

Биология внутренних вод, 2020, № 6, стр. 584-591
Рост и раннее половое созревание кумжи Salmo trutta р. Алатсоя (Республика Карелия)
Е. Д. Павлов a, *, А. Г. Буш a, В. В. Костин a, Д. С. Павлов a
a Институт проблем экологии и эволюции им. А.Н. Северцова Российской академии наук
Москва, Россия
* E-mail: p-a-v@nxt.ru
Поступила в редакцию 19.03.2020
После доработки 15.06.2020
Принята к публикации 22.06.2020
Аннотация
Изучены особенности роста и раннего полового созревания кумжи реки Алатсоя (Республика Карелия). Возраст кумжи, обитающей в этой реке, как правило, не превышает трех лет, и однa из реализуемых жизненных стратегий – образование раносозревающих самцов. Раносозревающие самцы достигают половой зрелости в возрасте 1+, характеризуются бóльшими размерами по сравнению с пестрятками того же возраста и высоким темпом роста в первый год жизни. Самки созревают не раньше возраста 2+. Есть основание полагать, что формирование раносозревающих самцов в популяции кумжи р. Алатсоя начинается в течение первого лета жизни.
ВВЕДЕНИЕ
Кумжа Salmo trutta L., обычно достигающая половой зрелости к 3–4 годам (Hart, 1973), может созревать и раньше этого срока (Кузищин, 1997; Jonsson B., Jonsson N., 2011; Huusko et al., 2018). В реках Карелии, в том числе и в р. Алатсоя, встречаются раносозревающие особи кумжи (Павлов и др., 2019). По нашим наблюдениям, в р. Алатсоя раносозревающие особи отмечены чаще по сравнению с популяциями в других реках Карелии (Павлов и др., 2019).
Комплекс благоприятных факторов, сложившихся в речной системе или на отдельных ее участках в первый год жизни лососевых, создает условия для увеличения темпов роста рыб и стимулирует их на раннее созревание (Bohlin et al., 1994; Fleming, 1996; Jonsson B., Jonsson N., 2011; Metcalfe, 1998; Morgan, Metcalfe, 2001). Количество и доступность пищи относятся к важнейшим факторам, влияющим на формирование жизненной стратегии у лососевых рыб (Павлов, Савваитова, 2008; Павлов и др., 2007; Ferguson et al., 2019; Olsson et al., 2006). У кумжи раннее созревание самцов и самок рассматривается как одна из жизненных стратегий (Павлов, Савваитова, 2008).
На выбор жизненной стратегии в популяции оказывают влияние особенности роста рыб (Мина, 1986; Myers et al., 1986) и развития их половых желез (Груздева и др., 2013, 2017).
Цель работы ‒ изучение их связи и роли в формировании жизненной стратегии у кумжи из р. Алатсоя.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Длина р. Алатсоя 14 км, ее исток – оз. Алатунлампи (Государственный…). Она впадает в р. Янисйоки, которая в свою очередь впадает в Ладожское озеро. Река Янисйоки выше (в районе пос. Харлу) и ниже (в районе пос. Ляскеля) по течению от места впадения р. Алатсоя перекрыта плотинами (рис. 1). Нижняя плотина блокирует возврат половозрелой кумжи из Ладожского озера на нерест в р. Алатсоя, а верхняя – блокирует контранатантную миграцию кумжи на нагул в оз. Янисъярви. Таким образом, кумжа, обитающая в р. Алатсоя, изолирована от других популяций вида.
Рис. 1.
Карта района проведения работ (а) и схема озерно-речной системы (б): 1 – район работ, 2 – плотины.
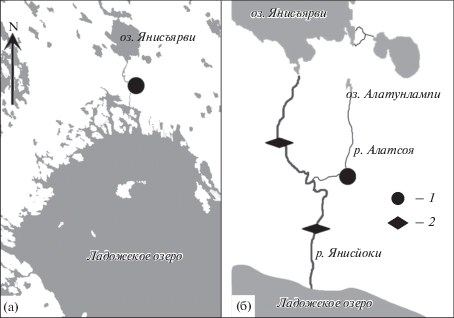
Кумжа разного возраста отловлена 28 и 29 августа 2015 г. в р. Алатсоя (61°48.02′ с.ш., 31°1.92′ в.д.) (бассейн Ладожского озера) щадящим методом с использованием электролова (Fa-2, Норвегия) в количестве 69 экз. Дополнительно 29 августа 2019 г. отловлены 18 экз. сеголетков (0+) кумжи из этой реки.
У рыб измеряли длину тела по Смитту (FL) и массу тела, определяли пол и принадлежность к той или иной группировке: пестрятки или раносозревающие рыбы. Пестрятки – неполовозрелые особи, у которых еще не началась дифференциация по жизненным стратегиям; раносозревающая кумжа – самцы в возрасте 1+ с гонадами на переходной III–IV или IV стадиях зрелости и самки в возрасте 2+ с гонадами на IV стадии зрелости.
Для определения возраста кумжи отбирали несколько десятков чешуй выше боковой линии между задним краем спинного плавника и началом анального (Мартынов, 1987). Возраст кумжи определяли по числу годовых колец на чешуе с цельной центральной зоной, не имеющей признаков регенерации и повреждения (Чугунова, 1959). У каждой особи просматривали ≥10 чешуй. Одинаковый участок тела рыб, с которого отбирали чешую, обеспечивал снижение ошибки определения возраста, поскольку число склеритов на чешуе, расположенной на разных участках тела вариативно (Чернова, Дгебуадзе, 2008). Радиусы склеритов измеряли в краниальной зоне чешуи с отклонением ≤45° от длиннейшей оси.
Обратное расчисление длины тела рыб в возрасте 1+ проводили по формуле Ли (Busaker et al., 1990): li – c = ri/R(FL – c), где li − вычисляемая длина тела особи в i-м возрасте, ri – радиус i-го годового кольца на чешуе, R – полный радиус чешуи, FL – конечная длина по Смитту, с – свободный член, вычисляемый по уравнению связи радиуса чешуи с длиной рыбы: ln FL = c + ln R.
Для оценки длины тела кумжи к концу первого лета жизни, измеряли сеголетков, пойманных в августе 2019 г. Затем сравнивали полученные значения с обратно расчисленной длиной годовиков в конце первого года жизни для определения доли прироста длины тела в летние месяцы.
Гонады для цитологического исследования фиксировали в жидкости Буэна. Гистологические препараты изготавливали с использованием полуавтоматического гистопроцессора TPC-15, заливочной станции TES-99, микротома Meditome M530 (“Medite”, ФРГ). Срезы толщиной 5 мкм окрашивали гематоксилином Эрлиха и эозином. Фотографии срезов половых желез получены при помощи моторизованного микроскопа Keyence Biorevo BZ-9000 (Япония). Пробы отобраны у 67 экз. рыб.
У самок для выявления цитологических изменений в генеративной ткани рассчитывали диаметр и ЯЦО на гистологических срезах (Павлов и др., 2014). ЯЦО рассчитывали как отношение площади ядра к площади цитоплазматического материала ооцита на срезе, прошедшем непосредственно вблизи от центральной части клетки. Измерения для определения ЯЦО, а также подсчет числа клеток в семеннике проводили при помощи программного обеспечения Image J. Ver. 1.8.0. Всего измерено 1227 ооцитов. У раносозревающих самок определяли относительную плодовитость – количество ооцитов, приходящихся на 1 г массы тела особи.
Статистическая обработка материала выполнена по индивидуальным значениям с использованием параметрического и непараметрического дисперсионных анализов, а также t-критерия Стьюдента.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Характеристика исследованных рыб. В р. Алатсоя были отловлены пестрятки, раносозревающие самцы и самки, а также недифференцированные по полу сеголетки (табл. 1). В период проведения работ смолты не встречались. Сеголетки имели темно-серую окраску, пестрятки – серую с оливковым оттенком и с ярко выраженными черными поперечными полосами вдоль тела. Раносозревающая кумжа сохраняла окраску, характерную для пестряток, но половые железы таких рыб были более развиты – находились на III–IV и IV стадиях зрелости, у пестряток – только на II и II‒III стадиях.
Таблица 1.
Возрастная структура и биологические показатели кумжи р. Алатсоя
| Пол, возраст | Длина, см | Масса, г | n, экз. |
|---|---|---|---|
| Сеголетки | |||
| 0+ | $\frac{{6.1 \pm 0.15}}{{5.2 - 7.1}}$ | $\frac{{2.5 \pm 0.19}}{{1.5 - 4.2}}$ | 18 |
| Пестрятки | |||
| ♂, 1+ | $\frac{{12.8 \pm 0.28}}{{10.8 - 15.3}}$ | $\frac{{22.4 \pm 1.57}}{{13.4 - 38.9}}$ | 21 |
| ♀, 1+ | $\frac{{12.7 \pm 0.24}}{{9.8 - 14.8}}$ | $\frac{{21.9 \pm 1.21}}{{9.9 - 33.5}}$ | 31 |
| ♀, 2+ | $\frac{{17.9}}{{15.8,{\text{ }}20.0}}$ | $\frac{{60.3}}{{39.8,\,80.8}}$ | 2 |
| Раносозревающие особи | |||
| ♂, 1+ | $\frac{{13.5 \pm 0.17}}{{12.4 - 14.5}}$ | $\frac{{29.0 \pm 1.31}}{{19.6 - 37.4}}$ | 13 |
| ♀, 2+ | $\frac{{20.5 \pm 0.5}}{{20.0,{\text{ }}21.0}}$ | $\frac{{91.8 \pm 3.95}}{{87.8,{\text{ }}95.7}}$ | 2 |
Пестрятки и раносозревающая кумжа имели сходный возраст – 1+ и 2+ (табл. 1). Две раносозревающие самки были в возрасте 2+, раносозревающие самцы пойманы только в возрасте 1+, на их долю приходилось 38% всех пойманных самцов.
Особенности роста рыб. Непараметрический дисперсионный анализ показал, что длина и масса рыб достоверно (p < 0.01, H-критерий Краскела–Уоллиса) различаются у пестряток и раносозревающих особей. Кроме того, различия в массе тела достоверны (p < 0.01) и по t-критерию Стьюдента.
Для оценки особенностей роста самцов кумжи в разные годы жизни определены расчисленные длины тела в первый год жизни, а по ним темпы роста рыб в первый и второй (неполный) годы жизни. Двухфакторный дисперсионный анализ темпов роста самцов кумжи из разных группировок и разного возраста (в разные годы жизни) показал, что они достоверно зависят от возраста рыб (p <0.001) и совместного влияния факторов (p = = 0.002). Последнее указывает на то, что темпы роста пестряток и раносозревающих самцов достоверно по-разному изменяются в первый и второй годы жизни. На первом году жизни будущие раносозревающие особи вырастают до 7.7 ± 0.15 см, пестрятки – до 6.7 ± 0.17 см, т.е. раносозревающие особи обогнали в росте пестряток – их рост был на 15% больше. К середине второго года жизни темп роста (прирост длины тела) у раносозревающих самцов снизился до 5.8 ± 0.21 см. У пестряток он тоже оказался ниже, чем в первый год, но выше, чем у раносозревающих самцов – 6.2 ± ± 0.21 см, т.е. на втором году жизни раносозревающие самцы отставали в росте от пестряток на 6%.
Длина сеголетков в августе достигала 89% расчисленной длины годовиков в конце первого года жизни.
Две раносозревающие самки на первом году жизни также характеризовались большей длиной (обратное расчисление) по сравнению с пестрятками самками – 8.5 ± 0.47 (7.9–9.5) см и 6.6 ± 0.15 (5.4–8.6) см соответственно.
Состояние половых желез самок. У пестряток в возрасте 1+ яичники находились на II стадии зрелости. На этой стадии зрелости гонады небольшие, имеют желтоватый цвет, представлены ооцитами периода превителлогенеза, которые характеризуются крупным ядром и цитоплазмой, имеющей гомогенную структуру (рис. 2а). Яичники одной из пестряток в возрасте 2+ также были на II стадии зрелости и по цитологическому строению не отличались от яичников пестряток в возрасте 1+. По периферии цитоплазмы превителлогенных ооцитов видны темные, часто прерывающиеся кольцевые зоны скопления РНК. В ядерном материале клеток локализованы ядрышки, в среднем 12 ± 1.4 (1‒33) штук на срезе. Диаметр превителлогенных ооцитов 97 ± 1.31 (16–231) мкм, а ЯЦО – 0.4 ± 0.01 (0.1–2.4).
Рис. 2.
Яичники кумжи Salmo trutta (показаны стрелкой). а ‒ ооциты превителлогенеза яичников на II стадии зрелости; б, в ‒ крупные ооциты вителлогенеза яичников на III стадии зрелости; г ‒ крупные ооциты, целиком заполненные гомогенным желтком (IV стадия зрелости). Масштаб: а – 300 мкм, б, г – 20 мм, в – 800 мкм.
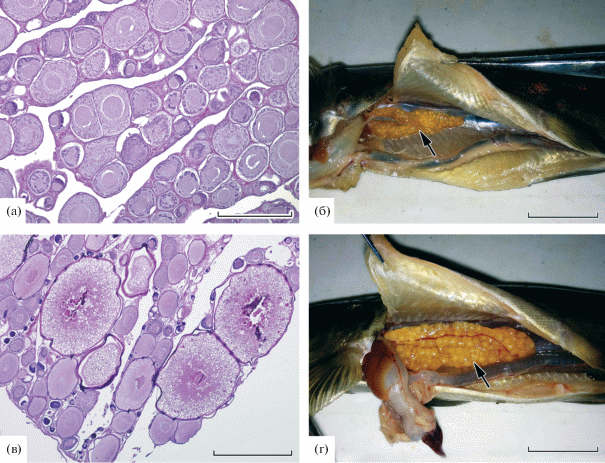
Яичники одной пестрятки в возрасте 2+ находились на III стадии зрелости (рис. 2б). Они имели желто-оранжевый цвет, в таких гонадах невооруженным глазом видны крупные светло-желтые ооциты. Половые железы были массой 0.3 г и содержали ооциты диаметром 108 ± 6.9 (29‒255) мкм периода превителлогенеза и диаметром 582 ± ± 64.2 (328‒802) мкм периода вителлогенеза. Бóльшая часть цитоплазмы вителлогенных ооцитов была вакуолизирована, в наиболее крупных клетках появлялись гранулы желтка (рис. 2в). Число ооцитов периода вителлогенеза в гонадах не превышало 10% всех половых клеток на срезе. Вероятно, половой зрелости самка достигла бы в возрасте 3+.
Две раносозревающие самки в возрасте 2+ имели крупные гонады IV стадии зрелости. Яичники занимали значительную часть полости тела, имели оранжево-красный цвет и содержали крупные ооциты, целиком заполненные гомогенным желтком (рис. 2г). Вдоль половых желез хорошо виден крупный кровеносный сосуд arteria ovarica. Масса гонад была 2.7 г (относительная плодовитость 0.22 ооцита) и 9.0 г (относительная плодовитость 0.31 ооцит).
У самок не обнаружено корреляции (p > 0.05) годовых приростов с ЯЦО и диаметром ооцитов.
Состояние половых желез самцов. У самцов степень созревания гонад положительно коррелирует (0.34, p < 0.05) с величиной прироста чешуи первого года жизни, массой (0.47, p < 0.01) и с принадлежностью к какой-либо группе (раносозревающие особи или пестрятки) (0.67, p < 0.001).
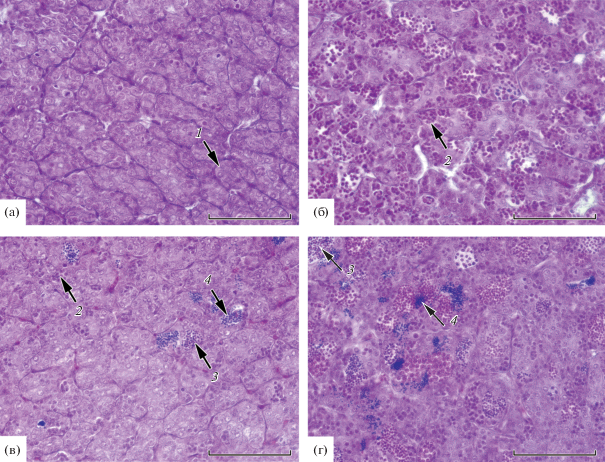
Семенники у пестряток были на II стадии зрелости, имели вид тонких полупрозрачных тяжей. Такие гонады содержат сперматогонии, которые локализованы в цистах и находятся еще в процессе митотического деления. Клетки следующей генерации либо отсутствуют, либо единичны (рис. 3а). Гонады других пестряток-самцов были на III стадии зрелости, кроме сперматогониев в них многочисленны сперматоциты I и II порядка (рис. 3б). Сперматоциты II порядка в ~2 раза мельче сперматоцитов I порядка. В некоторых семенниках сформированы немногочисленные сперматиды и сперматозоиды (рис. 3в), в других гонадах их количество превалирует над сперматоцитами (рис. 3г).
Рис. 3.
Семенники пестряток кумжи Salmo trutta на II (а) и III (б, в, г) стадиях зрелости: 1 – сперматогонии, 2 – сперматоциты, 3 – сперматиды, 4 – сперматозоиды. Масштаб: 100 мкм.
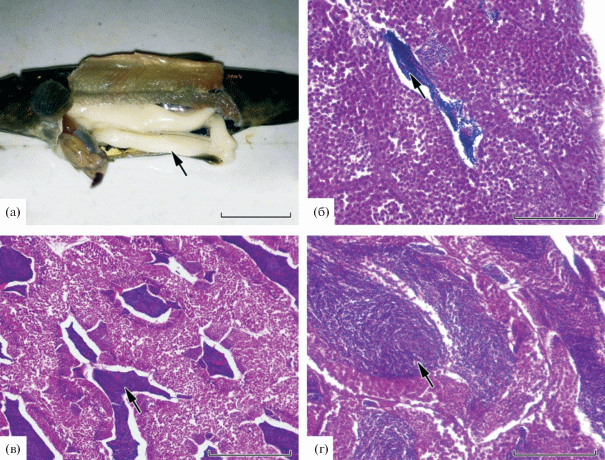
У раносозревающих самцов половые железы находились на переходной III–IV и IV стадиях зрелости. Семенники белого цвета, занимали бóльшую часть полости тела (рис. 4а), их масса – 2.2 ± 0.29 (0.7–3.4) г. В менее развитых гонадах бóльшую часть клеток представляли сперматоциты I и II порядка, а также единичные цисты со сперматидами и сперматозоидами (рис. 4б). Наличие в генеративной ткани сперматид свидетельствует о начале спермиогенеза. Размеры сперматид более мелкие по сравнению со сперматоцитами II порядка, окраска темнее (интенсивнее воспринимают гемотоксилиновый лак). Сперматозоиды отличаются от других половых клеток самцов наименьшими размерами, продолговатой формой и наличием жгутика. Начало IV стадии зрелости гонад характеризуется увеличением в генеративной ткани числа сперматоцитов II порядка и сперматид; сперматозоиды по-прежнему единичны. В дальнейшем количество сперматозоидов увеличивается (рис. 4в). Наиболее развитые гонады содержат семенные канальца со сперматозоидами и немногочисленные клетки раннего состояния ‒ сперматоциты и сперматиды (рис. 4г).
Рис. 4.
Гонады раносозревающих самцов кумжи Salmo trutta: a – внешний вид семенника (→); б – III–IV стадии зрелости, сперматозоиды немногочисленны (→); в, г – на IV стадии зрелости, по мере созревания число сперматозоидов (→) в семенных канальцах возрастает. Масштаб: а – 20 мм, б, г – 100 мкм, в – 200 мкм.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Полученные результаты свидетельствуют, что возраст кумжи в р. Алатсоя, как правило, не превышает 2+ лет, а одна из жизненных стратегий – образование раносозревающих особей. Отметим, что в других реках Фенноскандии (Huusko et al., 2018), в том числе в Карелии (Pavlov et al., 2019), возраст кумжи обычно достигает 3–5 лет. Вторая особенность кумжи в р. Алатсоя – высокая (38%) доля раносозревающих особей по сравнению с популяциями этого вида в других реках Карелии (Pavlov et al., 2019).
Зарегулирование стока р. Янисйоки привело к изоляции популяции кумжи из р. Алатсоя от крупных рек и озер, обеспечивающих нагул половозрелых рыб. Это обусловило уменьшение численности производителей в р. Алатсоя и, соответственно, снижение численности поколений. Увеличение доли раносозревающих самцов до 48% на фоне снижения численности популяции отмечено и в южной части ареала у черноморской кумжи Salmo trutta labrax (Pallas, 1814) – в р. Мзымта (Pavlov еt al., 2010). Снижение численности поколений, соответственно и пищевой конкуренции, привело к увеличению темпов роста и доли раносозревающих особей. Такое явление было отмечено у другого представителя сем. Salmonidae – нерке Oncorhynchus nerka (Walbaum, 1792) (Варнавская, Варнавский, 1988).
Период онтогенеза в первый год жизни является определяющим для раннего созревания лососевых (Jonsson B., Jonsson N., 2011). Темпы роста атлантического лосося Salmo salar (L., 1958) отрицательно коррелируют с возрастом достижения им половой зрелости (Hutchings, Jones 1998; Jonsson B., Jonsson N., 2011; Thorpe, 1994). У симы Oncorhynchus masou (Brevoort, 1856), как и у кумжи, раносозревающие самцы могут достигать половой зрелости в возрасте 1+ (Груздева и др., 2013), у мальмы Salvelinus malma (Walbaum, 1792) – только в возрасте 2+ (Груздева и др., 2017). При этом темпы роста раносозревающих особей симы и мальмы в первые годы жизни выше, чем у пестряток. Следовательно, в популяциях лососевых, относящихся к разным родам (атлантический лосось, кумжа, сима, мальма), прослеживается одинаковая закономерность – у наиболее быстрорастущих особей часто формируется траектория развития, направленная на раннее созревание. Сходные результаты получены и в нашей работе – выявлена группировка кумжи с созреванием в раннем возрасте, которое начинается на первом году жизни. К концу первого года будущие раносозревающие особи достоверно обгоняют пестряток по длине и массе тела.
Косвенные данные авторов позволяют уточнить период начала формирования указанной стратегии. Сравнение длин тела сеголетков (отловленных в конце августа) и обратно расчисленных длин тела у кумжи в конце первого года жизни показали, что прирост длины кумжи в период интенсивного роста (в летние месяцы) достигает 89% годового прироста. То есть, кумжа в течение первого года жизни преимущественно растет в летний период, осенью и зимой прирост длины тела незначительный, что соответствует литературным данным (Шустов и др., 2012; Jonsson B., Jonsson N., 2011). Наиболее вероятно, что различия в темпах роста пестряток и будущих раносозревающих особей закладываются в период интенсивного роста – летом. Это, в свою очередь, указывает на начало формирования у кумжи р. Алатсоя уже к концу первого лета траектории онтогенеза, направленного на раннее созревание и формирование жизненной стратегии раносозревающих самцов. Таким образом, первая особенность роста раносозревающей кумжи из р. Алатсоя – ускоренный (по сравнению с пестрятками) рост в начале первого года жизни.
Изучение состояния половых желез кумжи показало, что яичники у пестряток, как правило, развиваются с одной скоростью и, к возрасту особей 1+ находятся обычно на II–III стадиях зрелости. В связи с этим раносозревающие самки достигают половой зрелости на год позднее, чем раносозревающие самцы. Известно, что самки кумжи в отличие от самцов более склонны к миграциям (анадромии и потамодромии), которые связаны с их поздним созреванием (≥3+). Самцы, напротив, чаще остаются в пресной воде (García-Vega et al., 2018; Ferguson et al., 2017, 2019; Huusko et al., 2018; Jonsson et al., 2001). Не вызывает сомнений, что сложившийся половой диморфизм в проявлении миграционных жизненных стратегий у кумжи обусловливает частое формирование раносозревающих самцов в популяции и гораздо более редкое (единичное) раносозревающих самок.
Вторая характерная черта роста у самцов кумжи тесно связана с особенностями их полового созревания. На второй год жизни различия в темпах роста между раносозревающими особями и пестрятками сглаживаются – раносозревающие самцы растут медленнее, и самцы-пестрятки начинают догонять их по этому показателю. В то же время самцы в возрасте 1+ значительно различаются по скорости созревания гонад: медленное созревание (II стадия зрелости к возрасту 1+) характерно для пестряток, а быстрое (III–IV стадии зрелости к возрасту 1+) – для раносозревающих самцов. Известно (Jonsson B., Jonsson N., 2011), что созревающие рыбы расходуют свои ресурсы на созревание, соответственно соматический рост замедляется. Авторы указанной работы отмечают, что, очевидно, у лососевых присутствует связь между зависимыми от условий обитания темпами роста и последующей жизненной стратегией рыб. Быстрое созревание гонад у кумжи следует рассматривать в качестве ингибитора роста раносозревающих самцов на втором году жизни.
Выводы. Кумжа в р. Алатсоя характеризуется высокой долей (38%) раносозревающих самцов, достигающих половой зрелости в возрасте 1+, и наличием раносозревающих самок. Самки созревают не раньше возраста 2+. Особенность полового созревания двухлеток кумжи (1+) – синхронное развитие гонад у самок и асинхронное у самцов. Разделение самцов пестряток кумжи из р. Алатсоя на раносозревающих самцов и недифференцированную молодь обусловлено их различиями в темпах роста в первый год жизни и скоростью созревания половых желез. По-видимому, в р. Алатсоя формирование раносозревающих самцов кумжи начинается в первое лето жизни.
Список литературы
Варнавская Н.В., Варнавский В.С. 1988. О биологии карликовой формы нерки озера Дальнего (Камчатка) // Биол. моря. № 2. С. 16.
Государственный водный реестр. http://textual.ru/gvr.
Груздева М.А., Кузищин К.В., Павлов Е.Д. и др. 2017. Морфофизиологические закономерности формирования жизненных стратегий мальмы Salvelinus malma Камчатки // Вопр. ихтиологии. Т. 57. № 5. С. 587.
Груздева М.А., Малютина А.М., Кузищин К.В. и др. 2013. Закономерности формирования жизненной стратегии у симы Oncorhynchus masou реки Коль (западная Камчатка) в связи с процессами роста и полового созревания // Вопр. ихтиологии. Т. 53. № 5. С. 687.
Кузищин К.В. 1997. Особенности формирования внутривидовой разнокачественности у кумжи Salmo trutta L. Белого моря: Дис. … канд. биол. наук. Москва: МГУ.
Мартынов В.Г. 1987. Сбор и первичная обработка биологических материалов из промысловых уловов атлантического лосося. Сыктывкар: АН СССР. Уральское отд. Коми науч. центр.
Мина М.В. 1986. Микроэволюция рыб. Москва: Наука.
Павлов Д.С., Савваитова К.А. 2008. К проблеме соотношения анадромии и резидентности у лососевых рыб (Salmonidae) // Вопр. ихтиологии. Т. 48. № 6. С. 810.
Павлов Д.С., Лупандин А.И., Костин В.В. 2007. Механизмы покатной миграции молоди речных рыб. Москва: Наука.
Павлов Д.С., Павлов Е.Д., Ганжа Е.В. и др. 2014. Цитологическое состояние гонад и уровень тиреоидных и половых стероидных гормонов у двух фенотипических форм молоди черноморской кумжи Salmo trutta labrax // Вопр. ихтиологии. Т. 54. № 4. С. 470.
Павлов Д.С., Ганжа Е.В., Немова Н.Н. и др. 2019. Уровень тиреоидных и половых стероидных гормонов у кумжи Salmo trutta L. в реках Карелии // Биол. внутр. вод. Т. 12. №2, вып. 1. с. 87–92. https://doi.org/10.1134/S0320965219020116
Чернова О.Ф., Дгебуадзе Ю.Ю. 2008. Архитектоника чешуи костистых рыб и ее диагностическое значение: теоретические основы современных методов экспертного исследования. Москва: Наука.
Чугунова Н.И. 1959. Руководство по изучению возраста и роста рыб. Москва: Изд-во АН СССР.
Шустов Ю.А., Тыркин И.А., Белякова Е.Н. 2012. Особенности роста молоди кумжи Salmo trutta L. в водоемах европейского севера России // Труды Карельск. науч. центра РАН. № 1. С. 94.
Bohlin T., Dellefors C., Faremo U. 1994. Probability of first sexual maturation of male parr in wild sea-run brown trout (Salmo trutta) depends on condition factor 1 yr in advance // Can. J. Fish. and Aquat. Sci. V. 51. P. 1920. https://doi.org/10.1111/faf.12396
Busaker G.P., Adelman I.K., Goolish E.M. 1990. Growth // Methods for fish biology. V. 1. Maryland: Amer. Fish. Soc. P. 363.
Ferguson A., Reed T.E., Mcginnity P., Prodöhl P.A. 2017. Anadromy in brown trout (Salmo trutta): A review of the relative roles of genes and environmental factors and the implications for management and conservation // Sea Trout: Science and Management – Proceedings of the 2nd International Sea Trout Symposium Publisher: Matador, Leicestershire, UK.
Ferguson A., Reed T.E., Cross T.F. et al. 2019. Anadromy, potamodromy and residency in brown trout Salmo trutta: the role of genes and the environment // J. Fish Biol. P. 1. https://doi.org/10.1111/jfb.14005
Fleming I.A. 1996. Reproductive strategies of atlantic salmon: Ecology and evolution // Rev.: Fish Biol. Fish. V. 6. P. 379.
García-Vega A., Sanz-Ronda F.J., Fernandes Celestino L. et al. 2018. Potamodromous brown trout movements in the North of the Iberian Peninsula: Modelling past, present and future based on continuous fishway monitoring // Science of the Total Environment. V. 640–641. P. 1521. https://doi.org/10.1016/j.scitotenv.2018.05.339
Hart J.L. 1973. Pacific fishes of Canada // Bull. Fish. Res. Board Can. V. 180.
Hutchings J.A., Jones M.E.B. 1998. Life history variation in growth rate thresholds for maturity in atlantic salmon // Can. J. Fish. and Aquat. Sci. V. 55. № 1. P. 22.
Huusko A., Vainikka A., Syrjänen J.T. et al. 2018. Life-History of the adfluvial brown trout (Salmo trutta L.) in Eastern Fennoscandia // Brown Trout: Biology, Ecology and Management. P. 267. https://doi.org/10.1002/9781119268352.ch12
Jonsson B., Jonsson N. 2011. Ecology of atlantic salmon and brown trout: habitat as a template for life histories // Fish Fisheries Ser. V. 33.
Jonsson B., Jonsson N., Brodtkorb E., Ingebrigtsen P.-J. 2001. Life-history traits of Brown Trout vary with the size of small streams // Functional Ecology. V. 15. № 3. P. 310.
Metcalfe N.A. 1998. The interaction between behavior and physiology in determining life history patterns in atlantic salmon (Salmo salar) // Can. J. Fish. and Aquat. Sci. V. 55. P. 93.
Morgan I.J., Metcalfe N.B. 2001. Deferred costs of compensatory growth after autumnal food shortage in juvenile salmon // Proceedings of the Royal Society of London Series B-Biological Sciences. V. 268. P. 295.
Myers R.A., Hutchings J.A., Gibson R.J. 1986. Variation in male parr maturation within and among populations of Atlantic salmon, Salmo salar // Can. J. Fish. and Aquat. Sci. V. 43. № 6. P. 1242.
Olsson I.C., Greenberg L.A., Bergman E., Wysujack K. 2006. Environmentally induced migration: The importance of food // Ecology Letters. V. 9. P. 645.
Pavlov D.S., Kostin V.V., Nechaev I.V. et al. 2010. Hormonal Status in Different Phenotypic Forms of Black Sea Trout Salmo trutta labrax // J. Ichthyol. V. 50. № 11. P. 985.
Thorpe J.E. 1994. Reproductive strategies in Atlantic salmon Salmo salar L. // Aquacult. Res. V. 25. № 1. P. 77.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод