

Биология внутренних вод, 2020, № 5, стр. 495-501
Сезонная динамика соотношения типов реореакции у плотвы Rutilus rutilus
Д. С. Павлов a, В. Ю. Паршина a, *, В. В. Костин a
a Институт проблем экологии и эволюции им. А.Н. Северцова Российской академии наук
Москва, Россия
* E-mail: ponomareva_ipee@mail.ru
Поступила в редакцию 25.02.2020
После доработки 20.03.2020
Принята к публикации 26.03.2020
Аннотация
Исследована сезонная динамика мотивационной компоненты реореакции (соотношения типов рео-реакции) плотвы (Rutilus rutilus (L.)) Рыбинского водохранилища. Использована гидродинамическая установка ‒ кольцевой лоток с лимнозоной. Показано, что изменение соотношения типов реореакции у немигрирующей половозрелой плотвы адекватно отражает ее поведение в водохранилище. Выявлено слабое влияние пола рыб, степени зрелости их гонад и температуры воды на соотношение типов реореакции и стабильность их проявления.
ВВЕДЕНИЕ
Реореакция, или отношение рыб к течению, – это врожденная реакция, связанная с жизнью рыб в подвижной среде. Она проявляется в выборе рыбами участка водоема или водотока с оптимальными (для актуального поведения) скоростями течения и вектора собственного движения в нем. Проявление реореакции определяют три компоненты: ориентационная – восприятие стимула; мотивационная – выбор типа реореакции; локомоторная – реализация реореакции выбранного типа (Павлов, 1979; Павлов и др., 2020; Pavlov et al., 2010). К известным показателям мотивационной компоненты относятся: избираемые скорости течения в реоградиенте (Павлов, 1979), реопреферендум рыб – выбор течения или его отсутствия (Павлов и др., 2007), соотношение типов реореакции – соотношение экспериментальных оценок частот проявления реореакции разных типов (Павлов и др., 2020; Maclean, Gee, 1971).
Нами была разработана методика определения четырех типов реореакции: ПТР – движение против течения (реотаксис), ОТР – движение по течению (отрицательный реотаксис (Johnston et al., 2017)), КТР – сохранение своего положения относительно неподвижных ориентиров и ФТР – уход с потока в лимноусловия (лимнозону). В лимнозонах скорости течения ниже пороговых для реореакции или близки к ним, поэтому реореакция носит факультативный характер – течение не заставляет особь постоянно проявлять реореакцию для компенсации сноса (Павлов и др., 2020). Реореакцию этих типов мы наблюдаем (Павлов и др., 2019; Pavlov et al., 2010; Zvezdin et al., 2015) как в экспериментальных, так и в естественных условиях. Динамические типы реореакции преобладают у мигрирующих рыб: ПТР – при контранатантной миграции, ОТР – при денатантной (покатной) миграции; а статические – у немигрирующих: КТР – при сохранении места обитания (компенсации сноса) в речном потоке, ФТР – при нахождении рыб в зоне с пониженной скоростью течения или его отсутствия (лимнозоне).
На аквариумных рыбах данио рерио (Danio rerio Hamilton) показано, что эта методика позволяет более полно, чем при использовании рыбохода11, оценивать частоту проявления, прежде всего статических типов реореакции (Павлов и др., 2020). Для проверки этого положения на диких рыбах в данной работе использовали плотву из залива водохранилища. Так как мотивационное состояние рыб в естественных условиях может меняться в зависимости от целого ряда сезонных факторов, исследование проводили в течение трех сезонов – весны, лета, осени (с апреля по ноябрь).
Цель работы – экспериментально оценить сезонную динамику соотношения типов реореакции плотвы: определить основные факторы, влияющие на изменение реореакции плотвы в нерестовый период; оценить целесообразность использования методики определения не трех, а четырех типов реореакции.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Исследования проводили в апреле‒ноябре 2017 г. на прибрежной плотве, находящейся на разных стадиях зрелости гонад (табл. 1). Длина тела подопытных рыб тотальная (TL) составила 145–290 мм, стандартная (SL) ‒ 120–240 мм, возраст – от 3+ до 6+.
Таблица 1.
Объем и характеристика собранного материала (2017 г.)
| Месяц | Стадия зрелости гонад | Температура воды, °C | Пол | N | SL, мм | m, г | Коэффициент зрелости гонад |
|---|---|---|---|---|---|---|---|
| Апрель‒май | IV, V, VI–II | 3.0–11.0 | ♀ | 59 | 199.7 ± 2.1 | 163.9 ± 5.8 | 19.1 ± 1.1 |
| ♂ | 44 | 193.1 ± 2.4 | 141.1 ± 5.1 | 5.4 ± 0.4 | |||
| Июль | II–III | 17.0–17.5 | ♀ | 8 | 176.9 ± 6.1 | 114.7 ± 12.7 | 1.2 ± 0.1 |
| ♂ | 21 | 145.5 ± 4.8 | 56.6 ± 6.8 | 1.0 ± 0.2 | |||
| Август | III | 16.5–17.0 | ♀ | 17 | 166.2 ± 3.6 | 89.3 ± 6.8 | 5.5 ± 0.4 |
| ♂ | 12 | 161.7 ± 3.9 | 77.9 ± 6.0 | 2.1 ± 0.2 | |||
| Октябрь | III | 9.0–10.0 | ♀ | 19 | 158.2 ± 3.3 | 73.4 ± 5.0 | 9.0 ± 0.5 |
| ♂ | 10 | 147.5 ± 3.8 | 57.8 ± 5.1 | 2.7 ± 0.4 | |||
| Ноябрь | III–IV | 5.0–6.5 | ♀ | 16 | 163.1 ± 5.6 | 85.2 ± 9.4 | 10.1 ± 0.6 |
| ♂ | 12 | 157.1 ± 4.9 | 73.9 ± 8.5 | 4.6 ± 0.3 |
Рыб отлавливали ставными сетями и удочками на нерестилищах и местах откорма рыб – в середине залива Рыбинского водохранилища, образовавшегося в подпорной зоне рек Сунога и Шумаровка. Его ширина здесь была ~250–300 м, максимальная глубина, определяемая уровнем воды в водохранилище, увеличивалась от 2.0 м (15.04.2017) до 4.5 м (08.05.2017), в ноябре она медленно уменьшалась до 4.0 м.
Рыб (25–30 экз.) после отлова содержали без кормления в помещении в аэрируемом бассейне объемом воды 1.3 м3 в течение 1–2 сут до начала опыта. Вода в бассейн поступала из реки. Освещенность во время опытов (с 9:00 до 21:00) была 100–400 лк. В остальное время суток искусственное освещение в помещении, где проводили опыты и содержали рыб, выключали. Температуру в бассейнах поддерживали с помощью автоматических терморегуляторов в соответствии с температурой в реке во время отлова рыб.
Для экспериментов использовали воду из бассейнов, в которых содержали подопытных рыб, температура и освещенность в установке были такими же, как в бассейнах.
Экспериментальную установку ‒ кольцевой лоток с лимнозоной (Павлов и др., 2020) ‒ размещали в каркасном бассейне Intex Prism Frame 28710/26710. Дно лотка было сделано из покрашенной фанеры, стенки – из поликарбоната (рис. 1), высота стенок 50 см, внутренняя стенка на 33 см состояла из оргстекла, выше – из капронового сита, через которое вода стекала из рабочего коридора в общий бассейн; диаметр круга внешней стенки 300 см, внутренней – 240 см; ширина рабочего коридора по всей длине 30 см, длина коридора (по центральной линии) 850 см; глубина воды 40 см, объем всей воды в бассейне 5300 л, рабочий объем установки 1100 л. На стенки коридора были нанесены визуальные ориентиры в виде черных вертикальных полос шириной 1 см с промежутками в 8 см, на дне – границы секторов. Поток в рабочем канале создавали насосами. Точка подачи воды располагалась у внешней стенки. Обратный перелив воды происходил по всей длине внутренней стенки через сетку. Лимнозона (рис. 1, сектор 5) с шириной входа 50 см располагалась с внутренней стороны коридора.
Рис. 1.
Схема кольцевого гидродинамического лотка с лимнозоной: 1–5 – сектора установки (5 – лимнозона), Б – бассейн, Н – насос для создания течения, пунктирные линии – границы секторов и лимнозоны, стрелка – направление течения.
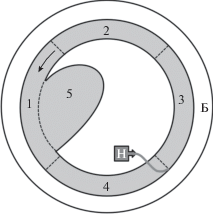
Скорость течения в лотке устанавливали в диапазоне крейсерских скоростей течения для плотвы – 43 см/с (2–4 длины тела особи в секунду). В рабочем канале установки создавался поперечный градиент скоростей течения (с максимальной скоростью у внешней стенки). Скорость течения измеряли при помощи специально разработанного измерительного комплекса, состоящего из датчика скорости (микровертушка с диаметром лопасти 8 мм), усилителя, преобразователя сигнала и контролера на базе однокристальной ЭВМ (Павлов и др., 2020).
Перед началом опыта лимнозону (стартовый сектор (5)) предварительно изолировали сеткой и в него помещали подопытную особь. Время акклимации было 20 мин, после чего сетку убирали. Далее на протяжении 15 мин визуально фиксировали время и направление пересечения рыбой границ секторов. Перемещения особи в пределах сектора не фиксировали.
Всего проведено 227 индивидуальных опытов.
После окончания опыта у каждой особи измеряли длину и массу тела; определяли пол, массу порки и гонад, визуально оценивали стадию зрелости гонад по универсальной шкале (Сакун, Буцкая, 1963); рассчитывали коэффициент зрелости гонад как отношение массы гонад к массе порки.
По экспериментальным данным рассчитывали частоты проявления разных типов реореакции, подвижность рыб и частоту смены типов реореакции.
Для объективного (не зависящего от оператора) способа оценки времени проявления КТР потребовалось использование дополнительного параметра – граничного времени (tg). Это расчетная оценка максимальной продолжительности прохода особью без остановок сектора кольцевого лотка (Павлов и др., 2020). Величину tg определяли по суммарной рабочей диаграмме частоты встречаемости времени, проведенного особями в секторе кольцевого лотка. Граничное время находили отдельно для движения рыбы по течению и против него. По диаграммам устанавливали время, при котором указанная частота резко падала до принятой нами величины – в ≥8 раз. Если такого падения частоты не наблюдалось при исходной величине классового интервала времени (0.5–1.0 с), его величину увеличивали до появления искомого резкого различия. Выполненные расчеты показали, что при перемещении рыб вверх по течению величина граничного времени равна 22 с, а вниз по течению – 14 с. Если рыба пребывала в отсеке свыше соответствующего значения граничного времени, мы принимали, что в течение этого времени (разница между реальным временем пребывания в отсеке и граничным временем) рыба проявляла КТР.
Частоту проявления разных типов реореакции (P) вычисляли по доле времени их проявления от длительности опыта:
РПТР = (ku tg + ∑tu+ ∑tr/2)/T,
РОТР = (kd tg + ∑td + ∑tr/2)/T,
РКТР = ∑(t – tg)/T,
РФТР = ∑tz/T,
где: ku – число переходов в верхний по течению сектор после проявления КТР; kd – число переходов в нижний по течению сектор после проявления КТР; T – длительность опыта, с; t – время пребывания в секторе длительностью >tg, с; td – время пребывания в секторе длительностью <tg, при сквозном проходе сектора по течению, с; tg – граничное время, с; tr – время пребывания в секторе длительностью <tg, при развороте (вход и выход через одну границу сектора); tu – время пребывания в секторе длительностью <tg, при сквозном проходе сектора против течения, с; tz – время пребывания в лимнозоне, с.
Подвижность рыб оценивали по общему пройденному пути (S), равному сумме путей, пройденных рыбами против (Su) и по течению (Sd): S = = Su + Sd.
Su = nu l + 0.5kl,
Sd = nd l + 0.5kl,
где k – число секторов, в которых особь изменила направление движения; l – длина центральной дуги сектора, м; nu и nd – число секторов установки, пройденных против и по течению.
Частоту смены типов реореакции рассчитывали как частное числа изменений типа реореакции на время опыта в мин. Чем меньше этот показатель, тем в среднем стабильнее у рыбы проявление реореакции исследованных типов.
Для статистического анализа использовали непараметрический дисперсионный анализ Краскела–Уоллиса, критерий Стьюдента для долей (Лакин, 1973) и регрессионный анализ. Соотношения типов реореакции достоверно различаются, как и любые соотношения, когда достоверно различаются частоты проявлений хотя бы одного из типов реореакции.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Сезонная динамика реореакции плотвы. Дисперсионный анализ показал, что в исследованный период изменения реореакции по месяцам наблюдения достоверны (p < 0.002, H критерий Краскела–Уоллиса) для всех показателей. При этом сезонная динамика показателей реореакции достоверно (pH < 0.005, H критерий Краскела–Уоллиса) различается у самцов и самок.
Во все исследованные периоды у плотвы преобладала реореакция статических типов (табл. 2, рис. 2), что указывает на отсутствие мотивации к миграции у исследованных рыб. Различия между самцами и самками наиболее резко были выражены в августе и октябре: у самок сохранялась стабильность показателей, у самцов она нарушалась (рис. 2). Во все месяцы, кроме ноября, подвижность самок превышала таковую у самцов – общий пройденный путь у самок в 1.3–3.4 раза длиннее, чем у самцов (табл. 2). За исключением периода нереста и ноября у самок частота проявления ОТР была больше, чем ПТР, у самцов наоборот, т.е. наиболее подвижные рыбы (самки) чаще двигались по течению, а менее подвижные (самцы) примерно одинаково двигались по и против течения. В ноябре у рыб обоих полов увеличилась частота проявления ФТР до 0.77 (самцы) и 0.97 (самки).
Таблица 2.
Сезонная динамика частоты проявления типов реореакции, частоты их смены и пройденного пути у плотвы Рыбинского водохранилища в апреле–ноябре 2017 г.
| Месяц | Пол | Число опытов | S, м | Типы реореакции | ЧСТР, мин–1 | |||
|---|---|---|---|---|---|---|---|---|
| динамические | статические | |||||||
| ПТР | ОТР | КТР | ФТР | |||||
| Апрель–май | ♀ | 59 | 71.2 ± 13.9 | 0.05 ± 0.01 | 0.20 ± 0.04 | 0.42 ± 0.05 | 0.33 ± 0.05 | 0.22 ± 0.03 |
| ♂ | 44 | 53.7 ± 9.1 | 0.07 ± 0.01 | 0.14 ± 0.03 | 0.54 ± 0.05 | 0.25 ± 0.05 | 0.47 ± 0.09 | |
| Июль | ♀ | 8 | 46.3 ± 10.6 | 0.10 ± 0.03 | 0.12 ± 0.03 | 0.28 ± 0.06 | 0.50 ± 0.09 | 0.68 ± 0.16 |
| ♂ | 21 | 31.1 ± 12.1 | 0.14 ± 0.07 | 0.06 ± 0.03 | 0.28 ± 0.09 | 0.51 ± 0.14 | 0.29 ± 0.10 | |
| Август | ♀ | 17 | 56.3 ± 17.0 | 0.09 ± 0.03 | 0.14 ± 0.04 | 0.25 ± 0.06 | 0.52 ± 0.08 | 0.71 ± 0.16 |
| ♂ | 12 | 16.5 ± 6.3 | 0.07 ± 0.04 | 0.04 ± 0.02 | 0.29 ± 0.09 | 0.60 ± 0.12 | 0.33 ± 0.11 | |
| Октябрь | ♀ | 19 | 50.7 ± 9.5 | 0.08 ± 0.02 | 0.16 ± 0.04 | 0.41 ± 0.07 | 0.35 ± 0.09 | 0.52 ± 0.10 |
| ♂ | 10 | 36.3 ± 13.6 | 0.13 ± 0.05 | 0.10 ± 0.05 | 0.44 ± 0.12 | 0.33 ± 0.15 | 0.36 ± 0.11 | |
| Ноябрь | ♀ | 16 | 5.6 ± 3.1 | 0.01 ± 0.01 | 0.02 ± 0.01 | 0.07 ± 0.04 | 0.91 ± 0.06 | 0.09 ± 0.06 |
| ♂ | 12 | 6.7 ± 3.0 | 0.00 ± 0.00 | 0.03 ± 0.03 | 0.20 ± 0.09 | 0.77 ± 0.09 | 0.18 ± 0.08 | |
Рис. 2.
Динамика частоты проявления статических (1) и динамических (2) типов реореакции у самок (а) и самцов (б) плотвы в апреле–ноябре 2017 г. Вертикальная линия – ошибка средней.
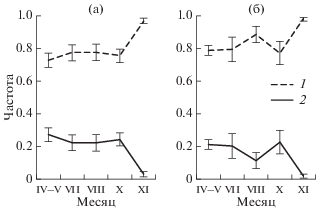
Залив Рыбинского водохранилища, в котором отлавливали рыб, мелководный, и его глубина зависит от уровня водохранилища. При низком уровне он превращается в узкое (2–10 м) русло р. Сунога. При таком падении уровня воды плотва совершает откочевку в более глубокие места водохранилища. В 2017 г. динамика уровня Рыбинского водохранилища проходила по второму типу (Буторин, 1963, 1969) – весной, летом, и осенью 2017 г. наполнение водохранилища было стабильным, близким к нормальному подпорному уровню (НПУ) (по данным РусГидро (www.rushydro.ru/hydrology/informer/?date=2017-03-18) падение уровня воды в водохранилище не превышало 77 см), и плотва все три сезона обитала в этом заливе. Наши наблюдения и отловы рыб не зафиксировали массовых перемещений (миграций) плотвы в исследованный период, что соответствует и литературным данным. Весной в Рыбинском водохранилище значительное время до и после нереста основная масса плотвы сосредоточена в прибрежной зоне и на прилегающей к ней пойме (Поддубный, 1966, 1971; Рыбопромысловый…, 1963; Рыбы…, 2015). Летом и осенью она также не совершает миграций. Для плотвы характерны только кочевки в поиске корма. Их интенсивность велика в мае–июне и снижается в июле–августе, в сентябре–октябре снова возрастает, а в ноябре опять резко снижается (Поддубный, 1971; Рыбопромысловый…, 1963). В целом прибрежная плотва в водохранилище ведет не мигрантный, а резидентный образ жизни.
Ранее показано, что для немигрирующих рыб, в том числе и плотвы, характерно преобладание статических типов реореакции над динамическими (Павлов и др., 2019; Pavlov et al., 2010). Именно такое соотношение типов реореакции мы наблюдали в 2017 г. Отмечены и более частные совпадения поведения плотвы в установке и водохранилище. В августе у самцов плотвы несколько снизилась частота проявления динамических типов реореакции, в октябре – повысилась (рис. 2б). Это совпадает с отмеченным выше снижением интенсивности питания в августе и его увеличением в октябре. В ноябре резко увеличивается частота проявления ФТР – плотва избегает течения (табл. 2). При этом существенно снижается частота смены типов реореакции (табл. 2), т.е. есть реореакция стабилизируется – из-за сильного преобладания одного ее типа. Это соответствует снижению подвижности рыб при зимовке – в начальный период зимовки распределение в целом очень похоже на осеннее, значительных миграций не происходит, в местах зимовки рыбы малоподвижны в пойме (Поддубный, 1971; Рыбопромысловый…, 1963; Рыбы…, 2015).
Таким образом, сезонная динамика соотношения типов реореакции, определенная в кольцевом лотке с лимнозоной, адекватно отражает сезонную динамику поведения плотвы в водохранилище – преобладание статических типов реореакции соответствует резидентному (немиграционному) поведению рыб.
Основные факторы, влияющие на реореакцию плотвы. В течение лета и осени у плотвы происходило постепенное созревание гонад, в местах нагула в заливе водохранилища изменялись температура воды, кислородный и уровенный режимы, обеспеченность рыб кормом и др. Проследить отдельно влияние каждого фактора на реореакцию плотвы по имеющимся данным невозможно, поскольку в один год наблюдений они меняются одновременно. Это удалось сделать только в период нереста (апрель–май), когда температура и созревание гонад менялись независимо друг от друга (при одной температуре воды были рыбы с разной степенью зрелости гонад). На IV стадии зрелости гонад особи находились в самом широком диапазоне температур – от 3 до 11.5°С, на V и VI–II стадиях – при температуре 8–8.5°С. Остальные факторы изменялись незначительно.
Дисперсионным анализом (H критерий Краскела–Уоллиса) установлено, что в период нереста соотношение типов реореакции различается у рыб разного пола (p < 0.001) с разной стадией зрелости гонад (p ≤ 0.007) и при разной температуре воды (p < 0.05) для всех показателей, кроме пути, пройденного рыбами в установке (табл. 3). Также достоверно и совместное влияние этих факторов (p ≤ 0.04), т.е. у самцов и самок по-разному изменяется реореакция при различных температуре воды и стадии зрелости гонад.
Таблица 3.
Уровни значимости влияния факторов на показатели реореакции плотвы по критерию (H) Краскела–Уоллиса
| Факторы | S, м | Типы реореакции | ЧСТР, мин–1 | |||
|---|---|---|---|---|---|---|
| динамические | статические | |||||
| ПТР | ОТР | КТР | ФТР | |||
| Пол | 0.13 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| t°C | 0.14 | 0.05 | 0.03 | 0.005 | 0.59 | 0.03 |
| ЗГ | 0.94 | 0.18 | 0.62 | 0.007 | 0.77 | 0.10 |
| Пол + t°C | 0.08 | 0.0001 | 0.0001 | <0.001 | 0.10 | <0.001 |
| ЗГ + t°C | 0.32 | 0.01 | 0.03 | 0.001 | 0.68 | 0.01 |
| Пол + ЗГ | 0.74 | <0.001 | 0.0002 | <0.001 | 0.012 | <0.001 |
| Пол + ЗГ + t°C | 0.18 | <0.001 | 0.001 | <0.001 | 0.14 | <0.001 |
Регрессионный анализ подтвердил достоверное влияние рассматриваемых факторов по отдельности на все показатели реореакции. Однако коэффициенты детерминации (R2) оказались в большинстве случаев <0.5 (0.14–0.38), что указывает на слабую зависимость реореакции плотвы от этих факторов. Относительно значимые (R2 > 0.5) уравнения регрессии, в которых достоверны и сами уравнения и все их коэффициенты (p ≤ 0.008), получены только для частоты проявления компенсаторного (РКТР) и статических (РСТР) типов реореакции в целом, а также для частоты смены ее типов (ЧСТР):
PКТР = 0.122$K_{{\text{s}}}^{2}$ + 0.013Kzg, R2 = 0.66;
PСТР = 0.222 t°C – 0.017$t_{{^\circ {\text{C}}}}^{2}$ + 0.030Kzg – 0.001$K_{{{\text{zg}}}}^{2}$, R2 = 0.89;
ЧСТР = 0.67Ks, R2 = 0.67;
где: Ks – код пола (Ks для самок принят равным 1, а для самцов – 2), Kzg – коэффициент зрелости гонад; t°C – температура воды, оC. Все надстрочные цифры – показатели степени.
Исходя из этих уравнений, частота проявления КТР и частота смены типов реореакции у самцов превышает таковые у самок, что соответствует реальному отношению частот смены типов реореакции в апроксимируемый период (апрель–май): 0.22 и 0.47 мин–1 у самок и самцов соответственно.
Как и в целом за период исследований, у плотвы в период нереста преобладала реореакция статических типов (рис. 3). Причем только весной у самцов частота КТР была достоверно больше, чем ФТР (p = = 0.005, критерий Стьюдента для долей).
Рис. 3.
Динамика частоты проявления ПТР (1), ОТР (2), КТР (3) и ФТР (4) у самок (а) и самцов (б) плотвы на IV, V и VI–II стадиях зрелости гонад весной 2017 г. Вертикальная линия – ошибка средней.
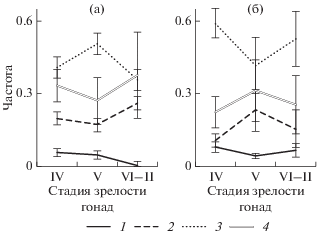
У самок статические типы реореакции (компенсаторный и факультативный) наиболее различаются на V стадии зрелости гонад, динамические (положительный и отрицательный) – на VI–II стадиях, у самцов – соответственно на IV и V стадиях.
У самок наиболее высокая доля динамических типов в период перед нерестом и сразу после нереста. Снижение активности наблюдается во время нереста (V стадия зрелости гонад). В эти периоды сниженной активности, соответственно, высока доля времени, в которое рыба не перемещается, проявляя ФТР или КТР. У самцов снижения активности (доли динамических типов реореакции) на V стадии зрелости гонад не происходит, наоборот, доля статических типов снижается.
Влияние весеннего повышения температуры в диапазоне 3–11°С на соотношение типов реореакции выражено слабо. Судя по коэффициентам уравнений регрессии, с повышением температуры частота динамических типов реореакции несколько увеличивается, статических – снижается. Увеличение частоты реже проявляемых типов реореакции (динамических) приводит и к увеличению частоты смены типов реореакции при увеличении температуры воды.
Для многих видов рыб известно, что весеннее повышение температуры воды вызывает их нерестовую контранатантную миграцию, а в экспериментах усиливает положительный реотаксис (ПТР) (Banks, 1969; Dodson, Young, 1977; Keenleyside, Hoar, 1954; Maclean, Gee, 1971). Поскольку исследованная плотва не находилась в состоянии миграции, то у нее не происходило значительного изменения соотношения типов реореакции при повышении температуры.
Ранее при определении соотношения трех типов реореакции в рыбоходе у немигрирующих рыб разных видов, в том числе и у плотвы, различие в соотношении типов реореакции было недостоверным (Павлов и др., 2019; Pavlov et al., 2010). В отличие от этого, в данной работе при определении соотношения четырех типов реореакции в кольцевом лотке с лимнозоной такие различия у немигрирующих рыб (при явном преобладании статических типов реореакции) были достоверны, т.е. предоставление рыбам возможности проявлять четыре типа реореакции, а не три, позволило и у немигрирующей плотвы выявить различия, не обнаруженные при использовании рыбохода. Это подтверждает целесообразность применения кольцевого лотка с лимнозоной для определения соотношения типов реореакции рыб.
Выводы. Сезонная динамика соотношения типов реореакции у немигрирующей плотвы из водохранилища, определенная в кольцевом лотке с лимнозоной, соответствует ее поведению в естественных условиях. Преобладают статические типы реореакции, частота их проявления колеблется от 0.62 до 0.97. Выявлено достоверное, но слабое влияние на соотношение типов реореакции пола плотвы, стадии зрелости гонад и температуры воды. По-видимому, слабое влияние этих факторов обусловлено немиграционным состоянием исследованных рыб. Подтверждена целесообразность применения кольцевого лотка с лимнозоной для определения соотношения типов реореакции рыб.
Список литературы
Буторин Н.В. 1963. Уровень Рыбинского водохранилища и его колебания (1948–1960 гг.) // Тр. Ин-та биол. водохранилищ АН СССР. Т. 5(8). Москва: Изд-во АН СССР. С. 303.
Буторин Н.В. 1969. Гидрологические процессы и динамика водных масс в водохранилищах Волжского каскада. Ленинград: Наука.
Лакин Г.Ф. 1973. Биометрия. Москва: Высшая школа.
Павлов Д.С. 1979. Биологические основы управления поведением рыб в потоке воды. Москва: Наука.
Павлов Д.С., Лупандин А.И., Костин В.В. 2007. Механизмы покатной миграции молоди речных рыб. Москва: Наука.
Павлов Д.С., Костин В.В., Звездин А.О. и др. 2019. Реореакция молоди некоторых карповых рыб (Cyprinidae) в период осенней контранатантной миграции // Вопр. ихтиологии. Т. 59. № 6. С. 716. https://doi.org/10.1134/S0042875219060122
Павлов Д.С., Паршина В.Ю., Костин В.В., Прозоров Д.А. 2020. Сравнение экспериментальных методов оценки мотивационной компоненты реореакции рыб (соотношения типов реореакции) // Вопр. ихтиол. Т. 60. № 4. С. 418.
Поддубный А.Г. 1966. Об адаптивном ответе популяций плотвы на изменение условий обитания // Тр. Института биол. внутр. вод АН СССР. Вып. 10(13). С. 131.
Поддубный А.Г. 1971. Экологическая топография популяций рыб в водохранилищах. Ленинград: Наука.
Рыбопромысловый атлас Рыбинского водохранилища. 1963. Ин-т биологии внутр. вод АН СССР. Рыбин. рыбопром. трест. Ярославль: Кн. изд-во.
Рыбы Рыбинского водохранилища: популяционная динамика и экология. 2015. Ярославль: Филигрань.
Сакун О.Ф., Буцкая Н.А. 1963. Определение стадий зрелости и изучение половых циклов рыб. Мурманск: ПИНРО.
Banks J.W. 1969. A review of the literature on the upstream migration of adult salmonids // J. Fish. Biol. V. 1. P. 85.
Dodson J.J., Young J.C. 1977. Temperature and photoperiod regulation of rheotropic behavior in prespawning common shniners, Nolropicso rnutus // J. Fish. Res. Board Can. V. 34. P. 341.
Johnston M.E., Kelly J.T., Lindvall M.E. et al. 2017. Experimental evaluation of the use of vision and barbels as references for rheotaxis in green sturgeon // J. Exp. Mar. Biol. Ecol. V. 496. P. 9. https://doi.org/10.1016/j.jembe.2017.04.002
Keenleyside M.H.A., Hoar W.S. 1954. Effects of temperature on the responses of young salmon to water currents // Behaviour. V. 7. P. 77.
Maclean J.A., Gee J.H. 1971. Effect of temperature on movements of pre-spawning brook sticklebacks, Culaea inconstans, in the Roseau River, Manitoba // J. Fish. Res. Board Can. V. 28. P. 919.
Pavlov D.S., Kostin V.V., Zvezdin A.O., Ponomareva V.Yu. 2010. On methods of determination of the rheoreaction type in fish // J. Ichthyol. V. 50. № 11. P. 977. https://doi.org/10.1134/s0032945210110020
Zvezdin A.O., Pavlov D.S., Kostin V.V. 2015. On the mechanism of orientation and navigation of sockeye salmon underyearlings (Oncorhynchus nerka Walb.) during feeding migration in the inlet–lake–outlet system // Inland Wat. Biol. V. 8. № 3. P. 287. https://doi.org/10.1134/s195082915030189
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод