

Биология внутренних вод, 2020, № 6, стр. 528-537
Конвергенции в архитектонике мускулатуры трематод и соотношение понятий “конвергенция” и “параллелизм”
М. В. Ястребов a, *, Е. Н. Глаголева a, И. В. Ястребова a
a Ярославский государственный университет им. П.Г. Демидова
Ярославль, Россия
* E-mail: mvy@uniyar.ac.ru
Поступила в редакцию 17.01.2020
После доработки 26.03.2020
Принята к публикации 12.05.2020
Аннотация
Описаны новые примеры конвергенций в архитектонике мускулатуры трематод. В их числе полная редукция диагональной мускулатуры на малоподвижных участках покровов, характер распределения дорсо-вентральных мышц в пределах вентральных впадин, наличие радиальной паренхимной мускулатуры в полых радиально-симметричных участках тела, пространственное распределение протракторов глотки, вынос периферических слоев мускулатуры ротовой присоски на поверхность данного органа, формирование в ротовой присоске хордальных мышц и другие. Обсуждена обоснованность разделения понятий “конвергенция” и “параллелизм”. Названы недостатки таких критериев разделения, как степень родства филогенетических групп, занятие адаптивных зон разного масштаба и формирование на основе эволюции аналогичных или гомологичных органов. Возведение параллелизма в ранг самостоятельного эволюционного явления оценено как нецелесообразное.
ВВЕДЕНИЕ
Конвергенции, понимаемые как независимое формирование сходных признаков в разных филогенетических группах организмов, широко распространены в живой природе. Известны десятки примеров конвергенций, многие из которых стали хрестоматийными. Примеры охватывают широкий круг объектов от одноклеточных форм до высших растений и животных. Однако среди объектов почти отсутствуют паразиты, в частности, крупные и бурно эволюционирующие классы плоских червей (Trematoda, Cestoda, Monogenea), внимание которым уделено в данном контексте всего в нескольких публикациях (Рыжиков, Ошмарин, 1981, 1985).
Одна из задач настоящей работы – представление новых примеров конвергенций в архитектонике мускулатуры трематод. Вторая задача – обсуждение вопроса о соотношении понятий “конвергенция” и “параллелизм”, который остается дискуссионным в течение десятилетий.
МАТЕРИАЛ И МЕТОДЫ
В основу работы положены выполненные нами описания мышечной системы более тридцати видов трематод из девяти отрядов и девятнадцати семейств (Ястребов и др., 2000; Ястребова и др., 2008; Смирнова и др., 2010; Бурдакова, Ястребова, 2011; Бурдакова и др., 2012; Ястребов, Ястребова, 2014; Burdakova et al., 2015; Glagoleva et al., 2019). Для световой микроскопии отобраны одноразмерные половозрелые черви каждого вида. Материал фиксирован 75%-ным этанолом. Тотальные препараты окрашены уксуснокислым кармином. Архитектоника мускулатуры покровов, присосок, глотки, а также пространственное распределение паренхимных мышц изучены по полным сериям сагиттальных, поперечных и фронтальных парафиновых срезов толщиной 7–10 мкм, изготовленных по стандартной методике и окрашенных методом Маллори.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
Примеры конвергенций
В покровах марит толщина отдельного слоя мускулатуры определяется не только величиной, но и формой поперечного сечения мышечных волокон. В частности, при одинаковой площади сечения овальных и поперечно-овальных волокон мышечный слой, состоящих из первых, будет толще. Изменение формы сечения волокон с типичной овальной или округлой на поперечно-овальную, по-видимому, – один из способов эволюционной регулировки толщины мышечных слоев в покровах трематод. Поперечно-овальная форма сечения мышц во всех или в части слоев независимо сформировалась у Halipegus ovocaudatus (Vulpian, 1859) (Halipegidae), Haplometra cylindracea (Zeder, 1800) (Plagiorchiidae), Brandesia turgida (Brandes, 1888) (Pleurogenidae) и Eucotyle cohni Skrjabin, 1924 (Eucotylidae). Эти виды относятся к так называемым статофильным гельминтам, использующим пассивную фиксацию (Ошмарин, 1959, 1960). Их общая экологическая черта – обитание в условиях, где сохранение локализации не требует больших мышечных усилий. В этих условиях мускулатура покровов становится тонкой на фоне размеров тела. У второго и третьего видов все мышцы покровов поперечно-овальные в сечении, у двух других – только кольцевые. Для реофильных трематод, живущих под угрозой утраты локализации, обсуждаемая форма сечения мышц не характерна.
В покровах трематод кольцевой слой почти всегда представлен одним правильным рядом мышечных волокон. Изредка слой утолщается на значительной части тела за счет многоярусности. У Clinostomum foliiforme Braun, 1899 (Clinostomidae) она наблюдается на спинной и брюшной сторонах тела от переднего конца до середины полового отдела. В заднем сегменте тела Сardiocephalus longicollis (Rudolphi, 1819) (Strigeidae) кольцевые мышцы также распределены в несколько “этажей”, и толщина слоя в 4–5 раз больше, чем в переднем сегменте. В данном случае конвергентное сходство архитектоники кольцевых мышц обусловлено необходимостью приложения мощных мышечных усилий при жизни в механически агрессивной среде, каковой является пищеварительная трубка птиц. Оно позволяет эффективно суживать и вытягивать соответствующие участки тела, которые довольно массивны. Интересно, что для продольной мускулатуры покровов многоярусность не характерна. Причина, по-видимому, в том, что у многих реофильных трематод хорошо развиты продольные паренхимные мышцы – синергисты аналогичных мышц покровов.
Конвергенциями являются не только приобретения, но и утраты. К ним можно отнести редукцию на отдельных участках покровов диагональной мускулатуры, которая считается характерным атрибутом плоских червей (Mair et al., 1998; Tyler, Hooge, 2004). Например, она отсутствует в адоральном диске Patagifer bilobus (Rudolphi, 1819) (Echinostomatidae), в хвосте Lecithochirium sp. (Lecithochiriidae) и в части покровов некоторых парамфистоматид (Krupenko, 2013). У ряда представителей семейства Strigeidae она не обнаружена в органе Брандеса и в лопастях стенки переднего сегмента тела. При этом в переднем сегменте тела Pseudoapatemon tiaratus Mamaev, 1959 ее нет совсем, а у Ichthyocotylurus platycephalus (Creplin, 1825) Odening, 1969 и Cardiocephalus longicollis (Rudolphi, 1819) Szidat, 1928 – на значительной части покровов. Общей чертой перечисленных участков является относительная малоподвижность, в особенности отсутствие скручиваний, которые обеспечиваются как раз диагональными мышцами. При малоподвижном образе жизни такого вида, как Collyriclum faba (Bremser in Schmalz, 1831) (Collyriclidae), наблюдается даже полная редукция диагональной мускулатуры (Быховская-Павловская, Хотеновский, 1964).
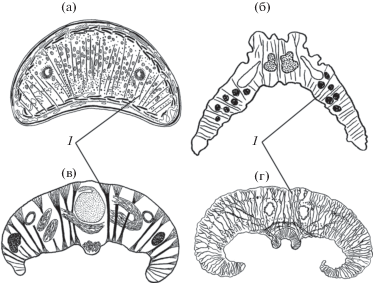
Во многих семействах трематод (Notocotylidae, Echinostomatidae, Cephalogonimidae, Clinostomidae, Mesometridae, Gastrodiscidae и другие) для усиления фиксации на теле конвергентно формируются вентральные впадины, работающие как большие присоски (Ошмарин, 1960; Bartoli, Ukoli, 1970; 1987; Ястребов, Фрезе, 1997). В отр. Strigeidida использование всего тела (подотряд Cyathocotylata) или его переднего сегмента (подотряд Strigeata) в качестве большой присоски – общий принцип фиксации (Судариков, 1959). Некоторые авторы (Ошмарин, Жарикова, 1984; MacKinnon, 1982) отмечают в пределах вентральных впадин хорошо развитую дорсо-вентральную мускулатуру. Она является функциональным аналогом радиальных мышц присосок, и ее архитектоника в разных группах трематод обретает ряд сходных черт: общее интенсивное развитие, расположение самых тонких мышечных волокон на боковых сторонах тела и увеличение толщины мышц и размеров их терминальных конусов по мере разреживания (рис. 1).
Рис. 1.
Архитектоника дорсо-вентральных мышц (1) трематод в вентральных впадинах на поперечных срезах тела: (а) Clinostomum complanatum, (б) Parafasciolopsis fasciolaemorpha, (в) Tetraserialis tscherbakovi, (г) Alaria alata.
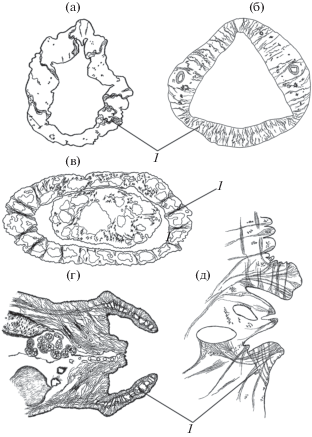
В ряде таксонов трематод тело мариты имеет радиально-симметричный участок с полостью внутри. К таким участкам относятся циркумкаудальная складка в основании хвостового придатка (представители разных семейств отр. Hemiurida), вентральная камера (Gasrtothylacidae), передний сегмент тела (многие Strigeidae) и половой атриум (разные семейства отрядов Paramphistomatida и Strigeidida). В подобных участках конвергентно формируются радиальные паренхимные мышцы, регулирующие объем полости (рис. 2).
Рис. 2.
Радиальные мышцы (1) в радиально-симметричных участках тела трематод: (а) циркумкаудальная складка Lecithochirium sp., (б) вентральная камера Gastrothylax crumenifer (Gastrothylacidae), (в) передний сегмент тела Pseudoapatemon tiaratus, (г) половой атриум Ophiosoma patagiatum (Strigeidae), (д) половой атриум Gastrodiscoides hominis (Gastrodiscidae); (а–в) на поперечных, (г–д) на срединных сагиттальных срезах тела.
Есть интересный пример конвергентного формирования коротких, обычно продольных мышц в различных фиксаторных структурах, имеющих форму двускатной крыши. Эти мышцы соединяют “скаты” и, сокращаясь, стягивают их, вызывая выступание соответствующей структуры (рис. 3). Они обнаружены в кольцевом мышечном валике на теле Eucotyle сohni Skrjabin, 1924, в гребневидных выступах на поверхности тела Typhlocoelium sisowi (Diesing, 1850) (Cyclocoelidae) и в адоральных дисках Potagifer bilobus (Rud., 1819) и Cortinasoma ocadiae Oshmarin & Zharikova, 1984 (Pronocephalidae) (Ошмарин, Жарикова, 1984). В перечисленных случаях обеспечивается прикрепление разными способами. Кольцевой валик распирает мочевой каналец хозяина. Гребневидные выступы, идущие в разных направлениях, формируют на теле гельминта ячеистый рельеф, усиливающий сцепление с тканями хозяина. Адоральные диски в ходе фиксации меняют форму, в частности, уплощаются и расширяются. В стенке ротовой присоски Diplodiscus mehrai Pande, 1937 (Diplodiscidae) на границе основной части и дивертикулов обнаружены такие же продольные мышцы, каждая из них соединяет две точки покровов полости органа. Одна точка находится в задней части собственно присоски, другая – в начальном участке дивертикула. Сокращение этих мышц замыкает вход в дивертикул, выдвигая участок стенки органа в его просвет.
Рис. 3.
Продольные мышцы (1) в “двускатных” фиксаторных структурах трематод на срединных сагиттальных срезах тела: (а) адоральный диск Patagifer bilobus, (б) кольцевой мышечный валик Eucotyle cohni, (в) гребневидные выступы покровов Typhlocoelum sisowi.

Трематоды Parafasciolopsis fasciolaemorpha Ejsmont, 1932 (Fasciolidae), Leucochloridium macrostomum (Rudolphi, 1802) (Leucochloridiidae), Bolbophorus confusus (Krause, 1914) (Diplostomidae), Haematotrephus lanceolatus (Wed’l, 1857) (Cyclocoelidae) и некоторые другие виды имеют сходную архитектонику протракторов глотки, обусловленную одинаковой функциональной нагрузкой. Заметим, что мы называем их протракторами условно, за неимением более точного термина (Ястребов, Ястребова, 2014). Сходство наблюдается по нескольким параметрам. Во-первых, протракторы крепятся не к самой глотке, а к пищеварительной трубке впереди и позади нее. Во-вторых, места крепления расположены предельно близко к глотке. Вследствие этого, мышечные волокна лежат прямо на органе. В-третьих, число протракторов велико, и они формируют на поверхности глотки правильный слой с регулярным расположением сократительных элементов. Лишь по местам крепления впереди и позади органа удается понять, что это паренхимные, а не собственно глоточные мышцы. Описанные особенности указывают, что перемещение органа вперед и назад стремится к нулю из-за близости точек крепления мышц к глотке. Протракторы производят продольное сжатие органа и являются синергистами наружных продольных мышц глотки. Это повышает эффективность накачивания пищи в кишечник.
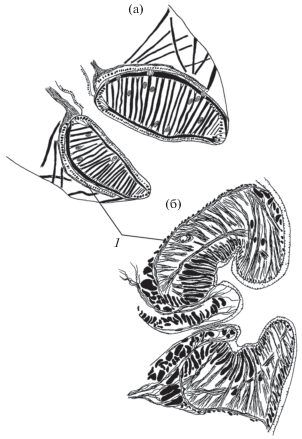
Число мышечных групп и особенности их пространственного распределения в ротовых присосках трематод заметно меняются в эволюции. Периферическая мускулатура ротовой присоски обычно находится под оболочкой органа. При этом в разных таксонах имеются виды, у которых наружные кольцевые мышцы располагаются поверх оболочки присоски, как это наблюдается у Parafasciolopsis fasciolaemorpha и Patagifer bilobus (рис. 4). Присоски обоих видов плотно заполнены мускулатурой. Поэтому возможная причина выноса мышечного слоя на поверхность органа – дефицит пространства внутри присоски для размещения мышечных элементов и площадей для их крепления. Данная особенность архитектоники встречается у трематод не только с мощными, но и с относительно слабыми ротовыми присосками, например, у Haptometra сylindracea (Zeder, 1800) и Eucotyle сohni. Слабость выражается в тонких мышечных слоях и в разреженности радиальной мускулатуры. Возможно, вынос наружного кольцевого слоя на поверхность органа произошел у предков этих видов, имевших более мощные присоски.
Рис. 4.
Наружные кольцевые мышцы (1) ротовых присосок трематод на срединных сагиттальных срезах тела: (а) Parafasciolopsis fasciolaemorpha, (б) Patagifer bilobus.
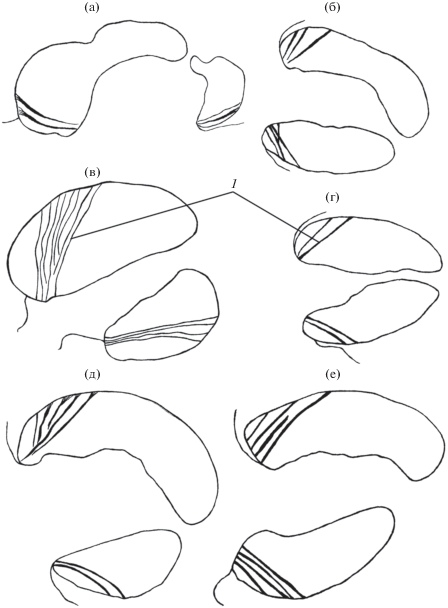
В работах обобщающего характера среди сведений об устройстве ротовых присосок трематод (Nasmark, 1937; Гинецинская, 1968; Nachtigall, 1974; Быховская-Павловская, 1983; Smyth, Halton, 1983) не встречается упоминаний о мышцах, названных нами хордальными (Ястребов, Ястребова, 2014). Подобно хорде окружности, каждая такая мышца соединяет две точки на внутренней стороне оболочки присоски. Одна из точек чаще всего расположена в устье органа, другая – позади него. Проекция хордальных мышц на ось симметрии присоски может быть как очень небольшой, так и более половины длины органа (рис. 5). Их функциональная нагрузка состоит в укорочении присоски и в изменении размера ее устья. Эта группа мышц найдена у одиннадцати видов из семейств Azygiidae, Clinostomidae, Echinostomatidae, Lecithochiriidae, Halipegidae, Leucochloridiidae, Plagiorchiidae, Notocotylidae, Eucotylidae, Strigeidae. Перечень семейств указывает на довольно широкое распространение хордальной мускулатуры. Однако она не является признаком класса, и есть сомнения в том, что это общий признак какого-либо отряда или семейства трематод. Например, она отсутствовала в ротовых присосках Paramphistomatida (шесть видов из пяти семейств), а среди девяти видов отр. Strigeidida из семейств Strigeidae, Diplostomidae и Alariidae была найдена лишь у Ichtyocotylurus platycephalus. По-видимому, хордальные мышцы конвергентно формировались у отдельных видов в разных отрядах или семействах и при дальнейшем видообразовании могли стать атрибутами видов одного рода или близких родов. Наличие хордальных мышц – показатель относительно высокого разнообразия движений ротовой присоски, связанного с обеспечением не только прикрепления, но и питания.
Рис. 5.
Схемы расположения хордальных мышц (1) в ротовых присосках трематод на срединных сагиттальных срезах тела: (а) Ichthyocotylurus platycephalus, (б) Leucochloridium macrostomum, (в) Halipegus ovocaudatus, (г) Tetraserialis tscherbakovi, (д) Azygia robusta, (е) Clinostomum complanatum.
Можно предположить, что у различных мягкотелых животных хордальная мускулатура – не редкость. Ее наличие ожидаемо в стенках полых органов, при работе которых расширение и сужение полостей или отверстий сочетается с определенной степенью продольного сжатия. Например, у трематод, она имеется в присоскоподобном органе Брандеса Alaria аlata (Goeze, 1782) (Alariidae), в половых атриумах Tylodelphys podicipina (Diplostomidae) и Pseudoapatemon tiaratus (Strigeidae) и в мышечном колпачке на переднем конце тела Stichorchis subtriquetrus (Rudolphi, 1814) (Cladorchidae).
Присоски трематод обычно имеют радиальную симметрию, но в некоторых случаях архитектоника мускулатуры придает им признаки билатеральной симметрии, которые конвергентно возникают у представителей разных таксонов. Так, общая черта Lecithochirium sp., Halipegus ovocaudatus и Azygia robusta Odhner, 1911 (Azygiidae) – наличие крупной ротовой присоски с субтерминальным устьем. В ее дорсальной части кольцевая мускулатура развита гораздо лучше, чем в вентральной. Продольная мускулатура, напротив, лучше развита в вентральной части присоски. Одновременное сокращение кольцевых мышц в дорсальной части органа и продольных – в вентральной поворачивает ротовое отверстие к субстрату, способствуя прикреплению. Устья брюшных присосок Lecithochirium sp. и Patagifer bilobus также субтерминальные. Как следствие, различия в степени развития кольцевых и продольных мышц в дорсальной и вентральной половинах этих органов такие же, как в ротовых присосках. Различия между частями брюшной присоски Lecithochirium sp. усиливает архитектоника хордальных мышц: они имеют кольцевой ход в дорсальной половине органа и продольный – в вентральной.
Глотка трематод анатомически довольно консервативна (Иоффе, Чубрик, 1988). Сфинктеры, если они имеются, обычно образованы утолщенными наружными продольными мышцами, внутренними продольными или теми и другими вместе. Известен один пример конвергентного формирования нетипично крупного переднего сфинктера в глотках Parafasciolopsis fasciolaemorfa Eismont, 1932 и Bolboforus сonfusus (Krause, 1914). У обоих видов мышечные волокна сфинктера с огромной суммарной площадью поперечного сечения диффузно распределены в стенке глотки не только в устье, но и позади него, заходя назад более чем на треть ее длины. У первого вида глотка сопоставима по размерам с ротовой присоской, у второго – крупней ее. Возможно, выдающееся развитие сфинктера – результат участия не только в накачивании пищи в кишечник, но и в фиксации, при которой ткани хозяина втягиваются глубоко в глотку и зажимаются мощной кольцевой мускулатурой.
Замечания о понятиях “конвергенция” и “параллелизм”
Описанные выше примеры конвергенций могут быть расценены другими исследователями как примеры эволюционных параллелизмов. Это не удивительно, поскольку соотношение понятий “конвергенция” и “параллелизм” не установлено точно и, как следствие, отсутствуют их однозначные общепринятые определения.
В основе обоих обсуждаемых понятий лежит одно и то же эволюционное явление – независимое формирование сходных признаков в разных филогенетических группах. Причиной сходства признаков считается существование групп в сходных условиях (Шмальгаузен, 1939; Георгиевский, 1985). Между тем существование в сходных условиях само по себе – не причина возникновения конвергенций или параллелизмов, поскольку к одинаковым условиям организмы обычно приспосабливаются разными способами. По всей видимости, причина конвергентного сходства – сходные способы освоения ресурсов среды (Иорданский, 2001). Однако представление о способах освоения ресурсов среды является по сути экологическим, отсылающим нас к разработке понятия “экологическая ниша” (Пианка, 1981). При этом имеется еще более привлекательный подход в терминах эволюционной теории: конвергенции и параллелизмы обусловлены реакцией организмов на сходные давления отбора (Майр, 1968). Последняя позиция считается наиболее широкой, вмещая в себя и наличие сходных условий среды, и сходные способы ее освоения.
До этого места в определениях конвергенции и параллелизма все более или менее совпадает. Вопросы начинаются с момента их разграничения, т.е. с необходимости определить, какие именно филогенетические группы попадают в сферу действия каждого из понятий. В этой связи нередко указывают, что параллелизмы возникают в родственных филогенетических группах, а конвергенции – в неродственных (Шмальгаузен, 1969; Иорданский, 1979; Грант, 1991). В данном контексте слово “неродственные” звучит странно, ибо является завуалированной формой отрицания естественной системы живой природы, основанной на эволюционном родстве биологических видов. Кроме того, в большинстве известных примеров конвергенций факт родства сравниваемых групп очевиден. Правильнее говорить о близком или дальнем родстве (Воробьева, 1980), хотя отличить одно от другого не представляется возможным, и Берг (1977) справедливо отмечает, что представления о близости родства зависят от точки зрения.
Иной способ разграничения понятий состоит во мнении, что конвергенции осуществляются на разной организационной основе, а параллелизмы – на общей (Северцов, 1981, 1987) однако смысл “организационная основа” автором не уточнялся. Другие авторы вместо организационной основы говорят об основе наследственной, которая, подобно организационной, является общей при параллелизмах и разной при конвергенциях (Майр, 1974; Парамонов, 1978). Замена слов не снимает проблему критериев общей и разной основ, и критерии появятся не раньше, чем будут введены определения понятий “организационная основа” или “наследственная основа”, применимые к филогенетическим группам от рода до царства.
Была предпринята попытка установить различия между конвергенциями и параллелизмами через понятие “адаптивная зона”, определяемое как комплекс условий внешней среды, в которой протекает эволюция конкретного таксона. Считается, что конвергенция происходит при независимом вселении конвергирующих таксонов в адаптивную зону. Появление параллелизма связано с эволюцией в исходной для анцестрального таксона адаптивной зоне при ее дроблении на подзоны (Северцов, 1990). К сожалению, термин “адаптивная зона” воспринимается в основном интуитивно и, по ряду причин, недоступен для практического использования. Во-первых, ни для одного таксона на уровне, к примеру отряда, такая зона не охарактеризована в измеряемых показателях. Какие-то измерения еще возможны для абиотических факторов среды. Однако неясно, как оценивать биотические взаимодействия всех видов таксона с пищевыми объектами, хищниками, конкурентами, паразитами и т.д. Это особенно сложно для групп, где виды на различных стадиях жизненного цикла освоили все среды жизни – наземно-воздушную, водную, почвенную и организменную (Plathelminthes, класс Nematoda, отряд Diptera и др.). Добавим, что введенное в определение слово “комплекс” подразумевает не просто набор факторов среды, а их сложную констелляцию, основанную на сопряженном действии. Возможность ее качественного описания вызывает большие сомнения. Во-вторых, наблюдаемые сейчас конвергенции и параллелизмы формировались зачастую в далеком геологическом прошлом, что исключает точную оценку условий среды того времени. В-третьих, учитывать все условия не нужно в принципе, поскольку лишь малая их часть связана с появлением конкретных конвергенций.
Некоторые авторы переводят понятие “параллелизм” в дочернее по отношению к понятию “конвергенция”. Параллелизм определяют сходным образом как “конвергентную эволюцию филогенетически родственных групп организмов” (Иорданский, 1979, с. 164), либо как форму конвергентного развития генетически близких групп (Яблоков, Юсуфов, 1989). Данный перевод оставляет нерешенным вопрос об оценке степени родства и лишь подчеркивает отсутствие различий между конвергенцией и параллелизмом. Заметим также, что в обеих цитируемых работах есть методологически неверные оговорки, а именно, “конвергентная эволюция” и “конвергентное развитие”. Ранее отмечалось, что эволюция любого таксона дивергентна по определению, а конвергенция – лишь следствие независимых дивергентных изменений, приводящих к сходству таксонов (Рыжиков, Ошмарин, 1985).
Перечисленные трудности предположительно лежат в основе суждения, что “по-видимому, ввести универсальный, не формальный критерий различения конвергенции и параллелизма нельзя” (Северцов, 1990, с. 163). Между тем такой критерий давно известен. Суть его в том, что конвергенции осуществляются на основе эволюции аналогичных органов, а параллелизмы – на основе эволюции гомологичных (Шмальгаузен, 1969; Тимофеев-Ресовский и др., 1977; Северцов, 1981; Георгиевский, 1985; Яблоков, Юсуфов, 1989). Наличие критериев гомологии (Remane, 1952) позволяет уйти от оценок степени родства организмов и открывает путь к новым формулировкам обсуждаемых понятий. В этом случае конвергенцию можно определить как независимое формирование в разных филогенетических группах сходных признаков на основе эволюции аналогичных структур. Для определения параллелизма требуется заменить слово “аналогичных” на “гомологичных”.
Если принять эти определения, потребуется переосмысление места и роли конвергенций и параллелизмов в эволюции органического мира. В частности, придется признать, что конвергенция – довольно редкое природное явление, поскольку формирование эволюционного сходства аналогичных структур иллюстрируется небольшим числом примеров. К ним относятся сходство крыльев насекомых и птиц, медуз и медузоподобного жгутиконосца Medusochloris phiale Pascher, 1917, растительных колючек листового и стеблевого происхождения, роющих конечностей крота европейского и медведки, сложно устроенных глаз Craniata и Cephalopoda и некоторые другие. Желающий расширить набор примеров легко убедится, что тот окажется относительно кратким. Напротив, подавляющее большинство классических примеров конвергенций потребуется перевести в разряд параллелизмов по причине гомологии сравниваемых органов. Так, известное сходство между акулами, ихтиозаврами и китообразными, между летающими ящерами и рукокрылыми млекопитающими, между современными Equidae и вымершими Litopterna, между многими видами сумчатых и их плацентарными эквивалентами, между стеблевыми суккулентами в разных группах цветковых растений основано на сходстве именно гомологичных структур. В итоге список параллелизмов станет огромен по сравнению со списком конвергенций.
На этом можно было бы остановиться, если бы не одно обстоятельство. Общие закономерности эволюции филогенетических групп должны быть, по логике вещей, применимы к любой группе, взятой в отдельности, а также к любым сравниваемым группам. Для формальных определений в данной области требуются только ключевые характеристики явлений без добавления частностей. В явлении конвергенции таковыми служат факт независимого формирования сходных признаков в разных группах и его причина – упоминавшиеся сходные давления отбора. Такие показатели как степень родства групп, основа формирования в виде аналогов или гомологов, временной фактор (синхронное или асинхронное формирование), структурный уровень (макромолекулы, ткани, морфология) – это как раз частности. Они показывают разнообразие проявлений конвергенции, но не определяют его сути, и потому от них можно абстрагироваться. На основании вышеизложенного, понятие конвергенции можно трактовать как независимое формирование сходных признаков в разных филогенетических группах под действием сходных давлений отбора. Разница между понятиями “конвергенция” и “параллелизм” отсутствует, и первое из них поглощает все явления, относящиеся ко второму.
Выводы. Гермафродитное поколение трематод обладает впечатляющей эволюционной пластичностью, выражающейся, в частности, в разнообразии внешнего строения. Одним из следствий разнообразия является большое число групп мышц (особенно паренхимных), зарегистрированных у трематод, и огромное число вариантов их пространственного распределения в теле марит. Несмотря на это, ряд сходных элементов внешнего строения появляется в эволюции разных таксонов трематод и влечет за собой конвергенции в архитектонике мускулатуры. Конвергенций, безусловно, больше, чем представлено в настоящей работе, поскольку мышечная система марит изучена крайне слабо. При сравнении мускулатуры представителей разных классов Plathelminthes число примеров возрастет, и исследования в этом направлении имеют хорошую перспективу.
В дискуссии о соотношении понятий “конвергенция” и “параллелизм” последняя точка не поставлена. У их разделения есть много сторонников, и наши суждения, несомненно, подвергнутся критике. Если критика позволит приблизиться к истине, значит эти суждения были небесполезны.
Список литературы
Берг Л.С. 1977. Номогенез, или эволюция на основе закономерностей // Труды по теории эволюции. Ленинград: Наука. С. 95.
Бурдакова Е.Н., Ястребова И.В. 2011. Ротовые присоски некоторых представителей отряда Paramphistomatida (Plathelminthes, Trematoda) // Вестник Ярославского регионального отделения РАЕН. Т. 5. № 3. С. 50.
Бурдакова Е.Н., Ястребова И.В., Ястребов М.В. 2012. Копулятивные аппараты некоторых представителей отряда Paramphistomatida (Plathelminthes, Trematoda) // Ярославский педагогический вестник. № 3. Т. III. С. 94.
Быховская-Павловская И.Е. 1983. Органы прикрепления трематод и их функции // Тр. Зоол. ин-та АН СССР. Т. 31. Ленинград: Наука. С. 76.
Быховская-Павловская И.Е., Хотеновский И.А. 1964. К морфологии сосальщика Collyriclum faba (Bremser, 1831) // Тр. Зоол. ин-та АН СССР. Т. 22. Москва: Наука. С. 207.
Воробьева Э.И. 1980. Параллелизмы и конвергенции в эволюции кистеперых рыб // Морфологические аспекты эволюции. Москва: Наука. С. 7.
Георгиевский А.Б. 1985. Дарвинизм. Москва: Просвещение.
Гинецинская Т.А. 1968. Трематоды, их жизненные циклы, биология и эволюция. Ленинград: Наука.
Грант В. 1991. Эволюционный процесс: критический обзор эволюционной теории. Москва: Мир. (Grant V. 1985. The evolutionary process. A critical review of evolutionary theory. New York: Columbia University Press.).
Иорданский Н.Н. 1979. Основы теории эволюции. Москва: Просвещение.
Иорданский Н.Н. 2001. Эволюция жизни. Москва: Академия.
Иоффе Б.И., Чубрик Г.К. 1988. Строение глотки трематод и филогенетические связи Trematoda с турбелляриями // Паразитология. Т. 22. № 4. С. 297.
Майр Э. 1968. Зоологический вид и эволюция. Москва: Мир. (Mayr E. 1965. Animal species and evolution. London: Oxford University Press).
Майр Э. 1974. Популяции, виды и эволюция. Москва: Мир. (Mayr E. 1970. Populations, species and evolution. Cambridge, Massachusetts: The Belknap Press of Harvard University Press).
Ошмарин П.Г. 1959. К изучению специфичной экологии гельминтов. Владивосток: Изд-во Дальневосточного научного центра АН СССР.
Ошмарин П.Г. 1960. К понятию “фиксация гельминтов” // Биологические ресурсы Дальнего Востока. Москва: Изд-во АН СССР. С. 182.
Ошмарин П.Г., Жарикова Т.И. 1984. Новый вид и род трематод Cortinasoma ocadiae gen. еt sp. nov. (Pronocephalata, Pronocephalidae) – паразит пресноводной черепахи из Вьетнама // Паразиты животных и растений. Владивосток: Изд-во Дальневосточного научного центра АН СССР. С. 44.
Парамонов А.А. 1978. Дарвинизм. Москва: Просвещение.
Пианка Э. 1981. Эволюционная экология. Москва: Мир. (Pianka E. 1978. Evolutionary ecology. New York: Harper and Row Publishers).
Рыжиков К.М., Ошмарин П.Г. 1981. Явление конвергенции у гельминтов // Современные проблемы эволюционной морфологии животных. Москва: Наука. С. 86.
Рыжиков К.М., Ошмарин П.Г. 1985. Конвергенция у гельминтов и вопрос о взаимоотношении понятий “конвергенция” и “параллелизм” // Морфологические исследования животных. Москва: Наука. С. 179.
Северцов А.С. 1981. Введение в теорию эволюции. Москва: Изд-во МГУ.
Северцов А.С. 1987. Основы теории эволюции. Москва: Изд-во МГУ.
Северцов А.С. 1990. Направленность эволюции. Москва: Изд-во МГУ.
Смирнова Д.А., Ястребов М.В., Ястребова И.В. 2010. Мышечная система Clinostomum foliiforme (Trematoda, Clinostomida) // Биоразнообразие и экология паразитов. Тр. Центра паразитологии ИПЭЭ РАН. Т. 46. Москва: Наука. С. 257.
Судариков В.Е. 1959. Отряд Strigeidida (LaRue, 1926) Sudarikov, 1959. Ч. 1. Морфологическая характеристика стригеидид и надсемейство Strigeoidea Railliet, 1919 // Трематоды животных и человека. Т. 16. Москва: Изд-во АН СССР. С. 219.
Тимофеев-Ресовский Н.В., Воронцов Н.Н., Яблоков А.В. 1977. Краткий очерк теории эволюции. Москва: Наука.
Шмальгаузен И.И. 1939. Пути и закономерности эволюционного процесса. Москва; Ленинград: Изд-во АН СССР.
Шмальгаузен И.И. 1969. Проблемы дарвинизма. Ленинград: Наука.
Яблоков А.В., Юсуфов А.Г. 1989. Эволюционное учение. Москва: Высшая школа.
Ястребов М.В., Фрезе В.И. 1997. Дифференцировки тела в эволюции трематод // Экологическое и таксономическое разнообразие паразитов. Труды Института паразитологии РАН. Т. 41. Москва. С. 214.
Ястребов М.В., Ястребова И.В. 2014. Мышечная система трематод (строение и возможные пути эволюции). Москва: Товарищество научных изданий КМК.
Ястребова И.В., Ястребов М.В., Фрезе В.И. 2000. Мускулатура стенки тела некоторых реофильных представителей класса Trematoda // Актуальные проблемы общей паразитологии. Труды Института паразитологии РАН. Т. 42. Москва: Наука. С. 348.
Ястребов М.В., Ястребова И.В., Филимонова Л.В. 2008. Об эволюционной пластичности мышечной системы гермафродитного поколения трематод (Plathelminthes, Trematoda) // Систематика и биология паразитов. Труды Центра паразитологии ИПЭЭ РАН. Т. 45. Москва: Наука. С. 184.
Bartoli P. 1987. Caracteres adaptatifs originaux des digenes intestinaux de Sarpa salpa (Teleostei, Sparidae) et leur interpretation en termes d’evolution // Ann. parasitol. hum. et comp., 1987. V. 62. № 6. P. 542.
Burdakova E., Yastrebova I., Yastrebov M. 2015. The Oral Sucker Muscles of Six Representatives of the Order Paramphistomatida (Plathelminthes, Trematoda) // Biology Bulletin. V. 42. № 2. P. 108.
Glagoleva E., Yastrebova I., Yastrebov M. 2019. Body Wall Musculature of Five Representatives of the Order Paramphistomatida (Plathelminthes, Trematoda) // Biology Bulletin. V. 46. № 2. P. 168.
Krupenko D. 2013. Muscle system of Diplodiscus subclavatus (Trematoda: Paramphistomida) cercariae, pre-ovigerous, and ovigerous adults // Parasitol. Res. V. 113. P. 941.
MacKinnon B. 1982. The histology, ultrastructure and histochemistry of the ventral surfaces of Catatropis verrucosa (Froelich, 1789) Odhner, 1905 and Paramonostomum alveatum (Mehlis in Creplin, 1846) Luhe, 1909 (Digenea: Notocotylidae) // Can. J. Zool. V. 60. № 10. P. 2434.
Mair G., Maule A., Shaw C., Halton D. 1998. Muscling in on parasitic flatworms // Parasitol. Today. V. 14. № 2. P. 73.
Nachtigall W. 1974. Biological mechanisms of attachment. Berlin: Springer.
Nasmark K. 1937. Revision of the trematode family Paramphistomatidae // Zoologiska Bidrag fran Uppsala. V. 16. P. 301.
Remane A. 1952. Die Grundlagen des Natürlichen Systems, der Vergleichenden Anatomie und der Phelogenetik. Leipzig: Geest & Portig K.-G.
Smyth J., Halton D. 1983. The physiology of trematodes. Cambridge: Cambridge Univ. Press.
Tyler S., Hooge M. 2004. Comparative morphology of the body wall in flatworms (Platyhelminthes) // Can. J. Zool. V. 82. P. 194.
Ukoli F. 1970. On the adhesive mechanisms of Apharyngostrigea simplex (Johnston, 1904) and Clinostomum tilapiae (Ukoli, 1966) // Niger. J. Sci. V. 4. № 1. P. 77.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод