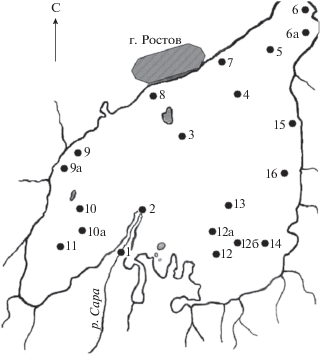
Вертикальное распределение содержания растительных пигментов в донных отложениях изучают для восстановления исторических изменений продуктивности водоемов (Guilizzoni et al., 1983). Результаты нашей работы показали, что гипертрофный статус характерен для оз. Неро продолжительное время. Исходя из средней скорости осадконакопления (1 мм/год), возраст донных отложений в колонках 40–80 см соответствует ~400–800 годам. Различия между минимальной и максимальной величинами Хл + Ф в кернах на отдельных станциях варьируют в 2–3 раза (рис. 2). Абсолютный максимум достигает 704 мкг/г. Наибольшую по вертикали величину не всегда отмечали в верхнем слое (рис. 2), что может быть связано с неравномерным накоплением ОВ в донных отложениях разнотипных участков. В целом значения средних концентраций Хл + Ф в коротких колонках (40–45 см) были выше (280–523 мкг/г сухого осадка), чем в длинных (70–80 см) – 261–341. Послойное осреднение концентраций Хл + Ф для всех кернов одного горизонтального уровня позволило получить единый вертикальный профиль, характеризующийся положительным трендом изменений от нижнего слоя к верхнему (рис. 3). Во многих случаях различия концентраций пигментов в слоях, отражающие возрастание первичной продукции в многолетнем аспекте, статистически достоверны (табл. 3).
Рис. 2.
Вертикальное распределение Хл (1) и Ф (2) (мкг/г сухого грунта) на разных глубинах (см) донных отложений оз. Неро: а, б, в, г, д, е, ж – ст. 4, 5, 7, 15, 2, 10, 12 соответственно.

Рис. 3.
Среднее (± стандартная ошибка) содержание и линия тренда Хл + Ф (мкг/г) в слоях кернов сапропеля оз. Неро в 2017 г. Коэффициент связи (R2) Хл + + Ф с глубиной донных отложений равен 0.82.
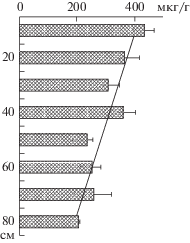
Таблица 3.
Оценка достоверности различий между концентрациями растительных пигментов (Хл + Ф) в слоях кернов по t-критерию Стьюдента
| Сравниваемые слои |
t-Стьюдента | Реальное значение | p, % |
|---|---|---|---|
| 1 и 2 | 2.18 | –1.08 | – |
| 1 и 3 | 2.18 | –2.35 | <5 |
| 1 и 4 | 2.18 | –1.28 | – |
| 1 и 5 | 2.26 | –4.85 | <0.1 |
| 1 и 6 | 2.26 | –3.81 | <1 |
| 1 и 7 | 2.26 | –2.46 | <5 |
| 1 и 8 | 2.37 | –6.18 | <0.1 |
| 2 и 3 | 2.18 | –0.94 | – |
| 3 и 4 | 2.18 | 0.93 | – |
| 4 и 5 | 2.26 | –2.71 | <5 |
| 5 и 6 | 2.45 | 0.43 | – |
| 6 и 7 | 2.45 | 0.04 | – |
| 7 и 8 | 2.78 | –0.77 | – |
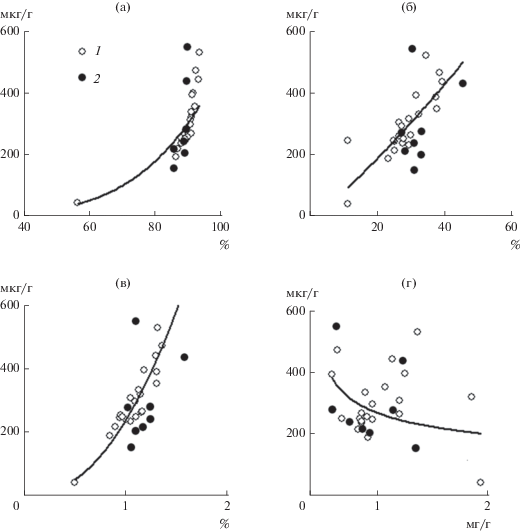
Одним из доказательств индикаторной значимости осадочных пигментов как продукционных показателей озера могут быть тесные зависимости пигментов с концентрациями органического вещества и биогенных элементов в донных отложениях. Корреляционный анализ показал, что концентрации Хл, Ф и Хл + Ф достоверно связаны с содержанием ОВ и общего азота (табл. 4), что отмечалось как по данным горизонтального распределения концентраций по площади дна, так и по вертикальному профилю донных отложений. Связь растительных пигментов с характеристиками донных отложений (за исключением общего фосфора) – степенная с высокими коэффициентами детерминации (рис. 4), это согласуется с соответствующими показателями, полученными для озер и водохранилищ (Timofeeva, Sigareva, 2004).
Таблица 4.
Коэффициенты линейной корреляции ОВ и суммы (Хл + Ф) с другими пигментными характеристиками, концентрациями общего азота и фосфора для верхнего слоя донных отложений (0–10 см) на станциях и в кернах (0–70 см)
| Показатели | 0–10 см, r0.05 = 0.43 | 0–70 см, r0.05 = 0.37 | ||
|---|---|---|---|---|
| ОВ, % | Хл + Ф | ОВ, % | Хл + Ф | |
| Хл, мкг/г | 0.57 | 0.76 | 0.38 | 0.72 |
| Ф, мкг/г | 0.81 | 0.99 | 0.67 | 0.99 |
| Хл + Ф, мкг/г | 0.81 | 1.00 | 0.66 | 1.00 |
| D480/D665 | 0.04 | 0.00 | –0.10 | –0.18 |
| D480/1.7D665к | 0.02 | 0.00 | –0.15 | –0.17 |
| Ф, % | 0.17 | 0.08 | 0.23 | –0.02 |
| Хл, % | –0.17 | –0.08 | –0.23 | 0.02 |
| Влажность грунта, % | 0.67 | 0.72 | 0.60 | 0.63 |
| Воздушно-сухая объемная масса, г/см3 | –0.64 | –0.69 | –0.58 | –0.59 |
| Хл + Ф, мг/(м2 · мм) | 0.52 | 0.79 | 0.37 | 0.75 |
| Nобщ, % | 0.79 | 0.91 | 0.82 | 0.72 |
| Pобщ, мг/г | –0.27 | –0.17 | –0.20 | –0.19 |
Рис. 4.
Связи содержания Хл + Ф с влажностью (а), c содержанием ОВ (б), общего азота (в) и общего фосфора (г). 1 – слой 0–10 см, 2 – 30–70 см. Коэффициенты детерминации (R2) для верхнего слоя составили 0.86, 0.61, 0.92 и 0.10 для а, б, в, г соответственно.
Коэффициенты вариации содержания Хл + Ф в донных отложениях сравнительно невысокие (2–50%) и в среднем составляют по горизонтальному распределению 29%, по вертикали – 31%, по отдельным слоям кернов – 27% (табл. 5). Такие величины заметно ниже коэффициентов вариации содержания осадочных пигментов в менее продуктивных искусственных водоемах – водохранилищах, где они достигают 200% (Сигарева, 2012). Даже в высокопродуктивном Влоцлавском водохранилище (на р. Висла) изменчивость концентраций осадочных пигментов выражена сильнее (57% по вертикали и 79% по площади дна) (Сигарева и др., 2017), чем в оз. Неро. Невысокие коэффициенты варьирования концентраций пигментов в донных отложениях озера – признак однородности абиотических условий и сходства продуктивности на отдельных участках водоема.
Таблица 5.
Коэффициенты вариации (%) показателей содержания растительных пигментов в донных отложениях оз. Неро
| Варианты | Хл + Ф, мкг/г | Хл + Ф, мг/(м2 · мм) | Ф от (Хл + Ф), % | D480/D665 |
|---|---|---|---|---|
| По площади дна (0–10 см) | ||||
| Все станции (1–16) | 29.4 | 10.4 | 5.0 | 10.3 |
| без макрофитов | 21.8 | 9.2 | 4.6 | 7.5 |
| с макрофитами | 25.8 | 9.7 | 4.4 | 9.5 |
| По вертикали в кернах | ||||
| Ст. 2, 0–40 см | 43.3 | 24.7 | 7.2 | 10.5 |
| Ст. 4, 0–80 | 36.6 | 6.5 | 4.7 | 7.4 |
| Ст. 5, 0–70 | 30.1 | 22.3 | 5.7 | 8.5 |
| Ст. 7, 0–80 | 23.4 | 26.2 | 3.5 | 7.2 |
| Ст. 10, 0–45 | 18.1 | 11.6 | 5.9 | 8.6 |
| Ст. 12, 0–40 | 19.1 | 21.8 | 4.8 | 4.8 |
| Ст. 15, 0–70 | 47.0 | 37.6 | 8.5 | 16.7 |
| По слоям кернов (ст. 2, 4, 5, 7, 10, 12, 15)* | ||||
| Толщина слоя, см | ||||
| 0–10 | 22.6 | 11.7 | 7.0 | 10 |
| 10–20 | 36.1 | 16.7 | 7.2 | 10.2 |
| 20–30 | 33.8 | 26.3 | 8.8 | 14.4 |
| 30–40 | 31.6 | 30.6 | 4.6 | 14.9 |
| 40–50 | 14.6 | 22.1 | 2.6 | 7.6 |
| 50–60 | 24.5 | 32.5 | 3.6 | 10.3 |
| 60–70 | 50.1 | 36.4 | 4.2 | 11.3 |
| 70–80 | 1.6 | 25.8 | 5.1 | 0.8 |
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Эвтрофирование – ключевой природный процесс, приводящий к сукцессии сообществ и всей экосистемы (Лазарева, 2010; Трифонова, 1986; Alimov, Golubkov, 2014). Однако, выявить признаки естественного эвтрофирования сложно, поскольку этот процесс имеет низкую скорость и сочетается с антропогенным эвтрофированием и цикличностью, обусловленной периодичностью климатических факторов (Даценко, 2007; Пырина и др., 2006; Alimov, Golubkov, 2014; Datsenko, 1998; Izmest’eva et al., 2011). В зависимости от абиотических условий обычно отмечают чередование увеличения и уменьшения продуктивности организмов первичного звена и других сообществ. При этом изменчивость интенсивности развития звеньев трофической цепи иногда необоснованно называют эвтрофированием и деэвтрофированием. Считается, что основные факторы эвтрофирования, приводящего к сукцессии сообществ и экосистемы в целом, – тип ландшафта и генезис территории (Черевичко, 2017). В то же время, выявлена эвтрофирующая роль внутренней биогенной нагрузки и гидробионтов, которые стимулируют продукционные процессы за счет экскреции фосфора (Alimov, Golubkov, 2014).
Рассмотреть вопрос о естественном эвтрофировании как о процессе, характеризующемся возрастанием интенсивности первичного продуцирования, можно лишь в геологических масштабах времени. Единственный подход, позволяющий проследить признаки естественного эвтрофирования – изучение вертикального профиля палеоиндикаторов в донных отложениях. В этом аспекте большое значение имеют растительные пигменты как показатели реализованной продукции органического вещества, в отличие от концентрации биогенных элементов – показателей потенциальной продуктивности. Количественные зависимости осадочных пигментов от концентрации Хл в воде и первичной продукции фитопланктона свидетельствуют о связи трофии ярусов экосистемы – бентали и пелагиали (Guilizzoni et al., 1983; Sigareva et al., 2013; Сигарева, 2012; Sigareva, Timofeeva, 2014; Cremona et al., 2016). При эвтрофировании наблюдается повышение накопления ОВ на дне, приводящее к преобладанию илов в грунтовом комплексе высокопродуктивных водоемов, что показано на примере озер и водохранилищ Волжского бассейна (Сигарева, 2012).
Материалы настоящей работы, выполненной на оз. Неро, позволили утвердительно ответить на вопрос, имеет ли место естественное эвтрофирование озер. Другие изменения продуктивности оз. Неро, связанные с антропогенным эвтрофированием и динамикой климатических условий, были выявлены ранее по показателям развития фитопланктона и макрофитов (Сиделев и др., 2008; Состояние…, 2008).
Выводы. Полученные результаты по осадочным пигментам в оз. Неро позволили выявить ряд признаков естественного эвтрофирования. Содержание растительных пигментов в донных отложениях незначительно варьирует во времени и пространстве и характеризует озеро как гипертрофное. По концентрации Хл + Ф в донных отложениях оз. Неро продолжает занимать лидирующее положение среди изученных водоемов Верхне-Волжского бассейна. Экосистема озера по-прежнему находится на последней стадии олиготрофно-эвтрофной сукцессии, после которой следует заболачивание. Длительный период последнего этапа сукцессии озера и положительный тренд к настоящему времени свидетельствуют о преимущественном влиянии на продуктивность естественных природных факторов, а не антропогенных. Оценка состояния продукционных свойств водоема и выявление типа эвтрофирования имеет особое значение, поскольку от этого зависит характер водоохранных мероприятий, направленных на оздоровление озера на ближайшее столетие.
Список литературы
Аринушкина Е.В. 1970. Руководство по химическому анализу почв. Москва: МГУ.
Борисов В.А., Успенская О.Н., Гренадеров Н.В., Васючков И.Ю. 2014. Состав органического вещества разных видов сапропелей // Агрохимия. № 8. С. 51.
Буторин Н.В., Зиминова Н.А., Курдин В.П. 1975. Донные отложения верхневолжских водохранилищ. Ленинград: Наука.
ГОСТ 26107-84. 1984. Почвы. Методы определения общего азота.
Даценко Ю.С. 2007. Эвтрофирование водохранилищ. Гидролого-гидрохимические аспекты. Москва: ГЕОС.
Лазарева В.И. 2010. Структура и динамика зоопланктона Рыбинского водохранилища. Москва: Товарищество науч. изданий КМК.
Мальцев А.Е., Леонова Г.А., Бобров В.А., Кривоногов С.К. 2014. Геохимические особенности голоценового разреза сапропеля озера Минзелинское (Западная Сибирь) // Известия Томского политехнического университета. Т. 325. № 1. С. 83.
Мартынова М.В., Шмидеберг Н.А. 1983. О методах определения различных форм фосфора в донных наносах // Гидрохимические материалы. Т. 85. С. 49.
Платонов В.А., Хадарцев А.А., Фридзон К.Я., Чуносов С.Н. 2014. Химический состав и биологическая активность сапропеля оз. Глубокое (Татарстан) // Вестник новых медицинских технологий. Т. 21. № 3. С. 199.
Пырина И.Л., Литвинов А.С., Кучай Л.А. и др. 2006. Многолетние изменения первичной продукции фитопланктона Рыбинского водохранилища в связи с действием климатических факторов // Состояние и проблемы продукционной гидробиологии. Москва: Товарищество науч. изданий КМК. С. 36.
Сигарева Л.Е. 2012. Хлорофилл в донных отложениях волжских водоемов. Москва: Товарищество науч. изданий КМК.
Сигарева Л.Е., Тимофеева Н.А., Законнов В.В., Гершевский П. 2017. Оценка вариабельности трофического состояния Влоцлавского водохранилища по осадочным пигментам // Вода: химия и экология. № 2. С. 3.
Сиделев С.И., Бабаназарова О.В., Рычкова Е.Н. 2008. Многолетние изменения концентраций хлорофилла а в мелководном высокоэвтрофном озере // Водные экосистемы: трофические уровни и проблемы поддержания биоразнообразия. Вологда: Центр оперативной полиграфии “Коперник”. С. 102.
Состояние экосистемы озера Неро в начале XXI века. 2008. Москва: Наука.
Трифонова И.С. 1986. Сезонная и основная сукцессия озерного фитопланктона // Гидробиол. ж. Т. 22. № 3. С. 21.
Хендерсон-Селлерс Б., Маркленд Х. 1990. Умирающие озера. Причины и контроль антропогенного эвтрофирования. Ленинград: Гидрометеоиздат.
Черевичко А.В. 2017. Сукцессия зоопланктона заболоченных озер и болот озерного происхождения // Труды Института биологии внутренних вод РАН. № 79(82). С. 278.
Alimov A.F., Golubkov M.S. 2014. Lake eutrophication and community structure // Inland Water Biology. V. 7. № 3. P. 185. https://doi.org/10.1134/S1995082914030031
Bul’on V.V. 1998. Does natural eutrophication of lakes occur? // Water Resources. V. 25. № 6. P. 701.
Cremona F., Laas A., Arvola L. et al. 2016. Numerical exploration of the planktonic to benthic primary production ratios in lakes of the Baltic Sea catchment // Ecosystems. V. 19. P. 1386. https://doi.org/10.1007/s10021-016-0006-y
Datsenko Yu.S. 1998. Comment on “Does natural eutrophication of lakes occur?” by V.V. Bul’on // Water Resources. V. 25. № 6. P. 707.
Eutrophication of waters. Monitoring, assessment and control. 1982. Paris: OECD.
Gebrehiwot M., Kifle D., Stiers I., Triest L. 2017. Phytoplankton functional dynamics in a shallow polymictic tropical lake: the influence of emergent macrophytes // Hydrobiologia. V. 797. № 1. P. 69. https://doi.org/10.1007/s10750-017-3161-z
Guilizzoni P., Bonomi G., Galanti G., Ruggiu D. 1983. Relationship between sedimentary pigments and primary production; evidence from core analyses of twelve Italian lakes // Hydrobiologia. V. 103. № 1. P. 103. https://doi.org/10.1007/BF00028436
Izmest’eva L.R., Silow E.A., Litchman E. 2011. Long-term dynamics of lake Baikal pelagic phytoplankton under climate change // Inland Water Biology. V. 4. № 3. P. 301. https://doi.org/10.1134/S1995082911030102
Kurzo B.V., Navosha Yu.Yu., Strigutskii V.P. 2017. Formation of sapropel in the lakes of Belarus // Solid Fuel Chem. V. 51. № 5. P. 326. https://doi.org/10.3103/S0361521917050032
Lorenzen C.J. 1967. Determination of chlorophyll and phaeo-pigments: spectrophotometric equations // Limnol., Oceanogr. V. 12. № 2. P. 343. https://doi.org/10.4319/lo.1967.12.2.0343
Möller W.A.A., Scharf B.W. 1986. The content of chlorophyll in the sediment of the volcanic maar lakes in the Eifel region (Germany) as an indicator for eutrophication // Hydrobiologia. V. 143. № 1. P. 327. https://doi.org/10.1007/BF00026678
Sánchez M.L., Rodríguez P., Torremorell A.M. et al. 2017. Phytoplankton and periphyton primary production in clear and turbid shallow lakes: Influence of the light environment on the interactions between these communities // Wetlands. V. 37. № 1. P. 67. https://doi.org/10.1007/s13157-016-0840-x
Sigareva L.E., Timofeeva N.A. 2014. The phytoplankton role in formation of bottom sediment productivity in a large reservoir in the years with different temperature conditions // Phytoplankton: biology, classification and environmental impacts. New York: Nova Sci. Ch. 6. P. 151.
Sigareva L.E., Zakonnov V.V., Timofeeva N.A., Kas’yanova V.V. 2013. Sediment pigments and silting rate as indicators of the trophic condition of the Rybinsk reservoir // Water Resources. V. 40. № 1. P. 54. https://doi.org/10.1134/S0097807813010090
Søndergaard M., Jensen J.P., Jeppesen E. 2003. Role of sediment and internal loading of phosphorus in shallow lakes // Hydrobiologia. V. 506. № 1–3. P. 135. https://doi.org/10.1023/B:HYDR.0000008611.12704.dd
Stankevica K., Klavins M., Rutina L., Cerina A. 2014. Lake Sapropel: a valuable resource and indicator of lake development // Advances in Environment, Computational Chemistry and BioScience. Montreux: WSEAS Press. P. 247.
Strakhovenko V.D., Taran O.P., Ermolaeva N.I. 2014. Geochemical characteristics of the sapropel sediments of small lakes in the Ob’-Irtysh interfluve // Russian Geology and Geophysics. V. 55. № 10. P. 1160. https://doi.org/10.1016/j.rgg.2014.09.002
Timofeeva N.A., Sigareva L.E. 2004. Relationships between the concentrations of phyto-pigments, nitrogen, and phosphorus in bottom deposits of water reservoirs // Water Resources. V. 31. № 3. P. 303. https://doi.org/10.1023/B:WARE.0000028700.50699.5f
Yermolaeva N.I., Zarubina E.Yu., Romanov R.E. et al. 2016. Hydrobiological conditions of sapropel formation in lakes in the south of Western Siberia // Water Resources. V. 43. № 1. P. 129. https://doi.org/10.1134/S0097807816010073
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод