

Биология внутренних вод, 2020, № 3, стр. 232-242
Свободноживущие нематоды искусственных водоемов Вьетнама
В. Г. Гагарин a, *, Нгуен Динь Ты b
a Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
пос. Борок, Некоузский р-н, Ярославская обл., Россия
b Институт экологии и биологических ресурсов, Вьетнамская академия наук и технологий
Ханой, Вьетнам
* E-mail: gagarin@ibiw.yaroslavl.ru
Поступила в редакцию 26.02.2018
После доработки 01.08.2018
Принята к публикации 24.12.2018
Аннотация
В пробах из четырех искусственных водоемов во Вьетнаме обнаружено 62 вида свободноживущих нематод, из них один род и 11 видов – новые для науки. По численности доминировали Dichromadora geophila (de Man, 1876), Sabatieria foetida Gagarin, Nguyen Vu Thanh, 2008, Parodontophora fluviatilis Gagarin, Nguyen Vu Thanh, 2008 и Terschellingia longicaudata de Man, 1922. Приведены видовой состав нематод и иллюстрированное описание двух новых для науки видов из сем. Sphaerolaimidae: Parasphaerolaimus stagnalis sp. n. и Megalamphis vietnamicus sp. n.
ВВЕДЕНИЕ
Фауна свободноживущих нематод водоемов Вьетнама изучена сравнительно слабо. Только с начала XXI в. в связи с составлением банка данных по гидрофауне водоемов и водотоков Вьетнама стали интенсивно исследовать эту группу червей различных водных объектов.
Цель работы – изучить видовой состав свободноживущих нематод в искусственных водоемах Вьетнама.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
В 2014 и 2015 гг. пробы нематод отобраны в четырех искусственных водоемах (площадью от 1000 до 12 000 м2), используемых для выращивания двух видов креветок Penaeus monodon Fabricius, 1798 и Litopenaeus vannamei (Boone, 1931). Водоемы расположены в провинции Куанг Нинь (Qung Ninh) на острове в Южно-Китайском море вблизи побережья Вьетнама. Они густо заросли травами Halophila beccarii Ascherso, 1871 и Ruppia maritima L., 1953. Пробы отбирали на глубине 0.3–0.7 м с помощью цилиндра диаметром 3.5 см, длиной 10 см и промывали через сачок из газа с диаметром ячеи 0.08 мм. Пробы фиксировали горячим (60–70°С) 4%-ным раствором формалина. Затем пробы помещали в емкость объемом 200 мл, добавляли раствор Ludox-TM50 (1 : 1) и центрифугировали 5 раз по 40 мин. Нематод переводили в чистый глицерин по методу Зайнхорста (Seinhorst, 1959), затем монтировали в капле глицерина на предметных стеклах и опечатывали кольцом из парафина (воска). Для измерения, определения, фотографирования и изготовления рисунков червей использовали световой микроскоп Nicon Eclipse 80i, оборудованный принадлежностями для наблюдения методом ДИК-контраста, цифровой камерой Nikon DS-Fil и ПК, оснащенной программой NIS-Elements D 3.2 для анализа и документирования.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Фауна. В пробах из водоемов за два года исследования обнаружено 62 вида свободноживущих нематод, из них один новый для науки род Paradaptonema securum Gagarin, 2018 и 11 новых для науки видов (табл. 1). До вида не определены 12 групп червей, так как они представлены или малым количеством особей (1–4 экз.), или личиночными стадиями, или особями одного пола.
Таблица 1.
Видовой состав и количество особей нематод в пробах из искусственных водоемов Вьетнама
| Таксон | 2014 г. | 2015 г. |
|---|---|---|
| Отряд Enoplida Filipjev, 1929 | ||
| Сем. Anoplostomidae Gerlach, Riemann, 1974 | ||
| Anoplostoma dubium Gagarin, 2015 | 2♂♂, 8♀♀, 12juv | – |
| A. nhatrangensis Tchesunov, Nguyen Vu Thanh, 2010 | – | 4♂♂, 2♀♀, 5juv |
| Сем. Oxystominidae Chitwood, 1935 | ||
| Oxystomina minor Nguyen Dinh Tu et al., 2016 | 3♂♂, 8♀♀, 12juv | – |
| Halalaimus vietnamicus Gagarin, 2016 | 1♂, 5♀♀, 11juv | 4♂♂, 2♀♀, 2juv |
| H. gracilis de Man, 1888 | – | 1♂, 1♀, 3juv |
| H. durus Gagarin, Nguyen Vu Thanh, 2004 | – | 1♂, 2juv |
| Сем. Tripyloididae Filipjev, 1928 | ||
| Tripyloides minor Gagarin, Nguyen Vu Thanh, 2012 | 1♂, 1juv | – |
| T. stagnalis Gagarin, 2019 | – | 6♂♂, 5♀♀, 9juv |
| Сем. Oncholaimidae Filipjev, 1926 | ||
| Adoncholaimus minor Gagarin, Nguyen Dinh Tu, 2016 | 1♂, 1♀, 4juv | – |
| Viscosia timmi Gagarin, Nguyen Thi Thu, 2008 | 1♀, 1juv | 3♂♂, 1♀, 3juv |
| Сем. Enchelidiidae Filipjev, 1928 | ||
| Belbolla gracilis Gagarin, Nguyen Vu Thanh, 2016 | 1♀, 3juv | 5♂♂, 1♀, 2juv |
| Отряд Monhysterida Filipjev, 1929 | ||
| Сем. Monhysteridae de Man, 1876 | ||
| Thalassomonhystera parva (Bastian, 1885) | 7♂♂, 8♀♀, 11juv | 4♂♂, 7♀♀, 13juv |
| Th. pygmea Gagarin, 2012 | – | 10♂♂, 20♀♀, 27juv |
| Th. longisoma Gagarin, 2018 | – | 3♂♂, 8♀♀, 15juv |
| Th. sp. | – | 1♂, 3♀♀, 7juv |
| Monhystera sp. | 1♀, 2juv | – |
| Сем. Xyalidae Chtwood, 1920 | ||
| Daptonema securum Nguyen Thi Xuan Phuong et al., 2016 | 2♂♂, 4♀♀, 2juv | 8♂♂, 5♀♀, 9juv |
| D. pumilus Nguyen Vu Thanh et al., 2005 | 2♂♂, 3♀♀, 1juv | – |
| D. curvatum Gagarin, Nguyen Thi Thu, 2008 | – | 12♂♂, 7♀♀, 15juv |
| D. sp. | 3♀♀, 2juv | – |
| Paradaptonema securum Gagarin, 2018 | – | 9♂♂, 8♀♀, 17juv |
| Theristus gigas Gagarin, 2009 | – | 1♂, 7♀♀ |
| Steineria vietnamica Gagarin, 2013 | – | 4♂♂, 2♀♀, 2juv |
| Sphaerotheristus sp. | 1♂, 1♀, 4juv | – |
| Сем. Sphaerolaimidae Filipjev, 1918 | ||
| Sphaerolaimus pacificus Allgen, 1947 | 1♂, 3♀♀, 8juv | – |
| S. rivalis Gagarin, 2014 | 1♂, 5♀♀, 12juv | – |
| Parasphaerolaimus stagnalis Gagarin, Nguyen Dinh Tu sp. n. | – | 2♂♂, 2♀♀, 3juv |
| Megalamphis vietnamicus Gagarin, Nguyen Dinh Tu sp. n. | – | 3♂♂, 1♀, 3juv |
| Subsphaerolaimus sp. | 2♀♀, 2juv | – |
| Сем. Linhomoeidae Filipjev, 1922 | ||
| Terschellingia longicaudata de Man, 1922 | 58♂♂, 75♀♀, 106juv | 8♂♂, 7♀♀, 27juv |
| T. yenensis Gagarin, 2018 | 36♂♂, 38♀♀, 78juv | 38♂♂, 45♀♀, 18juv |
| T. sp. | 1♂, 11juv | 2♀♀, 3juv |
| Valvalaimoides leptus Gagarin, 2017 | 25♂♂, 33♀♀, 47juv | 7♂♂, 3♀♀, 6juv |
| Paraterschelingoides vietnamicus Gagarin, 2018 | 7♂♂, 7♀♀, 14juv | 22♂♂, 30♀♀, 32juv |
| Desmolaimus minor Gagarin, 2019 | 1♂, 2♀♀, 1juv | 2♂♂, 1♀, 3juv |
| Megadesmolaimus sp. | 4♀♀, 5juv | – |
| Отряд Araeolaimida de Coninck, Sch. Stekhoven, 1933 | ||
| Сем. Axonolaimidae Filipjev, 1918 | ||
| Paradontophora fluviatilis Gagarin, Nguyen Vu Thanh, 2006 | 50♂♂, 52♀♀, 89juv | 22♂♂, 38♀♀, 62juv |
| Pseudolella tenuis Gagarin, Nguyen Dinh Tu, 2018 | 9♂♂, 2♀♀, 12juv | 2♂♂, 6♀♀, 25juv |
| Сем. Comesomatidae Filipjev, 1918 | ||
| Sabatieria foetida Gagarin, Nguyen Vu Thanh, 2008 | 33♂♂, 34♀♀, 78juv | 62♂♂, 67♀♀, 135juv |
| Paracomesoma leptum Gagarin, Nguyen Vu Thanh, 2018 | 3♂♂, 10♀♀, 32juv | 13♂♂, 2♀♀, 3juv |
| Hopperia mira Gagarin, Nguyen Vu Thanh, 2006 | 1♂, 5juv | – |
| Сем. Diplopeldidae Filipjev, 1918 | ||
| Campylaimus gerlachi Timm, 1961 | 1♂, 1♀, 1juv | 1juv |
| Отряд Chromadorida Chitwood, 1933 | ||
| Сем. Cyatholaiimidae Filipjev, 1918 | ||
| Paracanthonchus securus Nguyen Dinh Tu, Gagarin, 2018 | 12♂♂, 23♀♀, 41juv | – |
| Paracanthonchus bravicaudatus Gagarin, Nguyen Vu Thanh, 2016 | 8♂♂, 10♀♀, 23juv | – |
| Marylinnia sp. n. | 1♂, 1♀, 2juv | – |
| Сем. Ethmolaimidae Filipjev, Sch. Stekhoven, 1941 | ||
| Paraethmolaimus minor Nguyen Dinh Tu, Gagarin, 2018 | 9♂♂, 5♀♀, 8juv | – |
| Сем. Neotonchidae Wieser, Hopper, 1966 | ||
| Gomphionema parvum Gagarin, Nguyen Vu Thanh, 2009 | 1♂, 15♀♀, 52juv | 5♂♂, 6♀♀, 27juv |
| Сем. Chromadoridae Filipjev, 1917 | ||
| Chromadorita orientalis Gagarin, Nguyen Vu Thanh, 1918 | 3♂♂, 1♀, 1juv | 9♂♂, 4♀♀, 7juv |
| Dichromadora geophila (de Man, 1876) | 86♂♂, 83♀♀, 144juv | 17♂♂, 15♀♀, 57juv |
| D. simplex Timm, 1961 | 8♂♂, 4♀♀, 1juv | 4♂♂, 9♀♀, 9juv |
| Rhips sp. | 1♀, 1juv | – |
| Spilophorella bidentata Platonova, 1971 | 9♂♂, 26♀♀, 28juv | 8♂♂, 9♀♀, 11juv |
| Actinonema dolichurum Gagarin, Phan Ke Long, 2017 | – | 33♀♀, 8juv |
| Сем. Selachinematidae Cobb, 1915 | ||
| Halichoanolaimus vietnamicus Gagarin, Phan Ke Long, 2017 | 2♂♂, 4♀♀, 8juv | – |
| H. sp. | 1♂, 1 juv | – |
| Отряд Plectida Malakhov, 1985 | ||
| Сем. Leptolaimidae Oerley, 1880 | ||
| Halaphanolaimus congiensis Gagarin, Nguyen Vu Thanh, 2007 | 11♂♂, 33♀♀, 25juv | 18♂♂, 30♀♀, 24juv |
| Antomicron intermedius Gagarin, Nguyen Vu Thanh, 2005 | 1♂, 2♀♀, 3juv | – |
| Сем. Camacolaimidae de Man, 1889 | ||
| Camacolaimus sp. | 6juv | 1♂, 3juv |
| Отряд Desmodorida de Coninck, 1965 | ||
| Сем. Desmodoridae Filipjev, 1922 | ||
| Pseudochromadora parva Gagarin, Nguyen Vu Thanh, 2008 | 5♂♂, 7♀♀, 14juv | 13♂♂, 27♀♀, 32juv |
| Metachromadoroides sp. | 1♂, 2♀♀, 5juv | – |
| Сем. Microlaimidae Micoletzky, 1922 | ||
| Microlaimus orientalis Gagarin, Nguyen Vu Thanh, 2011 | 5♂♂, 4♀♀, 8juv | – |
| Отряд Dismoscolecida Filipjev, 1922 | ||
| Сем. Desmoscolecidae Shipley, 1838 | ||
| Tricoma sp. | 3♂♂, 3♀♀, 4juv | – |
Выявленные виды входят в состав 7 отрядов и 21 семейства. Более разнообразная фауна представлена в пробах 2014 г. Наибольшее количество видов (25) относится к отр. Monhysterida, наименьшее (1 вид) – к отр. Desmoscolecida (табл. 1). Подавляющее количество видов – солоновато-водные формы нематод, что объясняется сравнительно высокой соленостью воды (12.9–15.3‰) в местах отбора проб и близостью моря. Наиболее высокое количество особей в пробах обнаружено у пяти видов: Paradontophora fluviatilis Gagarin, Nguyen Vu Thanh, 2008, Dicromadora geophila (de Man, 1876), Sabatieria foetida Gagarin, Nguyen Vu Thanh, 2008, Terschellingia longicaudata de Man, 1922, Terschellingia yenensis Gagarin, 2018 (табл. 1).
Описание видов. Отряд Monhysterida Filipjev, 1929. Семейство Sphaerolaimidae Filipjev, 1918. Род Megalaimophis de Coninck, 1965. Megalamphis vietnamicus Gagarin, Nguyen Dinh Tu sp. n. (рис. 1, 2).
Рис. 1.
Строение самца (а, б) и самки (в) Megalamphis vietnamicus sp. n.: a – голова, б, в – задний конец тела.
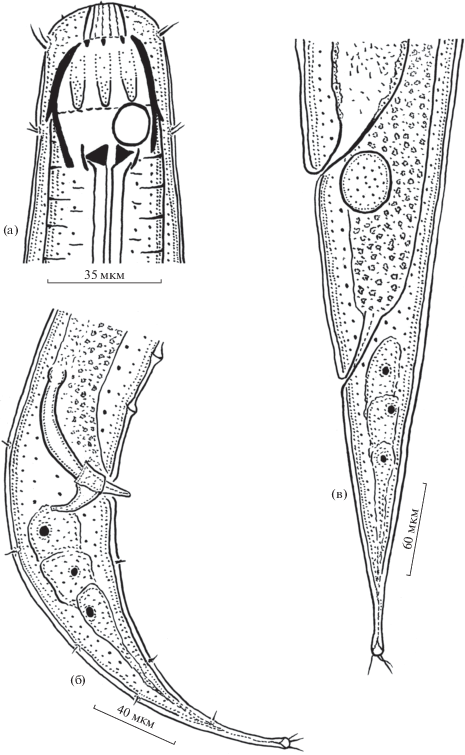
Рис. 2.
Самец (а, в, г, е, з, и) и самка (б, д, ж, к) Megalamphis vietnamicus sp. n.: a, б – общий вид, в, г, д – голова, е – передний конец тела, ж – тело в области вульвы, з – тело в области клоаки, и, к – задний конец тела.
Mатериал. Голотип ♂, инвентарный номер препарата PL 2.2.3; паратипы: 2♂♂, 1♀. Препараты голотипа и паратипов хранятся в коллекции Музея природы Вьетнамской академии наук и технологий (г. Ханой, Вьетнам).
Местонахождение. Вьетнам, провинция Куанг Нинь (Quang Ninh). Координаты: 20°48′18′′–20°48′84′′с.ш., 106°53′21′′–106°55′48′′ в.д.
Искусственный водоем для выращивания креветок, глубина 0.3–0.7 м, грунт – заиленный песок, соленость воды 12.9–15.3‰, сборы в июне 2014 г.
Описание. Морфометрическая характеристика голотипа и паратипов приведена в табл. 2.
Таблица 2.
Морфометрическая характеристика Megalamphis vietnamicus sp. n.
| Признак | Голотип ♂ | Паратипы | |
|---|---|---|---|
| 2♂♂ | 1♀ | ||
| L, мкм | 1154 | 1129, 1482 | 1466 |
| a | 15 | 12, 15 | 13 |
| b | 4.7 | 4.6, 5.4 | 5.5 |
| c | 7.3 | 6.8, 7.9 | 7.8 |
| c' | 3.1 | 3.0, 2.7 | 3.7 |
| V, % | – | – | 78.5 |
| Ширина, мкм: | |||
| области губ | 43 | 34, 34 | 43 |
| стомы | 32 | 32, 33 | 33 |
| тела в его среднем отделе | 78 | 92, 97 | 114 |
| тела в области ануса или клоаки | 51 | 54, 71 | 51 |
| Длина, мкм: | |||
| внешних губных щетинок | 4 | 4, 5 | 4 |
| головных щетинок | 6 | 7, 6 | 7 |
| субцефалических щетинок | 10 | 10, 12 | 11 |
| стомы | 48 | 48, 46 | 47 |
| фаринкса | 248 | 247, 275 | 267 |
| хвоста | 158 | 165, 187 | 187 |
| спикул (по дуге) | 85 | 82, 90 | – |
| дорсального отростка рулька | 22 | 19, 20 | – |
| Расстояние, мкм: | |||
| от фовеи амфидов до переднего конца тела | 32 | 31, 32 | 35 |
| от заднего конца фаринкса до вульвы | – | 717, 1020 | 884 |
| от заднего конца фаринса до клоаки | 748 | – | – |
| от вульвы до ануса | – | – | 128 |
| Диаметр фовеи амфидов, мкм | 13 | 12, 13 | 11 |
| Количество супплементарных органов | 8 | 8, 10 | – |
Самцы. Черви средней длины, сравнительно толстые. Кутикула тонко-кольчатая. Соматические щетинки редкие, длиной 2.0–3.5 мкм. Губы плохо выражены. Внутренние губные сенсиллы в форме папилл. Внешние губные сенсиллы, головные и субцефалические сенсиллы в форме тонких щетинок. Длина внешних губных щетинок 4–5 мкм, головных щетинок – 6–7 мкм, субцефалических щетинок – 10–12 мкм. Головные щетинки и субцефалические расположены в один круг. На уровне середины длины стомы расположены шейные щетинки длиной 5.0–6.0 мкм. Хейлостома маленькая, в форме воронки, со слабо кутикулизированными стенками и 12 продольными ребрами. Гимностома обширная, с сильно кутикулизированными стенками и разделена на два отдела. В переднем отделе гимностомы находятся шесть продольно расположенных толстых пластинок. Стегостома в форме узкого кольца, имеет три треугольных зубовидных образования. Общая длина стомы равна 1.0–1.4 ширины области губ. Фовеи амфидов в форме круга, диаметр которого 34–36% соответственно диаметру тела, расположены на уровне заднего отдела стомы. Фаринкс сравнительно длинный, мускулистый. Кардий маленький. Клетка ренетты и ее выводная пора не обнаружены. Семенники парные, противопоставленные. Спикулы изогнутые, с головками. Их длина в 1.3–1.7 раза превышает диаметр тела в области клоаки. Рулек состоит из основного тела и довольно длинного дорсального отростка, длина которого равна 0.2–0.3 длины спикул. Перед клоакой расположено 8–10 плохо заметных поровидных супплементов. Хвост удлиненно-конический. Каудальные щетинки имеются. На кончике хвоста находятся три субтерминальные щетинки длиной 13–15 мкм. Каудальные железы и спиннерета хорошо развиты.
Самки. По общей морфологии подобны самцам. Строение кутикулы и переднего конца тела как у самцов. Кутикула тонко-кольчатая. Внутренние губные сенсиллы в форме папилл. Внешние губные сенсиллы, головные сенсиллы и субцефалические сенсиллы в форме тонких щетинок. Головные и субцефалические щетинки расположены в один круг. На уровне середины стомы расположены шейные щетинки длиной 5.0–6.0 мкм. Хейлостома в форме маленькой воронки. Гимностома обширная, с сильно кутикулизированными стенками, разделена на два отдела. В переднем отделе 6 пластинок. Стегостома в форме узкого кольца и содержит три зубовидных образования. Общая длина стомы примерно равна диаметру области губ. Фовеи амфидов в форме круга, расположены на уровне заднего отдела стомы. Фаринкс сравнительно длинный, мускулистый. Длина ректума примерно равна диаметру тела в области ануса. Яичник один, передний, гомодромный, находится справа от кишки. Вульва постэкваториальная, в форме поперечной щели. Стенки вагины утолщены. Матка обширная, заполнена спермиями. Задняя матка отсутствует. Поствульварная железистая клетка присутствует. Хвост удлинено-конический. На кончике хвоста имеются три субтерминальные щетинки. Каудальные железы и спиннерета хорошо развиты.
Дифференциальный диагноз. Род Megalamphis содержит один вид – М. sphaerolaimoides (Timm, 1961), найденный в Бенгальском заливе (Timm, 1961), Megalamphis vietnamicus sp. n. отличается от него наличием субцефалических щетинок, относительно более короткими внешними губными щетинками (30–35% ширины области губ против 95% ширины области губ у M. sphaerolaimoides), иной формой спикул и рулька и наличием преклоакальных супплементарных органов (Timm, 1961).
Этимология. Видовое название означает “вьетнамский”, “из Вьетнама”.
Род Parasphaerolaimus Lorenzen, 1978. Parasphaerolaimus stagnalis Gagarin, Nguyen Dinh Tu sp. n.
Материал. Голотип ♂, инвентарный номер препарата CH 1.1.1a; паратипы: 1♂, 2♀♀. Препарат голотипа хранится в коллекции Музея природы Вьетнамской академии наук и технологий (г. Ханой, Вьетнам). Препараты паратипов хранятся в коллекции нематод Института экологии и биологических ресурсов (г. Ханой, Вьетнам).
Местонахождение. Вьетнам, провинция Куанг Нинь (Quang Ninh). Координаты: 20°48′18′′–20°48′84′′ с.ш., 106°53′21′′–106°55′48′′ в.д. Искусственный водоем для выращивания креветок, соленость воды 12.9–15.3‰, сборы в июне 2014 г.
Описание. Морфометрическая характеристика голотипа и паратипов приведена в табл. 3.
Самцы. Черви средней длины, сравнительно толстые. Кутикула тонко-кольчатая. Соматические щетинки редкие, длиной ~2 мкм. Губы плохо выражены. Внутренние губные сенсиллы в форме едва видных щетинок длиной 1.5–2.0 мкм. Внешние губные сенсиллы в форме тонких щетинок длиной 5–6 мкм. Четыре головные сенсиллы в форме щетинок длиной 5–6 мкм. В круг головных щетинок входят восемь субцефалических щетинок длиной 11–12 мкм. Четыре шейные щетинки длиной 5 мкм расположены на уровне середины стомы. Хейлостома сравнительно крупная, с 12 кутикулизированными ребрами. Гимностома обширная, с сильно кутикулизированными стенками, разделена на два отдела. В переднем отделе шесть небольших пластинок. В заднем отделе три едва заметные пластинки. Стегостома в форме широкой воронки. Длина стомы примерно равна ширине области губ. Фовеи амфидов в форме круга, расположены на уровне заднего конца стомы. Фаринкс мускулистый. Кардий сравнительно маленький. Семенники парные, противопоставленные. Спикулы тонкие и очень длинные. Их длина в 4.2–4.9 раза превышает ширину тела в области клоаки. Рулек длиной 44–46 мкм, в форме желоба, без дорсального отростка. Супплементарные органы отсутствуют. Хвост удлиненно-конический. Каудальные щетинки имеются. На кончике хвоста расположены три субтерминальные щетинки 12–13 мкм длиной. Каудальные железы и спиннерета хорошо развиты.
Таблица 3. Морфометрическая характеристика Parasphaerolaimus stagnalis sp. n.
| Признак | Голотип ♂ |
Паратипы | |
|---|---|---|---|
| 1♂ | 2♀♀ | ||
| L, мкм | 1834 | 1869 | 2593, 2230 |
| a | 18 | 20 | 25, 15 |
| b | 4.2 | 5.0 | 5.4, 4.7 |
| c | 8.3 | 8.8 | 10.0, 8.6 |
| c' | 3.2 | 3.6 | 4.9, 3.8 |
| V, % | – | – | 81.8, 80.9 |
| Ширина, мкм: | |||
| области губ | 46 | 43 | 43, 48 |
| стомы | 35 | 34 | 36, 38 |
| тела в его среднем отделе | 102 | 95 | 102, 144 |
| тела в области ануса или клоаки | 70 | 60 | 53, 68 |
| Длина, мкм: | |||
| внутренних губных щетинок | 1.5 | 2.0 | 1.5, 2.0 |
| внешних губных щетинок | 5 | 6 | 5, 5 |
| головных щетинок | 6 | 5 | 6, 5 |
| субцефалических щетинок | 12 | 11 | 12, 10 |
| стомы | 46 | 45 | 54, 53 |
| фаринкса | 435 | 371 | 476, 476 |
| хвоста | 221 | 213 | 258, 260 |
| спикул (по дуге) | 296 | 292 | – |
| рулька | 46 | 44 | – |
| Расстояние, мкм: | |||
| от фовеи амфидов до переднего конца тела | 26 | 25 | 28, 29 |
| от заднего конца фаринкса до вульвы | – | – | 1646, 1329 |
| от заднего конца фаринса до клоаки | 1178 | 1285 | – |
| от вульвы до ануса | – | – | 213, 165 |
| Диаметр фовеи амфидов, мкм | 12 | 12 | 10, 10 |
Примечаниe. Сокращения, как в табл. 2.
Самки. По общей морфологии подобны самцам. Строение кутикулы и переднего конца тела, как у самцов. Кутикула тонко-кольчатая. Внутренние губные сенсиллы в форме коротких, едва заметных щетинок. Шесть внешних губных сенсилл и четыре головные сенсиллы в форме тонких щетинок длиной 5–6 мкм расположены в два разобщенных круга. Восемь субцефалических щетинок длиной 10–12 мкм внедрены в круг головных щетинок. Четыре шейные щетинки находятся на уровне середины стомы. Хейлостома с 12 ребрами. Гимностома обширная, разделена на два отдела. В переднем отделе имеются шесть пластинок, в заднем – только три и они едва заметны. Стегостома в форме широкой воронки. Длина стомы немного больше ширины области губ. Фовеи амфидов в форме круга, расположены на уровне заднего конца стомы. Фаринкс мускулистый. Длина ректума примерно равна ширине тела на уровне ануса. Яичник один, передний, голодромный, расположен справа от кишки. Вульва постэкваториальная, в форме поперечной щели. Вагина наклонена к переднему концу тела, ее стенки утолщены. Задняя матка и поствульварная клетка отсутствуют. Хвост удлиненно-конический. Каудальные железы и спиннерета хорошо развиты.
Дифференциальный диагноз. Род Parasphaerolaimus Lorenzen, 1978 в настоящее время содержит восемь валидных видов (Гагарин, 2014; Zograf et al., 2017). Parasphaerolaimus stagnalis sp. n. отличается от всех видов наличием очень длинных и тонких спикул. Морфологически он наиболее близок к P. crassus (Timm, 1961), обнаруженному в Бенгальском заливе (Timm, 1961). Отличается от него более короткими головными и субцефалическими щетинками (их длина соответственно равна 5–6 и 10–12 мкм против 10 и 16–30 мкм длины у P. crassus), более далеко от переднего конца тела расположенной вульвой (80.9–81.8% против 63% у P. crassus) и более длинной спикулой (292–296 мкм против 51 мкм у P. crassus) (Timm, 1961).
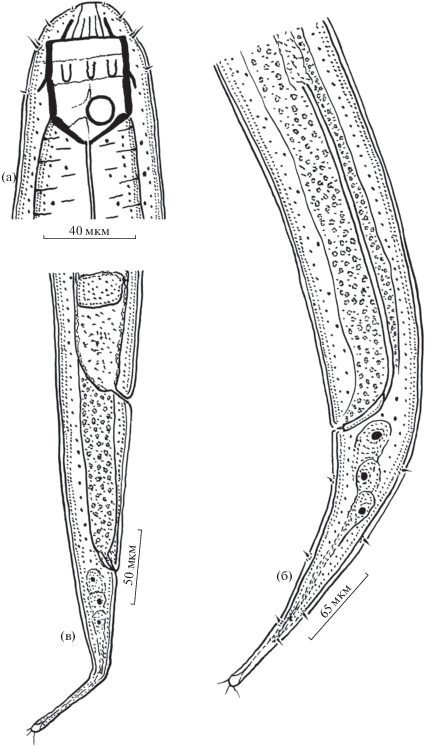
Рис. 3.
Строение самца (а, б) и самки (в) Parasphaerolaimus stagnalis sp. n.: a – голова, б, в – задний конец тела.
Этимология. Видовое название означает “прудовой”, “из пруда”.

Список литературы
Гагарин В.Г. 2014. Два новых вида свободноживущих нематод (Nematoda, Sphaerolaimidae) из мангровых зарослей в дельте реки Красной, Вьетнам // Амурский зоол. журн. Т. 6. № 1. С. 3.
Fonseca G., Bezerra T.N. 2014. Order Monhysterida Filipjev 1929 // Handbook of Zoology. V. 2. P. 435.
Seinhorst J.V. 1959. A rapid method for the transfer of nematodes from fixative to anhydrous glycerin // Nematologica. V. 4. P. 67.
Timm R.W. 1961. The marine nematodes of the Bay of Bengal // Proceedings of the Pakistan Academy of Science. V. 1. № 1. P. 1.
Zograf J.K., Pavlyuk O.N., Trebukhova Y.A., Nguyen Dinh Tu. 2017. Revision of the genus Parasphaerolaimus (Nematoda: Sphaerolaimidae) with description of new species // Zootaxa. V. 4232(1). P. 58. https://doi.org/10.11646/zootaxa.4232.1.4
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод