

Биология внутренних вод, 2020, № 6, стр. 573-582
Состав, структура и распределение фитопланктона высокоминерализованного карстового озера
Е. Л. Воденеева a, b, *, А. Г. Охапкин a, С. И. Генкал c, П. В. Кулизин a, Е. М. Шарагина a, К. О. Скамейкина d
a Нижегородский государственный университет им. Н.И. Лобачевского, Институт биологии и биомедицины
Нижний Новгород, Россия
b Нижегородский филиал “Всероссийского научно-исследовательского института рыбного хозяйства
и океанографии”
Нижний Новгород, Россия
c Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
пос. Борок, Некоузский р-н, Ярославская обл., Россия
d ОАО “Гидроагрегат”
Павлово, Россия
* E-mail: vodeneeva@mail.ru
Поступила в редакцию 11.11.2019
После доработки 14.02.2020
Принята к публикации 27.03.2020
Аннотация
Рассмотрены особенности состава, развития и пространственного распределения фитопланктона высокоминерализованного карстового оз. Ключик (Нижегородская обл.), имеющего выраженный экотон по гидролого-гидрохимическим показателям. В западной части озера и в зоне экотона выявлен уникальный альгоценоз с несвойственным для водоемов средней полосы высоким количественным развитием (биомасса >130 г/м3) и преобладанием центрической диатомеи Cyclotella distinguenda Hustedt., редкого вида, нехарактерного для водных объектов Средней Волги. Вертикальное распределение фитопланктона показало наличие придонного максимума в зоне экотона и в районе максимальных глубин.
ВВЕДЕНИЕ
В условиях интенсивного антропогенного воздействия и глобального изменения климата изучение биологического разнообразия и структурно-функциональной организации водной биоты по-прежнему относится к актуальным направлениям современных гидробиологических исследований. Особую значимость эти исследования приобретают для природных охраняемых территорий, которые являются прототипами ненарушенных биогеоценозов и выполняют глобальную функцию сохранения биогеохимических взаимосвязей между биотической и абиотической компонентами экосистемы. Водоемы, которые находятся в первозданном состоянии, представляют собой идеальные объекты для оценки изменений в сообществах гидробионтов. К таким водоемам относятся карстовые озера, образовавшиеся в местах карстовых провалов и характеризующиеся разнообразием морфометрических, гидрологических и гидрохимических параметров, а также специфическим гидробиологическим режимом (Корнева, 2000; Уникальные…, 2001; Палагушкина, 2004; Гусев, 2008а, 2008б; Баянов и др., 2009, 2014; Горохова, Паутова, 2009; Ternjej et al., 2010; Варгот и др., 2015).
Солоновато-водные сульфатные озера, азональные для европейской части России, редко встречаются среди карстовых водоемов. В Среднем Поволжье, где распространены карстовые явления, они составляют ≤0.1% общего числа озер (Уникальные…, 2001). Примером служат так называемые “голубые” озера окрестностей г. Казань (Уникальные…, 2001), Самарской обл. (Тарасова, 2010), Красноярского края (Кусковский, Кривошеев, 1989), Кавказа (Ефремов, 1988). Химический состав их вод определяется не поверхностным стоком, а особенностями карстовых провалов, образовавшихся при вымывании подземными водами растворимых горных пород.
К таким водоемам относится оз. Ключик (Доскинское Святое) – природно-исторический памятник Поволжья, расположенный в Павловском р-не Нижегородской обл. (Бакка, Киселева, 2009; Баянов, 2019). Уникальность озера определяется характером питания, которое осуществляется водами подземной р. Суринь, впадающей в озеро в виде ручья и вытекающей сформированным водотоком метровой глубины (Козлов и др., 2017); наличием экотона – переходной зоны, где зарегистрированы изменения гидрологических и гидрохимических параметров; сменой цвета воды от голубого в западной части до зеленоватого в центральной и восточной котловинах (Баянов, 2019).
В перечень приоритетных показателей качества воды и экологического статуса озер входят состав, численность и интенсивность развития фитопланктона (John, 2012; Rousseaux, Gregg, 2014; Salmaso et al., 2014), исследования которого позволяют расширить представления о структурно-функциональной организации сообществ гидробионтов карстовых озер.
Цель работы – определить особенности состава и пространственного распределения фитопланктона высокоминерализованного оз. Ключик под действием факторов среды и их градиентов, в том числе – экотонного эффекта.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Карстовое оз. Ключик (56°58′ с.ш., 43°20′ в.д.) относится к солоновато-водным (минерализация ~2 г/дм3, электропроводность 1515–1640 мкСм/см), малым (площадь 11 га) озерам с глубиной от 2 до 11.7 м. Озеро имеет котловину сложной формы (рис. 1). В поверхностных слоях воды отмечена высокая концентрация растворенного кислорода (10–11 мг/дм3 или 120–140% насыщения), снижающаяся в районе воклины до 4–6 мг/дм3 (~40% насыщения). Вода в западной части озера окрашена в голубоватый цвет, в центральной и восточной частях – в зеленоватый. Цветность воды постепенно нарастает от 40 град. в западной части озера до 62 и 80 град. в центральной и восточной частях. Западная часть озера, которая не замерзает даже в наиболее холодные зимы, характеризуется равномерным вертикальным распределением температуры, содержания кислорода и величины рН в течение всего года (Баянов, 2019). Летняя температура воды здесь не превышает 8–10°С, рН 7.4–7.5, прозрачность достигает 9.5 м. В центральной и восточной частях прозрачность снижается до 4.5 и 3.6 м соответственно, рН увеличивается до 8.1–8.2, поверхностный слой воды прогревается до 22–25°С, но ниже 1 м температура снижается, достигая 7.4–11.5°С у дна (Баянов, 2019). В переходной зоне (от западной части к центральной) регистрируются максимальные изменения гидрологических и гидрохимических показателей.
Рис. 1.
Карта-схема оз. Ключик. Ст. 1‒Ст. 5 – станции отбора проб. 2 м, 4 м, 6 м, 8 м, 10 м, 13.5 м ‒ глубины.
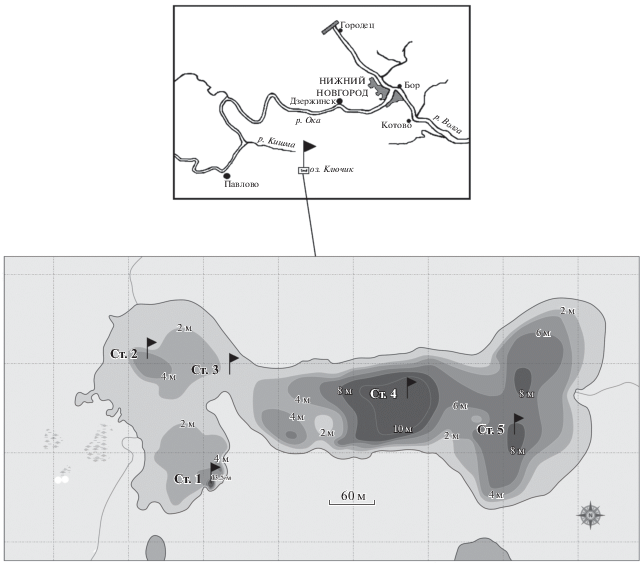
Материалом работы послужили 76 проб фитопланктона, собранных в озере в июне–августе 2017 г. Пробы отбирали батометром Руттнера на пяти станциях, где особенно ярко проявляются гидрохимические особенностей озера (рис. 1). Станции 1 и 2 располагались в западной части водоема; ст. 4 и 5 – в центральной и восточной котловинах; ст. 3 – в переходной экотонной зоне. На всех станциях анализировали интегральные пробы, на ст. 1, 3, 5 в июле и августе дополнительно отбирали пробы через каждый метр от поверхности до дна. Пробы фиксировали йодно-формалиновым раствором, концентрировали до 5 мл сочетанием осадочного метода и прямой фильтрации (Okhapkin et al., 2014) и просматривали под световым микроскопом (MEIJI Techno, увеличение ×1000) с масляной иммерсией. Для учета фитопланктона использовали камеру Нажотта объемом 0.01 мл, биомассу вычисляли счетно-объемным методом. Для характеристики состава диатомовых водорослей готовили постоянные препараты с применением смолы Naphrax (Методика…, 1975). Качественный и количественный учет центрических диатомей проводили с помощью СЭМ JSM-25S (Genkal et al., 2019). Перечень руководств, использованных при идентификации видового состава фитопланктона, дан в работе Воденеевой, Кулизина (2019). К доминирующим отнесены виды с численностью или биомассой ≥10% общей величины, к субдоминантам – с численностью (биомассой) 5–10%. Сходство альгоценозов по видовому составу и биомассе оценивали методом попарных сравнений с помощью индексов Серенсена (качественная мера) и Серенсена–Чекановского (количественная мера) (Шитиков и др., 2003). При кластеризации данных использовали метод Варда, в качестве метрики – нормированное эвклидово расстояние, выраженное в процентах ((Dlink/Dmax) × 100).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
В альгофлоре оз. Ключик из девяти отделов водорослей выявлено 133 таксона (видовые, внутривидовые и определенные до рода). В сложении общего видового богатства ведущая роль принадлежит диатомовым водорослям (Bacillariophyta) – 46% общего списка видов. Другие систематические группы представлены беднее: Chlorophyta – 24%, Chrysophyta –11%, Cyanoprokaryota – 8%, Xanthophyta, Raphidophyta, Euglenophyta, Cryptophyta и Dinophyta – <3% каждая. Удельное видовое богатство фитопланктона варьировало по станциям от 4 до 30 видовых и внутривидовых таксонов в пробе. В западной части озера оно было 13–20 таксонов для всей водной толщи и 4–15 для отдельных глубин, в зоне экотона (ст. 3) увеличивалось до 26–30 и 10–29 таксонов соответственно.
Среди диатомовых водорослей наибольшим видовым богатством характеризовались пеннатные (36.8% общего состава), доминировали роды Pinnularia, Gomphonema и Navicula. Центрические диатомовые были представлены 12 видовыми таксонами (9%), наиболее часто встречались Cyclotella distinguenda Hustedt, C. meneghiniana Kütz., Stephanodiscus hantzschii Grunow, S. minutulus (Kütz.) Cleve et J.D. Möller, S. neoastraea (Håkansson et Hickel) emend Casper, Scheffler et Augsten, S. triporus Genkal et Kuzmin, Handmannia comta (Ehrenb.) Kociolek et Khursevich, Discostella pseudostelligera (Hust.) Houket Klee, Melosira varians C. Agardh (рис. 2).
Рис. 2.
Центрические диатомовые водоросли альгофлоры оз. Ключик: а – Cyclotella distinguenda, б – Disctostella pseudostelligera, в – Handmannia comta, г – Melosira varians, д – Stephanodiscus hantzschii, е – S. minutulus, ж – S. neoastraea, з – S. triporus; а, в–е – створки с наружной поверхности, б, ж, з – с внутренней.

В биотопическом отношении основу флористического списка представляли бентосные виды (33.3% общего видового состава), вторую позицию занимали литоральные и истинно-планктонные виды (по 25.4%), третью – обитатели обрастаний различных субстратов (15.9%). Анализ соотношения географических элементов флоры выявил преобладание космополитов (87.1% числа видов с известной характеристикой) и невысокую долю бореальных видов (12.9%). По отношению к солености воды превалировали индифференты (83.3%), по отношению к pH – алкалифилы (68.2%). Среди индикаторов органического загрязнения преобладали показатели β-мезосапробной зоны (50%), показатели более чистых вод – олигосапробы и o–β-мезосапробы составляли по 14.3%, α-мезосапробных и β–α-мезосапробных вод – 21.4%.
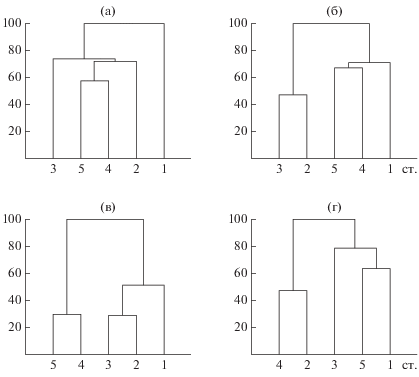
Наибольшее сходство видового состава водорослей выявлено для станций центральной и восточной акваторий озера (индекс Серенсена 0.62–0.72), максимальные различия с другими – для самой глубокой ст. 1 (индекс Серенсена 0.47–0.61). Специфичность количественного развития фитопланктона и состава ценозообразующих видов на этой станции подтверждают низкие величины индекса Серенсена–Чекановского (2–19%), а также результаты кластерного анализа (рис. 3). Максимальное своеобразие альгоценозов на ст. 1 было отмечено в июне и июле.
Рис. 3.
Дендрограммы сходства видового состава (а) и количественных показателей альгоценозов оз. Ключик в июне, июле и августе (б, в, г, соответственно). По оси ординат – расстояние объединения, по оси абсцисс – станции отбора проб.
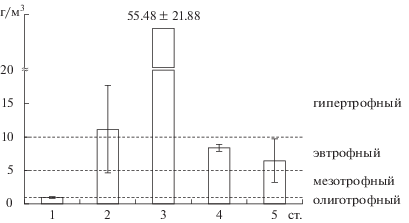
Пространственная неоднородность таксономического состава сопровождалась варьированием показателей количественного развития фитопланктона на станциях. В водной толще западной части озера (ст. 1 и 2) численность была 0.16–0.41 и 1.46–7.77 млн кл./л, биомасса – 0.71–1.25 и 5.21–23.14 г/м3 соответственно. В центральной (ст. 4) и восточной (ст. 5) части эти показатели изменялись в пределах 1.36–2.72 и 0.44–2.71 млн кл./л, 7.51–9.17 и 2.41‒12.46 г/м3. Максимальное обилие фитопланктона (97 г/м3) отмечено на ст. 3. В течение летнего сезона биомасса колебалась в диапазоне величин, свойственных водам разной трофии – от олиготрофных до гипертрофных (рис. 4).
Рис. 4.
Средняя за период наблюдения биомасса фитопланктона на станциях оз. Ключик. Справа от диаграммы указан трофический статус водоема, соответствующий данным значениям биомасс.
Основу численности и биомассы формировали диатомовые водоросли. На всей акватории озера развивалась центрическая диатомея Cyclotella distinguenda, которая всегда входила в состав доминантов и часто преобладала по биомассе над другими видами. В ее популяциях встречались только одиночные крупные клетки с диаметром створок от 11.4 до 40 мкм, достигая 34–40 мкм в нижних слоях воды. Число штрихов в 10 мкм менялось от 11 до 16, краевые выросты с тремя опорами располагались на каждом шестом интерштрихе, имелся один двугубый вырост.
В западной части озера наиболее значимыми видами были также диатомовые Melosira varians и Navicula radiosa Kütz., виды рода Pinnularia и рафидофитовые водоросли Vacuolaria sp. В зоне экотона формировались олигодоминантные комплексы, в них преобладала Cyclotella distinguenda в сопровождении бентосных форм Navicula radiosa и видов рода Pinnularia. В центральной и восточной частях сохранялось доминирование центрических диатомей с господством Cyclotella distinguenda, в группе содоминантных и сопутствующих пеннатных диатомовых наряду с видами рода Pinnularia, отмечены представители родов Ulnaria и Nitzschia. Заметной численности (до 20% общей) в этом районе достигали золотистые водоросли из рода Dinobryon, а по биомассе (до 40% общей) – динофлагелляты Ceratium hirundinella (O.F. Müller) Dujardin.
Вертикальное распределение фитопланктона в западной части озера (ст. 1) также отражало ведущую роль диатомовых водорослей, их доля в общих показателях обилия по всей глубине достигала 60–100% (рис. 5). В июле биомасса варьировала от 0.05 до 4.24 г/м3 с максимумом в зоне эпилимниона, в августе – от 0.01 до 1.52 г/м3 с максимумом у дна. От поверхности до глубины 6–7 м основу альгоценозов (до 89% биомассы) создавала Cyclotella distinguenda с незначительными показателями обилия (от 0.04 до 3.81 г/м3 и от 0.01 до 0.12 г/м3 в июле и августе соответственно); на глубине 8–9 м ее вклад снижался до 6–17%, уступая видам Melosira varians (до 72% общей биомассы) и Meridion circulare (Grev.) C. Agardh (до 18%).
Рис. 5.
Вертикальное распределение биомассы фитопланктона в оз. Ключик 9 июля (а, в, д) и 12 августа (б, г, е) 2017 г.: а, б – ст. 1, в, г – ст. 3, д, е – ст. 5. 1 – Chlorophyta, 2 – Bacillariophyta, 3 – Cyanophyta, 4 – Chrysophyta, 5 – Cryptophyta, 6 – Dinophyta, 7 – Raphidophyta, 8 – Euglenophyta.
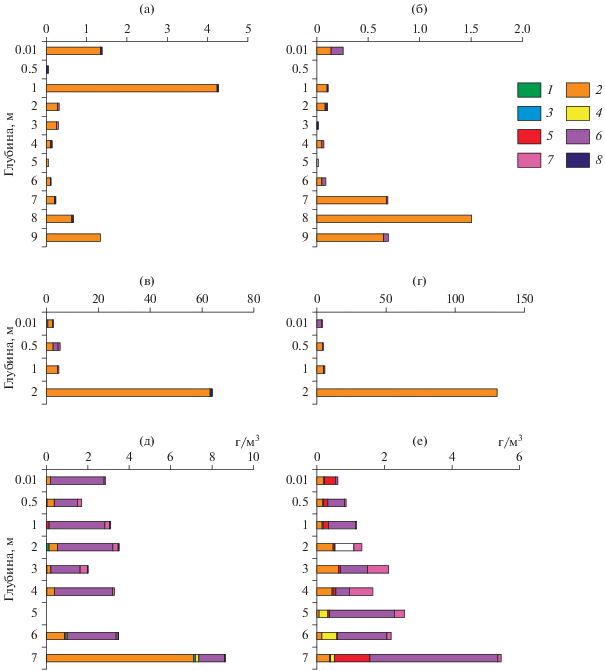
В восточной части (ст. 5) отмечена более сложная структура сообщества (рис. 5). На диатомовые водоросли приходилось 2–40% общей численности и 3–50% биомассы, лишь в июле в придонных слоях биомасса была до 80%. Cyclotella distinguenda сохраняла свое лидирующее положение, но при более низких количественных показателях. Заметного развития достигали жгутиковые динофитовые Ceratium hirundinella и Peridinium cinctum Ehrenb. В июле их присутствие зарегистрировано на всех глубинах, доля в биомассе изменялась от 14 до 89%. В августе динофитовые водоросли с биомассой 1.5–3.8 г/м3 (65–72% общей) отмечены на глубине 5–7 м, в поверхностном и придонном слоях зафиксировано высокое обилие криптофитовых. На глубине 2–5 м обнаружен редкий для планктона водоемов бассейна Средней Волги вид рафидофитовых водорослей Vacuolaria sp., достигающий заметного развития.
В мелководной переходной зоне (ст. 3) вертикальное распределение фитопланктона было неоднородным. У поверхности численно преобладали диатомовые водоросли, по биомассе, суммарные величины которой достигали 2.9–3.6 г/м3, – динофлагелляты. Ниже 0.5 м общие показатели развития фитопланктона и вклад диатомовых резко возрастали, и на глубине 2 м (у дна) получены рекордно высокие для водоемов бассейна Средней Волги величины. В июле и августе численность достигала 29 и 65 млн кл./л соответственно, биомасса – 63 и 130 г/м3 при полном господстве (99–100%) диатомовых водорослей и массовом развитии Cyclotella distinguenda.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
К ведущим факторам формирования и развития альгоценозов карстовых водоемов относятся их морфометрические характеристики, минерализация (Уникальные…, 2001), рН воды, трофический статус (Гусев, 2011), а также особенности окружающего ландшафта (Палагушкина, 2004). Известно, что видовой состав водорослей связан со степенью минерализации водоема (Воронихин, 1953). В солоноватых озерах одного и того же региона качественный состав беднее, чем в пресных, но количественные показатели развития водорослей могут быть гораздо выше (Уникальные…, 2001).
Фитопланктон оз. Ключик существенно отличается от других карстовых водоемов лесной и лесостепной ландшафтно-географических зон Европейской России (Гусев, 2011; Палагушкина, 2004) по составу крупных таксонов (отделов). При относительно небогатом видовом разнообразии альгофлора оз. Ключик характеризуется высокой представленностью диатомовых и золотистых, но меньшим участием зеленых водорослей и цианопрокариот. Высокая доля диатомовых и низкая доля зеленых водорослей отмечена в Голубом озере Самарской обл. (Тарасова, 2010). Альгофлора оз. Ключик сформирована донными формами и водорослями обрастаний при бедности истинно планктонных видов. Преобладают виды, индифферентные к степени минерализации, а также алкалифилы, развивающиеся в олигощелочных водах. Удельное видовое богатство альгоценозов было невысоким в западной части озера и увеличивалось в зоне экотона.
Восточная часть озера, имеющая зеленоватые воды, характеризуется однородным составом фитопланктона. В голубоцветной западной части он распределен по акватории более дискретно, при этом четко выделяется самая глубокая зона – воклина (ст. 1). Слабое сходство альгоценозов разных станций западной и восточной частей озера, выявленное в середине лета (июль), может свидетельствовать о формировании двух водных масс с различным гидрохимическим составом и физико-химическими свойствами.
Черты уникальности просматриваются в составе преобладающих компонентов альгоценозов оз. Ключик и выделяют его в ряду карстовых озер разных регионов (Уникальные…, 2001; Горохова, Паутова, 2009; Ternjej et al., 2010). Относительное постоянство температуры придонного слоя при отсутствии ветрового перемешивания водной толщи малого лесного озера с котловиной сложной формы, создают благоприятные условия для массового развития диатомовых водорослей. Основной структурообразующий вид – Cyclotella distinguenda редко встречается в волжском бассейне. Морфология ее створок близка к литературному описанию (Genkal et al., 2019), но диаметр створок больше и расположение краевых выростов более редкое. Это наиболее репрезентативный представитель диатомовых, включенных в функциональную группу кодона B (Padisák et al., 2009), предпочитающих мезотрофные условия и чувствительных к развитию стратификации. Обычно C. distinguenda не достигает высокого обилия и не относится к доминантам озерного планктона, а в озерах, имеющих связь с водотоками, и речных системах Венгрии отмечен как редкий вид, требующий охраны (Kiss et al., 2012). Форма C. distinguenda var. distinguenda обнаружена в пелагической зоне озер с высоким содержанием биогенных веществ (Krammer, Lange-Bertalot, 1991); в литоральной и пелагической зонах щелочных олиготрофно-мезотрофных озер и прудов (Houk et al., 2010); в глубоководных карстовых озерах Хорватии (Udovič et al., 2017); в донных отложениях ледникового меромиктического озера Карантийской низменности в Австрии (Huber et al., 2007) и слабощелочного с повышенной жесткостью вод оз. Сименит в Турции (Ersanli, Gönülol, 2003).
Показатели обилия C. distinguenda в оз. Ключик характерны для интенсивного “цветения” воды гипертрофных водоемов, что не сочетается с физико-химическими характеристиками низко- или среднепродуктивных вод ‒ высокой прозрачностью и низким содержанием биогенов. Наличие большого количества инициальных створок на подъеме численности популяции свидетельствует об оптимальных условиях для развития вида, которые позволяют ему формировать в зоне экотона биомассу, свойственную высокопродуктивным водам.
Распределение фитопланктона по акватории озера было неоднородным. Варьирование численности и биомассы в 4.4–6.2 раза отмечено на станциях 2 и 5, в меньшей степени (в 1.2–2.6 раза) – на станциях 1 и 4, что может свидетельствовать о формировании здесь более стабильных условий для существования альгоценозов.
Вертикальное распределение фитопланктона в западной части озера характеризовалось наличием подповерхностного и придонного максимумов в июле и придонного в августе. На станциях с зеленоватыми водами биомасса возрастала с глубиной, а структура альгоценозов усложнялась. В мелководной переходной зоне, хорошо освещенной до дна, выявлена вертикальная стратификация фитопланктона с заглубленным максимумом биомассы, которая в десятки раз выше, чем на других станциях. При этом отмечено упрощение структуры и формирование олигодоминантных сообществ с массовым развитием одного вида – Cyclotella distinguenda.
Биомасса фитопланктона характеризует уровень трофии восточной части озера чаще как эвтрофный, реже – мезотрофный и заметно выше, чем в западной части, где статус зоны максимальных глубин меняется от олиготрофного до олиго-мезотрофного. Вертикальное распределения биомассы фитопланктона свидетельствуют о преобладании мезотрофных вод в основной водной массе озера, эвтрофных – в придонных слоях и гипертрофных – в зоне экотона.
Выводы. В альгофлоре карстового оз. Ключик выявлено 133 таксона водорослей из девяти отделов. Основу видового богатства, численности и биомассы фитопланктона формировали диатомовые водоросли при доминировании Cyclotella distinguenda. Основу флористического списка составляли бентосные, литоральные и истинно-планктонные формы с преобладанием космополитов (87.1%) и невысокой долей бореальных видов (12.9%). Распределение таксономического состава и показателей количественного развития фитопланктона характеризовалось пространственной неоднородностью. Максимальное обилие фитопланктона (130.1 г/м3) отмечено в зоне экотона (ст. 3), где формировались олигодоминантные комплексы с преобладанием Cyclotella distinguenda. Морфометрические характеристики оз. Ключик, высокая степень минерализации вод, благоприятные световые условия, низкий температурный фон и термостабильность водной толщи при наличии выраженного экотонного эффекта создавали условия для формирования высокопродуктивных (с уникально высокой для водоемов бассейна Волги биомассой) диатомовых альгоценозов, образованных не типичными для зоны хвойно-широколиственных лесов видами водорослей.
Список литературы
Бакка С.В., Киселева Н.Ю. 2009. Особо охраняемые природные территории Нижегородской области. Аннотированный перечень. Нижний Новгород: Мин-во экологии и природных ресурсов Нижегородской области.
Баянов Н.Г. 2019. Озеро Ключик – уникальный природный объект Нижегородского Поволжья // Тр. Гос. природ. биосфер. заповед. “Керженский”. Т. 9. С. 65.
Баянов Н.Г., Макеев И.С., Воденеева Е.Л. 2009. Фитопланктон и продукционно-деструкционные процессы в озере Светлояр // Вестн. Мордовского гос. ун-та. Сер. биол. Саранск. № 1. С. 218.
Баянов Н.Г., Логинов В.В., Кривдина Т.В. 2014. Озера юго-запада Нижегородской области // Труды Мордовского государственного заповедника им. П.Г. Смидовича. Саранск: Изд-во Мордов. ун-та. Вып. 12. С. 241.
Варгот Е.В., Гришуткин О.Г., Артаев О.Н. 2015. Результаты исследований водно-болотных комплексов окрестностей озера Крячек, Ульяновская область // Самарский науч. вестн. № 2 11. С. 41.
Воденеева Е.Л., Кулизин П.В. 2019. Водоросли Мордовского заповедника аннотированный список видов. Флора и фауна заповедников. Москва. Вып. 134.
Воронихин Н.Н. 1953. Растительный мир континентальных водоемов. Москва, Ленинград: Изд-во АН СССР.
Горохова О.Г., Паутова В.Н. 2009. Фитопланктон слабоминерализованных карстовых озер юга лесостепного Поволжья // Изв. Самар. научн. центра РАН. № 14. С. 671.
Гусев Е.С. 2011. Вертикальное распределение фитопланктона в небольших карстовых озерах центральной России // Водоросли: таксономия, экология, использование в мониторинге. Екатеринбург: УрО РАН. С. 160.
Ефремов Ю.В. 1988. Голубое ожерелье Кавказа. Ленинград: Гидрометеоиздат.
Козлов А.В., Тарасов И.А., Дедык В.Е. 2017. Эколого-гидрохимическая характеристика акватории озера “Ключик” Павловского района Нижегородской области // Совр. пробл. науки и образов. № 1. С. 126.
Корнева Л.Г. 2000. Разнообразие и структура фитопланктона некоторых слабоминерализованных лесных озер Вологодской области // Гидробиологические вопросы. Якутск: Изд-во Якутск. ун-та. Ч. 2. С. 94.
Кусковский B.C., Кривошеев A.C. 1989. Минеральные озера Сибири (юг Красноярского края). Новосибирск: Наука. Сибир. отд.
Методика изучения биогеоценозов внутренних водоемов. 1975. Москва: Наука.
Палагушкина О.В. 2004. Экология фитопланктона карстовых озер Среднего Поволжья: Автореф. дис. … канд. биол. наук. Казань.
Тарасова Н.Г. 2010. Состав альгофлоры планктона озера Голубое Самарская область // Самарская Лука: проблемы региональной и глобальной экологии. Т. 19. № 2. С. 157.
Уникальные экосистемы солоновато-водных карстовых озер Среднего Поволжья. 2001. Казань: Изд-во Казан. ун-та.
Шитиков В.К., Розенберг Г.С., Зинченко Т.Д. 2003. Количественная гидроэкология: методы системной идентификации. Тольятти: Ин-т экологии Волжского бассейна РАН.
Ersanli E., Gönülol A. 2003. Study on the phytoplankton and seasonal variation of lake Simenit Terme – Samsun, Turkey // Turkish J. Fisheries and Aquatic Sciences. V. 3. № 1. P. 29.
Genkal S.I., Okhapkin A.G., Vodeneeva E.L. 2019. To the morphology and taxonomy of Cyclotella distinguenda Bacillariophyta // Novosti sistematiki nizshikh rastenii. V. 53. № 2. P. 247‒254. https://doi.org/10.31111/nsnr/2019.532.247
Gusev E.S. 2008a. Photosynthetic pigments of plankton in some karst lakes of central Russia. Inland Wat. Biol. V. 1. № 3. 217–224. https://doi.org/10.1134/S1995082908030048
Gusev E.S. 2008б. Phytoplankton primary production in several karst lakes in central Russia. Inland Wat. Biol. V. 1. № 4. 356–361. https://doi.org/10.1134/S199508290804007X
Houk V., Klee R., Tanaka H. 2010. Atlas of freshwater centric diatoms with a brief key and descriptions. Part III. Stephanodiscaceae A: Cyclotella, Tertiarius, Discostella // Fottea. V. 10. P. 1.
Huber K., Klee R., Schmidt R. 2007. Distribution and morphological variability of Cyclotella-taxa in the late glacial of Längsee Austria // Proceedings of the 1st Central European Diatom Meeting. BGBM, Berlin. P. 59. https://doi.org/10.3372/cediatom.112
John J. 2012. A beginner’s guide to diatoms. Liechtenstein: A. R. G. Gantner Verlag.
Kiss K.T., Klee R., Ector L., Ács É. 2012. Centric diatoms of large rivers and tributaries in Hungary: morphology and biogeographic distribution // Acta Bot. Croat. V. 71. № 2. P. 311. https://doi.org/10.2478/v10184-011-0067-0
Krammer K., Lange-Bertalot H. 1991. Bacillariophyceae. 3. Teil: Centrales, Fragilariaceae, Eunotiaceae // Süsswasserflora von Mitteleuropa. Stuttgart: Gustav Fisher. V. 2. № 3. P. 1.
Okhapkin A.G., Genkal S.I., Scharagina E.M., Vodeneeva E.L. 2014. Structure and dynamics of phytoplankton in the Oka river mouth at the beginning of the 21th century // Inland Water Biology. V. 7. № 4. P. 357. https://doi.org/10.1134/S1995082914040142
Padisák J., Crossetti L. O., Naselli-Flores L. 2009. Use and misuse in the application of the phytoplankton functional classification: a critical review with updates // Hydrobiologia. V. 621. № 1. P. 1. https://doi.org/10.1007/s10750-008-9645-0
Rousseaux C.S., Gregg W.W. 2014. Interannual Variation in Phytoplankton Primary Production at a Global Scale // Remote Sensing. V. 6. № 1. P. 1. https://doi.org/10.3390/rs6010001
Salmaso N., Buzzi F., Cerasino L., et al. 2014. Influence of atmospheric modes of variability on the limnological characteristics of large lakes South of the Alps: a new emerging paradigm // Hydrobiologia. V. 731. № 1. P. 31. https://doi.org/10.1007/s10750-013-1659-6
Ternjej I., Plenkoviċ-Moraj A., Mihaljeviċ Z., Kerovec M. 2010. Spatial and temporal variation of plankton in a Mediterranean, karstic lake // Ekológia Bratislava. V. 29. № 1. P. 65. https://doi.org/10.4149/ekol_2010_01_65
Udovič M.G., Cvetkoska A., Žutinić P. et al. 2017. Defining centric diatoms of most relevant phytoplankton functional groups in deep karst lakes // Hydrobiologia. V. 788. № 1. P. 169. https://doi.org/10.1007/s10750-016-2996-z
Žutinić P., Udovič M.G., Borojević K., et al. 2014. Morpho-functional classifications of phytoplankton assemblages of two deep karstic lakes // Hydrobiologia. V. 740. № 1. P. 147. https://doi.org/10.1007/s10750-014-1950-1
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод