

Биология внутренних вод, 2020, № 1, стр. 87-93
Погрешность оценки численности поколений популяций рыб при неполной исходной информации
В. Г. Терещенко a, *, В. А. Кузнецов b, Ф. М. Шакирова c, Л. И. Терещенко a
a Институт биологии внутренних вод им. И.Д. Папанина РАН
Некоузcкий р-н, Ярославская обл., пос. Борок, Россия
b Казанский государственный (федеральный) университет
Казань, Россия
Ñ Всероссийский научно-исследовательский институт рыбного хозяйства и океанографии, ТатарстанНИРО
Казань, Россия
* E-mail: tervlad@ibiw.yaroslavl.ru
Поступила в редакцию 29.06.2018
После доработки 23.10.2018
Принята к публикации 27.11.2018
Аннотация
Проведен анализ влияния объема отсутствующей информации о возрастной структуре рыб на погрешность оценки относительной численности поколений их популяций. На примере леща Куйбышевского водохранилища показано, что при потере информации за 1–8 лет погрешность оценки численности поколений в среднем 5–15%. Это позволяет применять данную модификацию метода убыли от лова для оценки численности поколений популяций. С помощью предложенного метода можно анализировать популяции не только рыб, но и других групп гидробионтов.
ВВЕДЕНИЕ
Анализ состояния и изменений в популяциях животных основан на различных подходах, в том числе и оценке численности их поколений. Для этого в ихтиологии широко применяется метод убыли от лова (Монастырский, 1952; Никольский, 1974; Рикер, 1979). Во всех вариантах данного метода численность поколений рассчитывают на основании непрерывных многолетних данных о возрастной структуре исследуемой популяции. Изменение экономической ситуации в России в начале 90-х гг. XX в. привело к тому, что за ряд лет данные наблюдений за состоянием популяций отсутствуют. Вместе с тем, увеличение антропогенной нагрузки на пресноводные экосистемы оказало существенное влияние на популяции многих видов рыб (Стрельников, 1996). В создавшихся условиях возросла важность информации о численности поколений как массовых, так и охраняемых видов. Ранее была предложена и апробирована модификация биостатистического метода (убыли от лова), пригодная для оценки относительной численности поколений популяций рыб в условиях неполной исходной информации (Tereshchenko, Zuyanova, 2006). Она основана на двух положениях. Во-первых, для решения многих задач необходимо знать не столько абсолютную, сколько относительную численность поколения. Поэтому, мы воспользовались предложением Стрельникова (1996) подсчитывать для оценки численности сумму процентов данного поколения в уловах от момента вступления в промысел до фактического предельного их возраста в уловах. Второе положение метода предполагает использование неполной исходной информации. Поскольку при наличии пропусков в исходных данных классический метод убыли от лова не может быть использован, то для оценки численности поколения сравнивали суммарное процентное соотношение данной генерации в уловах с суммарным процентным соотношением этих же возрастных групп в равновесной возрастной структуре, т.е. характерной для постоянных условий среды, когда все поколения данного вида одинаковы по численности.
В ранее вышедшей работе (Tereshchenko, Zuyanova, 2006) не было возможности оценить погрешность метода. Его адекватность проверяли по соответствию результатов расчетов численности поколений популяций леща и судака оз. Воже реальному положению в водоеме. Так, поколения леща, родившиеся в середине 1990-х гг., все были малочисленными, что соответствует низкому уровню воды в эти годы во многих озерах Вологодской обл. Это свидетельствует в пользу адекватности предложенного метода. Сходные результаты дал и прогноз уловов леща в период вступления этих малочисленных поколений в промысел. Оценка относительной численности поколений судака показала, что поколение, родившееся в 1995 г., было наиболее многочисленным, что отмечает в своей работе и Коновалов (2004).
Сейчас, имея в наличии непрерывный ряд возрастной структуры леща за 17 лет, можно на примере этого вида оценить погрешность предложенной модификации метода убыли от лова, что необходимо при любом исследовании, проводимом на количественном уровне (Поддубный, 1982; Reshetnikov, Tereshchenko, 2017). Кроме того, появилась возможность решения задачи влияния объема отсутствующей информации на погрешность оценки относительной численности поколений популяции рыб.
Цель работы – анализ влияния объема отсутствующей информации на погрешность оценки относительной численности поколений популяции рыб.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Материалом для исследования послужили данные возрастной структуры леща Куйбышевского водохранилища в контрольных уловах 1961–1977 гг. (табл. 1). Рыбу отлавливали сетями с размером ячеи 36–90 мм (Анохина и др., 2013; Кузнецов, 1980). Возраст рыб определяли по чешуе, используя стандартную методику (Чугунова, 1959). Метод оценки относительной численности поколений рыб при неполной исходной информации о возрастной структуре популяции был подробно изложен ранее (Tereshchenko, Zuyanova, 2006).
Таблица 1.
Возрастной состав леща контрольных уловов в Куйбышевском водохранилище в 1961–1977 гг.
| Год | Доля (% в уловах) возрастной группы, лет | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | |
| 1961 | 6.3 | 44.0 | 40.4 | 4.0 | 3.0 | 1.5 | 0.8 | 0 | 0 | 0 | 0 |
| 1962 | 1.0 | 7.7 | 35.4 | 38.4 | 8.0 | 6.5 | 3.0 | 0 | 0 | – | – |
| 1963 | 1.3 | 2.6 | 7.9 | 32.7 | 36.7 | 10.9 | 6.2 | 1.7 | – | – | – |
| 1964 | 5.8 | 2.8 | 3.1 | 7.0 | 28.0 | 42.4 | 7.8 | 1.9 | 0.8 | 0.4 | – |
| 1965 | 3.0 | 19.0 | 8.0 | 4.0 | 8.0 | 21.0 | 28.0 | 8.0 | 1.0 | – | – |
| 1966 | 20.3 | 2.8 | 9.9 | 4.4 | 3.8 | 11.8 | 22.2 | 16.8 | 6.3 | 1.7 | – |
| 1967 | 8.5 | 27.1 | 1.1 | 8.5 | 2.3 | 1.1 | 5.1 | 19.7 | 20.4 | 5.6 | 0.6 |
| 1968 | 2.3 | 17.0 | 29.9 | 1.8 | 11.3 | 2.0 | 0.9 | 4.8 | 13.2 | 15.0 | 1.8 |
| 1969 | 23.4 | 6.3 | 16.8 | 23.8 | 1.6 | 5.4 | 2.1 | 1.2 | 2.5 | 8.6 | 8.3 |
| 1970 | 0.6 | 8.4 | 3.7 | 19.2 | 35.3 | 3.7 | 13.4 | 1.6 | 1.8 | 2.8 | 9.5 |
| 1971 | 7.4 | 1.0 | 10.6 | 7.1 | 29.5 | 34.8 | 1.8 | 4.3 | 1.0 | 0.5 | 2.0 |
| 1972 | 6.7 | 1.9 | 0.5 | 25.3 | 9.6 | 24.0 | 28.1 | 1.9 | 1.4 | 0.3 | 0.3 |
| 1973 | 0.7 | 6.9 | 3.2 | 0.7 | 30.9 | 7.928.7 | 23.3 | 23.0 | 1.6 | 1.1 | 0.7 |
| 1974 | 2.0 | 1.5 | 7.1 | 3.0 | 1.5 | 7.5 | 22.0 | 23.7 | 2.0 | 1.0 | |
| 1975 | 1.0 | 2.0 | 4.1 | 7.3 | 3.4 | 2.0 | 22.5 | 8.0 | 21.7 | 27.0 | 1.0 |
| 1976 | 1.6 | 1.8 | 1.3 | 1.6 | 5.7 | 1.3 | 15.6 | 23.7 | 8.5 | 19.5 | 19.4 |
| 1977 | 1.0 | 0.2 | 0.2 | 1.5 | 1.5 | 3.6 | 6.8 | 20.4 | 26.5 | 12.8 | 25.5 |
Для нахождения равновесной возрастной структуры популяции строили объединенную кривую улова (Анохина и др., 2013; Тereshchenko, Zuyanova, 2006) за 1961–1977 гг., которую для упрощения моделирования изменений численности рыб разного возраста переводили в полулогарифмические координаты: по оси ординат – натуральный логарифм процента возрастной группы, по оси абсцисс – возраст.
Необходимо отметить, что на получаемую с помощью контрольных орудий лова возрастную структуру популяции действуют три фактора. Численность младших возрастных групп занижается из-за селективности орудий лова, а обилие старших возрастных групп уменьшается с возрастом из-за естественной и промысловой убыли. И, наконец, наличие высоко- и малочисленных поколений вносят дополнительные искажения в величину относительной численности рыб, поэтому далее полученную объединенную кривую возрастной структуры сглаживали, применяя стандартный регрессионный анализ.
Периоды кусочной аппроксимации обусловлены биологическими причинами. По правилам рыболовства, минимальный размер разрешенной ячеи определяется размером впервые созревающих рыб. И хотя в популяции особей меньшего возраста по численности больше, чем старших, но из-за уменьшения эффективности их поимки в возрастной структуре уловов численность младших возрастных групп меньше, чем в водоеме. Для моделирования действия факторов, ограничивающих численность рыб младшего возраста, применяли полином второй степени. Границы аппроксимации младших возрастных групп – от минимального до времени полового созревания. Для моделирования уменьшения с возрастом обилие старших возрастных групп проводили линейную регрессию.
Точность метода обусловлена погрешностью оценки исследуемого параметра. На погрешность оценки относительной численности поколений влияют как минимум три фактора: аппроксимация данных, неполнота временного ряда и величина выборки, взятой для анализа возрастной структуры. В настоящее время анализ возрастной структуры основан, как правило, на меньшем объеме выборки, чем до начала 1990-х гг. Относительно других факторов нет оснований полагать, что что-то изменилось с прежних времен. Поэтому материалы 1960–1980-х годов могут быть хорошей основой для решения ряда методических вопросов, в том числе и влияния неполноты исходных данных на погрешность оценки численности поколений. Часто для оценки погрешности количественного показателя применяют метод Декарта, т.е. разложение сложной проблемы (оценки погрешности) на более простые составляющие, которые имеют решение (Кондрашов, Шестопалов, 1977; Рабинович, 1978; Reshetnikov, Tereshchenko, 2017).
Авторы воспользовались редкой в практике экологических исследований возможностью найти истинное значение исследуемого параметра (относительную численность поколения). В качестве истинного значения численности поколения брали ее значение при отсутствии пропусков в исходной информации о возрастной структуре популяции. По определению погрешность – это отклонение результата от его истинного значения (Кондрашов, Шестопалов, 1977; Рабинович, 1978), в данном случае – это разность результата оценки численности поколения при неполной исходной информации и его истинного значения.
где rN – погрешность оценки относительной численности поколения, NIncomplate – численность поколения по неполным данным, NReal – численность поколения по полным данным.Для моделирования пропусков 10 раз в исследовании повторяли формирование случайных подвыборок для каждого из числа исключаемых лет (от 1 до 8) в исходной информации по таблице случайных чисел.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Оценка численности поколений рыб на основе модифицированной методики убыли от лова предполагает построение, прежде всего, равновесной возрастной структуры популяции в период исследования. Для этого проводили усреднение данных возрастной структуры леща Куйбышевского водохранилища за 1961–1977 гг. (рис. 1).
Рис. 1.
Усредненная возрастная структура леща Куйбышевского водохранилища в 1961–1977 гг. 1 – исходные данные; 2 – сглаженные данные (равновесная возрастная структура). По оси ординат – натуральный логарифм процента численности возрастной группы.
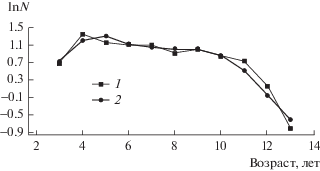
По имеющимся данным, лещ Куйбышевского водохранилища становится половозрелым при длине тела 260–270 мм, что соответствует его возрасту 5–6 лет (Лукин, 1986; Цыплаков, 1972). Поэтому аппроксимацию относительной численности младших возрастных групп (3–6 лет) проводили полиномом второй степени:
где ln N – натуральный логарифм процентного соотношения возрастной группы, T – возраст, лет (3–6 лет). R2 – коэффициент детерминации (R2 = 78.4%).Известно, что коэффициент естественной смертности рыб может зависеть от возраста (Chen, Watanabe, 1989). На рис. 1 видно, что смертность леща Куйбышевского водохранилища (тангенс угла наклона) в возрасте 6–10 лет существенно меньше, чем у более старших особей. Поэтому для описания изменения численности более старших возрастных групп взяты следующие линейные функции:
где Т =6–10 лет (R2 = 74.7%)
где Т =10–13 лет (R2 = 90.2%)
Коэффициент детерминации (R2) показал, что полином описывает 78.4% изменения относительной численности особей младших возрастных групп (3–6 лет), а линейные функции описывают 74.7–90.2% изменения обилия более старших особей. Это указывает на хорошую аппроксимацию полиномом и линейными функциями возрастной структуры леща Куйбышевского водохранилища.
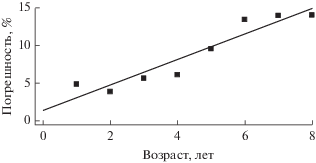
Погрешность оценки относительной численности поколений рыб (рис. 2, табл. 2) при потере информации о возрастной структуре популяции от 1 до 8 лет в среднем составляет 5–15%, хотя максимальное значение погрешности при этом может быть 20–50%. При увеличении объема недостающей информации о возрастной структуре рыб погрешность оценки численности поколения возрастает (рис. 3).
Рис. 2.
Гистограммы относительной погрешности численности поколений леща при потере информации о возрастной структуре за 1 (а), 2 (б), 3 (в), 4 (г), 5 (д), 6 (е), 7 (ж) и 8 (з) лет. По оси ординат – частота случаев с положительным и отрицательным отклонением.
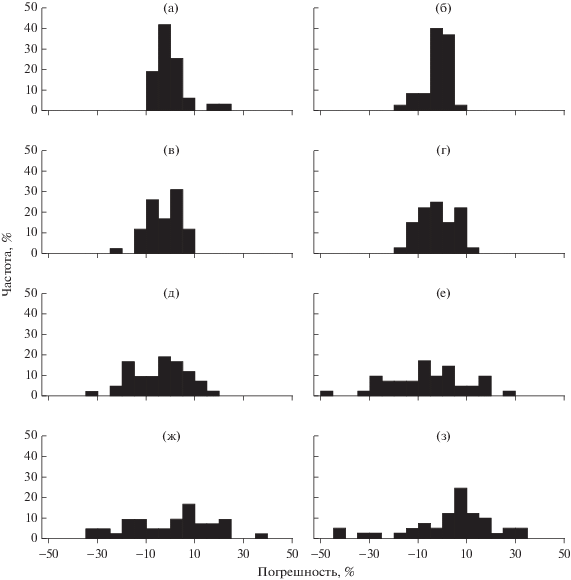
Таблица 2.
Погрешность оценки численности поколений леща Куйбышевского водохранилища при десятикратной повторности отсутствия информации о возрастной структуре для каждого из числа исключаемых лет (от 1 до 8)
| Число лет с отсутствующей информацией | Относительная погрешность, % | |||
|---|---|---|---|---|
| средняя | ошибка средней | минимальная | максимальная | |
| 1 | 4.9 | 4.6 | 0.2 | 22.0 |
| 2 | 3.9 | 4.0 | 0.1 | 16.4 |
| 3 | 5.7 | 4.8 | 0.1 | 22.6 |
| 4 | 6.2 | 4.3 | 0.1 | 15.2 |
| 5 | 9.6 | 7.6 | 0.2 | 32.5 |
| 6 | 13.5 | 10.4 | 0.1 | 45.6 |
| 7 | 14.0 | 8.7 | 0.9 | 36.3 |
| 8 | 14.1 | 10.9 | 1.1 | 44.7 |
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
На примере леща Куйбышевского водохранилища оценена погрешность численности поколений популяций рыб, возникающая из-за потери информации об ее возрастной структуре за 1–8 лет и достигающая 50% исходного ряда. С увеличением объема отсутствующей информации погрешность численности поколения возрастает. Однако, уже наличие данных о возрастной структуре за 4 года позволяет в первом приближении получить количественную информацию о численности поколения популяции, что ранее нельзя было сделать. Погрешность такой оценки достигает в среднем 15%, т.е. данная модификация метода убыли от лова применима. Особенно это важно для мониторинга состояния популяций охраняемых видов рыб. Полученную при этом информацию по степени точности можно отнести к приближенной (Поддубный, 1982). При дополнении данных о возрастной структуре оценка численности поколения может быть переведена в класс обычных исследований, а при вылове всего поколения – оценка численности поколения переходит в разряд точных.
Необходимо учитывать, что в целом метод убыли от лова дает надежную оценку величины численности поколений популяций рыб в водоемах с мало изменчивым промысловым усилием (Никольский, 1974; Рикер, 1979). Вместе с тем, в Куйбышевском, как и в ряде других равнинных водохранилищ, поколения леща и других фитофильных видов рыб первых лет его заполнения (1956–1957 гг.) были многочисленными (Лукин, 1986; Цыплаков, 1972). По мере их созревания изменялся и промысел. Тем не менее, даже в таких условиях модификация метода показала свою работоспособность.
Метод убыли от лова разработан для рыб. И неудивительно, что ихтиологи оказались впереди в плане оценки состояния и анализа изменений в популяциях животных, поскольку потребности практики способствуют продвижению науки. Кроме рыб, среди гидробионтов возрастную структуру популяции и ее динамику исследуют также у длинноцикловых ракообразных (Gongora, 2010; Maxwell et al., 2013; Roa et al., 1995; Roa, Erns, 1996; Sharifan et al., 2017 и др.) и млекопитающих (Огнетов, Светочева, 2000; Трухин, 1996; Чакилев, Кочнев, 2017; Boyd et al., 1995; Dabin et al., 2004; Gibbens, Arnould, 2009; Kuzin, 2010; Taylor et al., 2018 и др.), поэтому данный метод оценки численности поколений может быть полезен при анализе и этих гидробионтов.
Анализ изменений в популяциях рыб требует в дальнейшем решения ряда методических вопросов, в частности, какой минимальный ряд возрастной структуры достаточен для оценки относительной численности поколения рыб и какая при этом будет погрешность, как изменится погрешность оценки численности поколения, если информация о возрастной структуре этого поколения была представлена младшими или старшими возрастными группами.
Необходимо отметить, что данная модификация метода оценки относительной численности поколения рыб применима только к случаю отсутствия за некоторые годы информации о возрастной структуре популяции. Предполагается, что имеющиеся у исследователя данные за другие годы репрезентативны по объему собранного материала и охвату проанализированных биотопов, а при определении возраста отсутствует систематическая погрешность, связанная с определением возраста различными операторами. Если за некоторые годы информация о возрастной структуре популяции не достоверна, т.е. собрана с большими погрешностями, то ее необходимо исключить из расчетов.
Выводы. Оценена погрешность относительной численности поколений рыб для случая отсутствия полной исходной информации о возрастной структуре исследуемой популяции. Для длинноцикловых видов, даже при наличии информации о возрастной структуре популяции за 4 года, средняя погрешность оценки мощности поколений не превышает 15%. Оценка погрешности метода показала возможность его применения для количественной оценки численности поколения рыб.
Список литературы
Анохина О.К., Гончаренко К.С., Говоркова Л.К. 2013. Промыслово-биологическая характеристика, состояние промысловых запасов и допустимые уловы рыб в Куйбышевском водохранилище // Гидробиологические и ихтиологические исследования водоёмов Среднего Поволжья. № 13. С. 152.
Кондрашов А.П., Шестопалов Е.В. 1977. Основы физического эксперимента и математическая обработка результатов измерения. Москва: Атомиздат.
Коновалов А.Ф. 2004. Роль судака в экосистемах крупных озер Вологодской области: Автореф. дис. … канд. биол. наук. Петрoзаводск.
Кузнецов В.А. 1980. Флюктуации численности промысловых рыб в условиях зарегулированного стока реки (на примере Куйбышевского водохранилища) // Вопр. ихтиол. Т. 20. Вып. 5(124). С. 805.
Лукин А.В. 1986. Основные этапы формирования ихтиофауны и состояние запасов рыб. Состояние запасов леща // Экологические особенности рыб и кормовых организмов Куйбышевского водохранилища. Казань: Изд-во КГУ. С. 5.
Монастырский Г.И. 1952. Динамика численности промысловых рыб // Тр. ВНИРО. Москва: ВНИРО. Т. 21. С. 3.
Никольский Г.В. 1974. Теория динамики стада рыб. Москва: Пищевая промышленность. 447 с.
Огнетов Г.Н., Светочева О.Н. 2000. Исследование половой и возрастной структуры беломорской нерпы // Вопросы рыболовства. Т. 1. № 2–3. С. 78.
Поддубный А.Г. 1982. Значение работ по оценке точности результатов биологических исследований // Оценка погрешностей методов гидробиологических и ихтиологических исследований. Рыбинск: Ин-т биологии внутр. вод АН СССР. С. 3.
Рабинович С.Г. 1978. Погрешности измерений. Ленинград: Энергия.
Рикер У.Е. 1979. Методы оценки и интерпретации биологических показателей популяций рыб. Москва: Пищевая промышленность.
Стрельников А.С. 1996. Состояние популяции судака Stizostedion lucioperca (L.) Рыбинского водохранилища в условиях новых коммерческих отношений // Вопр. ихтиол. Т. 36. Вып. 4. С. 481.
Стрельников А.С., Орлова С.С., Терещенко В.Г. 1997. Стратегия охраны запасов и прогнозирование уловов рыбы в Рыбинском водохранилище в условиях новых коммерческих отношений // Современное состояние рыбных запасов Рыбинского водохранилища. Ярославль: Ин-т биологии внутр. вод РАН. С. 178.
Трухин А.М. 1996. Межгодовая динамика возрастной структуры кольчатой нерпы северо-западной популяции Охотского // Известия Тихоокеанского научно-исследовательского рыбохозяйственного центра. Т. 121. С. 127.
Цыплаков Э.П. 1972. Лещ // Тр. Татар. отд. ГосНИОРХ. Вып. XII. Казань. С. 68.
Чакилев М.В., Кочнев А.А. 2017. Половозрастная структура тихоокеанского моржа Odobenus rosmarus divergens в районе мыса Сердце-Камень (Чукотское море) в летне-осенний период 2009–2014 гг. // Известия ТИНРО. Т. 190. С. 72. https://doi.org/10.26428/1606-9919-2017-190-72-78
Чугунова Н.И. 1959. Руководство по изучению возраста и роста рыб. Москва: Изд-во АН СССР.
Boyd I.L., Croxall J.P., Lunn N.J., Reid K. 1995. Population demography of Antarctic fur seals: the costs of reproduction and implications for life-histories // J. Anim. Ecol. V. 64. P. 505.
Chen S., Watanabe S. 1989. Age dependence of natural mortality coefficient in fish population dynamics // Nippon Suisan Gakkaishi. V. 55. P. 205.
Dabin W., Beauplet G., Crespo E.A., Guinet Ch. 2004. Age structure, growth, and demographic parameters in breeding-age female subantarctic fur seals, Arctocephalus tropicalis // Can. J. Zool. V. 82(7). P. 1043. https://doi.org/10.1139/z04-079
Gibbens J., Arnould J.P.Y. 2009. Age-specific growth, survival, and population dynamics of female Australian fur seals // Can. J. Zool. V. 87. P. 902. https://doi.org/10.1139/Z09-080
Gongora M. 2010. Assessment of Spiny Lobster (Panulirus argus) of Belize based on fishery dependent data. United Nations University Fisheries Training Programme, Iceland [final project]. http://www.unuftp.is/static/fellows/document/mauro09prf.pdf
Kuzin A.E. 2010. The Intrapopulation Structure of the Northern Fur Seal (Callorhinus ursinus L.) on Tyuleniy Island during the Post-Depression Years (1993–2009) // Russ. J. Mar. Biol. V. 36. № 7. P. 507. https://doi.org/10.1134/S1063074010070047
Maxwell K.E., Matthews T.R., Bertelsen R.D., Derby Ch.D. 2013. Age and size structure of Caribbean spiny lobster, Panulirus argus, in a no-take marine reserve in the Florida Keys, USA // Fisheries Research. V. 144. P. 84.
Reshetnikov Yu.S., Tereshchenko V.G. 2017. Quantitative level of research in fish ecology and errors associated with it // Russ. J. Ecol. V. 48. № 3. P. 233. https://doi.org/10.1134/S1067413617030146
Roa R., Erns B. 1996. Age structure, annual growth, and variance of size-at-age of the shrimp Heterocarpus reedi // Mar. Ecol.: Prog. Ser. V. 137. P. 59.
Roa R., Gallardo V.A., Ernst B. et al. 1995. Nursery ground, age structure and abundance of juvenile squat lobster Pleuroncodes monodon on the continental shelf off central Chile // Mar. Ecol.: Prog. Ser. V. 116. P. 47.
Sharifian S., Kamrani E., Safaie M., Sharifian S. 2017. Population structure and growth of freshwater crab Sodhiana iranica from the south of Iran // Fundam. Appl. Limnol. V. 189/4. P. 341. https://doi.org/10.1127/fal/2017/0965
Taylor R.L., Udevitz M.S., Jay Ch.V. et al. 2018. Demography of the Pacific walrus (Odobenus rosmarus divergens) in a changing Arctic // Mar. Mammal Sci. V. 34(1). P. 54. https://doi.org/10.1111/mms.12434
Tereshchenko V.G., Zuyanova O.V. 2006. The method of assessment of relative abundance of commercial fish species generations under conditions of incomplete initial information // Biol. Vnutr. Vod. № 1. P. 88.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод