

Биология внутренних вод, 2020, № 2, стр. 116-123
Строение клетки хищного жгутиконосца Metromonas grandis (Cercozoa)
А. А. Мыльников a, К. И. Прокина a, b, *, А. П. Мыльников a
a Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
Некоузский р-н, Ярославская обл., пос. Борок, Россия
b Зоологический институт Российской академии наук
Санкт-Петербург, Россия
* E-mail: kristin892@mail.ru
Поступила в редакцию 29.03.2018
После доработки 26.01.2019
Принята к публикации 09.04.2019
Аннотация
Изучено ультратонкое строение хищного жгутиконосца Metromonas grandis Larsen et Patterson, 1990. Цитостом в виде клеточного углубления не найден. Клетка окружена волокнистым слоем толщиной 0.05–0.12 мкм, который состоит из тонких ворсинок (волокон) и тонких извитых палочек (стержней). Ниже этого слоя располагается плазмалемма. Два неравных жгутика выходят из небольшого кармана. Длинный жгутик несет многочисленные очень тонкие мастигонемы (волоски) длиной 0.8–1.0 мкм, короткий жгутик редуцирован и покрыт немногочисленными мастигонемами. Поперечная пластинка приподнята над поверхностью клетки и окружена осмиофильной муфтой, спираль отсутствует. Корешковая система жгутиков упрощена и насчитывает две ленты микротрубочек, которые начинаются вблизи кинетосомы длинного жгутика. Кинетосомы лежат почти параллельно друг другу и связаны, по крайней мере, одним фиброзным мостиком. Пузырьковидное ядро, аппарат Гольджи и эндоплазматический ретикулум имеют обычное строение. Овальные митохондрии содержат пластинчатые кристы. В цитоплазме обнаружены цилиндрические экструсомы (трихоцисты) длиной 1.0–1.4 мкм и диаметром 0.12–0.08 мкм. Трихоцисты разделены перемычкой и содержат колесовидную структуру, состоящую из 13 спиц или секторов, заметных на поперечных срезах. Обсуждается сходство Metromonas grandis с M. simplex (Griessmann, 1913) Larsen et Patterson, 1990, Metopion fluens Larsen et Patterson, 1990, криомонадами и другими хищными жгутиконосцами.
ВВЕДЕНИЕ
Существует небольшая группа облигатно хищных гетеротрофных жгутиконосцев, занимающих пока неопределенное систематическое положение, так как они сочетают в своем строении признаки неродственных таксонов и имеют уникальные клеточные структуры. Исследование этих протистов в настоящее время становится особенно актуальным, когда пересматривается макросистема одноклеточных (Adl et al., 2019; Bass, Cavalier-Smith, 2004; Bass et al., 2005). Установлено, что многие такие хищные виды имеют всесветное распространение (Chantangsi, Leander, 2010), но пока они не выделены в чистые культуры. Малоизученными остаются недавно описанные хищные жгутиконосцы, которые известны из морских и пресных вод. Систематическое положение именно таких жгутиконосцев определяют, в основном, молекулярно-генетическими методами (Bass et al., 2004, 2005). Однако эти работы необходимо дополнять ультраструктурными исследованиями. Известно, что для хищных протистов характерно наличие крупных видоспецифичных экструсом, цитостома или цитостомальной щели, а также дополнительных структур в покровах (Мыльников и др., 1999; Chantangsi, Leander, 2010; Clay, Kugrens, 1999; Hoppenrath, Leander, 2006). Прогресс в изучении строения этих организмов связан с возможностью получения чистых культур на основе подходящего пищевого объекта. Обнаружено, что рассматриваемые хищники обычно погибают в отсутствие эукариотной жертвы (жгутиконосцев или инфузорий) (Мыльников и др., 1999; Mylnikov, 1986; Mylnikova, Mylnikov, 2011). Бактерии в их пищеварительных вакуолях обнаруживаются лишь вместе с клетками эукариотной жертвы. Обычно хищные жгутиконосцы – быстро плавающие протисты, активно нападающие на клетки жертвы с последующим поглощением их клеток целиком или частичным “высасыванием” их цитоплазмы (Clay, Kugrens, 1999; Mylnikov, 1986). Ларсеном и Паттерсоном (Larsen, Patterson, 1990) описано несколько новых родов и видов хищных жгутиконосцев. Причем представители родов Metopion Larsen et Patterson, 1990 и Metromonas Larsen et Patterson, 1990 относятся к ползающим или прикрепленным формам, неспособным к активному плаванию, что необычно для хищных флагеллят. В пробах из водоема Вьетнама были обнаружены прикрепленные бесцветные жгутиконосцы Metromonas grandis Larsen et Patterson, 1990.
Цель работы – описание ультратонкого строения морского хищного жгутиконосца Metromonas grandis.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Выделенный в чистую культуру (25.02.2016 г.) клон Metr-3 (Metromonas grandis) получен из пробы с архипелага Кондао (Южный Вьетнам) в лагуне небольшого острова Байкань, отобранной сотрудником Института биологии внутренних вод РАН, пос. Борок Ярославской обл. Е.С. Гусевым 3 мая 2015 г. Данный клон хранится в коллекции живых культур протистов группы протозоологии указанного института. Жгутиконосцев культивировали в среде Шмальца–Пратта. Пищей служили бодониды Procryptobia sorokoni (Zhukov, 1975) Frolov et al., 2001, выращенные в той же среде с добавлением бактерий Pseudomonas fluorescens Migula, 1895.
Для светомикроскопических наблюдений использовали микроскопы AxioScope A1 (Carl Zeiss, Германия) с фазово-контрастной и дифференциально-контрастной насадками и объективами водной иммерсии (×1000). Микроскопы были оборудованы цифровой фотокамерой AxioCam ERc 5s и аналоговой видеокамерой AVT HORN MC-1009/S. Для более четкой идентификации обнаруженных жгутиконосцев проводили прямую запись видеофильма с оцифровкой изображений и сохранением фрагментов видеофильма в виде файлов формата WMV.
Для электронно-микроскопического изучения взвесь жгутиконосцев сгущали центрифугированием. Клетки фиксировали в смеси 2%-ного раствора OsO4 и 0.6%-ного раствора глутаральдегида (1 : 1), приготовленных на среде Шмальца–Пратта в течение 15–30 мин при 1°С. После дегидратации в серии спиртов и безводного ацетона материал заключали в смесь Аралдита и Эпона. Для трансмиссионной электронной микроскопии (ТЭМ) получали срезы с помощью ультрамикротома LKB (Швеция) и исследовали в микроскопе JEM-1011 (Jeol, Япония). Для сканирующей электронной микроскопии (СЭМ) материал обезвоживали в аппарате критической точки и просматривали в микроскопе JSM-6510LV (Jeol, Япония).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
M. grandis относится к пассивным хищникам, он поглощает только те клетки, которые случайно соприкасаются с поверхностью хищника и прилипают к ней. При достижении максимума численности клетки M. grandis начинают быстро ползать, контактируя с субстратом длинным жгутиком, который направлен назад. В отсутствие жертвы (бодониды) хищники гибнут. Деление клетки начинается с удвоения жгутиков в стадии прикрепления жгутиконосца. Цист не обнаружено.
Форма клетки жгутиконосца овальная или сердцевидная, реже грушевидная. Верхний конец немного заострен. Клетка значительно уплощена. Диаметр клетки достигает 8–10 мкм (рис. 1а, 1б). Длинный жгутик (13–15 мкм) направлен всегда назад и у ползающих клеток целиком контактирует с дном чашки Петри (рис. 1а). У прикрепленных клеток он имеет форму крючка, конец которого прикрепляется к твердому субстрату. Эти клетки постоянно совершают маятникообразные движения (рис. 1в, 1г). Короткий жгутик заметен как на светомикроскопических снимках, так и в СЭМ (рис. 1д, 1е). На рис. 1ж показан продольный срез клетки, и видны основные ее комноненты. Вся клетка, кроме жгутиков и участков складок, окружена волокнистым слоем, который состоит из тонких ворсинок (волокон) и тонких извитых палочек (стержней) (рис. 1з–1к). Суммарная толщина волокнистого слоя 0.05–0.12 мкм. Ниже этого слоя располагается плазмалемма, имеющая обычное трехслойное строение. Волокнистый слой отсутствует в складках тела и бороздках (рис. 1к). Короткий жгутик покрыт немногочисленными тонкими малоконтрастными мастигонемами (волосками) длиной 0.8–1.0 мкм (рис. 1л). В свою очередь длинный жгутик густо покрыт такими же мастигонемами на всем своем протяжении (рис. 1м). Переходная зона жгутиков имеет обычное строение, спираль отсутствует. Поперечная пластинка приподнята над поверхностью клетки и окружена осмиофильной муфтой (рис. 1л). Кинетосомы обычного строения и длиной 0.45–0.60 мкм чаще располагаются параллельно друг другу (рис. 2). Кинетосомы соединены, по крайнем мере, одним фибриллярным мостиком (рис. 2). От кинетосомы длинного жгутика отходят две микротрубочковые ленты (рис. 2). Лента 1 состоит из 9–10 микротрубочек, которые сближены друг с другом и окружены аморфным осмиофильным материалом. Перпендикулярно этой ленте отходит лента 2 в виде расширяющегося веера из 7–8 микротрубочек, вероятно, поверхностно под плазмалеммой (рис. 1и). Фибриллярного жгутикового корешка (ризопласт), обычно направленного к ядру, не найдено.
Рис. 1.
Общий вид, строение покровов и жгутиков Metromonas grandis: а, б – ползающие клетки, видны оба жгутика (фазовый контраст и ДИК), в, г – прикрепленные раскачивающиеся клетки (фазовый контраст), д, е – прикрепленные клетки (СЭМ), ж – продольный срез клетки, з–к – покровы клетки, л – короткий жгутик, м – длинный жгутик. Обозначения: а.г – аппарат Гольджи, бр – бороздка, в.с – волокнистый слой, д.ж – длинный жгутик, ж.к – жгутиковый карман, к.ж – короткий жгутик, мн – мастигонемы, мф – муфта, п.в – пищеварительная вакуоль, пл – плазмалемма, пс – псевдоподия, тц – трихоциста, я – ядро, яд – ядрышко.

Рис. 2.
Расположение кинетосом и лент микротрубочек на серии срезов одной клетки Metromonas grandis (а–д) и на отдельном срезе другой клетки (е). Обозначения: 1 л.мкт – первая лента микротрубочек, 2 л.мкт – вторая лента микротрубочек, к.д.ж – кинетосома длинного жгутика, к.к.ж – кинетосома короткого жгутика, мс – мостик между кинетосомами. Остальные обозначения, как на рис. 1.
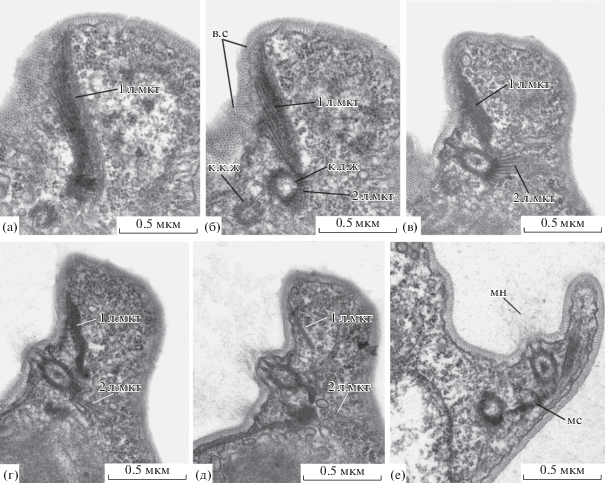
У M. grandis не обнаружено цитостомальной бороздки или цитостома. Псевдоподии, которые, вероятно, участвуют в поглощении клеток жертв, образуются на заднем конце клетки вблизи батареи трихоцист (рис. 1д). На срезах клетки эти псевдоподии не выявлены.
Пузырьковидное ядро содержит центральное ядрышко (рис. 1ж, 3а, 3б). Аппарат Гольджи (0.4 × 2.5 мкм) в виде стопки уплощенных пузырьков обычно располагается вблизи ядра (рис. 3б).
Рис. 3.
Компоненты клетки Metromonas grandis: а – ядро и ядрышко, б – аппарат Гольджи, в, г – митохондрии, д – микротельце и трихоциста, е – пищеварительная вакуоль, ж, з – триоцисты, и – предшественники мастигонем и гранула запасного вещества. Обозначения: аг – аппарат Гольджи, з.в – гранула запасного вещества, кп – кинетопласт, ми – микротельце, мт – митохондрия, пт – перетяжка. Остальные обозначения, как на рис. 1 и 2.

Овальные митохондрии с пластинчатыми кристами равномерно рассеяны в цитоплазме (рис. 3в, 3г). Длина митохондрий на срезах достигает 0.8 мкм, диаметр 0.4–0.6 мкм. Эндоплазматический ретикулум слабо выражен, обычного строения. Микротельце в виде небольшой структуры (0.5 × 0.8 мкм), имеющей неровные очертания и содержащей аморфный осмиофильный материал и мелкие пузырьки, примыкает к пищеварительной вакуоли (рис. 3д).
Клетка содержит стрекательные органеллы (два ряда) – трихоцисты, длиной 1.0–1.4 мкм и диаметром 0.12–0.08 мкм (рис. 1ж, 3д, 3ж, 3з), которые хорошо заметны при наблюдении в СЭМ в виде выпячиваний (рис. 1д, 1е). Эти органеллы имеют цилиндрическую форму и разделены на две почти равные части перемычкой (рис. 3ж). Трихоциста погружена в пузырек. На одном из концов трихоцисты заметна колесовидная структура (рис. 3з). Последняя содержит 13 спиц или секторов. Зачатки этих экструсом обнаружены в дериватах аппарата Гольджи (рис. 3б).
Сократительная вакуоль не обнаружена. Клетки бодонид (жертвы) заглатываются хищником целиком и попадают в пищеварительную вакуоль диаметром 1.5–3.0 мкм (рис. 3е). Внутри заметен кинетопласт жертвы. Момент заглатывания жертв клеткой хищника на срезах не зафиксирован. Запасные вещества представлены гранулами (рис. 3и). Предшественники мастигонем обнаруживаются в цитоплазматических пузырьках (рис. 3и).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Форма, размеры клетки и характер движения клона Metr-3 соответствуют светомикроскопическим признакам вида Metromonas grandis (Larsen, Patterson, 1990).
Ультраструктура этого хищника имеет следующие характерные признаки: волокнистый слой, покрывающий плазмалемму; редукция одного из жгутиков; многочисленные тонкие мастигонемы, покрывающие длинный жгутик и немногочисленные мастигонемы – короткий жгутик; две микротрубочковые ленты; муфта в переходной зоне жгутика; отсутствие цитостома; цилиндрические трихоцисты; параллельное расположение кинетосом; пластинчатые кристы митохондрий, небольшие микротельца. Отнесение клона Metr-3 к виду M. grandis не вызывает сомнений. Все перечисленные признаки ранее обнаружены у другого более мелкого вида этого рода M. simplex (Mylnikov A.A., Mylnikov A.P., 2011). Клон Metr-3 отличается от M. simplex наличием второй микротрубочковой ленты, мастигонем на коротком жгутике, отсутствием тонкого слоя внутри волокнистого слоя и большим размером клетки. Кроме того, у клона Metr-3 найдены псевдоподии, возможно, участвующие в захвате клеток жертвы. Существует еще один хищный жгутиконосец Metopion fluens Larsen et Patterson, 1990, ультраструктура которого почти идентична видам рода Metromonas (Мыльников и др., 1999). У них почти одинаковые покровы, мастигонемы на жгутиках, редукция одного жгутика, палочковидные трихоцисты с 13 секторами, видимыми на поперечных срезах (спицы), слабая развитость микротрубочковых лент и фибриллярных структур. Metopion fluens при отсутствии цитостома образует сетчатые псевдоподии. Кроме того, этот вид ведет ползающий образ жизни в отличие от прикрепленного у Metromonas.
Внешние структурированные или волокнистые слои, дополняющие снаружи плазмалемму, известны у некоторых жгутиконосцев. Уплотненный осмиофильный слой, примыкающий к плазмалемме снаружи, характерен для хищного почвенного жгутиконосца Hemimastix amphikineta Foissner, Blatterer et Foissner, 1988 (Foissner et al., 1988), однако, в отличие от Metromonas simplex, он не содержит волокнистых компонентов. Волокнистый слой оболочки клетки обнаружен у недавно описанного жгутиконосца Mataza hastifera Yabuki et Ishida, 2011 (Yabuki, Ishida, 2011). На поверхности клетки бактериотрофного жгутиконосца Ancyromonas sigmoides Kent, 1880 (Мыльников, 1990) располагается тонкий слой уплотненного гликокаликса, который отсутствует в области жгутикового кармана. У мелкого бактериотрофного жгутиконосца Discocelis saleuta Vørs, 1988 найден тонкий паракристаллический слой, который, в отличие от Metromonas simplex, лежит под плазмалеммой (Vørs, 1988).
Волокнистые покровы обнаружены также у хищных жгутиконосцев рода Cryothecomonas Thomsen et al., 1991 и Protaspa grandis (Hoppenrath et Leander, 2006) Cavalier-Smith in Howe et al., 2011 (Hoppenrath, Leander, 2006; Thomsen et al., 1991), относящихся к криомонадам (Chantangsi, Leander, 2010). Эти покровы состоят из трех слоев: внутренних аморфных (светлого и плотного) и внешнего волокнистого, и больше напоминают таковые Metromonas grandis. Выявлено также частичное сходство волокнистых покровов M. grandis с внешним структурированным ламеллярным (пластинчатый) слоем морских хищных Kathablepharis Skuja, 1932 и Leucocryptos Butcher, 1967 (Clay, Kugrens, 1999), покрывающим не только всю клетку, но и жгутики. У представителей этих родов он состоит из тонких чешуек, которые лежат рядами, направленными косо по отношению к продольной оси клетки. Под чешуйками располагаются две тонкие пластины и плазмалемма обычного строения. Толщина ламеллярного слоя, покрывающего всю клетку и жгутики, равна 30–45 нм. У рассматриваемого клона Metr-3 волокнистые покровы не образуют чешуек.
Длинный жгутик Metromonas grandis покрыт густым слоем тонких мастигонем, которые напоминают тонкие жгутиковые волоски монадных протистов (бодониды, эвглениды, гаптомонады, зеленые водоросли и др.) (Карпов, 1987). У большинства плавающих жгутиконосцев жгутиковые волоски участвуют в создании потока воды около клетки, однако, хищный M. grandis относится к прикрепленным протистам, и длинный жгутик не участвует в процессе захвата пищи. Похожие волоски покрывают длинный жгутик Metopion fluens.
Переходная зона жгутиков Metromonas grandis устроена просто, какие-либо добавочные структуры (спирали и цилиндры), за исключением небольшой муфты, позволяющие сравнить эти жгутики с таковыми у других хищников, отсутствуют. В отличие от большинства хищных жгутиконосцев цитоплазматические корешковые структуры M. grandis значительно упрощены и содержат одну фибриллу, соединяющую кинетосомы, и две отходящие микротрубочковые ленты, тогда как у M. simplex и Metopion fluens найдена лишь одна такая лента (Мыльников и др., 1999; Mylnikov A.A., Mylnikov A.P., 2011). Похожее строение корешковых структур жгутиков характерно для видов рода Cryothecomonas (Thomsen et al., 1991).
Экструсомы Metromonas simplex отнесены нами к цилиндрическим (палочковидным) трихоцистам (по: Hausmann, 1978). При выстреливании размеры и форма такой экструсомы не изменяются. Эти экструсомы представляют собой вытянутую палочковидную структуру, в отличие от концентрических или грибовидных экструсом некоторых солнечников (Bardele, 1972), амебоидных жгутиконосцев (Мыльников, 1986) и Discocelis saleuta Vørs, 1988 (Vørs, 1988); и от микротоксицист и токсицист церкомонад (Мыльников, 1986), инфузорий и хищного жгутиконосца Colponema loxodes Stein, 1878 (Hausmann, 1978). Экструсомы Metromonas grandis отличаются также от веретеновидных трихоцист инфузорий и динофитовых (Hausmann, 1978). Последние имеют кристаллоидную структуру и при выстреливании увеличиваются в длине. Цилиндрические трихоцисты известны в клетках хищных жгутиконосцев кинетопластид Pseudophyllomitus apiculatus (Skuja, 1948) Lee, 2002 (Mylnikov, 1986), церкозоев Mataza hastifera (Yabuki, Ishida, 2011), Cryothecomonas armigera Thomsen Buck, Bolt et Garrison, 1991 (Thomsen et al., 1991), Commation cryoporinum Thomsen et Larsen, 1993 (Thomsen, Larsen, 1993). На поперечных срезах колесовидная часть экструсом Metromonas grandis напоминает среднюю часть крупных трихоцист жгутиконосца Hemimastix amphikineta (обнаружено 11 спиц) (Foissner et al., 1988) и Metopion fluens (15 спиц) (Мыльников и др., 1999).
Пузырьковидное ядро у Metromonas grandis соответствует ядру большинства групп жгутиконосцев (Карпов, 2001). Аппарат Гольджи, как у многих флагеллят, лежит в переднем конце клетки между кинетосомами и ядром. Эндоплазматический ретикулум имеет обычное строение. Микротельца напоминают таковые у многих церкомонад, M. simplex и Metopion fluens (Мыльников, 1986; Мыльников и др., 1999; Mylnikov A.A., Mylnikov A.P., 2011). Митохондрии Metromonas grandis и M. simplex содержат пластинчатые кристы, в отличие от трубчатых у других хищных флагеллят Metopion fluens (Мыльников и др., 1999), Hemimastix Foissner et al., 1988 (Foissner et al., 1988), Aulacomonas Skuja, 1939 (Brugerolle, Patterson, 1990), Kathablepharis (Clay, Kugrens, 1999), Cryothecomonas (Thomsen et al., 1991) и Commation Thomsen et Larsen, 1993 (Thomsen, Larsen, 1993).
По основным признакам (два жгутика, волокнистые покровы, цилиндрические экструсомы, малое развитие корешковой системы) виды рода Metromonas сходны с представителями хищных Metopion и Cryothecomonas (Мыльников и др., 1999; Thomsen et al., 1991). В отличие от Metopion и Metromonas, виды рода Cryothecomonas имеют отстающие в некоторых местах от поверхности клетки волокнистые слои, небольшие псевдоподии содержат микротрубочки, жгутики одинаковой длины и не покрыты волосками, переходная зона включает спираль и добавочное электронно-плотное тельце. Известно, что представители рода Cryothecomonas – холодолюбивые плавающие организмы, в отличие от Metromonas grandis, хорошо размножающихся при комнатной температуре и не способных к активному плаванию.
Полученные данные сближают роды Metromonas и Metopion. Их объединяет общее строение клетки и наличие волосков на жгутиках. Выявлено сходство строения и размеров трихоцист, не встречающихся у других жгутиконосцев. В частности, колесовидная структура со спицами у представителей обоих родов совпадает по строению. Отличаются они в основном по форме крист митохондрий (трубчатые у Metopion и пластинчатые у Metromonas).
Исследование сиквенсов РНК малой субъединицы рибосом показало близкородственные связи этих двух родов хищников и родов Cryothecomonas (Thomsen et al., 1991) и Protaspa (Chantangsi, Leander, 2010). В свою очередь, два последних рода сейчас входят в отряд Cryomonadida Cavalier-Smith, 1993, который отнесен к крупной группе Cercozoa Cavalier-Smith, 1998 (Adl et al., 2019; Bass et al., 2005).
Таким образом, морфологические данные, полученные нами, подтверждают близкие родственные связи Metromonas, Metopion, Protaspis и Cryothe-comonas, ранее установленные по сиквенсам генов малой субъединицы рибосом этих организмов. Эти организмы могут быть отнесены к отряду Cryomonadida (криомонады).
Выводы. Впервые исследована морфология хищного жгутиконосца Metromonas grandis. Найдены волокнистые покровы, сильная опушенность длинного жгутика, пластинчатые кристы митохондрий, микротельца и палочковидные трихоцисты, что позволяет сблизить этот вид с другими хищными жгутиконосцами из группы криомонад.
Список литературы
Карпов С.А. 1987. Строение аксонемы и наружных элементов жгутика у подвижных клеток водорослей и у бесцветных жгутиконосцев // Бот. журн. Т. 72. № 1. С. 3.
Карпов С.А. 2001. Строение клетки протистов: Уч. пособие. Санкт-Петербург: ТЕССА.
Мыльников А.П. 1986. Биология и ультраструктура амебоидных жгутиконосцев Cercomonadida ord. n. // Зоол. журн. Т. 65. Вып. 5. С. 683.
Мыльников А.П. 1990. Особенности ультраструктуры бесцветного жгутиконосца Heteromita sp. // Цитология. Т. 32. № 6. С. 567.
Мыльников А.П., Мыльникова З.М., Цветков А.И. 1999. Ультраструктура хищного морского жгутиконосца Metopion fluens // Цитология. Т. 41. № 7. С. 581.
Adl S.M., Bass D., Lane C.E. et al. 2019. Revisions to the classification, nomenclature, and diversity of eukaryotes // J. Eukaryotic Microbiol. V. 66. № 1. P. 4. https://doi.org/10.1111/jeu.12691
Bardele C.F. 1972. Cell cycle, morphogenesis and ultrastructure in the pseudo-helizoan Clathrulina elegans // Z. Zellforsch. Mikrosk. Anat. V. 130. P. 219.
Bass D., Cavalier-Smith T. 2004. Phylum-specific environmental DNA analysis reveals remarkably high global biodiversity of Cercozoa (Protozoa) // Int. J. Syst. Evol. Microbiol. V. 54. № 6. P. 2393. https://doi.org/10.1099/ijs.0.63229-0
Bass D., Moreira D., López-García L. et al. 2005. Polyubiquitin insertions and the phylogeny of Cercozoa and Rhizaria // Protist. V. 156. P. 149. https://doi.org/10.1016/j.protis.2005.03.001
Brugerolle G., Patterson D.J. 1990. A cytological study of Aulacomonas submarina Skuja 1939, a heterotrophic flagellate with a novel ultrastructural identity // Eur. J. Protistology. V. 25. № 3. P. 191. https://doi.org/10.1016/S0932-4739(11)80170-7
Chantangsi C., Leander B.S. 2010. An SSU rDNA barcoding approach to the diversity of marine interstitial cercozoans, including descriptions of four novel genera and nine novel species // Int. J. Syst. Evol. Microbiol. V. 60. № 8. P. 1962. https://doi.org/10.1099/ijs.0.013888-0
Clay B., Kugrens P. 1999. Systematics of the enigmatic Kathablepharids, including EM characterization of the type species, Kathablepharis phoenikoston, and new observations on K. remigera comb. nov. // Protist. V. 150. P. 43. https://doi.org/10.1016/S1434-4610(99)70008-8
Foissner W., Blatterer H., Foissner I. 1988. The Hemimastigophora (Hemimastix amphikineta nov. gen., nov. spec.), a new protistan phylum from Gondwanian soils // Eur. J. Protistol. 1988. V. 23. № 4. P. 361. https://doi.org/10.1016/S0932-4739(88)80027-0
Hausmann K. 1978. Extrusive organelles in protists // Int. Rev. Cytol. V. 52. P. 197.
Hoppenrath M., Leander B.S. 2006. Dinoflagellate, Euglenid or Cercomonad? The ultrastructure and Molecular Phylogenetic Position of Protaspis grandis n. sp. // J. Eukaryotic Microbiol. V. 53. P. 327. https://doi.org/10.1111/j.1550-7408.2006.00110.x
Larsen J., Patterson D.J. 1990. Some flagellates (Protista) from tropical marine sediments // J. Natural History. V. 24. P. 801. https://doi.org/10.1080/00222939000770571
Mylnikov A.P. 1986. Ultrastructure of a colourless flagellate Phyllomitus apiulatus Skuja, 1948 (Kinetoplastida) // Archiv für Protistenkunde. V. 132. P. 1. https://doi.org/10.1016/S0003-9365(86)80001-X
Mylnikov A.A., Mylnikov A.P. 2011. Ultrastructure of the marine predatory flagellate Metromonas simplex Larsen et Patterson, 1990 (Cercozoa) // Inland Water Biology. V. 4. № 2. P. 105. https://doi.org/10.1134/S1995082911020155
Thomsen H.A., Buck K.R., Bolt P.A., 1991. Fine structure and biology of Cryothecomonas gen. nov. (Protista incertae sedis) from the ice biota // Can. J. Zool. V. 69. P. 1048.
Thomsen H.A., Larsen J. 1993. The ultrastructure of Commation gen. nov. (Stramenopiles incertae sedis), a genus of heterotrophic nanoplanktonic flagellates from antarctic waters // Eur. J. Protistology. V. 29. P. 462. https://doi.org/10.1016/S0932-4739(11)80409-8
Vørs N. 1988. Discocelis saleuta gen. nov. et sp. nov. (Protista incertae sedis) // Eur. J. Protistology. V. 23. P. 297. https://doi.org/10.1016/S0932-4739(88)80019-1
Yabuki A., Ishida K.-I. 2011. Mataza hastifera n. g., n. sp.: a possible new lineage in the Thecofilosea (Cercozoa) // J. Eukaryotic Microbiol. V. 58. № 2. P. 94. https://doi.org/10.1111/j.1550-7408.2010.00524.x
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод