

Биология внутренних вод, 2020, № 5, стр. 431-438
Микроморфология реликтовых гидрофитов Нижнего Приамурья
Д. Ю. Цыренова *
Тихоокеанский государственный университет
Хабаровск, Россия
* E-mail: Duma@mail.ru
Поступила в редакцию 03.12.2019
После доработки 10.02.2020
Принята к публикации 26.03.2020
Аннотация
Исследована микромофология реликтовых гидрофитов Brasenia schreberi J.F. Gmel. (Cabombaceae), Nuphar japonica DC. (Nymphaeaceae) и Nelumbo komarovii Grossh. (Nelumbonaceae) с Нижнего Приамурья. Приведены подробные описания анатомического строения листьев, контактирующих с воздушной средой. Общая черта – дорзовентральность листовой пластинки и атактостелия черешков. Развитие системы диффузных межклетников и аэренхимы, редукция ксилемы у изученных видов – эффективные адаптации к водной среде обитания. Предположено, что изученные виды сохранились на юге Дальнего Востока России благодаря максимальной защищенности в зимний период. Компенсационные адаптации в микроструктуре листа – развитие многофункциональной паренхимы, повышенное содержание вторичных метаболитов и гидрофобных соединений, армированные листья.
ВВЕДЕНИЕ
Современная водная флора Нижнего Приамурья богата реликтовыми представителями, чьи предковые формы существовали на данной территории с третичного периода. Они достигают ~30% флоры водоемов (Крюкова, 2005, 2013). Объектами настоящего исследования, как примеры очевидного реликтового эндемизма, послужили виды бразения Шребера Brasenia schreberi J.F. Gmel. (Cabombaceae), кубышка японская Nuphar japonica DC. (Nymphaeaceae) и лотос Комарова Nelumbo komarovii Grossh. (Nelumbonaceae) (Харкевич, Качура, 1981; Шлотгауэр, Крюкова, 2005). Все виды находятся под угрозой исчезновения на северном и северо-восточном пределах распространения и внесены в список охраняемых растений (Красная книга Российской…, 2008; Красная книга Хабаровского…, 2008).
Исследованные виды относятся к гидрофитам, или настоящим водным растениям из двух экологических групп: плейстогидрофиты с плавающими на поверхности воды листьями (Brasenia schreberi) и аэрогидрофиты с плавающими и надводными листьями (Nuphar japonica и Nelumbo komarovii) (Папченков, 1985; Распопов, 1985).
Brasenia schreberi – почти космополитный вид, распространенный в Южной и Восточной Азии, Северной Америке, Африке и Австралии (Ohwi, 1965; Цвелев, 1987; Lee, 2007; www.efloras.org). В России встречается в Восточной Сибири (единственное местонахождении в Иркутской обл.) и на юге Дальнего Востока (Цвелев, 1987; Чепинога, 1999; Малышев, 2012). Хромосомное число 2n = 80 (Соколовская, Пробатова, 1985). Растение культивируют (Пшенникова, 2005). На территории Нижнего Приамурья популяции бразении Шребера произрастают в старицах с низкими заболоченными берегами, часто переходящими в сплавины, на глубине 1.5–2.5 м. Образуют монодоминантные куртины различных размеров с примесью Myriophyllum verticilliatum L., Potamogeton maackianus A. Benn., Trapa maximowiczii Korsh., Utricularia vulgaris L. (Нечаев, Павленко, 1967; Мельникова, Мурдахаев, 1979).
Nuphar japonica – амуро-корейско-японский вид (Ohwi, 1965; Цвелев, 1987; Lee, 2007). На Дальнем Востоке России встречается лишь в нижнем течении р. Кия – притока р. Уссури (Нечаев, Павленко, 1969; Цвелев, 1987; Крюкова, 2013). Узколокальные дальневосточные популяции этого вида представляют собой самую северную материковую часть ареала преимущественно островного японского вида. Хромосомное число 2n = 34 (Пробатова и др., 2006). В Японии известны культурные формы вида (Харкевич, Качура, 1981). Дальневосточные популяции кубышки японской обитают в старицах и заводях реки с медленнотекущей водой. Сопутствующие виды: Ceratophyllum demersum L., Hydrilla verticilliata Rich., Potamogeton maackianus A. Benn., P. natans L., P. malainus Miq. (Нечаев, Павленко, 1969).
Nelumbo komarovii – амуро-уссурийско-маньчжурский вид (Цвелев, 1987). На юге Дальнего Востока России встречается по долине р. Амур в пределах Амурской обл. и Хабаровского края, охватывая низовья рек Зея и Бурея. По долине р. Уссури его ареал переходит на Приханкайскую низменность и о. Путятино Приморского края. Хромосомное число 2n = 16 (Probatova et al., 2011). Китайские исследователи синонимизируют N. komarovii с N. nucifera Gaertner (www.efloras.org). По мнению Н.Н. Цвелева (1987), дальневосточный лотос – наиболее северный по распространению восточноазиатский дикорастущий представитель древнего третичного рода Nelumbo, тогда как южноазиатский лотос N. nucifera и каспийский лотос N. caspica (DC.) Fisch. – культивируемые и уже подвергнувшиеся длительной селекции формы. На территории Нижнего Приамурья обнаружено около двух десятков лотосовых озер в пойме р. Амур как естественного, так и искусственного происхождения (Крюкова, 2009). Популяции образуют в пойменных озерах и старицах монодоминатные ценозы с примесью Trapa japonica Fler., Trapella sinensis Oliv. и Ceratophyllum demersum L. (Крюкова, 2009; Дулин, Цыренова, 2017).
Известно, что для оценки адаптивного потенциала видов, наряду с морфо-физиологическими и биологическими особенностями, используются и анатомические признаки (Барыкина, Чубатова, 2005; Таршис, 2007; Efremov, 2016; Tsyrenova et al., 2018). Анализ литературных сведений показал, что микроморфологические особенности выбранных видов и их приспособления к естественным условиям существования на северном пределе распространения ранее не рассматривали.
Цель работы – изучить микроморфологию ассимилирующих листьев реликтовых гидрофитов Nuphar japonica, Brasenia schreberi и Nelumbo komarovii из Нижнего Приамурья.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Исследования проведены на материале, собранном нами в естественных популяциях видов на территории Нижнего Приамурья (табл. 1).
Таблица 1.
Места сбора материала
| Вид | Водный объект | Координаты: с.ш., в.д. | Дата сбора |
|---|---|---|---|
| Еврейская автономная область | |||
| Brasenia schreberi | Озеро Ледяное | 48°37′32.80″, 134°19′02.94″ | 10 VIII 2018 г. |
| Хабаровский край | |||
| Nuphar japonica | Река Кия | 47°57′22.35″, 135°03′39.58″ | 6 VIII 2014 г. |
| Nelumbo komarovii | Лотосовое озеро | 48°32′11.23″, 135°23′20″ | 21 VII 2016 г. |
Общий объем сборов, включая гербарные образцы для коллекций и фиксированный в спирте материал для микроморфологического анализа, составил ~20 экз. растений. Материалы хранятся в Тихоокеанском государственном университете (г. Хабаровск). Определение собранных видов проводили по работе Н.Н. Цвелева (1987), анатомические исследования – по методике Г.Г. Фурст (1979). Поперечные срезы органов делали лезвием бритвы. Для окрашивания использовали спиртовой раствор флороглюцина с каплей концентрированной соляной кислоты. Готовые препараты просматривали с помощью микроскопа Биолам-ЛОМО и цифрового микрокомплекса Альтами БИО 8. При определении размеров клеток и пропорции гистологических зон применяли окуляр-микрометр М-ЛОМО и объект-микрометр ОМО. Визуализацию микропрепаратов проводили с помощью цифровой камеры для микроскопа ToupCam с использованием программного обеспечения ToupView.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Листовая пластинка. Анатомические особенности листа у всех изученных видов коррелируют со степенью экспозиции их в воздушной среде. Листовая пластинка плавающих листьев у плейстогидрофита Brasenia schreberi эпистоматического типа (рис. 1а, 1б). В группе аэрогидрофитов у Nuphar japonica листья гипостоматического типа (рис. 2а, 2б). Плавающие и воздушные листья у Nelumbo komarovii одинаково эпистоматические (рис. 3а, 3в).
Рис. 1.
Строение листа Brasenia schreberi: а – адаксиальная эпидерма, б – абаксиальная эпидерма, в – поперечный срез листовой пластинки (фрагмент), г – поперечный срез черешка (фрагмент), ed – эндодерма, ic – внутренняя кутикула, it – межклетник, ph – элементы флоэмы, pp – палисадная паренхима, ps – воздухоносная полость, sp – губчатая паренхима, t – трабекула, tr – слизевыделительная железка, x – элементы ксилемы.

Рис. 2.
Строение листа Nuphar japonica: а – адаксиальная эпидерма, б – абаксиальная эпидерма, в – поперечный срез листовой пластинки (фрагмент), г – поперечный срез черешка (фрагмент), д – проводящий пучок черешка, cb – проводящий пучок, sc – астросклереида, tr – головчатая трихома. Остальные обозначения, как на рис. 1.
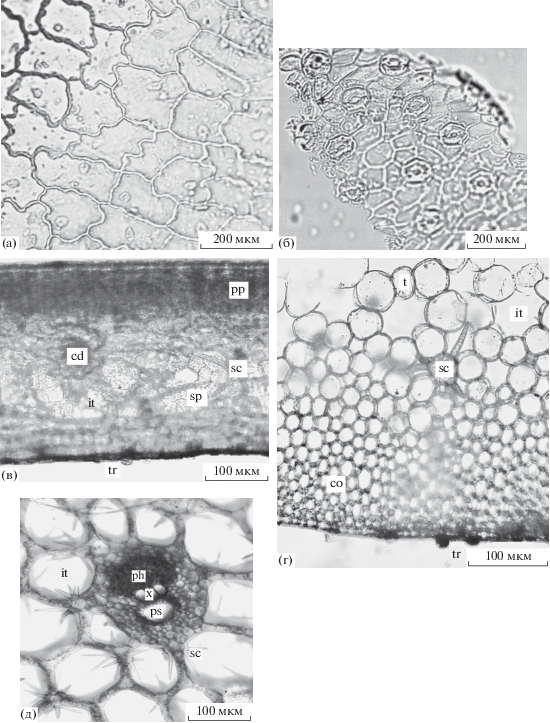
Рис. 3.
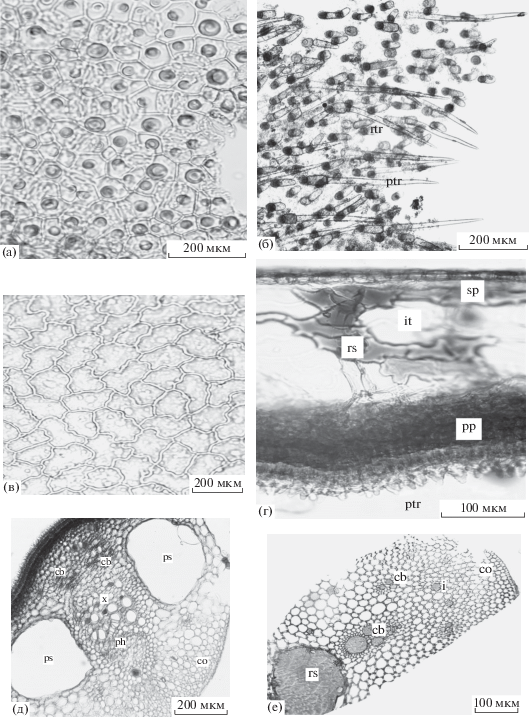
Строение листьев Nelumbo komarovii: а – адаксиальная эпидерма плавающего листа, б – опушение верхней эпидермы плавающего листа, в – абаксиальная эпидерма надводного листа, г – поперечный срез листовой пластинки (фрагмент), д – поперечный срез крупной жилки, е – поперечный срез черешка (фрагмент), i – идиобласт, ptr – остроконечная трихома, rtr – тупоконечная трихома, rs – секреторное вместилище. Остальные обозначения, как на рис. 1 и 2.
Для всех изученных видов устьичные аппараты однотипные – аномоцитные. Устьица крупные (длиной ~160 мкм, шириной ~130 мкм), замыкающие клетки почковидно-бобовидные, устьичные щели широко открытые. Редкие устьица наблюдаются у Brasenia schreberi (~44 экз./мм2), их значительно больше у Nuphar japonica (~160 экз./мм2) и Nelumbo komarovii (~116 экз./мм2).
Для видов характерны различные выросты эпидермального происхождения, обеспечивающие гидрофобность листьев в воде. Абаксиальная эпидерма Brasenia schreberi образует слизевыделительные железки (рис. 1в, tr). Каждая железка имеет ножку из двух мелких клеток и более крупную и округлую секретирующую окрашенную головку. Благодаря этому, обращенная в воду нижняя сторона листа фиолетового цвета покрыта толстым слоем гидрофобной слизи. В толще слизи присутствуют микроскопические одноклеточные и нитчатые водоросли. У Nuphar japonica (особенно вдоль жилок на нижней стороне листовой пластинки) найдены редкие короткие головчатые трихомы (рис. 2в, tr). У Nelumbo komarovii адаксиальная эпидерма сплошь покрыта одноклеточными трихомами двух типов: короткими сосочковидными тупоконечными (рис. 3б, rtr) и длинными остроконечными (рис. 3б, ptr).
Мезофилл плавающих и надводных листьев имеет дорзовентральное строение. Палисадная паренхима фотосинтезирующая, относительно компактная и многослойная, с продольно вытянутыми узкими клетками. У эпистоматических листьев они прерываются подъустьичными воздушными полостями (Brasenia schreberi и Nelumbo komarovii). Губчатая паренхима аэренхиматизирована. В ней у Brasenia schreberi (рис. 1в, it) и Nuphar japonica (рис. 2в, it) между рыхло расположенными клетками находятся диффузные межклетники диаметром 120–500 мкм. У Brasenia schreberi межклетники изнутри выстланы внутренней кутикулой. Более дифференцирована губчатая паренхима у Nelumbo komarovii (рис. 3г, sp). К палисадной паренхиме изнутри прилегает компактная зона с проводящими пучками. За ней следует ряд крупных лизигенных воздухоносных полостей с ярко-коричневым содержимым диаметром 2000–2400 мкм, лакуны непосредственно контактируют с адаксиальной эпидермой.
В области крупных жилок листа строение губчатой паренхимы значительно усложняется. Это особенно заметно у надводных листьев Nuphar japonica и Nelumbo komarovii, которым необходима дополнительная жесткость для их нахождения в воздушной среде. У Nuphar japonica формируется типичная аэренхима в виде ажурной сетки с шестиугольными воздушными полостями и армируется астросклереидами (рис. 2д, sc). Стенки полостей (трабекулы) образованы однорядными клетками, соединенными друг с другом одной более крупной опорной клеткой. К опорной клетке тесно прижата многоветвистая астросклереида, располагающая свои ответвления в трех смежных воздухоносных полостях аэренхимы. Астросклереиды живые, с зернистым содержимым внутри, утолщения стенок пронизаны поровыми каналами. Наружная поверхность астросклереид неровная. Проводящие пучки коллатеральные, с воздухоносной полостью в ксилеме и с выраженной паренхимной обкладкой. Они дополнительно армируются колленхимой уголкового типа. У Brasenia schreberi крупные жилки заметно выступают на нижней стороне листовой пластинки. В них находится один проводящий пучок коллатерального типа и один-два слоя колленхимы под абаксиальной эпидермой. Армированные листья Nuphar japonica обладают достаточной жесткостью и способны не только возвышаться высоко над водной поверхностью, но и противостоять течению речной воды. Из исследованных нами видов только кубышка японская встречается в медленнотекущих водах малой реки. Крупные жилки Nelumbo komarovii представлены колленхиматизированной паренхимой, двумя лизигенными вместилищами округлой формы, одним крупным проводящим пучком нормального коллатерального типа (рис. 3д, ph, x) и несколькими мелкими редуцированными пучками (рис. 3д, cb).
Черешок. Эпидерма черешка у изученных видов однослойная. Выполняет секретирующую функцию: слизевыделительные железы у Brasenia schreberi (рис. 1в, tr) и головчатые трихомы у Nuphar japonica (рис. 2г, tr).
Зона первичной коры дифференцирована на внешний и внутренний участки по-разному. У Brasenia schreberi внешняя кора представлена 3–4 слоями компактной колленхиматизированной паренхимы. Внутренняя кора – это аэренхима с крупными воздухоносными полостями, располагающимися в один или несколько кругов и отделяющимися друг от друга однослойными перегородками. Включает многочисленные крахмальные зерна. Стенки клеток, обращенные к воздухоносной полости, кутинизированы (рис. 1в, 1г, ic).
У Nuphar japonica внешняя кора представлена 6–7 (8)) слоями колленхиматизированной паренхимы. Внутренняя кора в виде аэренхимы с ажурной сетью крупных воздухоносных полостей. Воздухоносные полости армированы немногочисленными астросклереидами (рис. 2г, sc).
У Nelumbo komarovii в черешках у плавающих и надводных листьев вся кора одинаково компактная, без межклетников. Внешняя кора состоит из 1–2 параллельных слоев радиально вытянутых паренхимных клеток, отличающихся от нижележащих клеток своей упорядоченностью и светло-коричневым содержимым. Внутренняя кора несет 4–5 слоев уголковой колленхимы (рис. 3е, co) и одиночные секретирующие идиобласты (рис. 3е, i).
У изученных видов в черешках присутствует атактостела. Она образована проводящими пучками коллатерального типа. У Brasenia schreberi наблюдаются два проводящих пучка, расположенных по обе стороны от узкопросветного воздухоносного канала в центре органа. Стела отграничена от первичной коры эндодермой в виде однослойного крахмалоносного влагалища (рис. 1г, ed). У двух других видов проводящие пучки многочисленные и разбросанные среди аэренхиматизированной паренхимы. У Brasenia schreberi (рис. 1г, ps) и Nuphar japonica (рис. 2д, ps) сосудистые элементы ксилемы частично или полностью замещены одной–двумя воздухоносными полостями. У Nelumbo komarovii многие из сосудистых элементов пучков становятся секреторными вместилищами (3е, rs).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Листовая пластинка. У изученных видов в листьях проявляются микроморфологические адаптации, характерные для гидроморфной эволюции растений. К типичным гидроморфным признакам относятся: развитие полифункциональной аэренхимы, выполняющей функции фотосинтеза, транспорта газов, вместилища секретов и воздуха; слабое развитие ксилемы, частично замещенной воздухоносными полостями; диффузное расположение механических элементов.
Несмачиваемость листьев в воде обеспечивается гидрофобными соединениями, вырабатываемыми секреторными образованиями эпидермального происхождения. При исследовании Brasenia schreberi обнаружено присутствие внутренней кутикулы, выстилающей межклетники в губчатой паренхиме. Это относится к редким явлениям, поскольку у большинства растений образуются только поверхностные кутикулярные слои (Эсау, 1980).
У всех трех видов листья контактируют в той или иной степени с воздушной средой. Общая черта в их строении – дорзовентральность мезофилла. Надводные листья у аэрогидрофитов Nuphar japonica и Nelumbo komarovii имеют больше признаков наземных растений по локализации устьиц, их количеству, очертанию антиклинальных стенок основных клеток и секреции эпидермы. При понижении уровня воды эти виды могут входить в сообщество околоводных растений, что часто наблюдается у дальневосточных популяций (Дулин, Цыренова, 2017; Крюкова, 2009).
Черешок изученных видов характеризуется микроструктурой осевого органа с типичными гистолого-топографическими комплексами. Пропорции между этими комплексами видоспецифичны. В черешках Brasenia schreberi крахмалоносная и аэренхиматизированная первичная кора превалирует над сильно смещенной к центру стелой, т.е. черешки у этого вида представляют собой запасающие органы. В черешках Nuphar japonica и Nelumbo komarovii стела, наоборот, топографически преобладает над первичной корой. Запасающие ткани не развиты. Черешки у этих видов функционируют как типичные несущие органы. Дополнительную жесткость и упругость черешкам придают колленхиматизация и аэренхиматизация тканей.
Всем изученным видам присуща атактостелия. Этот признак – эволюционная особенность представителей двудольных из древних сем. Cabombaceae, Nymphaeaceae и Nelumbonaceae близкородственных с однодольными (Тахтаджян, 1987).
Не вызывает сомнения, что изученные виды сохранились на юге Дальнего Востока России благодаря своим биологическим особенностям на разных уровнях, в том числе, и на микроморфологическом. Особенно эффективны идиоадаптации видов для перенесения неблагоприятного существования в воде в зимний период. По-видимому, повышенное содержание вторичных метаболитов в тканях изученных видов обеспечивают максимальную защищенность живых тканей растений при контакте со льдом. Студенистая слизь и внутренняя кутикула воздухоносных каналов у Brasenia schreberi также способствуют холодостойкости вида.
Выводы. Проведенные микроморфологические исследования ассимилирующих органов у реликтовых видов водной флоры Нижнего Приамурья позволяют сделать некоторые предположения об эволюционной истории видов. Выработка внутренней кутикулы у Brasenia schreberi, гипостоматическая эпидерма и экскреторные структуры у Nuphar japonica, слабая аэрация коровой паренхимы, превалирование стелы в черешке у Nelumbo komarovii, свидетельствуют о существовании в прошлом у этих видов наземных предковых форм. По особенностям аэренхиматизации тканей листовой пластинки и черешка Brasenia schreberi и Nuphar japonica, вероятно, прошли более длительную эволюцию в водной среде, чем Nelumbo komarovii. Два первых вида уже выработали специализированную аэренхиму с упорядоченной системой межклетников и армированием их опорных клеток с помощью диффузных склереидов или уголковой колленхимой. Судя по значительной редукции проводящей системы листьев и черешка, степень гидроморфности Brasenia schreberi выше, чем у Nuphar japonica. Напротив, Nelumbo komarovii, по-видимому, освоил водную среду обитания в более позднее время, типичная аэренхима не развита, аэрация вегетативных органов происходит через дефинитивные лизигенные секреторные вместилища. Выявленные особенности микроморфологической структуры изученных видов, представителей реликтовой водной флоры региона, весьма устойчивы и консервативны, что при современных климатических и антропогенных изменениях окружающей среды может привести к вымиранию этих видов.
Список литературы
Барыкина Р.П., Чубатова Н.В. 2005. Экологическая анатомия цветковых растений: учебно-методическое пособие. Москва: Товарищество науч. изданий КМК.
Дулин А.Ф., Цыренова Д.Ю. 2017. Исследование фенологии и семенной продуктивности лотоса Комарова Nelumbo komarovii Grossh. в популяции вида вблизи Хабаровска (Дальний Восток, Россия) // Ученые записки Забайкальского госуниверситета. Серия Биологические науки. Т. 12. № 1. С. 13.
Красная книга Российской Федерации (растения и грибы). 2008. Москва: Товарищество науч. изданий КМК.
Красная книга Хабаровского края: Редкие и находящиеся под угрозой исчезновения виды растений и животных. 2008. Хабаровск: Изд. дом “Приамурские ведомости”.
Крюкова М.В. 2005. Флора водоемов Нижнего Амура. Владивосток: Дальнаука.
Крюкова М.В. 2009. Эколого-биологические особенности лотоса Комарова на северо-восточной границе ареала // Вест. Дальневост. отд. РАН. № 3. С. 27.
Крюкова М.В. 2013. Сосудистые растения Нижнего Приамурья. Владивосток: Дальнаука.
Малышев Л.И. 2012. Сем. Cabombaceae. Сем. Nymphaeaceae. Сем. Nelumbonaceae // Конспект флоры Азиатской России: Сосудистые растения. Новосибирск: Изд-во СО РАН. С. 28.
Мельникова А.Б., Мурдахаев Ю.М. 1979. Бразения Шребера на севере своего ареала и при интродукции в бассейны Ботанического сада УзССР // Интродукция и акклиматизация растений. Ташкент: ФАН. Вып. 16. С. 41.
Нечаев А.П., Павленко Г.Е. 1967. Brasenia schreberi J.F. Gmel. в Хабаровском крае // Бот. журн. Т. 52. № 12. С. 1795.
Нечаев А.П., Павленко Г.Е. 1969. Nuphar japonica DC. на юге Хабаровского края // Бот. журн. Т. 54. № 6. С. 931.
Папченков В.Г. 1985. О классификации макрофитов водоемов и водной растительности // Экология. № 6. С. 8.
Пробатова Н.С., Рудыка Э.Г., Баркалов В.Ю. и др. 2006. Числа хромосом сосудистых растений из заповедников Приморского края и Приамурья // Бот. журн. Т. 91. № 7. С. 1117.
Пшенникова Л.М. 2005. Водные растения российского Дальнего Востока. Владивосток: Дальнаука.
Распопов И.М. 1985. Высшая водная растительность больших озер Северо-Запада СССР. Ленинград: Наука.
Соколовская А.П., Пробатова Н.С. 1985. Числа хромосом сосудистых растений из Приморского края, Камчатской области, Приамурья и Сахалина // Ботан. журн. Т. 70. № 7. С. 997.
Таршис Л.Г. 2007. Анатомия подземных органов высших сосудистых растений. Екатеринбург: Изд-во УрО РАН.
Тахтаджян А.Л. 1987. Система магнолиофитов. Ленинград: Наука.
Фурст Г.Г. 1979. Методы анатомо-гистохимического исследования растительных тканей. Москва: Наука.
Харкевич С.С., Качура Н.Н. 1981. Редкие виды растений советского Дальнего Востока и их охрана. Москва: Наука.
Цвелев Н.Н. 1987. Сем. Кабомбовые – Cabombaceae. Сем. Кувшинковые – Nymphaeaceae. Сем. Лотосовые – Nelumbonaceae // Сосудистые растения советского Дальнего Востока. Ленинград: Наука. Т. 2. С. 21.
Чепинога В.В. 1999. Brasenia schreberi (Cabombaceae) – новый вид для флоры Сибири // Бот. журн. Т. 84. № 6. С. 144.
Шлотгауэр С.Д., Крюкова М.В. 2005. Редкие и исчезающие виды сосудистых растений Хабаровского края // Комаровские чтения. Владивосток: Дальнаука. Вып. 51. С. 57.
Эсау К. 1980. Анатомия семенных растений. Москва: Мир. Кн. 1, 2. (Esau Katherine. Anatomy of Seed Plants. N.Y.: John Wiley and Sonsю. 1977).
Efremov A.N. 2016. Anatomy and morphology of vegetative organs and inflorescence of Stratiotes aloides L. (Hydrocharitaceae) // Inland Water Biology. V. 9. № 1. P. 27. https://doi.org/10.1134/S1995082916010041
Lee Yong No. 2007. Flora of Korea. Seoul: Kyo-Hak Publishing Co.
Ohwi J. 1965. Flora of Japon. Washington: Smithsonian Institution.
Probatova N.S., Kazanovsky S.G., Rudyka E.G. et al. 2011. IAPT/IOPB chromosome data 12 // Taxon. V. 60. № 6. P. 1790.
Tsyrenova D.Ju., Varfolomeeva A.S., Goroshko Ju.M. 2018. Micromorphology of Hydrocharis dubia (Blume) Backer (Hydrocharitaceae) plants from the Far East of Russia // Inland Water Biology. V. 11. № 3. P. 264. https://doi.org/10.1134/S1995082918030227
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод