

Биология внутренних вод, 2021, № 3, стр. 286-301
Жизненный цикл паразита амфибий нематоды Cosmocerca ornata (Nematoda: Cosmocercidae)
Н. Ю. Кириллова a, А. А. Кириллов a, *
a Самарский федеральный исследовательский центр Российской академии наук, Институт экологии Волжского бассейна Российской академии наук
Тольятти, Россия
* E-mail: parasitolog@yandex.ru
Поступила в редакцию 23.06.2020
После доработки 29.07.2020
Принята к публикации 21.10.2020
Аннотация
Впервые изучен жизненный цикл паразита амфибий нематоды Cosmocerca ornata (Dujardin, 1845). Проведено экспериментальное заражение сеголетков озерной лягушки Pelophylax ridibundus инвазионными личинками Cosmocerca ornata. Изучены стадии развития паразитов в организме хозяина. Установлено, что заражение амфибий инвазионными личинками (III возраст) нематод происходит в приповерхностном слое воды путем проникновения паразитов через глаза на коньюктиву под нижнее веко, где личинки претерпевают третью и четвертую линьки. Затем паразиты мигрируют в ротоглоточную полость и по пищеварительному тракту к месту постоянной локализации – задней трети кишечника. Приведены описания и оригинальные рисунки личинок III и IV возрастов, личинок в стадиях линек и половозрелых особей C. ornata. Стадии развития нематод диагностировали по размерам тела, расположению полового зачатка, формирующимся наружным половым органам. Пол нематод можно определить на стадии третьей линьки. Половозрелые особи отличаются от личинок наличием вульвы у самок и спикул у самцов. Выявлены сроки отдельных стадий развития С. ornata и общая продолжительность жизни паразита в хозяине. Установлено влияние температуры окружающей среды на период развития нематод в амфибиях.
ВВЕДЕНИЕ
Нематода Cosmocerca ornata (Dujardin, 1845) паразитирует у широкого круга бесхвостых и хвостатых амфибий, пресмыкающихся и распространена в Палеарктике повсеместно. Основными хозяевами служат бесхвостые земноводные (Anderson, 2000; Yildirimhan et al., 2009; Dusen et al., 2010). Цикл развития C. ornata до настоящего времени не был изучен. Развитие паразита, как и других представителей сем. Cosmocercidae, осуществляется прямым путем без участия промежуточных хозяев (Anderson, 2000).
Популяция C. ornata состоит из ларвальной гемипопуляции (личинки I‒III возрастов), находящейся во внешней (водной) среде, и гемипопуляции паразитов в амфибиях, которая делится на ларвальную (личинки III и IV возраста) и адультную (половозрелые самцы и самки) группировки (Кириллов, Кириллова, 2016).
Ранее нами экспериментально изучен личиночный этап развития свободноживущих личинок C. ornata (Кириллова, Кириллов, 2015). Установлено, что у этого вида развитие личинок происходит по открытому типу, при котором из яйца выходит личинка I возраста (Шульц, Гвоздев, 1970). Развитие личинок C. ornata до инвазионной стадии (III возраст) протекает в водной среде. Заражение амфибий паразитом происходит в воде, где инвазионные личинки C. ornata сохраняют жизнеспособность до месяца и более, не претерпевая последующих линек (Кириллова, Кириллов, 2015).
Цель работы ‒ изучить стадии цикла развития нематоды C. ornata в организме хозяина ‒ озерной лягушки Pelophylax ridibundus (Pallas, 1771) (Anura: Ranidae).
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
С 13 июля по 14 сентября 2014 г. на базе стационара “Кольцовский” Института экологии Волжского бассейна РАН (пос. Мордово) проведено экспериментальное заражение 318 сеголетков озерных лягушек инвазионными личинками C. ornata. В эксперименте использовали инвазионных личинок C. ornata III возраста, полученных в лабораторных условиях (Кириллова, Кириллов, 2015).
Для заражения брали стерильных (свободных от C. ornata) сеголетков, выращенных в лаборатории из головастиков, отловленных в протоке Студенка Мордовинской поймы Саратовского водохранилища (53°10′ с.ш., 49°26′ в.д.). Ранее нами установлено, что головастики неспособны заражаться C. ornata (Кириллова, Кириллов, 2015).
Сеголетков амфибий заражали личинками паразитов (по 5‒7 экз. на каждого) с помощью стеклянной пастеровской пипетки перорально, перназально и перанально. Проведено шесть серий эксперимента (по две серии на каждый путь заражения) на 48 сеголетках (по восемь особей на серию). Вскрытие сеголетков проводили непосредственно после заражения спустя 1 и 2 ч. Проверена возможность естественного проникновения личинок нематод в хозяев. Для этого сеголетков лягушек помещали в отдельные емкости (по три особи) с личинками C. ornata III возраста (по 20 экз.). Экспериментальное заражение проводили в стеклянных емкостях объемом 0.1 л. Кроме того, проведены наблюдения за возможным перкутанным проникновением личинок нематод при контакте с кожей сеголетков в воде. Для этого личинок нематод наносили в капле воды на кожу амфибий и наблюдали с помощью бинокуляра МБС-10.
При экспериментальном заражении изучено влияние температуры окружающей среды на поступление инвазионных личинок в хозяев и на сроки развития отдельных возрастных групп нематод. В лаборатории стационара среднесуточная температура воздуха в период наблюдений изменялась от 15 до 29°С. Температуру фиксировали 4 раза в сутки лабораторным термометром ЛТ-2. Первые вскрытия сеголетков проводили спустя 1 ч с момента посадки в емкости с личинками нематод, последующие – в течение 45 сут (табл. 1). Выполнено 10 серий эксперимента на 270 сеголетках (одна серия ‒ в девяти емкостях по три сеголетка).
Таблица 1.
Экстенсивность инвазии (ЭИ) и индекс обилия (ИО) Cosmocerca ornata в сеголетках озерной лягушки
| Время эксперимента, сут | N | Личинки III–IV возрастов | Половозрелые нематоды | Общая зараженность | |||
|---|---|---|---|---|---|---|---|
| ЭИ, % | ИО | ЭИ, % | ИО | ЭИ, % | ИО | ||
| 1 | 12 | 100 | 6.8 ± 1.3 | 0 | 0 | 100 | 6.8 ± 1.3 |
| 2 | 12 | 100 | 6.8 ± 0.8 | 0 | 0 | 100 | 6.8 ± 0.8 |
| 3 | 12 | 100 | 6.7 ± 0.5 | 0 | 0 | 100 | 6.7 ± 0.5 |
| 4 | 12 | 100 | 5.8 ± 0.6 | 0 | 0 | 100 | 5.8 ± 0.6 |
| 5 | 15 | 100 | 4.3 ± 0.4 | 13.3 ± 9.1 | 0.2 ± 0.1 | 100 | 4.5 ± 0.4 |
| 7 | 15 | 86.7 ± 9.1 | 3.3 ± 0.5 | 40.0 ± 13.1 | 1.2 ± 0.3 | 100 | 4.5 ± 0.4 |
| 9 | 12 | 66.7 ± 14.2 | 1.8 ± 0.6 | 83.3 ± 11.2 | 2.6 ± 0.5 | 100 | 4.3 ± 0.4 |
| 11 | 12 | 33.3 ± 14.2 | 0.4 ± 0.2 | 100 | 3.8 ± 0.3 | 100 | 4.2 ± 0.4 |
| 14 | 15 | 0 | 0 | 100 | 3.6 ± 0.3 | 100 | 3.6 ± 0.3 |
| 17 | 15 | 0 | 0 | 100 | 3.2 ± 0.3 | 100 | 3.2 ± 0.3 |
| 20 | 15 | 0 | 0 | 100 | 2.8 ± 0.3 | 100 | 2.8 ± 0.3 |
| 23 | 12 | 0 | 0 | 100 | 2.9 ± 0.3 | 100 | 2.9 ± 0.3 |
| 26 | 12 | 0 | 0 | 100 | 3.0 ± 0.3 | 100 | 3.0 ± 0.3 |
| 29 | 12 | 0 | 0 | 100 | 2.6 ± 0.3 | 100 | 2.6 ± 0.3 |
| 33 | 15 | 0 | 0 | 100 | 2.3 ± 0.2 | 100 | 2.3 ± 0.2 |
| 36 | 15 | 0 | 0 | 100 | 2.2 ± 0.2 | 100 | 2.2 ± 0.2 |
| 38 | 12 | 0 | 0 | 83.3 ± 11.2 | 2.1 ± 0.3 | 83.3 ± 11.2 | 2.1 ± 0.3 |
| 40 | 15 | 0 | 0 | 60.0 ± 13.1 | 1.1 ± 0.3 | 60.0 ± 13.1 | 1.1 ± 0.3 |
| 42 | 15 | 0 | 0 | 33.3 ± 12.6 | 0.5 ± 0.2 | 33.3 ± 12.6 | 0.5 ± 0.2 |
| 45 | 15 | 0 | 0 | 0 | 0 | 0 | 0 |
Примечание. Здесь и в табл. 3, 5: N ‒ количество исследованных сеголетков амфибий.
Разновозрастных личинок и половозрелых особей C. ornata изучали в живом состоянии на временных препаратах путем варьирования количества воды между предметным и покровным стеклами. Измерение паразитов проводили после их предварительного обездвиживания нагреванием в воде, при котором нематоды выпрямлялись. Всего измерено 176 личинок и 150 половозрелых паразитов.
Прослежено развитие 455 личинок C. ornata: 148 личинок III возраста с момента заражения до линьки, 92 личинки III возраста на стадии линьки в IV (76 самок и 16 самцов), 92 личинки IV возраста (77 самок и 15 самцов) и 123 личинки IV возраста на стадии линьки в половозрелую особь (103 самки и 20 самцов). Развитие половозрелых нематод изучено на 404 самках C. ornata: 139 ‒ I возрастной группы, 131 ‒ II, 85 ‒ III, 49 ‒ IV, и на 54 самцах: 28 ‒ I возрастной группы и 26 ‒ II. Возрастные группы половозрелых паразитов устанавливали по степени развития их половой системы (Кириллов, Кириллова, 2016а).
Наблюдения за развитием паразитов проводили с помощью микроскопов МБС-10, МБИ-9 и цифровой камеры-окуляра DCM-300. Рисунки нематод выполнены с временных препаратов с использованием рисовального аппарата РА-7.
Для характеристики зараженности озерных лягушек нематодами использовали следующие параметры: экстенсивность инвазии, встречаемость (ЭИ, %), интенсивность инвазии (ИИ, экз.), индекс обилия гельминтов (ИО), ошибка среднего (mx). Обработку данных проводили с использованием статистических программ. Для сравнения темпов поступления личинок нематод в хозяев и сроков развития отдельных возрастных групп нематод при разной среднесуточной температуре лаборатории использовали критерий Краскела–Уоллиса (H) и ранговый коэффициент корреляции Спирмена (rs). Различия считали достоверными при p < 0.05.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Пути инвазии хозяев C. ornata
Результаты эксперимента по заражению сеголетков перорально, перназально и перанально показали, что личинки нематод III возраста, поступившие в хозяина одним из перечисленных путей, не приживаются в сеголетках амфибий. Личинок C. ornata обнаруживали в ротовой полости, желудке и прямой кишке непосредственно после введения. В течение 1‒2 ч от момента заражения сеголетки оказались свободными от нематод. Наблюдения за возможным перкутанным проникновением личинок C. ornata в сеголетков также показали отрицательный результат ‒ личинки нематод при контакте с кожей амфибий в капле воды не пытались проникать через кожу.
При посадке сеголетков в емкости с водой, содержащей личинок III возраста, спустя 1 ч под нижним веком и мигательной перепонкой сеголетков обнаружены живые личинки нематод. Спустя 3 ч с момента посадки личинки C. ornata отсутствовали в воде. При вскрытии сеголетков личинки найдены в глазах амфибий на конъюктиве под нижним веком.
Описание личинок C. ornata
Личинка III возраста (рис. 1). Губы неразличимы. Стома вытянутая, узкая, без четкой границы с глоткой. Глотка тонкая, удлиненная; заканчивается колбовидным бульбусом. Нервное кольцо окружает глотку примерно на уровне ее середины. Вокруг глотки в области нервного кольца и несколько кпереди от него хорошо заметны шесть–восемь образований – аркадные клетки. Хвост длинный, острый. Примерно на уровне середины тела просматривается компактный овальный половой зачаток. Пол личинок на данной стадии по морфологическим признакам не идентифицируется.
Рис. 1.
Личинки Cosmocerca ornata III возраста: а – 1–2 ч с момента заражения, б – 1 сут с момента заражения. Масштаб: 0.2 мм.
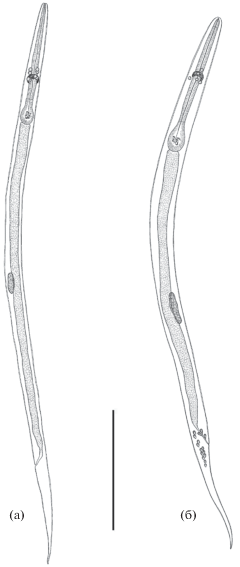
Личинка на стадии линьки III–IV (рис. 2). Губы слабо выражены. Стома укороченная, без четкой границы с глоткой. Число аркадных клеток сокращено до трех–четырех. Половой зачаток ≤1/8 длины тела. На этой стадии развития личинки самок морфологически отличаются от самцов – они значительно крупнее самцов, половой зачаток расположен примерно на уровне середины длины тела (рис. 2а). У самцов половой зачаток расположен постэкваториально, ближе к хвостовому концу, внутри клоаки отмечено формирование рулька, спикулы не просматриваются (рис. 2б).
Рис. 2.
Личинки Cosmocerca ornata на стадии линьки III–IV, 26 ч с момента заражения: а – самка, б – самец. Масштаб: 0.2 мм.

Таблица 2.
Морфометрическая характеристика (размеры, мм) личинок Cosmocerca ornata III–IV возрастов
| Показатель | III возраст (25 экз.) |
Линька III в IV | IV возраст | Линька IV в adult | |||
|---|---|---|---|---|---|---|---|
| самки (35) | самцы (16) | самки (25) | самцы (15) | самки (40) | самцы (20) | ||
| Длина тела | $\frac{{0.60 - 1.01{\text{*}}}}{{0.811 \pm 0.021{\text{**}}}}$ | $\frac{{0.94 - 1.03}}{{0.977 \pm 0.004}}$ | $\frac{{0.61 - 0.78}}{{0.692 \pm 0.011}}$ | $\frac{{0.97 - 1.05}}{{1.012 \pm 0.005}}$ | $\frac{{0.69 - 0.82}}{{0.750 \pm 0.008}}$ | $\frac{{0.99 - 1.88}}{{1.090 \pm 0.022}}$ | $\frac{{0.75 - 0.83}}{{0.796 \pm 0.005}}$ |
| Ширина тела | $\frac{{0.026 - 0.033}}{{0.0294 \pm 0.0003}}$ | $\frac{{0.038 - 0.046}}{{0.0410 \pm 0.0004}}$ | $\frac{{0.028 - 0.036}}{{0.0313 \pm 0.0004}}$ | $\frac{{0.041 - 0.049}}{{0.0430 \pm 0.0004}}$ | $\frac{{0.028 - 0.037}}{{0.0315 \pm 0.0005}}$ | $\frac{{0.043 - 0.062}}{{0.0512 \pm 0.0010}}$ | $\frac{{0.035 - 0.043}}{{0.0394 \pm 0.0005}}$ |
| Длина стомы | $\frac{{0.010 - 0.110}}{{0.0132 \pm 0.0020}}$ | $\frac{{0.012 - 0.015}}{{0.0132 \pm 0.0002}}$ | $\frac{{0.011 - 0.013}}{{0.0115 \pm 0.0001}}$ | $\frac{{0.013 - 0.015}}{{0.0138 \pm 0.0002}}$ | $\frac{{0.011 - 0.013}}{{0.0122 \pm 0.0002}}$ | $\frac{{0.014 - 0.018}}{{0.0159 \pm 0.0002}}$ | $\frac{{0.012 - 0.014}}{{0.0131 \pm 0.0002}}$ |
| Длина глотки | $\frac{{0.162 - 0.236}}{{0.203 \pm 0.003}}$ | $\frac{{0.213 - 0.258}}{{0.2314 \pm 0.0021}}$ | $\frac{{0.168 - 0.209}}{{0.1869 \pm 0.0023}}$ | $\frac{{0.238 - 0.273}}{{0.2580 \pm 0.0022}}$ | $\frac{{0.175 - 0.226}}{{0.2016 \pm 0.0031}}$ | $\frac{{0.248 - 0.294}}{{0.2738 \pm 0.0024}}$ | $\frac{{0.200 - 0.244}}{{0.2194 \pm 0.0027}}$ |
| Ширина глотки | $\frac{{0.010 - 0.014}}{{0.0117 \pm 0.0002}}$ | $\frac{{0.013 - 0.016}}{{0.0141 \pm 0.0002}}$ | $\frac{{0.011 - 0.013}}{{0.0116 \pm 0.0001}}$ | $\frac{{0.013 - 0.013}}{{0.0150 \pm 0.0003}}$ | $\frac{{0.011 - 0.013}}{{0.0121 \pm 0.0001}}$ | $\frac{{0.014 - 0.018}}{{0.0160 \pm 0.0002}}$ | $\frac{{0.012 - 0.014}}{{0.0129 \pm 0.0001}}$ |
| Длина бульбуса | $\frac{{0.038 - 0.056}}{{0.0444 \pm 0.0008}}$ | $\frac{{0.041 - 0.074}}{{0.0600 \pm 0.0013}}$ | $\frac{{0.041 - 0.055}}{{0.0480 \pm 0.0009}}$ | $\frac{{0.065 - 0.083}}{{0.0747 \pm 0.0011}}$ | $\frac{{0.048 - 0.066}}{{0.0543 \pm 0.0011}}$ | $\frac{{0.074 - 0.094}}{{0.0844 \pm 0.0010}}$ | $\frac{{0.058 - 0.083}}{{0.0692 \pm 0.0017}}$ |
| Ширина бульбуса | $\frac{{0.019 - 0.028}}{{0.0267 \pm 0.0038}}$ | $\frac{{0.027 - 0.033}}{{0.0289 \pm 0.0003}}$ | $\frac{{0.021 - 0.029}}{{0.0240 \pm 0.0006}}$ | $\frac{{0.027 - 0.036}}{{0.0310 \pm 0.0006}}$ | $\frac{{0.025 - 0.032}}{{0.0288 \pm 0.0004}}$ | $\frac{{0.030 - 0.038}}{{0.0338 \pm 0.0004}}$ | $\frac{{0.028 - 0.034}}{{0.0303 \pm 0.0003}}$ |
| Длина полового зачатка |
$\frac{{0.026 - 0.079}}{{0.0478 \pm 0.0021}}$ | $\frac{{0.067 - 0.116}}{{0.0902 \pm 0.0028}}$ | $\frac{{0.051 - 0.094}}{{0.0724 \pm 0.0028}}$ | $\frac{{0.185 - 0.252}}{{0.2228 \pm 0.0043}}$ | $\frac{{0.113 - 0.150}}{{0.1322 \pm 0.0020}}$ | $\frac{{0.262 - 0.319}}{{0.2939 \pm 0.0023}}$ | $\frac{{0.208 - 0.251}}{{0.2326 \pm 0.0023}}$ |
| Ширина полового зачатка |
$\frac{{0.010 - 0.014}}{{0.0115 \pm 0.0002}}$ | $\frac{{0.013 - 0.016}}{{0.0143 \pm 0.0002}}$ | $\frac{{0.012 - 0.014}}{{0.0127 \pm 0.0002}}$ | $\frac{{0.017 - 0.021}}{{0.0186 \pm 0.0002}}$ | $\frac{{0.014 - 0.017}}{{0.0156 \pm 0.0002}}$ | $\frac{{0.018 - 0.022}}{{0.0200 \pm 0.0002}}$ | $\frac{{0.018 - 0.020}}{{0.0191 \pm 0.0002}}$ |
| Длина рулька | – | – | $\frac{{0.020 - 0.030}}{{0.026 \pm 0.001}}$ | – | $\frac{{0.043 - 0.052}}{{0.048 \pm 0.001}}$ | – | $\frac{{0.062 - 0.069}}{{0.064 \pm 0.001}}$ |
| Длина спикул | – | – | – | – | $\frac{{0.031 - 0.039}}{{0.035 \pm 0.001}}$ | – | $\frac{{0.044 - 0.052}}{{0.049 \pm 0.001}}$ |
| Длина хвоста | $\frac{{0.110 - 0.196}}{{0.1504 \pm 0.0043}}$ | $\frac{{0.17 - 0.209}}{{0.1928 \pm 0.0016}}$ | $\frac{{0.115 - 0.134}}{{0.1270 \pm 0.0010}}$ | $\frac{{0.190 - 0.222}}{{0.2057 \pm 0.0021}}$ | $\frac{{0.120 - 0.141}}{{0.1327 \pm 0.0011}}$ | $\frac{{0.200 - 0.231}}{{0.2160 \pm 0.0016}}$ | $\frac{{0.122 - 0.143}}{{0.134 \pm 0.0014}}$ |
Примечание. Здесь и в табл. 4: над чертой ‒ min‒max, под чертой – среднее и его ошибка, в скобках – число исследованных личинок нематод.
Личинка IV возраста (рис. 3). Рот с тремя губами. Глотка разделена на стому и бульбус. Аркадные клетки отсутствуют. Половой зачаток удлиненный, занимает >1/8 тела. У самок в средней части полового зачатка имеется уплотнение – зачаток вульвы (рис. 3а). У самцов в клоаке просматривается слабо склеротизированный рулек. Спикулы слабо различимы. Задняя часть полового зачатка не соединена с клоакой. Впереди клоаки хорошо видны формирующиеся кутикулярные бляшки, на хвостовом конце становятся заметными сосочки (рис. 3б).
Рис. 3.
Личинка Cosmocerca ornata IV возраста, в конце 2-х сут: а – самка, б – самец. Масштаб: 0.2 мм.

Рис. 4.
Личинки Cosmocerca ornata на стадии линьки IV–adult, 60 ч с момента заражения: а – самка, б – самец. Масштаб: 0.2 мм.

Личинка на стадии линьки IV–adult (рис. 4). Строение переднего конца и глотки соответствует таковому личинок предыдущей стадии. У самок половой зачаток разделен на переднюю, среднюю и заднюю части. Удлинение полового зачатка происходит за счет передней и задней частей (рис. 4а). У самцов хорошо видны пять пар кутикулярных бляшек впереди клоаки и половые сосочки на хвостовом конце. В клоаке хорошо просматриваются склеротизированный рулек. Спикулы плохо видны. Имеется изгиб полового зачатка по направлению к заднему концу тела. Задняя часть полового зачатка соединена с клоакой (рис. 4б). Морфометрические характеристики личинок C. ornata приведены в табл. 2.
Развитие личинок C. ornata в организме хозяина
При исследовании сеголетков в первые сутки с момента посадки в емкости с инвазионными личинками зарегистрировано их 100%-ное заражение живыми личинками C. ornata (табл. 1). Морфологически эти личинки не отличались от свободноживущих личинок III возраста11. К концу первых суток увеличивается ширина личинок, стома укорачивается, глотка удлиняется. Губы личинок слабо различимы. Половой зачаток увеличивается в три раза (рис. 1б). Число и размеры аркадных клеток вокруг глотки (в области нервного кольца) уменьшаются по сравнению с только что поступившими в амфибий (рис. 1а, 1б). Следует отметить, что одни личинки заметно короче других и у них сравнительно короткий хвостовой конец (будущие самцы); другие ‒ значительно длиннее и у них более вытянут хвостовой конец (будущие самки).
Начиная со вторых суток от начала заражения (через 26 ч), в глазах лягушек зафиксирована третья линька личинок (рис. 2а, 2б). К концу вторых суток единично появляются личинки IV возраста (самцы и самки) (табл. 3, рис. 3а, 3б).
Таблица 3.
Экспериментальные данные распределения личинок Cosmocerca ornata III–IV возраста в сеголетках озерных лягушек
| Время от начала заражения, сут | N | III возраст | Линька III в IV | IV возраст | Линька IV в adult | |||
|---|---|---|---|---|---|---|---|---|
| самки | самцы | самки | самцы | самки | самцы | |||
| 1 | 12 | $\frac{{100.0\left( {12} \right)}}{{6.8 \pm 1.5\left( {1 - 17} \right)}}$ | 0 | 0 | 0 | 0 | 0 | 0 |
| 2 | 12 | $\frac{{75.0 \pm 13.1\left( 9 \right)}}{{4.6 \pm 1.0\left( {2 - 10} \right)}}$ | $\frac{{83.3 \pm 11.3\left( {10} \right)}}{{1.6 \pm 0.3\left( {1 - 3} \right)}}$ | $\frac{{33.3 \pm 14.2\left( 4 \right)}}{{0.3 \pm 0.1\left( 1 \right)}}$ | $\frac{{16.7 \pm 11.3\left( 2 \right)}}{{0.2 \pm 0.1\left( 2 \right)}}$ | $\frac{{8.3 \pm 8.3\left( 1 \right)}}{{0.1 \pm 0.1\left( 1 \right)}}$ | 0 | 0 |
| 3 | 12 | $\frac{{33.3 \pm 14.2\left( 4 \right)}}{{0.7 \pm 0.3\left( {1 - 3} \right)}}$ | $\frac{{100.0{\text{ }}\left( {12} \right)}}{{3.4 \pm 0.4\left( {2 - 6} \right)}}$ | $\frac{{58.3 \pm 14.9\left( 7 \right)}}{{0.6 \pm 0.2\left( 1 \right)}}$ | $\frac{{58.3 \pm 14.9\left( 7 \right)}}{{1.4 \pm 0.4\left( {2 - 3} \right)}}$ | $\frac{{16.7 \pm 11.3\left( 2 \right)}}{{0.2 \pm 0.1\left( 1 \right)}}$ | $\frac{{25.0 \pm 13.1\left( 3 \right)}}{{0.3 \pm 0.2\left( {1 - 2} \right)}}$ | $\frac{{8.3 \pm 8.3\left( 1 \right)}}{{0.1 \pm 0.1\left( 1 \right)}}$ |
| 4 | 12 | $\frac{{16.7 \pm 11.3\left( 2 \right)}}{{0.3 \pm 0.2\left( {1 - 2} \right)}}$ | $\frac{{33.3 \pm 14.2\left( 4 \right)}}{{1.1 \pm 0.5\left( {2 - 6} \right)}}$ | $\frac{{33.3 \pm 14.2\left( 4 \right)}}{{0.3 \pm 0.1\left( 1 \right)}}$ | $\frac{{91.7 \pm 8.3\left( {11} \right)}}{{2.6 \pm 0.4\left( {1 - 4} \right)}}$ | $\frac{{50.0 \pm 15.1\left( 6 \right)}}{{0.5 \pm 0.2\left( 1 \right)}}$ | $\frac{{41.7 \pm 14.9\left( 5 \right)}}{{0.8 \pm 0.3\left( {1 - 3} \right)}}$ | $\frac{{16.7 \pm 11.3\left( 2 \right)}}{{0.2 \pm 0.1\left( 1 \right)}}$ |
| 5 | 15 | 0 | $\frac{{13.3 \pm 9.1\left( 2 \right)}}{{0.2 \pm 0.1\left( {1 - 2} \right)}}$ | $\frac{{6.7 \pm 6.7\left( 1 \right)}}{{0.1 \pm 0.1\left( 1 \right)}}$ | $\frac{{60.0 \pm 13.1\left( 9 \right)}}{{1.4 \pm 0.3\left( {1 - 3} \right)}}$ | $\frac{{33.3 \pm 12.6\left( 5 \right)}}{{0.3 \pm 0.1\left( 1 \right)}}$ | $\frac{{80.0 \pm 10.7\left( {12} \right)}}{{1.9 \pm 0.4\left( {1 - 4} \right)}}$ | $\frac{{33.3 \pm 12.6\left( 5 \right)}}{{0.3 \pm 0.1\left( 1 \right)}}$ |
| 7 | 15 | 0 | 0 | 0 | $\frac{{33.3 \pm 12.6\left( 5 \right)}}{{0.4 \pm 0.2\left( {1 - 2} \right)}}$ | $\frac{{6.7 \pm 6.7\left( 1 \right)}}{{0.1 \pm 0.1\left( 1 \right)}}$ | $\frac{{100{\text{ }}\left( {15} \right)}}{{2.3 \pm 0.4\left( {1 - 5} \right)}}$ | $\frac{{46.7 \pm 13.3\left( 7 \right)}}{{0.5 \pm 0.1\left( 1 \right)}}$ |
| 9 | 12 | 0 | 0 | 0 | 0 | 0 | $\frac{{66.7 \pm 14.2\left( 8 \right)}}{{1.5 \pm 0.4\left( {1 - 5} \right)}}$ | $\frac{{25.0 \pm 13.1\left( 3 \right)}}{{0.3 \pm 0.1\left( 1 \right)}}$ |
| 11 | 12 | 0 | 0 | 0 | 0 | 0 | $\frac{{25.0 \pm 13.1\left( 3 \right)}}{{0.3 \pm 0.2\left( {1 - 2} \right)}}$ | $\frac{{8.3 \pm 8.3\left( 1 \right)}}{{0.1 \pm 0.1\left( 1 \right)}}$ |
Примечание. Здесь и в табл. 5: над чертой – количество зараженных паразитами сеголетков, %: в скобках – число сеголетков (экз.), в которых встречены нематоды; под чертой – индекс обилия гельминтов, в скобках – интенсивность инвазии сеголетков.
На третьи сутки заражения встречаемость личинок на стадии третьей линьки достигает в сеголетках 100%, личинки III возраста присутствуют единично в глазах амфибий (табл. 3). Спустя 60 ч с момента заражения начинают встречаться личинки на стадии четвертой линьки (табл. 3). Эти личинки отличаются от личинок предыдущего возраста по форме и размерам полового зачатка (рис. 4а, 4б).
На 4–5-е сутки личинки стадии четвертой линьки обнаружены не только в глазах, но и в ротоглоточной полости и пищеводе лягушек. С этого времени постепенно снижается общий индекс обилия нематод в амфибиях, а в воде, в которой содержались сеголетки, единично отмечаются личинки этой стадии развития (табл. 1, 3). В глазах сеголетков еще присутствуют личинки C. ornata на стадии третьей линьки (табл. 3).
Начиная с 7-х суток, личинки III возраста (линяющие и не линяющие) в глазах сеголетков не обнаружены. В это время в амфибиях встречаемость личинок на стадии четвертой линьки достигает 100% (табл. 3). На 9-е сутки большая часть личинок этой стадии отмечена в ротоглоточной полости и пищеводе амфибий, единично – в глазах лягушек (табл. 3).
На 11-e сутки с момента заражения в глазах сеголетков личинки C. ornata не отмечаются, а в начале пищеварительного тракта лягушек регистрируются единично. На 14-е сут с момента заражения личинки C. ornata в сеголетках не регистрируются (табл. 1).
Описание половозрелых особей C. ornata22
Общая морфология (табл. 4). Вдоль всего тела кутикула с несколькими продольными рядами сосочков. Латеральные крылья начинаются несколько позади переднего конца, простираются вдоль всего тела и заканчиваются позади клоаки, не достигая кончика хвоста. Ротовое отверстие треугольное, рот с тремя губами. Глотка состоит из стомы и бульбуса. Позади бульбуса начинается кишечник. Экскреторное отверстие открывается на уровне или несколько впереди бульбуса. Нервное кольцо располагается примерно в средней части глотки.
Таблица 4.
Морфометрическая характеристика половозрелых самок и самцов C. ornata
| Показатель, мм | Возраст самки | Возраст самца | ||||
|---|---|---|---|---|---|---|
| I | II | III | IV | I | II | |
| Длина тела | $\frac{{1.405 - 3.064}}{{2.157 \pm 0.108}}$ | $\frac{{3.000 - 4.571}}{{3.901 \pm 0.085}}$ | $\frac{{3.800 - 4.990}}{{4.385 \pm 0.080}}$ | $\frac{{4.000 - 5.414}}{{4.644 \pm 0.102}}$ | $\frac{{0.814 - 1.443}}{{1.077 \pm 0.051}}$ | $\frac{{1.170 - 2.210}}{{1.729 \pm 0.071}}$ |
| Ширина тела | $\frac{{0.083 - 0.180}}{{0.127 \pm 0.006}}$ | $\frac{{0.176 - 0.269}}{{0.229 \pm 0.005}}$ | $\frac{{0.213 - 0.276}}{{0.247 \pm 0.005}}$ | $\frac{{0.206 - 0.279}}{{0.239 \pm 0.005}}$ | $\frac{{0.040 - 0.099}}{{0.072 \pm 0.005}}$ | $\frac{{0.100 - 0.186}}{{0.141 \pm 0.005}}$ |
| Длина стомы | $\frac{{0.021 - 0.046}}{{0.032 \pm 0.002}}$ | $\frac{{0.017 - 0.027}}{{0.022 \pm 0.001}}$ | $\frac{{0.021 - 0.028}}{{0.031 \pm 0.001}}$ | $\frac{{0.023 - 0.032}}{{0.027 \pm 0.001}}$ | $\frac{{0.012 - 0.014}}{{0.0130 \pm 0.0002}}$ | $\frac{{0.012 - 0.014}}{{0.0130 \pm 0.0002}}$ |
| Длина глотки | $\frac{{0.204 - 0.444}}{{0.313 \pm 0.016}}$ | $\frac{{0.333 - 0.508}}{{0.429 \pm 0.009}}$ | $\frac{{0.384 - 0.504}}{{0.443 \pm 0.008}}$ | $\frac{{0.377 - 0.510}}{{0.438 \pm 0.010}}$ | $\frac{{0.221 - 0.256}}{{0.240 \pm 0.003}}$ | $\frac{{0.244 - 0.331}}{{0.289 \pm 0.006}}$ |
| Ширина глотки | $\frac{{0.022 - 0.049}}{{0.034 \pm 0.002}}$ | $\frac{{0.031 - 0.047}}{{0.040 \pm 0.001}}$ | $\frac{{0.039 - 0.051}}{{0.046 \pm 0.001}}$ | $\frac{{0.041 - 0.056}}{{0.049 \pm 0.001}}$ | $\frac{{0.013 - 0.016}}{{0.0150 \pm 0.0003}}$ | $\frac{{0.016 - 0.021}}{{0.0180 \pm 0.0004}}$ |
| Длина бульбуса | $\frac{{0.070 - 0.153}}{{0.108 \pm 0.005}}$ | $\frac{{0.104 - 0.148}}{{0.134 \pm 0.003}}$ | $\frac{{0.135 - 0.177}}{{0.155 \pm 0.003}}$ | $\frac{{0.132 - 0.179}}{{0.154 \pm 0.003}}$ | $\frac{{0.077 - 0.100}}{{0.087 \pm 0.002}}$ | $\frac{{0.089 - 0.115}}{{0.092 \pm 0.006}}$ |
| Ширина бульбуса | $\frac{{0.047 - 0.102}}{{0.072 \pm 0.004}}$ | $\frac{{0.074 - 0.112}}{{0.096 \pm 0.002}}$ | $\frac{{0.079 - 0.104}}{{0.108 \pm 0.002}}$ | $\frac{{0.092 - 0.124}}{{0.104 \pm 0.002}}$ | $\frac{{0.031 - 0.048}}{{0.039 \pm 0.001}}$ | $\frac{{0.050 - 0.063}}{{0.057 \pm 0.001}}$ |
| Длина вагины | $\frac{{0.040 - 0.054}}{{0.044 \pm 0.001}}$ | $\frac{{0.083 - 0.127}}{{0.108 \pm 0.002}}$ | $\frac{{0.144 - 0.189}}{{0.166 \pm 0.003}}$ | $\frac{{0.178 - 0.241}}{{0.206 \pm 0.005}}$ | – | – |
| Длина матки | $\frac{{0.142 - 0.388}}{{0.267 \pm 0.015}}$ | $\frac{{0.600 - 0.914}}{{0.775 \pm 0.018}}$ | $\frac{{1.218 - 1.599}}{{1.406 \pm 0.026}}$ | $\frac{{1.316 - 1.781}}{{1.528 \pm 0.033}}$ | – | – |
| Длина яичников | $\frac{{0.400 - 1.091}}{{0.752 \pm 0.042}}$ | $\frac{{1.818 - 2.770}}{{2.364 \pm 0.051}}$ | $\frac{{2.715 - 3.564}}{{3.132 \pm 0.057}}$ | $\frac{{2.740 - 3.708}}{{3.182 \pm 0.070}}$ | – | – |
| Длина яйцеводов | $\frac{{0.088 - 0.266}}{{0.182 \pm 0.010}}$ | $\frac{{0.652 - 0.994}}{{0.848 \pm 0.019}}$ | $\frac{{0.826 - 1.085}}{{0.954 \pm 0.018}}$ | $\frac{{0.952 - 1.289}}{{1.106 \pm 0.024}}$ | – | – |
| Длина семяприемников | $\frac{{0.038 - 0.089}}{{0.062 \pm 0.003}}$ | $\frac{{0.204 - 0.311}}{{0.264 \pm 0.006}}$ | $\frac{{0.258 - 0.339}}{{0.298 \pm 0.006}}$ | $\frac{{0.238 - 0.322}}{{0.279 \pm 0.006}}$ | – | – |
| Ширина семяприемников | $\frac{{0.017 - 0.040}}{{0.027 \pm 0.001}}$ | $\frac{{0.091 - 0.138}}{{0.117 \pm 0.003}}$ | $\frac{{0.110 - 0.144}}{{0.127 \pm 0.002}}$ | $\frac{{0.080 - 0.107}}{{0.093 \pm 0.002}}$ | – | – |
| Расстояние от вульвы до ануса | $\frac{{0.468 - 1.021}}{{0.719 \pm 0.036}}$ | $\frac{{0.923 - 1.406}}{{1.201 \pm 0.026}}$ | $\frac{{1.545 - 2.028}}{{1.786 \pm 0.033}}$ | $\frac{{1.695 - 2.294}}{{1.968 \pm 0.043}}$ | – | – |
| Длина яиц | – | – | $\frac{{0.098 - 0.130}}{{0.114 \pm 0.002}}$ | – | – | – |
| Ширина яиц | – | – | $\frac{{0.063 - 0.084}}{{0.074 \pm 0.001}}$ | – | – | – |
| Длина рулька | – | – | – | – | $\frac{{0.063 - 0.079}}{{0.069 \pm 0.001}}$ | $\frac{{0.075 - 0.086}}{{0.081 \pm 0.001}}$ |
| Длина спикул | – | – | – | – | $\frac{{0.051 - 0.062}}{{0.057 \pm 0.001}}$ | $\frac{{0.055 - 0.069}}{{0.061 \pm 0.001}}$ |
| Длина хвоста | $\frac{{0.227 - 0.494}}{{0.350 \pm 0.018}}$ | $\frac{{0.352 - 0.537}}{{0.454 \pm 0.011}}$ | $\frac{{0.302 - 0.396}}{{0.337 \pm 0.014}}$ | $\frac{{0.339 - 0.459}}{{0.394 \pm 0.009}}$ | $\frac{{0.129 - 0.148}}{{0.138 \pm 0.002}}$ | $\frac{{0.151 - 0.202}}{{0.182 \pm 0.003}}$ |
Самец. Хвост всегда загнут вентрально, с игловидным отростком на конце. На вентральной и латеральной сторонах хвоста имеются сосочки. Впереди отверстия клоаки расположены в два продольных параллельных ряда 10 (по пять в каждом ряду) кутикулярных образований – бляшек, несколько смещенных по отношению друг к другу. От центра каждой бляшки кзади направлены шесть–восемь коротких выростов. Половая система самца представлена семенником, семяпроводом, семенным пузырьком, семяизвергательным каналом, рульком и спикулами. Рулек треугольной формы сильно склеротизирован; с расширенной проксимальной частью и конической дистальной. Спикулы рудиментарные, плохо просматриваются (рис. 5б).
Рис. 5.
Половозрелые Cosmocerca ornata, 5-е сут с момента заражения: а – самка, б – самец. Масштаб: 0.2 мм.

Самка. Размеры тела больше, чем у самцов. Половая система продельфная; включает парные яичник, яйцевод, семяприемник, матку и вагину с вульвой. Яичники располагаются в передней части тела, образуют несколько петель и извивов и не доходят до основания пищевода. Яйцеводы с округлыми семяприемниками, один из которых лежит в средней части тела, выше уровня вульвы; другой – в задней части. Передняя матка располагается на уровне вульвы и несколько выше нее; задняя матка простирается в заднюю часть тела позади уровня вульвы. В дистальной части обе матки сужаются и сливаются позади отверстия вульвы в короткий общий канал, который переходит в мышечную вагину. Вульва открывается примерно на уровне середины тела. Хвост шиловидно заостренный. На дистальной части хвоста имеются две пары рудиментарных шипиков (рис. 5а, рис. 6).

Развитие половозрелых особей C. ornata
К концу 5-х сут с момента заражения в последней трети кишечника (место постоянной локализации C. ornata) единично регистрировали половозрелых нематод I возрастной группы (самки и самцы) (табл. 5, рис. 5). Морфологически они отличаются от личинок последнего возраста наличием сформированных наружных половых органов (у самцов ‒ спикул, у самок ‒ вульвы) и развитой половой системой. Следует отметить, что самцы C. ornata, если они встречались в сеголетках, то только по одному в хозяине. Как и на III–IV личиночных стадиях – одновременно два и более самца нематод в одном сеголетке не были отмечены.
Таблица 5.
Распределение самок и самцов Cosmocerca ornata отдельных возрастных групп в сеголетках озерных лягушек
| Время заражения, cут | N | Возраст самки | Возраст самца | ||||
|---|---|---|---|---|---|---|---|
| I | II | III | IV | I | II | ||
| 5 | 15 | $\frac{{13.3 \pm 9.1\left( 2 \right)}}{{0.1 \pm 0.1\left( 1 \right)}}$ | 0 | 0 | 0 | $\frac{{6.7 \pm 6.7\left( 1 \right)}}{{0.1 \pm 0.1\left( 1 \right)}}$ | 0 |
| 7 | 15 | $\frac{{53.3 \pm 13.3\left( 8 \right)}}{{0.9 \pm 0.2\left( {1 - 2} \right)}}$ | 0 | 0 | 0 | $\frac{{33.3 \pm 12.6\left( 5 \right)}}{{0.3 \pm 0.1\left( 1 \right)}}$ | 0 |
| 9 | 12 | $\frac{{83.3 \pm 11.3\left( {10} \right)}}{{1.8 \pm 0.4\left( {1 - 5} \right)}}$ | 0 | 0 | 0 | $\frac{{75.0 \pm 13.1\left( 9 \right)}}{{0.8 \pm 0.2\left( 1 \right)}}$ | 0 |
| 11 | 12 | $\frac{{100.0\left( {12} \right)}}{{2.8 \pm 0.3\left( {1 - 4} \right)}}$ | 0 | 0 | 0 | $\frac{{66.7 \pm 14.2\left( 8 \right)}}{{0.7 \pm 0.2\left( 1 \right)}}$ | $\frac{{25.0 \pm 13.1\left( 3 \right)}}{{0.3 \pm 0.1\left( 1 \right)}}$ |
| 14 | 15 | $\frac{{100.0\left( {15} \right)}}{{2.5 \pm 0.3\left( {1 - 5} \right)}}$ | $\frac{{20.0 \pm 10.7\left( 3 \right)}}{{0.3 \pm 0.2\left( {1 - 2} \right)}}$ | 0 | 0 | $\frac{{33.3 \pm 12.6\left( 5 \right)}}{{0.3 \pm 0.1\left( 1 \right)}}$ | $\frac{{53.3 \pm 13.3{\text{ }}\left( 8 \right)}}{{0.5 \pm 0.2\left( 1 \right)}}$ |
| 17 | 15 | $\frac{{66.7 \pm 12.6\left( {10} \right)}}{{1.5 \pm 0.3\left( {1 - 4} \right)}}$ | $\frac{{4.6.7 \pm 13.3\left( 7 \right)}}{{1.1 \pm 0.4\left( {1 - 4} \right)}}$ | 0 | 0 | 0 | $\frac{{66.7 \pm 12.6\left( {10} \right)}}{{0.7 \pm 0.2\left( 1 \right)}}$ |
| 20 | 15 | $\frac{{33.3 \pm 12.6\left( 5 \right)}}{{0.6 \pm 0.3\left( {1 - 3} \right)}}$ | $\frac{{73.3 \pm 11.8\left( {11} \right)}}{{1.9 \pm 0.5\left( {1 - 6} \right)}}$ | 0 | 0 | 0 | $\frac{{33.3 \pm 12.6\left( 5 \right)}}{{0.3 \pm 0.1\left( 1 \right)}}$ |
| 23 | 12 | 0 | $\frac{{100.0\left( {12} \right)}}{{2.8 \pm 0.4\left( {1 - 5} \right)}}$ | $\frac{{8.3 \pm 8.3\left( 1 \right)}}{{0.1 \pm 0.1\left( 1 \right)}}$ | 0 | 0 | 0 |
| 26 | 12 | 0 | $\frac{{91.7 \pm 8.3\left( {11} \right)}}{{2.6 \pm 0.4\left( {1 - 5} \right)}}$ | $\frac{{25.0 \pm 13.1\left( 3 \right)}}{{0.4 \pm 0.2\left( {1 - 2} \right)}}$ | 0 | 0 | 0 |
| 29 | 12 | 0 | $\frac{{58.3 \pm 14.9\left( 7 \right)}}{{1.2 \pm 0.4\left( {1 - 3} \right)}}$ | $\frac{{83.3 \pm 11.3\left( {10} \right)}}{{1.4 \pm 0.3\left( {1 - 3} \right)}}$ | 0 | 0 | 0 |
| 33 | 15 | 0 | $\frac{{26.7 \pm 11.8\left( 3 \right)}}{{0.3 \pm 0.2\left( {1 - 2} \right)}}$ | $\frac{{100.0\left( {15} \right)}}{{1.9 \pm 0.2\left( {1 - 3} \right)}}$ | $\frac{{13.3 \pm 9.1\left( 2 \right)}}{{0.1 \pm 0.1\left( 1 \right)}}$ | 0 | 0 |
| 36 | 15 | 0 | 0 | $\frac{{100.0\left( {15} \right)}}{{1.5 \pm 0.1\left( {1 - 2} \right)}}$ | $\frac{{40.0 \pm 13.1\left( 6 \right)}}{{0.7 \pm 0.2\left( {1 - 2} \right)}}$ | 0 | 0 |
| 38 | 12 | 0 | 0 | $\frac{{50.0 \pm 15.1\left( 6 \right)}}{{0.7 \pm 0.2\left( {1 - 2} \right)}}$ | $\frac{{83.3 \pm 11.3\left( {10} \right)}}{{1.4 \pm 0.3\left( {1 - 4} \right)}}$ | 0 | 0 |
| 40 | 15 | 0 | 0 | $\frac{{20.0 \pm 10.7\left( 3 \right)}}{{0.2 \pm 0.1\left( 1 \right)}}$ | $\frac{{60.9 \pm 13.1\left( 7 \right)}}{{0.9 \pm 0.2\left( {1 - 2} \right)}}$ | 0 | 0 |
| 42 | 15 | 0 | 0 | 0 | $\frac{{33.3 \pm 12.6\left( 5 \right)}}{{0.5 \pm 0.2\left( {1 - 2} \right)}}$ | 0 | 0 |
| 45 | 15 | 0 | 0 | 0 | 0 | 0 | 0 |
На 7‒9-е сутки встречаемость половозрелых самок и самцов I возрастной группы в кишечнике лягушек возрастала (табл. 5, рис. 6а, 7а). На 11-е сут с момента заражения в кишечнике амфибий были отмечены половозрелые самки нематод только I возрастной группы. Их встречаемость достигла 100%, среди самцов C. ornata появились особи II возрастной группы (табл. 5, рис. 7б).
Рис. 7.
Самцы Cosmocerca ornata разного возраста (по: Кириллов, Кириллова, 2016а): а – I возрастная группа, б – II возрастная группа. Масштаб: 0.2 мм.
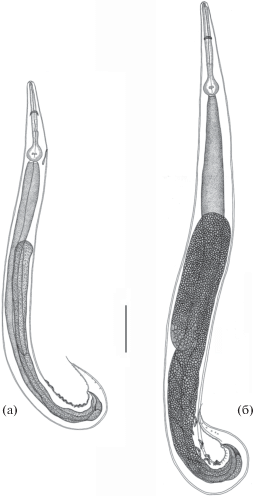
На 14-е сут встречаемость самок I возрастной группы в лягушках оставалась максимальной. С этого времени появились самки нематод, в матке которых формировались яйца (II возрастная группа); количество самцов II возрастной группы увеличилось (табл. 5, рис. 6б, рис. 7б).
С 17-х по 20-е сут эксперимента все обнаруженные в сеголетках самцы C. ornata относились к II возрастной группе; самки нематод представлены I и II возрастными группами.
С 17-х сут общая численность самцов в амфибиях снижалась, было отмечено их появление в воде (табл. 5).
На 23-и сут в кишечнике лягушек впервые была обнаружена самка C. ornata III возрастной группы, в яйцах матки отмечено формирование личинок (рис. 6в). Встречаемость самок II группы достигала 100%, самки I возрастной группы и самцы отсутствовали (табл. 5).
С этого времени и до 33-х сут в сеголетках возрастала встречаемость самок III возрастной группы с одновременным понижением доли самок II группы. На 33-и сут в кишечнике амфибий впервые зафиксированы самки с пустой растянутой маткой (IV возрастная группа) (табл. 5, рис. 6г), в воде, содержавшей сеголетков, отмечены личинки C. ornata.
На 36-е сут в сеголетках отсутствовали самки II возрастной группы; встречаемость самок III группы по-прежнему была 100%, доля самок IV возрастной группы увеличивалась (табл. 5).
На 38-е сут встречаемость в амфибиях самок C. ornata IV возрастной группы достигала максимума, доля самок III группы снизилась (табл. 5), появились сеголетки, свободные от паразитов.
На 40-е сут самки III возрастной группы в кишечнике лягушек встречались единично, а с 42-х сут уже не обнаруживались (табл. 5). В этот период встречаемость самок IV возрастной группы снизилась, число сеголетков, освободившихся от гельминтов, возросло.
На 45-е сут эксперимента нематоды C. ornata в лягушках отсутствовали (табл. 1, 5).
Влияние температуры окружающей среды на развитие нематод в амфибиях
При среднесуточной температуре воздуха лаборатории 27‒29°С третья линька личинок нематод была на 2-е–3-и сут, четвертая линька ‒ на 3‑и‒5-е сут (рис. 8а, 8б); период развития C. ornata в хозяине до половозрелого состояния и начала отрождения личинок самками нематод достигал 33‒36 сут (рис. 8в–8е). При 24‒26°С третью линьку личинок наблюдали на 3- и‒5-е сут, четвертую ‒ на 5‒9-е (рис. 8а, 8б); период развития нематод в хозяине занимал 38‒40 сут (рис. 8в–8е). При среднесуточной температуре 23°С на 4–5-е сут в глазах сеголетков еще встречались личинки III возраста, а третья линька отмечена только на 6–7-е сут. При этой температуре личинки четвертого возраста встречались в глазах лягушек на 9–10-е сут, четвертая линька наблюдалась на 12–13-е сут. Развитие нематод в сеголетках при 23°С продолжалось ~2 мес. Подобная тенденция была отмечена и для взрослых нематод в кишечнике сеголетков (рис. 8в–8е). Дальнейшее понижение температуры приводило к замедлению процессов развития личинок в глазах сеголетков и половозрелых паразитов в кишечнике лягушек.
Рис. 8.
Влияние среднесуточной температуры воздуха лаборатории на сроки развития отдельных стадий Cosmocerca ornata в амфибиях: а ‒ Третья линька личинок нематод, б ‒ Четвертая линька личинок нематод, в ‒ I возрастная группа самок нематод, г ‒ II возрастная группа самок нематод, д ‒ III возрастная группы самок нематод, е ‒ IV возрастная группа самок нематод.

При температуре ≤15°С развитие личинок в глазах и взрослых нематод в кишечнике амфибий не происходило, как и проникновение инвазионных личинок в глаза амфибий из воды. Заражение сеголетков инвазионными личинками нематод начиналось при среднесуточной температуре лаборатории 16‒18°С зарегистрированы единичные личинки в глазах амфибий (рис. 9). С повышением температуры поступление личинок в глаза лягушек увеличивалось. Для выявления роли температуры на процесс заражения амфибий инвазионными личинками C. ornata проведен статистический анализ инвазии лягушек личинками нематод при среднесуточной температуре от 15 до 29°С по критерию Краскела–Уоллиса, который показал рост поступления личинок в глаза сеголетков при повышении температуры (H = 268.1, p < 0.001).
Рис. 9.
Влияние среднесуточной температуры воздуха лаборатории на поступление инвазионных личинок Cosmocerca ornata в амфибий (за 1 ч).

Сравнение развития нематод и их личинок в сеголетках при разных среднесуточных температурах в лаборатории по критерию Краскела–Уоллиса также выявило зависимость сроков развития паразитов отдельных возрастных групп от температуры. Период развития личинок и половозрелых нематод в организме хозяина отрицательно коррелировал со среднесуточной температурой опыта лаборатории (рис. 8). Так, с повышением температуры воздуха в лаборатории отмечалось сокращение сроков наступления третьей (H = 140.3, rs = –0.862) и четвертой (H = 180.3, rs = –0.809) линек личинок C. ornata в глазах амфибий (рис. 8а, 8б). Чем выше была температура, тем раньше в кишечнике сеголетков появлялись половозрелые нематоды I (H = 219.8, rs = –0.755) и II (H = 101.0, rs = –0.768) возрастных групп (рис. 8в, 8г). Также с повышением температуры самки C. ornata в кишечнике амфибий быстрее достигали III и IV возрастов – H = 90.7, rs = –0.790 и H = 72.8, rs = –0.911 соответственно (рис. 8д, 8е). Различия в сроках развития нематод отдельных возрастных групп в сеголетках при разной температуре достоверны (p < 0.001).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Результаты экспериментального заражения сеголетков озерных лягушек показали, что заражение амфибий C. ornata происходит в воде путем проникновения инвазионных личинок III возраста через глаза на коньюктиву под нижнее веко, где личинки в секрете Гардеровой железы претерпевают третью и четвертую линьки. По-видимому, секрет служит своеобразным триггером, вызывающим начало линьки личинок. Подтверждением этого могут служить отрицательные результаты по приживаемости инвазионных личинок C. ornata, полученные при пероральном, перназальном и перанальном их введении в сеголетков. Личинки не претерпевали дальнейшего развития, а элиминировали из организма амфибий.
Проникновение личинок нематод через кожу амфибий не может происходить, что связано со строением ротового аппарата личинок C. ornata, который не приспособлен для перкутанного проникновения.
На стадии четвертой линьки паразиты мигрируют в ротоглоточную полость, вероятно, через носослезный канал и хоаны. Миграции личинок способствует способность глазных яблок амфибий вдвигаться внутрь ротовой полости при глотании пищи (Жданова, 2004). Линяющие личинки обнаруживаются в ротоглоточной полости и пищеводе амфибий и далее следуют к месту постоянной локализации (последней трети кишечника) по желудочно-кишечному тракту.
Войткова с соавт. (Vojtkova et al., 1972; Vojtkova, Moravec, 1973) при гельминтологическом исследовании амфибий Чехословакии обнаружили в соединительной ткани вокруг глаз личинок нематод, которых отнесли к виду C. ornata. Кроме того, авторы отнесли к данному виду личинок нематод, обнаруженных ими в легких, а также в цистах на поверхности пищеварительного тракта и почек у нескольких видов земноводных. В наших исследованиях (в экспериментальных и естественных условиях) личинки C. ornata кроме коньюктивы глаз (личинки III–IV возраста), ротоглоточной полости и пищевода (личинки в стадии линьки IV–adult) нигде не были зарегистрированы.
В.С. Магуза (1973) и И.В. Чихляев (2004) обнаружили у амфибий в тканях вокруг глаз личинок C. ornata, которых ошибочно определили как C. benyulensis Chabaud et Campana-Rouget, 1955 и Desmidocercella numidica Seurat, 1920 соответственно.
Проведенное исследование позволило выявить сроки развития отдельных стадий и общую продолжительность жизни C. ornata в хозяине, оценить влияние температуры на эти процессы. Оптимальная температура для поступления паразита и его развития в хозяине – 24‒29°С. Ранее было установлено, что поступление новых генераций C. ornata в амфибий весной в естественных условиях начинается, когда водоем прогревается до 16–17°С (как и отрождение личинок самками нематод). Показано достоверное влияние температуры воды на эти процессы. Максимальные показатели инвазии озерной лягушки нематодой C. ornata отмечены в летние месяцы, когда температура воды наиболее высока (Кириллов, Кириллова, 2016).
При оптимальной температуре (24‒29°С) нематодам для достижения половой зрелости необходимо ~1 нед; для достижения состояния II возрастной группы самкам C. ornata – ~2 нед, самцам ‒ <2 нед, для развития личинок в яйцах в матке самки и началу их отрождения – ~5 нед. Таким образом, продолжительность жизни самок C. ornata в организме хозяина достигает 45 сут, самцов нематод ‒ 14–23 сут, что подтверждает полученные данные в естественных условиях. Так, анализ ежемесячной динамики возрастной структуры гемипопуляции самок C. ornata в водоеме показал, что продолжительность их жизни в озерных лягушках – >1 мес (Кириллов, Кириллова, 2016а, 2016б).
Ранее установлено, что свободноживущие личинки достигают инвазионности (III возраст) за пять–шесть суток при температуре 24–28°С (Кириллова, Кириллов, 2015). Следовательно, весь цикл развития C. ornata, включающий развитие личинок I‒III возрастов в воде, и паразитов в организме хозяина (личинок III, IV возрастов и адультных нематод), проходит за ~50 сут. Проведенные лабораторные исследования показали, что продолжительность жизненного цикла зависит от температуры окружающей среды: при понижении температуры время нахождения нематод в хозяине увеличивается, при повышении – уменьшается.
Анализ численности личинок, поступивших в амфибий и достигших половозрелости нематод, выявил, что не все паразиты приживаются в лягушках (достигают места постоянной локализации). Часть мигрирующих личинок на стадии четвертой линьки элиминировали из организма хозяина (табл. 1). По-видимому, это связано со случайным вымыванием личинок нематод в воде (в процессе питания) при их миграции через ротоглоточную полость в пищевод. На 1–2-е сут эксперимента индекс обилия составлял 6.8 (табл. 1). Начиная с 4-х сут, когда начиналась миграция личинок через ротоглоточную полость, индекс обилия снижался до 5.8. На 14-е сут (индекс обилия 3.6) все нематоды достигли задней трети кишечника – места постоянной локализации в хозяине (табл. 1). Таким образом, места постоянной локализации достигли ~50% нематод.
Снижение индекса обилия с 14-х сут было обусловлено началом элиминации самцов C. ornata, которая продолжалась до 23-х сут, когда самцы нематод уже не обнаруживались в сеголетках. Снижение индекса обилия с 29–33-х сут было связано с элиминацией самок IV возраста, которая продолжалась до 45-х сут (нематоды уже не регистрировались в амфибиях) (табл. 1, 5).
Полученные экспериментальные данные подтверждают сделанное ранее предположение, что для C. ornata характерна только одна копуляция, после которой самец и самка больше не встречаются (Кириллов, Кириллова, 2016). В лабораторном эксперименте самцы C. ornata встречаются в сеголетках озерных лягушек во время присутствия самок I и II возрастов. После оплодотворения самок самцы элиминируют. Причем, самцы нематод, если и присутствуют в хозяине, то только по одному, как и в озерных лягушках из естественных условий (Кириллов, Кириллова, 2016а, 2016б).
Выводы. Результаты ранее проведенного и настоящего экспериментальных исследований по заражению озерной лягушки Pеlophylax ridibundus инвазионными личинками нематод позволили изучить все стадии и общую продолжительность жизненного цикла C. ornata. Развитие личинок паразита I–III возрастов и заражение амфибий инвазионными личинками (III возраст) происходит в водной среде. Личинки C. ornata в приповерхностном слое воды проникают в глаза амфибий (на конъюктиву нижнего века), где претерпевают третью и четвертую линьки. На стадии четвертой линьки нематоды мигрируют в ротоглоточную полость и далее по пищеварительному тракту к месту постоянной локализации в последней трети кишечника. Места постоянной локализации достигает только ~50% нематод, поступивших в хозяев. Выявлена зависимость поступления личинок в амфибий, продолжительности развития нематод во внешней среде и в организме хозяина от температуры окружающей среды.
Список литературы
Жданова Т.Д. 2004. Особенности строения и поведения земноводных. Москва: Мир.
Кириллова Н.Ю., Кириллов А.А. 2015. Роль головастиков озерных лягушек в реализации жизненного цикла Cosmocerca ornata (Nematoda: Cosmocercidae) // Паразитология. Т. 49. Вып. 1. С. 49.
Кириллов А.А., Кириллова Н.Ю. 2016а. Влияние зимовки озерной лягушки на репродуктивную структуру гемипопуляции Cosmocerca ornata (Nematoda, Cosmocercidae) // Паразитология. Т. 50. Вып. 1. С. 21.
Кириллов А.А., Кириллова Н.Ю. 2016б. Анализ репродуктивной структуры гемипопуляции Cosmocerca ornata (Dujardin, 1845) (Nematoda: Cosmocercidae) в озерных лягушках разного возраста // Биол. внутр. вод. №. 3 С. 93. https://doi.org/10.1134/S199508291603007X
Магуза В.С. 1973. Гельминты амфибий Полесья Украины: Автореф. дис. … канд. биол. наук. Киев.
Чихляев И.В. 2004. Гельминты земноводных (Amphibia) Среднего Поволжья (фауна, экология): Автореф. дис. … канд. биол. наук. Москва.
Шульц Р.С., Гвоздев Е.В. 1970. Основы общей гельминтологии. Т. 1. Москва: Наука.
Anderson R.C. 2000. Nematode Parasites of Vertebrates: Their Development and Transmission. Wallingford: CABI Publishing.
Dusen S., Ugurtas I.H., Aydogdu A. 2010. Nematode parasites of the two limbless lizards: Turkish worm lizards, Blanus strauchi (Bedriaga, 1884) (Squamata: Amphisbaenidae), and slow worm, Anguis fragilis Linnaeus, 1758 (Squamata: Anguidae), from Turkey // Helminthologia. V. 47. P. 158.
Vojtkova L., Moravec F., Krivanec K. 1972. Prispevek k poznani larvalnich stadii hlistic (Nematoda) z obojzivelniku v CSSR // Folia Fac. Sci. Nat. UJEP Brunensis. Biol. 36. Op. 7. Helminthol. sb. № 1. P. 79.
Vojtkova L., Moravec F. 1973. Die nematodenlarven (Nematoda) aus den amphibian in der Tschechoslowakei // Scripta Fac. Sci. Nat. UJEP Brunensis. Biol. 2(3). P. 81.
Yildirimhan H.S., Bursey C.R., Goldberg S.R. 2009. Helminth parasites of the Caucasian parsley frog, Pelodytes caucasicus, from Turkey // Comp. Parasitol. V. 76(2). P. 247. https://doi.org/10.1654/4376.1
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод