

Биология внутренних вод, 2021, № 5, стр. 439-448
Мускулатура тела и особенности фиксации Deropristis hispida (Trematoda, Deropristidae)
М. В. Ястребов a, *, И. В. Ястребова a, Д. И. Смирнова a
a Ярославский государственный университет им. П.Г. Демидова
Ярославль, Россия
* E-mail: mvy@uniyar.ac.ru
Поступила в редакцию 18.01.2021
После доработки 02.02.2021
Принята к публикации 12.02.2021
Аннотация
Описаны внешнее строение, покровы и паренхимная мускулатура Deropristis hispida. Локомоторный отдел тела представляет собой комплекс прикрепительных структур, включающий присоски, дорсальный горб, вентральную впадину сложной конфигурации и мощное вооружение, наиболее крупное на дорсальном горбе и на боковых сторонах тела. Мышечные слои покровов тонкие, диагональный слой частично редуцирован. Паренхимная мускулатура представлена дорсо-вентральными, хордальными и продольными мышцами, развитыми в основном в локомоторном отделе тела, а также немногочисленными волокнами, соединяющими дорсальную и латеральные стороны ротовой присоски с покровами. В передней трети полового отдела тела терминальные части половых протоков окружены редковолокнистым мышечным плексусом из хордальных и дорсо-вентральных мышц. Предполагается, что мелкие присоски не играют доминирующей роли в фиксации D. hispida. Их относительная слабость компенсируется прикреплением с помощью вооружения и вентральной впадины. Гельминт сохраняет локализацию в большей степени прикреплением, чем передвижением.
ВВЕДЕНИЕ
Большинство видов кишечных трематод относится к реофильным гельминтам, поскольку они живут в потоке веществ, находясь под угрозой утраты локализации. Обычно они сохраняют локализацию с помощью активной фиксации, которая требует мышечных усилий. Усилия расходуются на прикрепление и передвижение, используемые в разных пропорциях (Ошмарин, 1959, 1960). Различные способы сохранения локализации отчетливо проявляются во внешнем строении трематод. Самый распространенный способ – дифференцировка тела на локомоторный и половой отделы, в первом из которых сконцентрированы структуры, обеспечивающие прикрепление и передвижение гельминтов (Oshmarin, 1958). Данная эволюционно первичная дифференцировка существует в огромном разнообразии вариантов, отличающихся друг от друга относительным размером и формой отделов тела, а также набором, топографией и строением органов прикрепления. При этом крайне мало известно об одном их ключевых элементов дифференцировки тела трематод – архитектонике соматической мускулатуры.
Deropristis hispida отличается как резко выраженной дифференцировкой тела, так и весьма своеобразным строением его локомоторного отдела. Упомянутое своеобразие представляет интерес возможной архаичностью. Семейство Deropristidae включает всего три рода и является уникальной древней группой паразитов пищеварительного тракта осетровых рыб, эволюционно связанной со своими хозяевами (Choudhury, Dick, 1998). Как и осетровые, семейство находится под угрозой исчезновения, и потому требует особого внимания.
Цель настоящей работы – изучить пространственное распределение мышечных элементов в теле трематоды Deropristis hispida (Abildgaard in Rudolphi, 1819), паразитирующей в пищеводе и кишечнике русского и атлантического осетров, севрюги, стерляди, шипа и белуги (Скрябина, 1974).
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Материалом послужили мариты трематоды Deropristis hispida из кишечника севрюги (Acipenser stellatus Pallas, 1771), добытой в дельте р. Волги в 1970 г. Для изучения взяты половозрелые экземпляры, фиксированные 75%-ным этанолом. Тотальные препараты окрашены уксуснокислым кармином. Анатомия марит изучена под световым микроскопом Axioscop 40L (Carl Zeiss, Германия) по полным сериям парафиновых срезов толщиной 7 мкм, изготовленных по стандартной методике и окрашенных методом Маллори. Было изучено три серии срезов в сагиттальной проекции и пять – в поперечной. Рисунки выполнены с помощью рисовального аппарата РА–4 (“Прогресс”, СССР), промеры – с использованием программы анализа изображений AxioVision 4.8.2 (Carl Zeiss, Германия).
Размерные показатели частей тела приведены в миллиметрах, аналогичные показатели мышц и шипов – в микрометрах. Для присосок указаны длина по оси симметрии и диаметр, для шипов – длина и ширина в основании. При измерении диаметров волокон в мышечных слоях первый диаметр (вертикальный) перпендикулярен базальной пластинке, вдоль которой идет волокно, второй (горизонтальный) параллелен ей. Если первый диаметр больше второго, форма поперечного сечения мышечного элемента именуется овальной, если наоборот – поперечно-овальной, при равенстве диаметров – округлой. Расстояние между соседними мышцами слоя приводится в относительных величинах – в их горизонтальных диаметрах. По вертикальным диаметрам оценивается толщина слоя. Для мышечных элементов, расположенных неперпендикулярно плоскости среза, указана толщина, измеренная в одной плоскости.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Deropristis hispida – трематоды с удлиненным телом (у изученных экземпляров 8–9 мм), дифференцированным на локомоторный и половой отделы (рис. 1). Первый из них длиной 1–1.4 мм заканчивается у заднего края брюшной присоски. Часть данного отдела впереди брюшной присоски у фиксированных особей нередко загнута на спинную сторону почти под прямым углом. Обе присоски мелкие: ротовая 0.10–0.11 × 0.07–0.09, брюшная 0.1–0.11 × 0.13–0.14 мм. Устье ротовой присоски субтерминальное, брюшной – вентральное. Тело между присосками образует неглубокую вентральную впадину сложной конфигурации (рис. 1в). Передняя часть впадины на узкой головной части тела имеет вид продольной щели, начинающейся позади ротовой присоски. В районе центральной части впадины тело обретает форму диска шириной 0.41–0.42 мм, что почти в полтора раза больше ширины полового отдела тела. На данном участке тело несет невысокий дорсальный горб в форме усеченного конуса (рис. 1б). Задняя часть впадины в виде широкого канала доходит до переднего края брюшной присоски.
Рис. 1.
Deropristis hispida: а – общий вид, б, в – немасштабированные схемы строения локомоторного отдела тела со спинной (б) и брюшной (в) сторон. 1 – дорсальный горб, 2 – центральная часть вентральной впадины.

Покровы тела тонкие (рис. 2). В наружной пластинке тегумента находятся элементы вооружения, расположенные в шахматном порядке. Их форма и размер варьируют на разных участках тела. На уровне ротовой присоски шипы мелкие, игольчатые, слегка загнутые назад, размером 2.8 × 0.6 мкм. Позади ротовой присоски, в пределах локомоторного отдела тела шипы в основном конические, размером 7.8 × 1.9 мкм. Сравнительно немногочисленные шипы максимального размера (45.8–55.0 × 15.5–17.9 мкм) находятся на боковых сторонах тела в самой широкой части локомоторного отдела, а также на дорсальном горбе, где они единичны. На тотальных препаратах всегда отчетливо видны cемь-восемь крупных латеральных шипов на каждой стороне тела, хотя не исключено, что их число больше. В передней половине полового отдела тела элементы вооружения со слегка закругленными вершинами, размером 8.4 × 2.8 мкм. В задней половине данного отдела они вновь становятся остроконечными, сильно разреживаются и постепенно уменьшаются в направлении спереди назад до размеров, указанных для уровня ротовой присоски.
Рис. 2.
Покровы D. hispida на поперечных срезах тела: спинная (а) и боковая (б) стороны локомоторного отдела и спинная (в) сторона в задней половине полового отдела тела. 1 – наружная пластинка тегумента, 2 – шипы, 3 – кольцевые мышцы, 4 – продольные мышцы, 5 – диагональные мышцы, 6 – цитоны тегумента, 7 – окончания дорсо-вентральных мышц.

Мускулатура покровов представлена тремя мышечными слоями – кольцевым, продольным и диагональным. В первых двух слоях форма поперечного сечения мышц овальная или округлая, а расстояние между соседними мышцами слоя обычно больше их горизонтального диаметра, в особенности на боковых сторонах тела. Диаметр кольцевых мышц в локомоторном отделе тела 1.4–2.8 × 1.4–2.5, в передней половине полового отдела – 1.4–2.8 × 0.8–2.2, в его задней половине – 0.8–1.4 × 0.7–1.1 мкм. Для продольных мышц аналогичные показатели составляют 2.8–5.6 × × 2.2–3.4, 1.7–3.4 × 1.7–3.1 и 1.4–2.8 × 1.0–2.8 мкм. Диагональные мышцы пересекаются под прямым углом. Они очень редкие, толщиной <1 мкм и потому обнаруживаются с трудом, хотя имеются по всей длине тела.
Паренхимная мускулатура включает четыре группы мышц. Продольные паренхимные мышцы иногда представлены отдельными волокнами диаметром 1.1–1.7, но чаще мышечными пучками, диаметр которых варьирует в пределах 8.4–11.2 мкм (рис. 3–5). В локомоторном отделе тела они распределены в паренхиме диффузно, на уровне кишечных ветвей – в основном экстерцекально. Самые передние точки их крепления – покровы по бокам и дорсально от ротовой присоски. Часть обсуждаемых мышц, расположенная дорсально вблизи плоскости симметрии тела, заканчивается у оснований шипов дорсального горба. Кроме того, в боковых участках дисковидного расширения локомоторного отдела тела имеются мышечные элементы, которые соединяют переднюю и заднюю стороны диска. Позади бифуркации кишечника число продольных паренхимных мышц постепенно убывает в направлении спереди назад. В передней трети полового отдела тела данные мышцы формируют довольно регулярный периферический слой, расположенный сразу под стенкой тела, и лишь небольшая часть мышечных элементов рассеяна в паренхиме. Позади этого участка они очень редкие и на уровне яичника уже не прослеживаются.
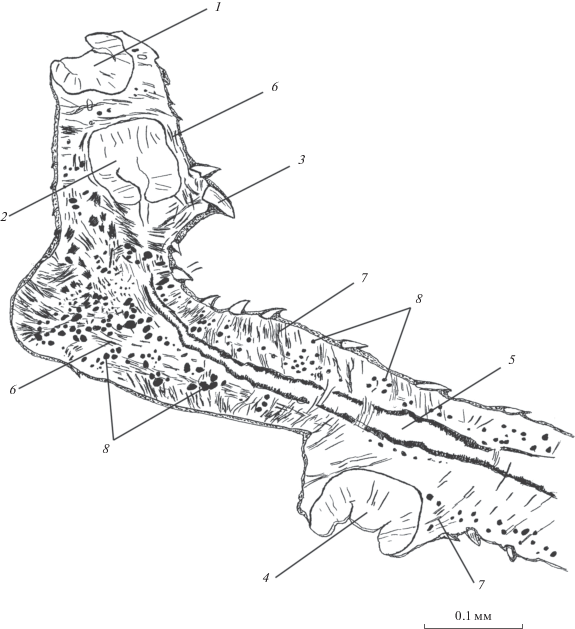
Рис. 3.
Паренхимная мускулатура D. hispida на поперечных срезах локомоторного отдела тела на уровне глотки (а) и бифуркации кишечника (б). 1 – глотка, 2 – шипы, 3 – ветвь кишечника, 4 – продольные паренхимные мышцы, 5 – дорсо-вентральные мышцы, 6 – хордальные мышцы.

Рис. 4.
Паренхимная мускулатура D. hispida на поперечных срезах тела: а – на уровне брюшной присоски, б – в передней трети полового отдела тела. 1 – брюшная присоска, 2 – ветвь кишечника, 3 – половые протоки, 4 – половой атриум, 5 – периферический слой продольных паренхимных мышц, 6 – продольные паренхимные мышцы в толще тела, 7 – дорсо-вентральные мышцы, 8 – хордальные мышцы.
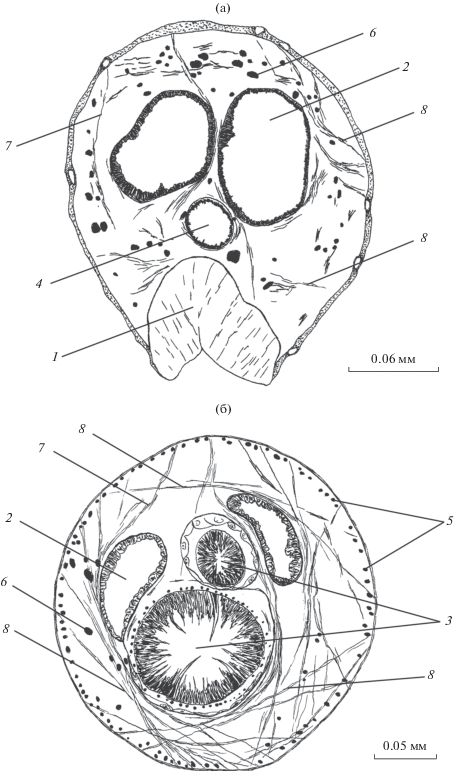
Рис. 5.
Паренхимная мускулатура D. hispida на сагиттальном срезе локомоторного отдела тела. 1 – ротовая присоска, 2 – глотка, 3 – шип на дорсальном горбе, 4 – брюшная присоска, 5 – ветвь кишечника, 6 – продольные паренхимные мышцы, 7 – дорсо-вентральные мышцы, 8 – хордальные мышцы.
Хордальные паренхимные мышцы на участке от глотки до брюшной присоски включительно имеют поперечное направление, соединяя боковые стороны тела над и под пищеварительной трубкой (рис. 3–5). Большая часть мышечных элементов диаметром ≤14 мкм сконцентрирована в передней половине названного участка, в основном под кишечником. Концевые ответвления мышц крепятся к покровам у оснований шипов. В передней трети полового отдела тела, до наполненных яйцами петель матки толщина нерегулярно расположенных хордальных мышц ≤2 мкм, а их архитектоника усложняется (рис. 4б). Помимо поперечного направления, они располагаются также в латеро-дорсальном и латеро-вентральном направлениях на каждой из боковых сторон тела, формируя в совокупности редковолокнистый мышечный плексус вокруг прямых терминальных частей половых протоков. В двух задних третях полового отдела тела данная группа мышц не обнаружена.
Дорсо-вентральные мышцы хорошо развиты в локомоторном отделе тела, особенно в широкой части вентральной впадины, где их толщина варьирует в пределах 4.2–11.2 мкм (рис. 3). На обоих концах мышц имеются терминальные конусы, достигающие иногда значительных размеров. Мышечные волокна, прилегающие к боковым сторонам ротовой присоски и глотки, заметно изогнуты в латеральных направлениях, поскольку их концы крепятся к покровам вблизи плоскости симметрии тела. Подобным образом они огибают с боков кишечник в районе его бифуркации. В пределах вентральной впадины толщина дорсовентральных мышц у боковых сторон тела не уменьшается. Мышцы, расположенные рядом с основаниями крупных латеральных шипов, нередко имеют дугообразный изгиб к плоскости симметрии тела. На уровне брюшной присоски дорсо-вентральные мышцы заметно разреживаются, а их толщина уменьшается до 1.5–1.7 мкм. Часть из них крепится вентральными концами к оболочке присоски. Указанная толщина сохраняется в передней трети полового отдела тела, где дорсо-вентральная мускулатура участвует в формировании упоминавшегося мышечного плексуса вокруг терминальных частей половой системы (рис. 4). В средней трети полового отдела мышцы становятся очень редкими, толщиной ≤1 мкм. Позади яичника они не обнаружены.
Последняя группа паренхимных мышц – это короткие волокна толщиной <1 мкм, которые соединяют дорсальную и боковые стороны ротовой присоски в ее средней части с покровами на спинной стороне тела. От поверхности присоски они расходятся веерообразно в дорсальном и дорсо-латеральных направлениях.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В строении покровов D. hispida обращает на себя внимание очень крупный размер элементов вооружения на дорсальном горбе и на боковых сторонах широкой части локомоторного отдела тела. Большие шипы подвергаются опасности выламывания из наружной пластинки тегумента при значительной механической нагрузке. Известно, что у некоторых видов трематод сем. Echinostomatidae опасность нивелируется расположением шипов адорального диска в толще базальной пластинки. При этом подвижность шипов обеспечивается либо паренхимными мышцами, крепящимися к покровам у их основания, либо собственной мускулатурой (Fujino et al., 1994; Ястребов, Ястребова, 2014). У крупных шипов D. hispida мы не обнаружено аналогичной оболочки, что косвенно свидетельствует об относительно слабых механических воздействиях. С основаниями шипов топографически связаны точки крепления дорсо-вентральных, хордальных и продольных паренхимных мышц. Их сокращение, несомненно, меняет угол наклона шипов к поверхности тела. Мускулатура покровов, напротив, слабая. Слабость выражается в относительно небольшой толщине всех мышечных слоев. Для кольцевой и продольной мускулатуры ранее отмечалось, что расстояние между соседними волокнами в мышечном слое варьирует у трематод незначительно. Оно редко превышает горизонтальный диаметр волокна и крайне редко достигает двух диаметров (Ястребов и др., 2008), поэтому все мышечные слои покровов D. hispida могут быть оценены как разреженные, в особенности диагональный слой, который, на наш взгляд, частично редуцирован.
Продольные паренхимные мышцы, лучше всего развитые в локомоторном отделе тела, имеют разную функциональную нагрузку. Они дополняют собой продольную мускулатуру покровов и укорачивают тело при перемещении гельминта. Помимо этого, мышечные элементы, соединяющие покровы на передней и задней сторонах дисковидного расширения тела, регулируют форму диска, делая его короче и шире. В последнем случае налицо очередной пример конвергентного формирования у трематод коротких продольных мышц, которые стягивают стороны различных фиксаторных структур и вызывают их выступание в том или ином направлении (Ястребов и др., 2020).
Дорсо-вентральные мышцы, расположенные в пределах вентральной впадины, регулируют ее объем, а также высоту дорсального горба. Их мощное развитие и особенности архитектоники однотипны у трематод с вентральной впадиной, функционирующей как большая присоска (Mac-Kinnon, 1982a, 1982b; Ошмарин, Жарикова, 1984). Единственным отличием является то, что латеральные мышечные волокна не уступают в толщине медиальным. По-видимому, мышцы, расположенные у боковых сторон дисковидной части тела, уплощают и расширяют ее, приводя крупные шипы в плотный контакт с субстратом. Позади вентральной впадины их роль в обеспечении двигательной активности гельминта гораздо меньше, на что указывают малая толщина, более редкое расположение и округлая форма поперечного сечения тела.
В локомоторном отделе тела регуляторами формы его дисковидной части, наряду с дорсо-вентральной и продольной паренхимной, служит хордальная мускулатура. Сокращение хордальных мышц, расположенных над пищеварительной трубкой, стягивает боковые стороны отдела, увеличивает высоту дорсального горба, приводя крупные шипы на выступе в контакт с тканями хозяина. Под пищеварительной трубкой хордальных мышц больше. Их сокращение предположительно не только стягивает стороны дисковидного расширения тела, но и сгибает тело на брюшную сторону, увеличивая объем вентральной впадины.
В передней трети полового отдела тела плексус из дорсо-вентральных и разнонаправленных хордальных мышц едва ли связан с процессами прикрепления и перемещения гельминта. Более вероятно, что сокращение компонентов плексуса обеспечивает перемещение половых продуктов и яиц по терминальным частям мужских и женских половых протоков. Периферический слой продольных паренхимных мышц обеспечивает продольное сжатие данного участка тела. Таким образом, архитектоника паренхимной мускулатуры дифференцирует половой отдел тела на две функционально разные части, задняя из которых более пассивна в двигательном плане.
По внешнему строению D. hispida сходен с видами трематод, прикрепительные структуры которых сконцентрированы на переднем конце тела. Эти структуры образуют в совокупности компактный комплекс, и его части дополняют друг друга, играя разную роль в обеспечении фиксации гельминтов. Как следствие, достигается весьма прочное прикрепление, а часть тела позади прикрепительного комплекса становится относительно длинной (Ошмарин, Егорова, 1978). Подобное строение имеют, например, трематоды некоторых родов сем. Echinostomatidae (Patagifer, Hypoderaeum, Nephrostomum). У представителей родов Patagifer и Hypoderaeum комплекс включает в себя присоски, вентральную впадину, адоральный диск и мелкие шипы покровов между присосками. У трематод рода Nephrostomum последний компонент отсутствует. В аналогичный комплекс D. hispida входят присоски, дорсальный горб, вентральная впадина и вооружение покровов.
Известно, что в эволюции функций существует явление компенсации. Суть его состоит в перераспределении нагрузки между структурами, обеспечивающими ту или иную функцию. Прогрессивное развитие одних структур компенсирует отставание в развитии других. Компенсирующие структуры не приобретают новых функций, а лишь усиливают имеющиеся (Воронцов, 1967). Так, в родах Patagifer и Nephrostomum мощное развитие адорального диска компенсирует относительную слабость мелкой ротовой присоски, особенно заметную у второго из названных родов. У трематод рода Hypoderaeum характер компенсации иной: ротовая присоска маленькая, адоральный диск частично редуцирован, и особую роль в фиксации гельминтов приобретают глубокая вентральная впадина и мускулистая брюшная присоска, субтерминальное устье которой направлено во впадину.
В случае с D. hispida основная фиксаторная нагрузка ложится не на маленькие присоски, а на участок тела между ними. Общее количество, разнообразие и пространственное распределение паренхимных мышц говорят о том, что данный участок может существенно изменять форму. Изменения формы необходимы для обеспечения контакта с тканями хозяина. При этом прикрепление происходит в основном с помощью подвижного вооружения. Его топография, не вполне типичная для трематод, указывает, что с субстратом наиболее надежно сцепляются боковые стороны локомоторного отдела тела и дорсальный горб, где шипы достигают максимальных размеров. Положение горба свидетельствует о глубоком проникновении переднего конца тела между микроворсинками кишечника рыб. Определенную прикрепительную работу выполняет также вентральная впадина. По-видимому, D. hispida сохраняет локализацию в основном за счет прикрепления, а возможности перемещения гельминтов ограничены.
Выводы. Морфология D. hispida показывает лишь один из множества вариантов приспособления к жизни в пищеварительной трубке позвоночных животных. Различные группы кишечных сосальщиков (Echinostomatidae, Notocotylidae, Strigeidae, Leucochloridiidae, Cladorchidae и многие другие) демонстрируют огромные эволюционные возможности класса Trematoda. На уровне внешнего строения диапазон возможностей известен, а способы организации гельминтов более или менее типизированы. На очереди стоит анатомический уровень исследований, в частности, изучение архитектоники мускулатуры марит. Оно позволит более точно объяснить механизмы функционирования структур, обеспечивающих прикрепление и передвижение этих интересных паразитических червей.
Список литературы
Воронцов Н.Н. 1967. Эволюция пищеварительной системы грызунов (мышеобразные). Новосибирск: Наука.
Ошмарин П.Г. 1959. К изучению специфичной экологии гельминтов. Владивосток: Изд-во Дальневост. науч. центра АН СССР.
Ошмарин П.Г. 1960. К понятию “фиксация гельминтов” // Биологические ресурсы Дальнего Востока. Москва: Изд-во АН СССР. С. 182.
Ошмарин П.Г., Егорова М.Н. 1978. Эколого-морфологические типы трематод // Экология гельминтов. Ярославль: Изд-во Ярослав. гос. ун-тa. С. 52.
Ошмарин П.Г., Жарикова Т.И. 1984. Новый вид и род трематод Cortinasoma ocadiae gen. et sp. nov. (Pronocephalata, Pronocephalidae) – паразит пресноводной черепахи из Вьетнама // Паразиты животных и растений. Владивосток: Изд-во Дальневосточного научн. центра АН СССР. С. 44.
Скрябина Е.С. 1974. Гельминты осетровых рыб (Acipenseridae Bonaparte, 1831). Москва: Наука.
Ястребов М.В., Ястребова И.В. 2014. Мышечная система трематод (строение и возможные пути эволюции). Москва: Товарищество науч. изданий КМК.
Ястребов М.В., Ястребова И.В., Филимонова Л.В. 2008. Об эволюционной пластичности мышечной системы гермафродитного поколения трематод (Plathelminthes, Trematoda) // Тр. Центра паразитологии Ин-та проблем экологии и эволюции РАН. Т. 45. Москва: Наука. С. 184.
Ястребов М.В., Глаголева Е.Н., Ястребова И.В. 2020. Конвергенции в архитектонике мускулатуры трематод и соотношение понятий “конвергенция” и “параллелизм” // Биология внутр. вод. № 6. С. 528. https://doi.org/10.31857/S0320965220060200
Choudhury A., Dick T. 1998. Systematics of the Deropristiidae Cable & Hunninen, 1942 (Trematoda) and biogeographical associations with sturgeons (Osteichthyes: Acipenseridae) // Systematic Parasitology. V. 41. P. 21.
Fujino T., Fried B., Hosier D. 1994. The expulsion of Echinostoma trivolvis (Trematoda) from ICR mice: extension/retraction mechanisms and ultrastructure of the collar spines // Parasitol. Res. V. 80. P. 581.
MacKinnon B. 1982a. The structure and possible function of the ventral papillae of Notocotylus triserialis Diesing, 1839 // Parasitology. V. 84. № 2. P. 313–332.
MacKinnon B. 1982b. The histology, ultrastructure and histochemistry of the ventral surfaces of Catatropis verrucosa (Froelich, 1789) Odhner, 1905 and Paramonostomum alveatum (Mehlis in Creplin, 1846) Luhe, 1909 (Digenea: Notocotylidae) // Can.J.Zool. V. 60. № 10. P. 2434.
Oshmarin P. 1958. On the differentiation of the body of trematodes into motor and genital parts and on the phylogenetical age of this property // Acta Veterinaria. V. 8. P. 257.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод