

Биология внутренних вод, 2020, № 3, стр. 276-288
Реофильные сообщества макрозообентоса Западного Тянь-Шаня
М. В. Чертопруд a, *, Д. М. Палатов a, b, Е. С. Чертопруд a, b
a Московский государственного университет им. М.В. Ломоносова
Москва, Россия
b Институт проблем экологии и эволюции им. А.Н. Северцова РАН
Москва, Россия
* E-mail: lymnaea@yandex.ru
Поступила в редакцию 25.04.2019
После доработки 23.05.2019
Принята к публикации 18.08.2019
Аннотация
На материале 370 оригинальных количественных проб из водотоков Западного Тянь-Шаня на территории Киргизии описаны региональное разнообразие и особенности реофильных сообществ макрозообентоса. Выявлены различия в составе и структуре сообществ трех высотно-температурных зон региона в высотном диапазоне от 1000 до 3700 м над уровнем моря. Выделено 23 типа сообществ, отнесенных к пяти ранее выделенным классам: ритраль (плотные субстраты на быстром течении), креналь (мозаичные субстраты малых водотоков), пелаль (мягкие грунты), рипаль (мозаичные субстраты береговой кромки) и эфемераль (временные биотопы). Показано, что сообщества низкогорной зоны структурно близки к сообществам других низкогорий южно-палеарктических регионов (Кавказ, Балканы), сообщества среднегорной зоны – к таковым низкоарктических регионов (Кольский п-ов, Ямал), сообщества высокогорной зоны – к высокоарктическим сообществам (арх. Шпицберген). Действие высотной и широтной поясности для горных сообществ реофильного макробентоса Палеарктики в общих чертах сходно и опосредовано главным образом летней температурой воды.
ВВЕДЕНИЕ
В серии публикаций последних лет (Chertoprud, 2011; Chertoprud, Palatov, 2013, 2017; Chertoprud et al., 2017, 2018; Palatov, Chertoprud, 2012, 2018; Palatov et al., 2016) рассматривались разнообразие реофильных сообществ макрофауны различных регионов Евразии: от apх. Шпицберген и п-ова Ямал до Балкан и Гималаев. Эти исследования показали, что все варианты таких сообществ в любом регионе укладываются в единую схему: большинство типов сообществ повторяются в разных регионах, сохраняя набор и, отчасти, количественное соотношение семейств и жизненных форм, но различаясь составом родов и видов. Таким образом, основной действующий здесь географический принцип – принцип параллельности сообществ разных регионов, многократно описанный для различных объектов (Чертопруд, Песков, 2003; Thorson, 1958). Однако, это не единственная выявленная закономерность. Климат, рельеф местности, характер растительности и другие элементы ландшафта также существенно воздействуют на структуру большинства типов сообществ. Еще большее влияние может оказывать биогеографический аспект: в ряде регионов, особенно на островах, таксономический состав бентосных беспозвоночных настолько своеобразен, что классического замещения близкородственных видов не наблюдается, и экологические ниши перехватываются неожиданными для них таксонами. Крайний вариант такой перестройки сообществ описан для арх. Шпицберген в Арктике, где почти все структурные типы донных сообществ в текучих и стоячих водах составлены из различных видов сем. Chironomidae. Это может быть следствием экстремальных условий арктической среды, но также резко обедненной островной фауны. Понятно, что эти факторы могут взаимно усиливать друг друга, но представляет особый интерес рассмотрение их действия по отдельности.
Экстремальные условия среды, близкие к высокоарктическим и перигляциальным, могут наблюдаться в любом регионе, где есть высокие горы с ледниками. В качестве примера мы рассматриваем один из высокогорных регионов Центральной Азии – горный массив Западный Тянь-Шань в пределах Киргизии. Помимо наличия высокогорных ландшафтов с ультрахолодноводными ледниковыми водотоками, он интересен как типичный для всей Центрально-азиатской биогеографической подобласти, реофильные сообщества которой до сих пор не были описаны нами, за исключением небольшой серии высокогорных проб из Южных Гималаев (Chertoprud et al., 2018). Учитывая засушливый континентальный климат, общее преобладание безлесных ландшафтов, повсеместное развитие скотоводства и существенную специфику таксономического состава макробентоса, сообщества подобласти могут существенно отличаться от таковых других регионов Палеарктики. Большой интерес представляет также исследование высотно-температурной поясности сообществ в регионе, где можно наблюдать все переходы условий – от высокогорных субнивальных до почти равнинных, близких к субтропическим.
Исследования фауны реофильных беспозвоночных водотоков Тянь-Шаня (как и большинства горных районов Азии) немногочисленны, к настоящему времени видовые определения доступны далеко не для всех таксонов (особенно много пробелов в изучении личинок насекомых). Экологические особенности бентосных организмов и их сообществ изучали здесь также не слишком активно. Несколько работ посвящено описанию бентосных сообществ водотоков Тянь-Шаня и окрестных равнин, с акцентом на предгорные и равнинные реки (Овчинников, 1936; Янковская, 1948; Кустарева, Иванова, 1980). Они кратко описывают состав и количественную характеристику лито-, пело- и псаммореофильных биоценозов. Особенности сообществ макробентоса горных потоков Тянь-Шаня (на примере двух рек Северного Прииссыккулья) обсуждаются в монографии К.А. Бродского (1976), но это анализ специфики типичного горного потока (очень быстрой малой горной реки), а не разнообразия местных сообществ. Ряд работ посвящен зональному и биотопическому распределению отдельных таксонов: некоторых двукрылых (Бродский, Оморов, 1972; Кустарева, Иванова, 1979), поденок (Оморов, Хаитов, 1992), веснянок (Оморов, 1977), ручейников (Оморов, 1975) и водных насекомых в целом (Бродский, 1935; Бродский, Оморов, 1973). Другие известные работы имеют таксономический или фаунистический характер.
Цель работы – на собственном материале описать многообразие сообществ реофильной макрофауны Западного Тянь-Шаня и проанализировать особенности сообществ для региона в целом и для высокогорных условий в частности.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
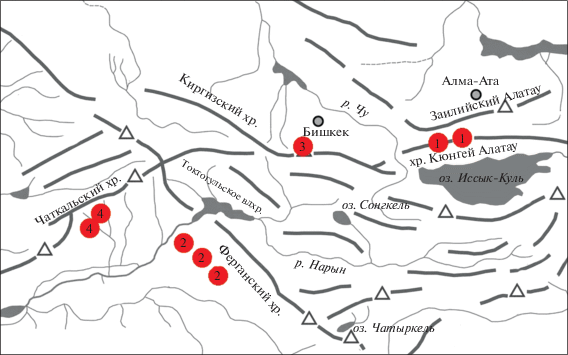
Район исследования. Материал собран в 127 водотоках разных районов Республики Киргизия, обследованных маршрутным методом. Cъемки охватили хребты Кюнгей Алатау (маршрут Каинды–Чолпон-Ата, август 2014 г., ~120 проб), Киргизский хребет (район Бишкек-Ала-Арча, август 2017 г., 40 проб), Ферганский хребет (маршрут Кара-Куль–Арсланбоб, август 2017 г., ~180 проб) и Чаткальский хребет (район Чаткал–Сары-Челек, май 2016 г., 30 проб) (рис. 1). Захвачены высотные зоны от 1000 до 3700 м над уровнем моря, обследованы все доступные размерные классы водотоков – от родников и ручьев первого порядка до рек шириной 15–20 м (Чон-Кемин, Карасу и др.).
Рис. 1.
Схема Западного Тянь-Шаня. Районы сбора проб: 1 – хр. Кюнгей Алатау, 2 – Ферганский хр., 3 – Киргизский хр., 4 – Чаткальский хр.
Высотная изменчивость ландшафтов на Тянь-Шане сглажена низкой лесистостью. В целом низкогорье в регионе можно охарактеризовать как засушливую степь с фрагментами зарослей кустарников и садов; среднегорье – степь с фрагментами лесов из ели Шренка; высокогорье – субальпийские и альпийские луга, постепенно замещающиеся к вершинам гор осыпями, скалами и ледниками. В одном районе (вокруг пос. Арсланбоб на южном макросконе Ферганского хребта) на высотах 1200–1800 м распространен широколиственный лес из грецкого ореха (вероятно, вторичный). В целом местность на высотах 1000, 2000 и 3000 м может выглядеть сходно – как склоновое пастбище овец и коров. Летом бо́льшая часть скота пасется в верхних эшелонах пастбищ, где трава растет лучше (местами до высоты 3500 м), а к зиме спускается в низкогорья.
Кроме того, на более высоких участках хребтов в долинах (высота от 3500–4000 м) располагаются многолетние ледники. Большинство водотоков на таких участках имеет ледниковые истоки, но некоторые ручьи уже на приледниковой морене образуют небольшие озера, другие уходят в толщу моренных отложений и выходят из нее вторичными родниками. Таким образом, даже вблизи ледников генезис и характер водотоков может заметно различаться. Ниже, вдали от ледников, большинство малых водотоков имеет родниковое питание, но могут встречаться реки, берущие начало в ледниковом высокогорье. Водотоки с ледниковым питанием обычно отличаются мутной, более холодной водой и характерной суточной динамикой водорасхода (максимум во второй половине дня, минимум к утру). Максимальная летняя температура в изученных водотоках варьировала от 0°С в ледниковых истоках до 22°С в ручьях низкогорий.
Методы сбора и обработки проб. Пробы отбирали полусферическим скребком с площадью 0.02 м2 и размером ячеи 1 мм раздельно для каждого типа биотопа – грунта, глубины и скорости течения. Каждая проба объединяла, как правило, 5 или 10 подпроб с одного биотопа на дистанции до пяти метров. В общей сложности собрано ~370 количественных проб. При описании местообитаний измеряли основные гидрологические характеристики: ширину и водорасход водотока, локальную глубину и скорость течения, тип субстрата, заиленность, освещенность, минерализацию и температуру воды.
Бо́льшую часть организмов определяли до уровня родов с помощью серии Определителей (Определитель…, 1994–2006), другую часть – до уровня видов с помощью ряда специальных работ по отдельным таксонам (Мартынов, 1935; Тесленко, Жильцова, 2009; Kluge, 2015). В качестве основного показателя обилия таксонов при выделении типов сообществ применяли относительный метаболизм, рассчитанный на основе численности и биомассы таксонов по стандартной формуле (Алимов, 1979).
Выделение типов сообществ. Принятая нами схема разделения сообществ на типы основана на методе Браун–Бланке (Braun–Blanquet, 1964), распространенном в геоботанике, но оперирующем с количественными данными. Исходной таблицей данных служила таблица относительного метаболизма определенных таксонов (роды или виды), достигающих 5% сообщества по обилию хотя бы в одной пробе. В этом массиве данных мы последовательно выявляли комплексы таксонов со сходным распространением, либо одиночные таксоны, доминирующие в скольких-то станциях, и группировали соответствующие станции. Эти группы рассматривали как потенциальные типы сообществ. Далее проводилась экологическая интерпретация этих групп: привязка их к биотопу, выявление обусловливающих их распространение факторов и оценка внутренней изменчивости. Если группа проб была достаточно устойчива по таксономической структуре, проводилось описание типа сообщества: его состав и структура доминирования, биотопическая приуроченность, вычисление средних показателей сообщества и т.д.
Номенклатура сообществ. Названия типов сообществ в данной работе – составные, как и в наших предыдущих работах (Chertoprud, 2011; Chertoprud, Palatov, 2013, 2017). Конечная часть слова обозначает крупный класс сообществ (креналь, ритраль, пелаль, рипаль), а префиксы – специфические факторы абиотической среды, определяющие данный тип, либо – сверхдоминирующий эврибионтный таксон. Как правило, корни в названии терминов имеют греческое происхождение и уже употребляются в гидробиологической литературе. Префикс “эу-” обозначает центральное сообщество данного класса, не связанное с особыми факторами, префикс “эпи-” – специфику, связанную с малым размером водотока. В остальных случаях ниже приводится этимология названий сообществ.
Высотно-температурное зонирование. В настоящей работе мы отказались от априорного выделения высотно-температурных зон и провели тотальную классификацию имеющихся данных. Тем не менее, для большинства выделенных типов сообществ (кроме двух) обнаружено четкое разделение на несколько вариантов, расходящихся, главным образом, по температурному признаку. В соответствии с этим, выделены три частично пересекающихся зоны, которым соответствуют разные варианты каждого типа сообществ (табл. 1–3).
Таблица 1.
Структура доминирования (% общего метаболизма) сообществ макрозообентоса низкогорной зоны Тянь-Шаня на уровне родов
| Тип сообщества | Макротаксон | Вид | % |
|---|---|---|---|
| Креналь | |||
| Гаммарокреналь* | AM | Gammarus | 76.4 |
| E | Baetis | 5.2 | |
| Ритраль | |||
| Эуритраль | TR | Hydropsyche | 45.0 |
| E | Baetis | 16.9 | |
| D | Tetisimulium | 11.2 | |
| D | Tianshanella | 5.6 | |
| E | Iron | 5.4 | |
| Эпиритраль | E | Baetis | 28.7 |
| P | Agnetina | 23.8 | |
| TR | Agapetus | 11.7 | |
| TR | Rhyacophila | 11.2 | |
| TR | Glossosoma | 6.1 | |
| D | Odagmia | 6.0 | |
| Химароритраль | D | Tianshanella | 49.7 |
| D | Tetisimulium | 15.1 | |
| E | Caucasiron | 8.0 | |
| Ch | Eukiefferiella | 6.2 | |
| E | Iron | 5.8 | |
| Ch | Diamesa | 5.3 | |
| E | Himalogena | 5.1 | |
| Лентиритраль | TR | Agapetus | 31.4 |
| E | Afghanurus | 14.5 | |
| TR | Glossosoma | 13.5 | |
| TR | Hydropsyche | 7.8 | |
| E | Drunella | 7.6 | |
| E | Baetis | 7.1 | |
| Фиторитраль | TR | Brachycentrus | 33.0 |
| D | Odagmia | 29.1 | |
| E | Baetis | 24.4 | |
| Пелаль | |||
| Псаммопелаль | Ch | Polypedilum | 73.1 |
| D | Hexatoma | 12.1 | |
| Эпипелаль | OL | Tubifex | 60.5 |
| Ch | Macropelopia | 6.2 | |
| Ch | Micropsectra | 5.9 | |
| Ch | Pagastia | 5.5 | |
| O | Cordulegaster | 5.2 | |
| Рипаль | |||
| Эурипаль | E | Baetis | 29.3 |
| TR | Limnephilus | 18.7 | |
| O | Cordulegaster | 12.5 | |
| P | Agnetina | 7.5 | |
| C | Agabus | 7.3 | |
| AM | Gammarus | 7.2 | |
| D | Dixa | 6.2 | |
| Эфемераль | |||
| Креноэфемераль* | C | Anacaena | 18.2 |
| C | Agabus | 16.1 | |
| C | Helophorus | 13.7 | |
| C | Laccobius | 9.3 | |
| Ch | Macropelopia | 7.6 | |
| GA | Galba | 5.8 | |
Примечание. Здесь и в табл. 2, 3 указана средняя доля каждого рода в сообществе по метаболизму, %. Даны роды со средней долей в сообществе ≥5%. Макротаксоны: BI – Bivalvia, C – Coleoptera, GA – Gasrtopoda, OL – Oligochaeta, AM – Amphipoda, Ch – Diptera Chironomidae, D – прочие Diptera, E – Ephemeroptera, P – Plecoptera, TR – Trichoptera, TU – Turbellaria. * Данные относятся и к среднегорной зоне.
Таблица 2.
Структура доминирования (% общего метаболизма) сообществ макрозообентоса среднегорной зоны Тянь-Шаня на уровне родов
| Тип сообщества | Макротаксон | Вид | % | Тип сообщества | Макротаксон | Вид | % |
|---|---|---|---|---|---|---|---|
| Ритраль | Креналь | ||||||
| Эпиритраль | E | Baetis | 20.8 | Гипокреналь | D | Tipula | 21.8 |
| Ch | Diamesa | 12.2 | P | Mesonemura | 19.5 | ||
| D | Tetisimulium | 8.9 | TU | Seidlia | 8.9 | ||
| P | Mesoperlina | 6.5 | E | Baetis | 8.4 | ||
| E | Iron | 5.7 | P | Amphinemoura | 7.1 | ||
| Химароритраль | D | Metacnephia | 44.8 | TR | Dinarthrum | 5.4 | |
| D | Asioreas | 21.1 | E | Ameletus | 5.3 | ||
| E | Baetis | 7.6 | Реокреналь | TR | Allomyia | 52.5 | |
| Ch | Iron | 6.4 | TR | Pseudostenophylax | 13.5 | ||
| D | Diamesa | 6.1 | E | Ameletus | 7.0 | ||
| E | Deuterophlebia | 5.3 | Ch | Pagastia | 6.2 | ||
| Химароэпиритраль | E | Iron | 46.4 | TU | Seidlia | 6.1 | |
| E | Caucasiron | 10.7 | Мадикреналь | D: | Thaumalea | 36.5 | |
| E | Baetis | 7.5 | D: | Oxycera | 15.7 | ||
| D | Tetisimulium | 5.2 | P: | Mesonemura | 8.5 | ||
| Лентиритраль | TR | Glossosoma | 54.3 | TU: | Seidlia | 8.5 | |
| E | Baetis | 9.1 | D: | Dixa | 6.9 | ||
| TU | Seidlia | 8.4 | D: | Tipula | 5.3 | ||
| Фиторитраль | E | Baetis | 40.3 | Пелаль | |||
| D | Tetisimulium | 18.6 | Эпипелаль | Ch | Pagastia | 49.8 | |
| P | Amphinemura | 8.0 | Ch | Micropsectra | 13.9 | ||
| D | Schoenbaueria | 7.2 | OL | Isochaetides | 11.0 | ||
| D | Montisimulium | 6.7 | Кренопелаль | Ch: | Macropelopia | 46.2 | |
| Бриоритраль | E | Baetis | 24.6 | BI | Euglesa | 11.1 | |
| P | Mesoperlina | 11.5 | Ch | Micropsectra | 6.6 | ||
| TR | Dinarthrum | 11.3 | Ch | Endochironomus | 5.2 | ||
| D | Tipula | 6.4 | Рипаль | ||||
| E | Ameletus | 5.2 | Эурипаль | E | Ameletus | 44.2 | |
| TU | Seidlia | 5.1 | E | Baetis | 12.7 | ||
| TR | Rhyacophila | 5.0 | Ch | Pagastia | 6.3 | ||
| Псефоритраль | TR | Dinarthrum | 49.4 | P | Mesoperlina | 5.3 | |
| D | Atherix | 8.9 | D | Tipula | 5.2 | ||
| AM | Gammarus | 6.9 | Гаммарорипаль | AM | Gammarus | 60.7 | |
| D | Tipula | 5.0 | E | Ameletus | 8.8 | ||
| Мадиритраль | TR | Tinodes | 64.7 | TR | Dinarthrum | 7.1 | |
| D | Thaumalea | 7.8 | E | Baetis | 5.5 | ||
| D | Pericoma | 6.7 | P | Mesonemura | 5.0 | ||
| Ch | Orthocladius | 5.6 | |||||
| Ch | Metriocnemus | 5.2 | |||||
Таблица 3.
Структура доминирования сообществ макрозообентоса высокогорной зоны Тянь-Шаня на уровне родов
| Тип сообщества | Макротаксон | Вид | % |
|---|---|---|---|
| Риталь | |||
| Криориталь | Ch | Diamesa | 34.9 |
| TR | Himalopsyche | 19.4 | |
| E | Iron | 12.5 | |
| P | Eucapnopsis | 7.0 | |
| P | Mesyatsia | 5.3 | |
| E | Baetis | 5.2 | |
| Ультракриориталь | Ch | Diamesa | 97.9 |
| Креналь | |||
| Криокреналь | Ch: | Diamesa | 53.4 |
| Ch | Metriocnemus | 17.4 | |
| D | Dicranota | 7.7 | |
| Ch | Chaetocladius | 6.7 | |
| D | Tipula | 6.0 | |
| Ch | Pagastia | 4.9 | |
| Пелаль | |||
| Криопелаль | OL | Enchytraeidae | 25.2 |
| Ch | Chaetocladius | 13.7 | |
| Ch | Orthocladius | 11.5 | |
| Ch | Pagastia | 11.4 | |
| Ch | Diamesa | 9.6 | |
| Ch | Rheocricotopus | 5.2 | |
Зона 1 (низкогорная): абсолютные высоты ~1000–2000 (в среднем 1740 м), летняя температура воды ~13–20°С (в среднем 15.3°С), 34 пробы.
Зона 2 (среднегорная): высоты ~1500–2800 (в среднем 2120) м, летняя температура 7–14 (в среднем 10.7°С), 282 пробы.
Зона 3 (высокогорная): высоты ~2500–3700 (в среднем 3100) м, летняя температура 0–8 (в среднем 4.3°С), 60 проб.
Видно, что основная часть материала относится к среднегорной зоне, в которой расположено большинство природных водотоков региона (табл. 2). В низкогорной зоне реки выходят на равнину и часто деградируют под антропогенным воздействием (табл. 1). Водотоки высокогорной зоны труднодоступны, при этом весьма однородны и бедны макробентосом (табл. 3). Поскольку эти зоны выделены эмпирически, на основе выделенных вариантов сообществ, границы между ними нечеткие: иногда в пробах с одной высоты и со сходными значениями температуры встречались сообщества разных вариантов, и наоборот. Абсолютизировать влияние как высоты, так и температуры воды нельзя, так как температура воды зависит не только от высоты, также влияние оказывает существенная суточная и погодная динамика (в измерениях не учитывалась).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Основные классы сообществ
В работах по различным регионам Палеарктики (Chertoprud, 2011; Chertoprud, Palatov, 2013, 2017; Chertoprud et al., 2018; Palatov, Chertoprud, 2012, 2018) мы выделили пять основных классов реофильных сообществ макрозообентоса, имеющих наиболее глубокие различия уже на уровне жизненных форм макрофауны, и довольно четко очерченных биотопически. Это креналь – сообщества ручьев и родников на мелкомозаичных субстратах, ритраль – сообщества плотных субстратов на быстром течении, пелаль – сообщества мягких грунтов, фиталь – сообщества зарослей макрофитов без явного течения, рипаль – сообщества мозаичных береговых субстратов. Этого разделения мы будем по возможности придерживаться и в данной работе.
Типизация данных по сообществам Тянь-Шаня позволила выделить 23 типа сообществ, шесть из них включают по два варианта, приуроченных к первой и второй высотно-температурным зонам. Остальные типы не обнаружили контрастные варианты в разных высотных зонах, либо принадлежали к одной зоне. Все сообщества третьей зоны оказались специфичны по структуре и выделены в собственные типы. В общей сложности к выделенным нами типам относится 357 описаний сообществ, еще 22 специфичных по структуре пробы типизировать не удалось.
Из выделенных нами 23 типов сообществ 11 относятся к классу ритрали, пять – к кренали, четыре – к пелали, два – к рипали. Класс фитали не представлен, но еще один тип сообществ отнесен в нетипичный для водотоков класс эфемерали (временных водоемов), более подробно описанный далее. Ниже приведено краткое описание всех типов: местообитания, доминирующие и характерные таксоны, распространение в других регионах. Из-за отсутствия определения многих видов, а также для компактности изложения описания сообществ в основном приводятся на уровне родов.
Сообщества ритрали
Класс сообществ, развивающихся на плотных грунтах водотоков, с явными адаптациями обитателей к течению. В горах Тянь-Шаня наиболее типичный класс, собрано 180 описаний из 11 (с учетом высотных вариантов – 15) типов. Обычно занимает большую часть площади дна водотоков, кроме маленьких ручьев и родников.
Эуритраль. Сообщество каменистых перекатов рек с хорошо освещенным руслом при течении, как правило, 0.3–0.6 м/с. На Тянь-Шане описано для немногих относительно крупных водотоков в первой (низкогорной) зоне (высота <1800 м). Имеет типичную для Палеарктики структуру: доминируют ручейники рода Hydropsyche, из субдоминантов типичны поденки Baetis, мошки (Tetisimulium, Metacnephia), гептагениды (род Iron), наиболее специфично появление эндемичных блефарицерид Tianshanella monstruosa (Brodskij, 1930). Близкие по структуре сообщества описаны нами для Гималаев (Chertoprud et al., 2018), где также характерны для предгорий и низкогорий (с субтропическим и тропическим климатом).
Эпиритраль. Сообщество каменистых перекатов малых рек и ручьев со слабо развитыми обрастаниями на камнях, при скорости течения 0.2–0.6 м/с. Встречается в регионе очень часто, описано для первой и второй высотных зон. Как и в других регионах, комплекс доминантов довольно обширен, включает реофильных поденок (Baetis, Heptageniidae), мошек и хирономид, различных веснянок, ручейников семейств Rhyacophilidae и Glossosomatidae. При увеличении высоты сообщество закономерно меняется: появляются холодноводные хирономиды рода Diamesa, сменяется состав ручейников, веснянок и мошек. В целом, сообщество первой зоны структурно близко к эпиритрали большинства регионов Палеарктики. Сообщество второй зоны специфичнее, но близко к таковым горно-лесного пояса Кавказа и Малой Азии (Palatov, Chertoprud, 2018).
Химароритраль (от греч. hiemarros – бурный поток). Сообщество экстремальных для бентоса скоростей течения (обычно 0.8–1.5 м/с) в реках. Встречается на Тянь-Шане нередко, в горных реках может занимать большую часть дна, оттесняя другие типы сообществ ритрали к берегам и отмелям. Описано для первой и второй зон, в обоих преобладают двукрылые из Simuliidae (типично для химароритрали многих регионов) и Blephariceridae встречается реже, но описано, в частности, для Кавказа и Балкан). Доминирующие роды обоих семейств сменяются в разных высотно-температурных зонах (табл. 1, табл. 2).
Химароэпиритраль. Вариант предыдущего сообщества, характерный для порогов в малых водотоках (в том числе с нестабильным грунтом), по-видимому, неблагоприятных для малоподвижных симулиид и блефарицерид. Описан для второй зоны, доминируют поденки, главным образом гептагенииды Iron gr. montanus и Caucasiron gr. guttatus. Тип известен для среднегорной зоны Гималаев (Chertoprud et al., 2018) и для Кавказа (Palatov, Chertoprud, 2018).
Лентиритраль (от греч. lentos – медленный). Сообщество каменистых грунтов при умеренном течении (обычно 0.1–0.4 м/с), в спокойной прозрачной воде. Характерно для водотоков с родниковым питанием, описано для первой и второй зон. Как и в других регионах, доминируют панцирные малоподвижные соскребатели – на Тянь-Шане это главным образом ручейники сем. Glossosomatidae (родов Glossosoma и Agapetus), среди прочих видов разнообразны поденки и веснянки (табл. 1, табл. 2). На Тянь-Шане род Agapetus более тепловоден и доминирует в первой зоне (как в тропиках и субтропиках Гималаев), а Glossosoma – во второй (как обычно на Кавказе).
Фиторитраль (от греч. phyton – растение). Сообщество макрофитов и затопленной наземной травянистой растительности при быстром и умеренном течении. На Тянь-Шане встречается изредка и только на затопленной траве, описано для первой и второй высотных зон. Имеет типичный для этого типа небольшой комплекс доминантов Baetis + Simuliidae (разные виды в разных регионах), в первой зоне к ним добавляется полуприкрепленный ручейник-хвататель Brachycentrus (Oligoplectrodes) sp. (род Brachycentus – характерный, но обычно не массовый в этом сообществе таксон).
Бриоритраль (от греч. bryon – мох). Сообщество, населяющее водные мхи на умеренном и быстром течении. В регионе встречается чаще предыдущего, в нем также доминируют поденки рода Baetis, но почти нет мошек и много щелевых ползающих таксонов (веснянки Mesoperlina, ручейники Dinarthrum и т.п.). В большинстве регионов отмечается редко, описано как минорный тип для Кавказа (Palatov, Chertoprud, 2018) и как массовый – для арктических районов: п-ова Ямал (Palatov, Chertoprud, 2012) и Кольского п-ова (Chertoprud, Palatov, 2013a).
Псефоритраль (от греч. psefos – галька). Сообщество гравийно-галечного и каменисто-галечного грунта рек. На Тянь-Шане встречается изредка, описано для второй зоны, среди макробентоса доминирует мелкий щелевой ручейник Dinarthrum (сем. Lepidostomatidae). Подобная структура описана для среднегорной зоны Гималаев (Chertoprud et al., 2018), в других регионах она существенно отличается.
Мадиритраль (от греч. madido – мокрый). Своеобразный тип сообществ тонкослойных водопадов и сливов по валунам (обычно в ручьях) при глубине от 1–2 мм до 3–5 см и скорости течения ≤0.3 м/с. Встречается редко, описано для второй зоны. Доминирует червеобразный галерейный ручейник Tinodes sp. (сем. Psychomiidae), иногда покрывающий прикрепленными домиками почти всю поверхность скалы. Как минорный тип сообщество наблюдалось для Кавказа, иногда смешивалось с мадикреналью.
Криоритраль (от греч. kryos – холодный). Специфичное по структуре сообщество холодных горных потоков высокогорной третьей зоны (температура воды 3–9, в среднем 5.5°С). Сообщество таксономически обеднено и практически лишено фильтраторов, а также панцирных альгофагов-соскребателей, в комплекс доминантов входят хирономиды Diamesa, хищные ручейники Himalopsyche, веснянки Eucapnopsis и Mesyatsia, поденки Iron и Baetis. Из известных нам ранее типов сообщество ближе всего к высокогорному сообществу эпиритрали Южных Гималаев (Chertoprud et al., 2018).
Ультракриоритраль. Крайне обедненное сообщество ультрахолодноводных приледниковых водотоков в высокогорной зоне (температура воды 0–4.5, в среднем 2.7°С). Полностью сформировано несколькими видами хирономид рода Diamesa, другие таксоны обычно отсутствуют. Данный тип описан для ледниковых водотоков арх. Шпицберген (Chertoprud et al., 2017) и горных ручьев Кольского п-ова (Chertoprud, Palatov, 2013).
В целом, класс ритральных сообществ в горах Тянь-Шаня развит хорошо, и региональные особенности природы (континентальность и засушливость климата, безлесность и скотоводство) сказываются на нем слабо. Налицо все типичные для ритрали жизненные формы и семейства макробентоса. Наличие субнивальных высокогорных условий дает два специфичных типа сообществ – криоритраль и ультракриоритраль, отсутствующих в большинстве других регионов.
Сообщества кренали
Класс сообществ, развивающихся на мозаичных субстратах малых водотоков (ручьев и родников), обычно с преобладанием детритофагов-разгрызателей. В пробах встречается часто, дано 80 описаний, отнесенных к пяти типам сообществ.
Гаммарокреналь. Сообщество малых постоянных водотоков и родников различных типов с большой плотностью и резким доминированием (обычно 60–90% по метаболизму) бокоплавов рода Gammarus (табл. 2), представленных в регионе большим количеством локально распространенных эндемичных видов, близких к G. turanus (Martynov, 1935). Другие таксоны не играют заметной роли. Встречается в первой и второй высотных зонах (отмечено на высоте ≤2360 м), не обнаруживая явной высотной изменчивости (хотя с высотой встречается реже). В низкогорье – наиболее типичное кренальное сообщество. Этот тип распространен по всей Южной Палеарктике (Chertoprud, Palatov, 2017; Palatov, Chertoprud, 2018).
Гипокреналь. Сообщество малых ручьев с мозаичным донным субстратом, где отсутствуют или редки бокоплавы (возможно, вследствие низкой температуры воды или большой вероятности зимнего промерзания). Описано для второй высотной зоны (табл. 2). Доминирование неустойчиво, обычно преобладают веснянки сем. Nemouridae (Mesonemoura, Amphinemura) и долгоножки (Tipula). В отличие от гипокренали большинства изученных нами регионов, малочисленны ручейники-лимнефилиды, которых лишь отчасти замещают Lepidostomatidae (род Dinarthrum).
Реокреналь. Сообщество холодных (5–8°С), но постоянных родниковых ручьев с заметным течением (0.1–0.2 м/с) и мозаичным грунтом. Описано для второй зоны. Структура своеобразна, преобладают ручейники: литофильные соскребатели Allomyia (сем. Apataniidae) и лимнефилиды рода Pseudostenophylax (табл. 2). Интересно, что Apataniidae – наиболее характерные обитатели этого типа сообщества, но редко доминируют в нем, обычно уступая по обилию лимнефилидам.
Мадикреналь. Сообщество “пленочных” водотоков: как правило, мокрых камней по краям родников. Описано для второй зоны. Доминируют в основном специфические мадидные двукрылые с воздушным дыханием: Thaumalea, Oxycera, Dixa, а также общекренальные двукрылые, веснянки и планарии (табл. 2). Сходные по структуре сообщества известны из самых разных регионов: от Европейской России (Chertoprud, 2011) до Гималаев (Chertoprud et al., 2018).
Криокреналь. Сообщество очень холодных (2–7, в среднем 4.6°С) малых ручьев со смешанным питанием и субстратами в третьей высотной зоне. Представлено, главным образом, хирономидами, из которых доминирует криофильный род Diamesa, также характерны Metriocnemus, Chaetocladius, Pagastia, из других групп присутствуют долгоножки Tipula и педицииды Dicranota (табл. 3).
Класс кренали выглядит несколько обедненным по разнообразию типов – многие варианты не встречены, несмотря на большое количество проб. Подобная ситуация имела место для арктических регионов (Кольский п-ов, п-ов Ямал) и может быть связана с общей суровостью климата (вызывающей зимнюю элиминацию сообществ кренали), а также безлесностью территорий и отсутствием в ручьях листового опада. С другой стороны, наблюдается и типичная черта южной части Палеарктики – оккупация большинства кренальных биотопов бокоплавами рода Gammarus и сообществом гаммарокренали, что также снижает распространение других типов сообществ.
Сообщества пелали
Класс сообществ мягких (илисто-песчаных) грунтов в водотоках различного типа, обычно с преобладанием инфауны. В горах относительно минорный класс сообществ, дано 53 описания, отнесенные к шести типам.
Псаммопелаль (от греч. psammon – песок). Сообщество песчаного грунта в реках при скорости течения 0.2–0.4 м/с. Описано для первой зоны, встречается на Тянь-Шане редко (5 описаний). Доминируют псаммофильные хирономиды рода Polypedilum, характерны также болотницы Hexatoma и ряд малочисленных хирономид (табл. 1). Подобные сообщества встречаются в большинстве регионов Евразии, но в данном случае их структура существенно обеднена (отсутствуют двустворчатые моллюски и олигохеты, состав двукрылых также беден).
Эпипелаль. Сообщество илистых, илисто-детритных и илисто-песчаных грунтов малых рек и ручьев. Встречается нередко (30 описаний), описано для первой и второй зон, в которых существенно различается. В первой зоне доминируют олигохеты рода Tubifex (сем. Tubificidae), среди субдоминантов преобладают различные хирономиды (табл. 1, табл. 2). Во второй зоне довольно резко доминирует крупная хирономида рода Pagastia, характерны также хирономиды Micropsectra и тубифициды Isochaetides, другие таксоны не достигают заметного обилия. В Евразии сообщества эпипелали, обычно с преобладанием хирономид и горошинок, описаны для многих регионов (Chertoprud, 2011; Chertoprud, Palatov, 2013, 2017; Palatov et al., 2016).
Кренопелаль. Сообщество илисто-детритного дна в родниках и родниковых ручьях без существенного течения, описано для второй зоны, встречается изредка. Преобладает хирономида Macropelopia, также характерны другие хирономиды и горошинки рода Euglesa (табл. 2). Близкие варианты известны для других регионов Палеарктики (Chertoprud, 2011; Palatov et al., 2016), но Macropelopia как первый доминант отмечается впервые.
Криоэпипелаль. Сообщество илисто-детритного дна холодных (2–8, в среднем 4.5°С) водотоков (в основном ручьев и родников) третьей высотной зоны. Доминирование неустойчиво, характерны олигохеты в целом почвенного сем. Enchytraeidae и ряд холодноводных хирономид. По-видимому, сообщество имеет эфемерный характер, элиминируя зимой, как и другие высокогорные сообщества. В сходных сообществах Кавказа и Кольского п-ова доминируют, главным образом, хирономиды рода Pseudodiamesa (Chertoprud, Palatov, 2013; Palatov et al., 2016), реофильные сообщества с преобладанием энхитреид отмечены в ручьях арх. Шпицберген (Chertoprud et al., 2017) и в рипали холодных водотоков Камчатки (Чебанова, 2009).
В целом класс пелали Тянь-Шаня значительно беднее по разнообразию сообществ и таксонов, по сравнению с большинством других горных регионов. В нем отсутствуют многие типичные пелофильные таксоны (перловицы, роющие поденки и ручейники, стрекозы), почти все сообщества представлены только хирономидами и олигохетами. Роль олигохет при этом повышается, что может быть связано и с отсутствием в таких сообществах ряда хищных таксонов, богатых биогенным питанием водотоков (за счет массового скотоводства), и пониженной температурой воды, обеспечивающей благоприятный кислородный режим даже при высокой органической нагрузке. В центрально-азиатских равнинных водотоках (Ферганская и Чуйская долины) сообщества пелали значительно богаче и содержат специфические группы крупных двустворчатых моллюсков (Anodonta) и роющих поденок (Palingenia) (Овчинников, 1936; Янковская, 1948).
Сообщества рипали
Сообщества мозаичных, главным образом органических субстратов береговой кромки водотоков. Встречаются нередко (52 описания), но занимают небольшую площадь.
Эурипаль. Сообщество мозаики органических субстратов береговой кромки – подмываемых водой корней, наземной травы при умеренном течении. Описано для первой и второй зон, встречается довольно часто (табл. 1, табл. 2). В первой зоне наиболее массовы общепалеарктические рода Baetis и Limnephilus, стрекоз представляет род Cordulegaster (центрально-азиатский вид C. coronata Morton, 1916). Эуритраль первой зоны близка по структуре с таковой равнинной Палеарктики, отличаясь, главным образом, отсутствием равнокрылых стрекоз. Во второй зоне доминируют холодноводные плавающие поденки Ameletus, что типично, например, для эурипали Кольского п-ова (Chertopud, Palatov, 2013) и высокогорной зоны Гималаев (Chertoprud et al., 2018).
Гаммарорипаль. Сообщество тех же биотопов, но с резким доминированием бокоплавов рода Gammarus, отчасти вытесняющих другие таксоны (табл. 2). Подобные варианты известны для многих регионов (Chertoprud, Palatov, 2017; Palatov, Chertoprud, 2018).
Сообщества эфемерали
Предварительно описываемый класс сообществ, в целом, не реофильный и характерный для временных стоячих водоемов, в основном, дождевых и снеговых луж. Подобным водоемам и их сообществам посвящена довольно обширная литература (например, Williams, 2006). Основные свойства этих сообществ: сезонность (существуют несколько недель или месяцев в году), быстрая динамика температуры, кислородного режима и сапробности, преобладание животных с короткими жизненными циклами (Chironomidae, Culicidae, Cladocera, Cyclopoida) или способных расселяться по воздуху (Coleoptera, Heteroptera). В некоторых случаях подобные сообщества могут развиваться и в слаботекучих водах, поэтому они нашли место среди наших данных.
Креноэфемераль. Сообщество луж со слабой родниковой подпиткой, которая создает некоторую проточность и пониженную температуру воды летом, но, по-видимому, не спасает от зимнего промерзания водоема. На Тянь-Шане подобные водоемы, как правило, служат водопоями скота и характеризуются повышенной сапробностью. Субстраты включают ил, глину, детрит, полузатопленную наземную растительность. Доминируют жесткокрылые, главным образом водолюбы и плавунцы (роды Anacaena, Laccobius, Helophorus, Agabus, Hydroporus), характерны также некоторые хирономиды (Macropelopia) и малый прудовик Galba. Данный тип не описывался нами ранее, хотя подобные сообщества иногда наблюдались в различных регионах.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Зоогеографические замечания
Взгляды на положение Тянь-Шаня в зоогеографических построениях и схемах различны, однако большинство из них рассматривает его в составе Центрально-азиатской подобласти, фауна которой связана своим происхождением с Гималаями (Иззатулаев, Старобогатов, 1985; Banarescu, 1992). Действительно, на уровне подродов и групп видов реофильных амфибиотических насекомых здесь обнаруживается немало сходства с другими регионами Центральной Азии (Гиссаро-Алай, Памир, Гиндукуш) и даже с южными склонами Гималаев. Так, от Тянь-Шаня вплоть до Северной Индии широко распространены поденки Ameletus gr. alexandrae Brodsky, 1930, Baetis (Rhodobaetis) gr. oreophilus Kluge, 1982, Baetiella muchei (Braasch, 1978), Iron gr. montanus, I. (Ironopsis) gr. rheophilus (Brodsky, 1930), Rhithrogena (Himalogena) spp.; веснянки Kyphopteryx spp., Mesoperlina spp., Mesyatsia tianshanica (Zhiltzova, 1972), Capnia gr. pedestris Kimmins, 1946, Eucapnopsis stigmatica Okamoto, 1922; ручейники Himalopsyche gr. gigantea (Martynov, 1914), Rhyacophila obscura Martynov, 1927, Pseudostenophylax micraulax (McLachlan, 1878); ряд видов амфибиотических двукрылых (Жильцова, 2003; Chertoprud et al., 2018; Gupta, Majumdar, 2013; Makarchenko et al., 2018). Впрочем, некоторые группы насекомых, обычные для Гиссаро-Алая (например, представители рода Tenuibaetis), здесь, по-видимому, отсутствуют. Доля широко распространенных, транспалеарктических видов в реофильной фауне региона очень мала. Из амфибиотических насекомых удалось обнаружить лишь два вида Baetis (B. buceratus Eaton, 1870 и B. rhodani (Pictet, 1843)), оба – в теплых водотоках с озерным питанием на территории заповедника Сары-Челек.
Фауна реофильных амфипод Тянь-Шаня слагается преимущественно рядом эндемичных, морфологически близких представителей рода Gammarus, видовой статус многих из которых нуждается в ревизии (Мартынов, 1935; Sidorov et al., 2018; Zhao et al., 2017). Помимо своеобразного эндемика оз. Иссык-Куль Issykogammarus hamatus Chevreux, 1908, отсюда известно два вида стигобионтных бокоплавов Stygobromus kazakhstanica Kulkina, 1992 и Tadzocrangonyx setiferus (Birstein et Ljovushkin, 1972). Последний принадлежит к центрально-азиатскому роду, ареал которого охватывает Тянь-Шань и Гиссаро-Алай (Sidorov et al., 2018). Однако, на Тянь-Шане не обнаружен целый ряд характерных центрально-азиатских родов Gammaridae (например, Sarothrogammarus, Comatogammarus или Barnardiorum), обычных уже в родниках Гиссарского и Зеравшанского хребтов.
Реофильные гастроподы на Тянь-Шане представлены лишь тремя эндемичными видами кренальных гидробиид, обитающими в низкогорьях северного склона. Это Martensamnicola kazakhstanica Izzatullaev, Sitnikova et Starobogatov, 1985 из родников близ г. Шымкент, Sibirobythinella almaatina Izzatullaev, Sitnikova et Starobogatov, 1985, описанная с Заилийского Алатау (род известен также на юге Сибири) и Chirgisia alaarchaensis Glöer, Boeters et Pešić, 2014, эндемичная для Киргизского хребта и встречающаяся в реокренах долины р. Ала-арча в составе гаммарокренальных сообществ (Glöer et al., 2014). Центральные районы Тянь-Шаня, видимо, лишены специфических реофильных гастропод, они появляются лишь в низкогорьях Алайского хребта (Valvatamnicola archangelskii (Zhadin, 1952), их разнообразие значительно увеличивается к юго-западу.
Таким образом, для водотоков Тянь-Шаня характерна типичная для Центральной Азии реофильная фауна, несколько обедненная по сравнению с Гиссаро-Алаем и более южными регионами за счет ряда родов ракообразных и моллюсков. Возможно, так проявляются последствия значительного оледенения Тянь-Шаня во времена последних ледниковых эпох.
Особенности сообществ региона: высотная зональность
При обсуждении особенностей реофильных сообществ региона, на наш взгляд, уместно разбить данные на выделенные ранее высотно-температурные зоны. Несмотря на внешнее сходство ландшафтов и водотоков в низко-, средне- и высокогорной зонах, состав и структура сообществ в них существенно варьируют.
Низкогорная зона включает довольно разнообразные сообщества, хотя материала по ним оказалось немного. Структура сообществ проявляет слабую специфику – во многих регионах Палеарктики (и равнинных, и горных) комплекс доминирующих родов в соответствующих типах сообществ примерно сходен. Особенно много параллелей выявлено с горными регионами Средиземноморья – Кавказа и Балкан. Налицо характерное преобладание в кренали бокоплавов рода Gammarus, стандартный комплекс доминантов в ритрали (Hydropsyche – Baetis – Simuliidae – Heptageniidae) и структурно бедная пелаль, представленная хирономидами и олигохетами. В равнинных регионах ритраль, как правило, беднее, пелаль богаче таксонами и сообществами, появляется фиталь.
Среднегорная зона объединяет большую часть собранного материала и большинство выделенных типов сообществ. Структурно сообщества этой зоны наиболее близки к таковым других холодноводных горных регионов (Кольский п-ов, Байкальский хребет, высокогорная зона Гималаев). От горных регионов с более мягким климатом (и от низкогорной зоны Тянь-Шаня) эти сообщества отличаются заметным таксономическим обеднением (выпадает много более тепловодных родов и семейств), появлением небольшого ряда новых таксонов (например, Ameletus в рипали) и довольно резким, во многих случаях, доминированием одного–двух таксонов. При этом типизация их также меняется: метод выделения комплексов совместно встречающихся видов редуцируется до классификации по первому доминанту. Возрастает сходство разных типов сообществ, особенно на уровне семейств, за счет увеличения роли Chironomidae, Simuliidae, Baetidae и Ameletidae.
Высокогорная зона Представлена криофильными сообществами, связанными с очень холодными приледниковыми ручьями. В трех типах сообществ резко доминируют различные хирономиды (наиболее типичен род Diamesa), в одном – также олигохеты-энхитреиды. Сходная структура сообществ (хотя с доминированием различных видов хирономид рода Diamesa) описана для водотоков арх. Шпицберген (Chertoprud et al., 2017), но также Альп и ряда других высокогорных районов (Milner et al., 2001). По-видимому, определяющим фактором для этих сообществ является очень низкая (до 4–5°С) летняя температура воды; хотя рассматриваются и другие факторы (Jacobsen, Dangles, 2012; Ilg, Castella, 2006; Milner, Petts, 1994). Описания видов хирономид рода Diamesa на Тянь-Шане и Памире, в том числе по нашим данным, еще только начались (Makarchenko et al., 2018), и многие личинки пока не определимы до вида.
Вопрос о региональных особенностях реофильных сообществ макробентоса Тянь-Шаня не имеет однозначного ответа: сообщества разных высотно-температурных зон близки по структуре к таковым в совершенно различных регионах. Низкогорная зона Тянь-Шаня примерно соответствует теплоумеренным и субтропическим регионам (Причерноморье, Балканы, среднегорные Гималаи), среднегорная зона Тянь-Шаня – низкоарктическим регионам (Кольский п-ов, п-ов Ямал), высокогорная зона – высокоарктическим регионам (арх. Шпицберген). По нашему мнению, ключевой структурирующий фактор в данном случае – летняя температура воды. Действительно, три описанные выше градации примерно соответствуют зонам летней температуры: 0–5, 5–12 и 12–20°С. Действие температуры, разумеется, может дополняться ландшафтными особенностями – характером увлажнения, растительности, почв, антропогенной нагрузки, сезонностью и т.п. Кроме того, высотные диапазоны одних и тех же температурных зон с характерными для них особенностями сооб-ществ закономерно меняются в зависимости от географической широты и общего климата региона. Например, для сообщества эурипали зона доминирования холодноводных поденок рода Ameletus в Гималаях находится на высотах 2000–3000 м (возможно, и выше), на Тянь-Шане – на 1500–2800 м, на Байкальском хребте – 500–1500 м, на Кольском п-ове – 300–1000 м.
Таким образом, действие высотной и широтной поясности для горных сообществ реофильного макробентоса в общих чертах сходно и опосредовано летней температурой воды. По мере нарастания как географической широты, так и абсолютной высоты, в горах температура падает, приводя к сходным изменениям сообществ. Наибольшее сходство наблюдается между высокоарктическими и высокогорными сообществами ледниковых ручьев, за счет резкого упрощения их структуры (повсеместно доминируют хирономиды одного рода Diamesa). Близкие эффекты уже были описаны для наземных растительных сообществ как “правило предварения” (Вальтер, Алехин, 1936). Для пресноводной биоты эти закономерности рассматривались лишь на уровне видового разнообразия (Jacobsen, Dangles, 2012). Впрочем, детали формирования структурно сходных сообществ в высокогорьях и Арктике еще предстоит исследовать.
Выводы. Реофильные сообщества донных беспозвоночных Западного Тянь-Шаня имеют особенности, типичные для горных регионов, а именно: большое разнообразие и встречаемость типов ритрали и кренали, угнетение типов пелали и рипали и отсутствие фитали. Структура большинства типов сообществ Западного Тянь-Шаня сильно варьирует в зависимости от высотно-температурных зон. В низкогорной зоне сообщества структурно сходны с большинством регионов Южной Палеарктики, в среднегорной зоне – с рядом холодноводных горных и гипоарктических регионов, в высокогорной приледниковой зоне (при резком таксономическом обеднении) – с сообществами водотоков Высокой Арктики (арх. Шпицберген). Ключевым фактором этой градации является, по-видимому, летняя температура воды. Характерно отсутствие в регионе многих типичных для Палеарктики реофильных таксонов и резкое доминирование одного-двух видов во многих сообществах. Фаунистически регион относится к Центральной Азии, представляя собой ее несколько обедненный северный форпост.
Список литературы
Алимов А.Ф. 1979. Интенсивность обмена у водных пойкилотермных животных // Общие основы изучения водных экосистем. Ленинград: Наука. С. 5.
Бродский К.А. 1935. Материалы к познанию фауны беспозвоночных горных потоков Средней Азии. 1. Река Иссык // Труды Среднеазиатского гос ун-та. Сер. VIIIa, Зоология. Вып. 15. С. 5.
Бродский К.А. 1976. Горный поток Тянь-Шаня. Эколого-фаунистический очерк. Ленинград: Наука.
Бродский К.А., Оморов Э.О. 1972. Распределение личинок комаров сем. Blepharoceridae и Deuterophlebiidae (Diptera) р. Акбура (Алайский хребет) // Энтомол. обозр. Т. 51. Вып. 1. С. 66.
Бродский К.А., Оморов Э.О. 1973. Вертикальная зональность горных потоков Тянь-Шаня по распределению характерных водных насекомых // Гидробиол. журн. Т. 9. № 2. С. 40.
Вальтер Г., Алехин В.В. 1936. Основы ботанической географии. Москва: Биомедгиз.
Жильцова Л.А. 2003. Веснянки (Plecoptera). Группа Euholognatha. Санкт-Петербург: Наука.
Иззатулаев З.И., Старобогатов Я.И. 1985. Зоогеографическая характеристика пресноводных моллюсков Центральной Азии и вопрос о существовании Нагорноазиатской подобласти Палеарктики // Зоол. журн. Т. 64. №. 4. С. 506.
Кустарева Л.А., Иванова Л.М. 1980. Бентос притоков озера Иссык-Куль. Фрунзе: Илим.
Кустарева Л.А., Иванова Л.М. 1979. Особенности распределения личинок хирономид (Chironomidae) в реке Чолпон-ата (Иссык-Кульская котловина) // Ихтиологические и гидробиологические исследования в Киргизии. Фрунзе: Илим. С. 68.
Мартынов А.В. 1935. К познанию Amphipoda текучих вод Туркестана // Тр. Зоол. ин-та АН СССР. Т. 2. Вып. 2–3. С. 409.
Овчинников И.Ф. 1936. Гидрологический и гидробиологический обзор главнейших водоёмов долины реки Чу // Труды Киргизской комплексной экспедиции АН СССР. Т. 3. Вып. 1. С. 115.
Оморов Э.О. 1975. К экологии ручейников (Trichoptera) горных потоков Южного Тянь-Шаня (на примере фауны р. Ак-буры) // Энтомол. обозр. 1975. Т. 54. № 4. С. 773.
Оморов Э.О. 1977. К экологии веснянок (Plecoptera) горных потоков Южного Тянь-Шаня (на примере фауны реки Акбуры) // Ихтиологические и гидробиологические исследования в Киргизии. Фрунзе: Илим. С. 45.
Оморов Э.О., Хаитов Х.Х. 1992. Вертикальное распределение поденок (Ephemeroptera) в реке Ак-Бура и ее притоке Кичик-Алае (Алайский хребет) // Энтомол. обозр. Т. 71. № 1. С. 17.
Определитель пресноводных беспозвоночных России и сопредельных территорий. 1994–2006. Т. 1–6. Санкт-Петербург: Зоол ин-т РАН.
Тесленко В.А., Жильцова Л.А. 2009. Определитель веснянок (Insecta, Plecoptera) России и сопредельных стран. Имаго и личинки. Владивосток: Дальнаука.
Чебанова В.В. 2009. Бентос лососевых рек Камчатки. Москва: ВНИРО.
Чертопруд М.В., Песков К.В. 2003. Географические параллели организации литореофильных сообществ малых рек Восточной Европы и Северной Азии // Журн. общ. биол. Т. 64. № 1. С. 78.
Янковская А.И. 1948. Материалы к гидробиологии бассейна Кара-Дарьи // Изв. АН УзССР. № 1. С. 60.
Banarescu P. 1992. Zoogeography of Fresh Waters. V. 2. Aula Verlag: Wiesbaden.
Braun-Blanquet J. 1964. Planzensociologie. Wien; N.Y.: Springer-Verlag.
Chertoprud M. 2011. Diversity and classification of rheophilic communities of macrozoobenthos in middle latitudes of European Russia // Biology Bull. Rev. V. 1. № 3. P. 165. https://doi.org/10.1134/S2079086411030017
Chertoprud M., Palatov D. 2013. Rheophilic communities of macrobenthos of the Southwestern Kola Peninsula // Inland Water Biology. № 6. P. 289. https://doi.org/10.1134/S1995082913040056
Chertoprud M., Palatov D. 2017. Stream Macrozoobenthic Communities of the Eastern Balkans // Inland Water Biology. V. 10. № 3. P. 286. https://doi.org/10.1134/S1995082917030051
Chertoprud M., Palatov D., Dimante-Deimantovica I. 2017. Macrobenthic communities in water bodies and streams of Svalbard, Norway // J. Natural History. V. 51. № 47–48. P. 2809. https://doi.org/10.1080/00222933.2017.1395092
Chertoprud M., Palatov D., Chertoprud E. 2018. Rheophilic Macrozoobenthos Communities of the Southern Himalayas // Inland Water Biology. V. 11. № 4. P. 435. https://doi.org/10.1134/S1995082918040041
Glöer P., Boeters H., Pešić V. 2014. Freshwater molluscs of Kyrgyzstan with description of one new genus and species (Mollusca: Gastropoda) // Folia Malacologica. V. 22. https://doi.org/10.12657/folmal.022.009
Gupta I.J., Majumdar M. 2013. Species of the Order Trichoptera Represented in the National Zoological Collection of the Zoological Survey of India, Kolkata // Records of the Zoological Survey of India. V. 113. № 4. P. 181.
Jacobsen D., Dangles O. 2012. Environmental harshness and global richness patterns in glacier-fed streams // Global Ecology and Biogeography. V. 21. P. 647.
Ilg C., Castella E. 2006. Patterns of macroinvertebrate traits along three glacial stream continuums // Freshwater Biol. V. 51. P. 840. https://doi.org/10.1111/j.1365-2427.2006.01533.x
Kluge N.J. 2015. Central Asian mountain Rhithrogenini (Ephemeroptera: Heptageniidae) with pointed and ephemeropteroid claws in the winged stages // Zootaxa. V. 3994. № 3. P. 301. https://doi.org/10.11646/zootaxa.3994.3.1
Makarchenko E.A., Semenchenko A.A., Palatov D.M. 2018. New data on taxonomy and systematics of the genus Diamesa Meigen (Diptera: Chironomidae: Diamesinae) from Tien Shan and Pamir mountains, with description of two new species // J. Limnol. V. 77. P. 26. https://doi.org/10.4081/jlimnol.2018.1783
Milner A., Petts G. 1994. Glacial rivers: physical habitat and ecology // Freshwater Biol. V. 32. P. 295.
Milner A., Brittain J., Castella E., Petts G. 2001. Trends of macroinvertebrate community structure in glacier-fed rivers in relation to environmental conditions: a synthesis // Freshwater Biol. V. 46. P. 1833.
Palatov D., Chertoprud M. 2012. The Rheophilic fauna and Invertebrate Communities of the Tundra Zone: A Case Study of the Southern Yamal // Inland Water Biology. V. 5. № 1. P. 19. https://doi.org/10.1134/S1995082912010117
Palatov D., Chertoprud M., Frolov A. 2016. Soft-bottom macrobenthic fauna and assemblages in watercourses of Eastern Black sea coast mountainous regions // Inland Water Biology. V. 9. № 2. P. 150. https://doi.org/10.1134/S1995082916020140
Palatov D., Chertopud M. 2018. Macrozoobenthic communities of compact grounds in streams of eastern Black Sea region // Russ. J. Ecol. V. 49. № 1. P. 80. https://doi.org/10.1134/S1067413618010125
Sidorov D., Hou Z., Sket B. 2018. Three new remarkable amphipod species (Crustacea: Gammaridae) from springs and subterranean waters of Central Asia // Zootaxa. V. 4444. № 4. P. 437. https://doi.org/10.11646/zootaxa.4444.4.5
Thorson G. 1958. Parallel level-bottom communities, their temperature adaptation, and their “balance” between predators and food animals // Perspectives in marine biology. Berkely: Univ. California Press. P. 67.
Williams D. 2006. The Biology of Temporary Waters. New York: Oxford Univ. Press.
Zhao S., Meng K., Hou Z. 2017. Two new Gammarus species and a new name (Crustacea: Amphipoda: Gammaridae) from Northwest China // Zootaxa. V. 4273. № 2. P. 195. https://doi.org/10.11646/zootaxa.4273.2.3
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод