

Биология внутренних вод, 2022, № 1, стр. 33-44
Фенетическая радиация особей генерализованной формы крупных африканских усачей комплекса Barbus intermedius в оз. Тана (Эфиопия)
a Институт проблем экологии и эволюции им. А.Н. Северцова Российской академии наук
Москва, Россия
b Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
пос. Борок, Некоузский р-н, Ярославская обл., Россия
* E-mail: adissa@mail.ru
Поступила в редакцию 25.01.2021
После доработки 12.02.2021
Принята к публикации 16.02.2021
- EDN: HEFAVG
- DOI: 10.31857/S0320965222010107
Аннотация
Выполнен анализ структуры фенетического разнообразия особей генерализованной формы в полиморфной популяции крупных африканских усачей оз. Тана. Показано, что в пространстве рассматриваемых признаков генерализованные усачи распределены неравномерно, образуя группы, в той или иной степени между собой разобщенные. Некоторые группы различаются без перекрывания, отношения других можно определить, как “гантелевидные структуры”. Это можно рассматривать как отражение разнонаправленной радиации, источником которой является одна из групп. Результаты исследования согласуются с гипотезой о подобии генерализованной формы предковой форме, давшей начало многообразию комплекса B. intermedius sensu Banister.
ВВЕДЕНИЕ
Озеро Тана, из которого берет начало р. Голубой Нил, расположено на Абиссинском нагорье в северной части Эфиопии (рис. 1). С годами уникальный по своему своеобразию животный мир озера все больше привлекает внимание зоологов весьма широкого круга интересов: от ихтиологов (Rüppell, 1836; Boulenger, 1902; Bini, 1940; Brunelli, 1940; Nagelkerke et al., 1994; Mina et al., 1996; Nagelkerke, Sibbing, 2000; Nagelkerke, Rossberg, 2014) до специалистов по планктонным организмам (Крылов и др., 2020а, 2020б). Ихтиофауна озера примечательна, прежде всего, обилием морфо-экологических форм (морфотипов) крупных африканских усачей комплекса Barbus (=Labeobarbus) intermedius (sensu Banister, 1973) (рис. 2). Считается, что комплекс танских морфотипов – результат адаптивной радиации в процессе разделения пищевых ресурсов озера (Sibbing et al., 1998; Sibbing, Nagelkerke, 2001). Морфологическое разнообразие усачей оз. Тана впечатляет: отмечается, что различия между некоторыми морфотипами сопоставимы с различиями видов и даже родов Cyprinidae Евразии (Nagelkerke et al., 1994). Таксономический статус танских морфотипов исследователи оценивают по-разному. Одни считают их видами (или подвидами) (Rüppell, 1836; Boulenger, 1902; Bini, 1940; Nagelkerke, Sibbing, 2000), другие – экоморфами одного полиморфного вида (Banister, 1973), третьи полагают, что некоторые формы представляют собой экоморфы (Brunelli, 1940; Mina et al., 1996), а некоторые – “безмерные виды” в понимании Майра (1971). Наряду с усачами специализированных морфотипов в оз. Тана обитают всеядные особи без выраженных признаков той или иной пищевой специализации. При этом, если большинство морфотипов – это эндемики озера, то всеядные особи, рассматриваемые в качестве генерализованной формы (GF), широко распространены в реках и озерах Эфиопии. Из них состоят мономорфные популяции крупных африканских усачей, а в полиморфных популяциях генерализованные особи доминируют по численности, как и в оз. Тана. Считается, что GF подобна предковой форме, в процессе диверсификации, давшей начало многообразию комплекса B. intermedius в ареале (Nagelkerke et al., 1994; Mina et al., 1996; Левин, 2003; Голубцов, 2010; Levin et al., 2019). Внешне (при визуальной оценке габитуса) особи GF производят впечатление весьма между собой схожих, образно говоря, “все на одно лицо”. Вместе с тем анализ изменчивости краниологических признаков показал, что взятые в разных водоемах выборки особей, одинаково именуемых генерализованными, различаются между собой не меньше, а в некоторых случаях – больше, чем выборки специализированных морфотипов оз. Тана (Мироновский, 2017). Это заставляет задуматься о том, сколь схожи между собой генерализованные особи одной популяции? Гомогенны ли слагаемые ими локальные совокупности? Или же в пространстве тех или иных признаков соплеменные особи генерализованных усачей распределены неравномерно, образуя феноны, т.е., группы, особи которых между собой сходны больше, чем с особями других подобных групп?
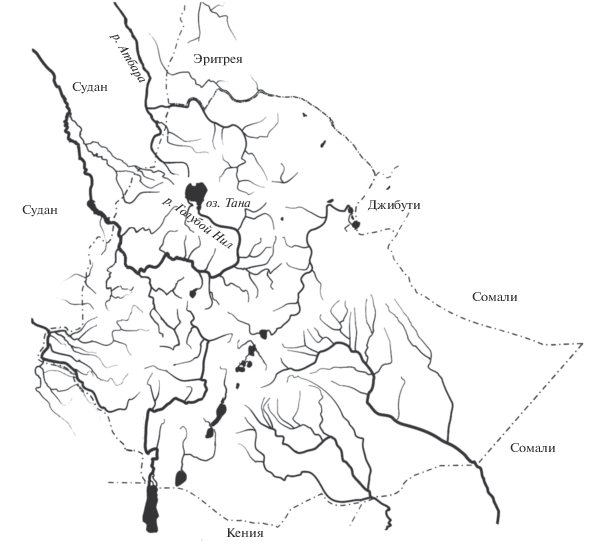
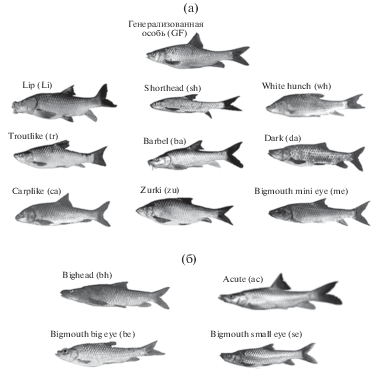
Рис. 2.
Морфо-экологические формы крупных африканских усачей комплекса Barbus intermedius оз. Тана: а – генерализованная особь и формы (морфотипы), внешне сравнительно сходные с генерализованной формой (GF); б – формы, внешне весьма отличающиеся от генерализованной.
Цель работы – дать анализ фенетического разнообразия усачей GF в оз. Тана.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Материал собирали на разных участках акватории оз. Тана в период с 1992 по 2010 гг. Рыб добывали жаберными и накидными сетями, брали из уловов местных рыбаков. Всего рассмотрено 165 особей GF стандартной длиной (SL) от 10 до 34 см. Отметим, что в оз. Тана GF крупных африканских усачей описана как морфотип intermedius11 (Nagelkerke et al., 1994; Мина и др., 2016). Для оценки морфологических отношений усачей разных форм, наряду с генерализованными, в исследовании рассматриваются особи специализированных морфотипов оз. Тана: acute – 75 особей; barbel – 17; bighead – 11; bigmouth big eye – 56; bigmouth mini eye – 10 особей; bigmouth small eye – 51; carplike – 12; dark – 35; troutlike – 41; white hunch – 48; zurki – 54; Lip – 27; shorthead – 13 особей. Принадлежность особи к тому или иному морфотипу (включая intermedius) определяли по диагностической таблице (Nagelkerke et al., 1994).
У каждой особи проведено 14 промеров черепа (рис. 3). Исследования изменчивости крупных африканских усачей показали, что обладая высокой разрешающей способностью и хорошей воспроизводимостью результатов измерений как одним, так и разными операторами, данный набор признаков позволяет достаточно уверенно оценивать фенетические отношения большинства морфотипов (Mina et al., 1993, 1996, 1998; Мина и др., 2005; Мироновский, 2006; Кожара и др., 2020).
Рис. 3.
Схема промеров черепа африканских усачей комплекса B. intermedius: а – признаки осевого черепа; б – признаки висцерального черепа; в – признаки висцерального черепа, отражающие морфологию челюстей. BL − базальная длина черепа; B1, B2, B3 – расстояние между внешними краями frontalia, pterotica и sphenotica соответственно; B4 – ширина черепа на уровне соединения frontale и pteroticum, HS1 и HS2 – высота черепа на уровне изгиба parasphenoideum и заднего края parasphenoideum соответственно, Hm – высота hyomandibulare, Pop − длина praeoperculum, Op – высота передней части operculum, lop и Pmx – длина interoperculum (межкрышечной кости) и praemaxillare, Mx – длина maxillare, De – длина dentale.
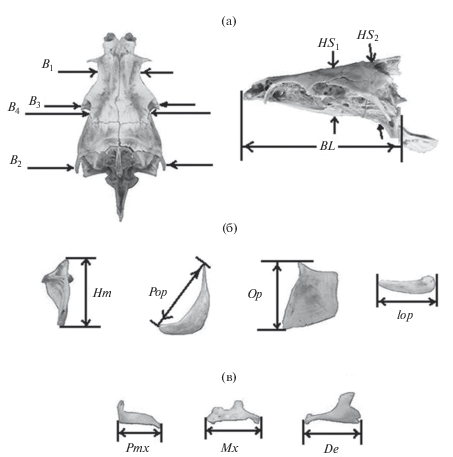
Статистическая обработка данных выполнена средствами пакетов NTSYS 2.02k (Rohlf, 1998) и Статистика 6. В расчетах использованы индексы, представляющие собой отношения абсолютных значений промеров к базальной длине черепа (BL). Далее в обсуждении, упоминая тот или иной признак, мы будем иметь в виду его индекс, а не сам промер. Значения индексов преобразовывали в натуральные логарифмы для нормализации распределений. В качестве показателя сходства использована обобщенная таксономическая дистанция (Rolf, 1998). Кластерный анализ матриц сходства между особями проводили методом complete linkage; его результаты иллюстрирует дендрограмма. При АГК учитывали рекомендацию Л. А. Животовского (1984) разделить признаки на определенные группы, для каждой группы АГК провести отдельно и далее анализировать ГК1 каждой группы. Ранее установлено (Мироновский, 2017), что в процессе диверсификации экологических форм танских усачей рассматриваемые признаки делятся на две корреляционные плеяды. В одну входят шесть признаков осевого черепа (B1, B2, B3, B4, HS1, HS2) и три признака висцерального черепа (Hm, Pop, Op). Вторую слагают четыре признака висцерального черепа, три из которых (Pmx, Mx и De) отражают морфологию челюстей, что дает основания рассматривать изменчивость данной плеяды как прямо связанную с особенностями питания22. Четвертый признак второй плеяды (lop) характеризует длину межкрышечной кости. В черепе усачей эта кость протянулась за зубной, что, по-видимому, определяет тесную корреляцию между нею и De. С учетом такого разделения АГК проводили раздельно для плеяды из девяти признаков и для плеяды из четырех признаков, затем анализировали распределение точек-объектов в координатах ГК1 “плеяды девяти” и ГК1 “плеяды четырех”. Такая ординация дает возможность наглядно соотнести вклад каждой плеяды в структуру распределения объектов на плоскости ГК. Собственные векторы рассчитывали по корреляционной матрице; длину вектора принимали равной 1.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Визуальный анализ дендрограммы сходства особей GF усачей комплекса B. intermedius (рис. 4) дает основания выделить пять кластеров – a, b, c, d и e, уровень различий между которыми заметно выше уровня различий внутри каждого из них. Наличие кластеров предполагает гетерогенность изучаемого множества, что требует объяснения при анализе отношений особей между собой весьма сходных (визуально производящих впечатление неразличимых). Одно из возможных объяснений проистекает из того, что установить принадлежность крупных танских усачей к определенному морфотипу можно лишь по достижению ими некоторой длины, в качестве которой Нагелькерке с соавт. (Nagelkerke et al., 1995) указывает SL ~10 см. Мина с соавт. (Mina et al., 1996) утверждает, что при SL = 10 см уверенно идентифицировать можно лишь два морфотипа – acute и bigmouth big eye, надежное же определение остальных возможно при SL 20–25 см. В составе четырех из пяти кластеров есть особи достаточно крупные (табл. 1), чтобы даже по более жестким критериям Мины с соавт. (1996) выявленную гетерогенность нельзя было объяснить тем, что в составе изучаемой совокупности присутствуют усачи не только GF, но и специализированных морфотипов, которые еще недостаточно выросли для надежной идентификации. Длины 20 см не достигают лишь особи кластера “b”.
Рис. 4.
Дендрограмма сходства особей генерализованной формы (GF) усачей комплекса B. intermedius в оз. Тана. D – обобщенная таксономическая дистанция. a, b, c, d и e – группы (кластеры).
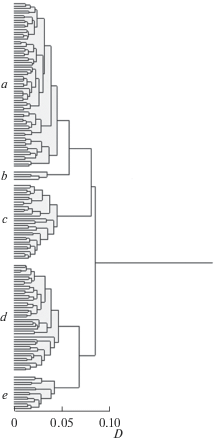
Таблица 1.
Особи разных размерных групп: распределение в кластерах дендрограммы
| Кластер дендрограммы | Размерная группа особей GF, см | ||||
|---|---|---|---|---|---|
| 10.0–15.0 | 15.0–20.0 | 20.0–25.0 | 25.0–30.0 | 30.0–35.0 | |
| a | 8 | 25 | 23 | 12 | 2 |
| b | 1 | 3 | – | – | – |
| c | 11 | 14 | 7 | – | – |
| d | 10 | 19 | 9 | 5 | 2 |
| e | – | 3 | 3 | 6 | 2 |
| Всего особей, экз. | 30 | 64 | 42 | 23 | 6 |
В качестве другого объяснения гетерогенности можно предположить, что в разные кластеры попали особи выборок разных лет, и наблюдаемая дифференциация имеет хронологический характер. Однако данные табл. 2 свидетельствуют, что это не так. В достаточно объемных кластерах (“a”, “c” и “d”) присутствуют особи сборов разных лет, и даже четыре особи наименьшего из всех кластера “b” пойманы в разные годы (1993, 1996, 1997). Обращает на себя внимание распределение особей, пойманных в 2010 г., после того как в результате чрезмерного промысла численность танских морфотипов резко снизилось, и некоторые морфотипы почти перестали встречаться (De Graaf et al., 2004; Dejen et al., 2017). В каждом из больших кластеров (“a”, “c” и “d”) особи 2010 г. присутствуют, а их отсутствие в кластерах “b” и “e” с большой вероятностью объясняется малым объемом двух этих кластеров в сочетании с малым объемом выборки 2010 г. Таким образом, нет оснований полагать, что после снижения численности специализированных морфотипов структура фенетического разнообразия особей GF в озере существенно изменилась.
Таблица 2.
Число особей в выборках разных лет: распределение в кластерах дендрограммы
| Год сбора | Кластеры дендрограмы | ||||
|---|---|---|---|---|---|
| a | b | c | d | e | |
| 1992 | 8 | – | 4 | 10 | 6 |
| 1993 | 4 | 1 | 2 | 7 | 3 |
| 1994 | 2 | – | 2 | 3 | – |
| 1996 | 37 | 2 | 10 | 12 | 5 |
| 1997 | 10 | 1 | 13 | 7 | – |
| 1999 | 5 | – | – | – | – |
| 2010 | 4 | – | 1 | 6 | – |
| Всего особей, экз. | 70 | 4 | 32 | 45 | 14 |
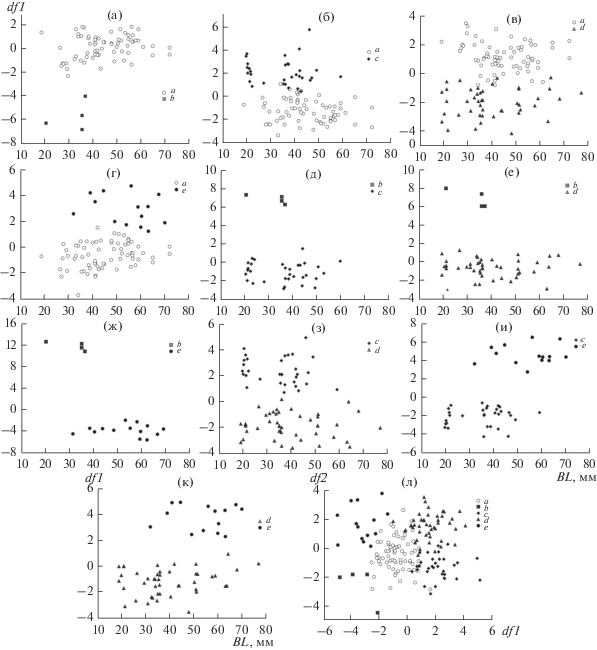
Еще одна из вероятных причин выявленной гетерогенности GF в оз. Тана – возможные различия разноразмерных групп в результате изменения пропорций черепа по мере роста рыб. На рис. 5 по оси ординат даны значения дискриминантной функции (df1), полученной по совокупности 14 рассматриваемых признаков, по оси абсцисс – базальная длина основания черепа (BL). Такая ординация дает возможность визуально оценить соотношение различий, обусловленных разными размерами усачей сравниваемых групп, с различиями, которые разницей размеров объяснить нельзя. В семи из десяти возможных сочетаний (рис. 5а, 5д–5к) одноразмерные особи сравниваемых групп по оси ординат различаются без перекрывания, при этом в шести сочетаниях (рис. 5а, 5д–5ж, 5и, 5к) различия с выраженным хиатусом. В трех сочетаниях (рис. 5б–5г) распределения перекрываются, однако тенденция к разделению одноразмерных особей сравниваемых совокупностей сомнений не вызывает. Следовательно, наблюдаемые различия нельзя объяснить постепенным изменением пропорций по мере роста рыб.
Рис. 5.
Результаты дискриминации пяти групп GF усачей комплекса B. intermedius из оз. Тана: а – а и b, б – а и с, в – а и d, г – а и e, д – b и с, е – b и d, ж – групп b и e, з – групп с и d, и – групп с и е, к – групп e и d, л – всех пяти выявленных групп одновременно; df1 и df2 – первая и вторая дискриминантные функции, BL – базальная длина основания черепа особей.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Полученные результаты характеризуют ситуацию противоречиво. По диагностическим признакам внешней морфологии каждая особь рассматриваемого множества отнесена к одной совокупности – морфотипу intermedius, т.е. к GF. Вместе с тем, анализ отношений особей этой совокупности по остеологическим признакам выявляет группы, различающиеся между собой без перекрывания, в большинстве сопоставлений – с отчетливым хиатусом. Ключ к пониманию ситуации дают рис. 5б–5г, где распределения можно рассматривать как в разной степени выраженные “гантелевидные структуры”, то eсть состояния, которые, согласно модели симпатрического видообразования Кондрашова и Мины (Kondrashov, Mina, 1986), предшествуют полному обособлению групп особей, дивергирующих в условиях симпатрии. Исходя из этого, наблюдаемое соотношение различий можно объяснить фенетической радиацией особей изучаемой совокупности. Центр и источник радиации – группа “а”. От нее дивергируют группы “с”, “d” и “e”. В результате дивергенции различия особей групп, расходящихся в разные направления, достигли уровня полной разобщенности вплоть до хиатуса, тогда как с общим центром у каждой группы фенетическая связь сохранилась. Особое положение занимают особи группы “b”, с хиатусом отличающиеся от прочих, включая особей группы “а” – предполагаемого источника радиации. На наш взгляд, возможная причина наблюдаемого хиатуса – это малая численность группы “b”. Согласно правилу убывания дискретности (Мина, 1986), с увеличением численности выборки “b”, хиатус между нею и группой “а” с большой вероятностью заполнится. Другое объяснение полного обособления особей группы “b” от прочих исходит из возможности полифилетического происхождения танского комплекса. В этом случае их можно рассматривать как представителей иной линии GF в процессе множественного вселения в озеро.
Результаты дискриминации пяти рассматриваемых групп одновременно отражены на рис. 5л. Распределение особей на плоскости первой (df1) и второй (df2) дискриминантных функций соответствует гипотезе о фенетической радиации GF в оз. Тана. В центре расположены особи группы “a”, в четырех направлениях от центра – особи групп-дериватов. Примечательно, что группы “с” и “d” на рис. 5л перекрываются, тогда как на рис. 5з их распределения разобщены. Это хорошо иллюстрирует большую надежность дискриминирующих алгоритмов при попарном сличении сравниваемых совокупностей. С увеличением числа сличаемых групп разделение в проекции на двумерную плоскость может не наблюдаться, хотя в n-мерном пространстве исходных признаков распределения находятся обособленно.
На рис. 6 приведены результаты АГК средних оценок признаков пяти обсуждаемых кластеров GF и 13 морфотипов озера. По оси абсцисс даны значения ГК1 “плеяды четырех”, по оси ординат – значения ГК1 “плеяды девяти”. Таким образом, ось абсцисс отражает изменчивость параметров, прямо связанных с процессом захвата и удержания пищевых объектов, т.е. изменчивость, сопряженную с диверсификацией особей в процессе разделения пищевых ресурсов. Ось ординат отражает изменчивость признаков, прямо с особенностями питания не связанных. Собственные векторы, собственные значения и процент связанной дисперсии приведены в табл. 3. Видно, что ось абсцисс отражает >80% изменчивости плеяды признаков, характеризующих морфологию челюстей, ось ординат >70% изменчивости признаков второй плеяды. При этом, в каждом векторе вклады признаков одинаковы по знаку и близки по абсолютной величине. Следовательно, двумерное распределение на рис. 6 с большой полнотой и минимальными искажениями отображает фенетические отношения танских морфотипов и пяти выявленных групп GF в многомерном пространстве исходных параметров. Графические символы (⚪, ◆, ▲, ◼ и ⚫) пяти групп особей GF соединены кратчайшей связывающей сетью. Видно, что конфигурация сети соответствует гипотезе о фенетической радиации, источник которой – группа “а”, а направления отрезков кратчайшей связывающей сети позволяет судить о направлениях дивергенции дериватов.
Рис. 6.
Результаты АГК средних оценок признаков 13 специализированных морфотипов (обозначения см. рис. 2) и пяти групп (a, b, c, d, e) GF комплекса B. intermedius из оз. Тана. Обозначение осей разъяснено в тексте.
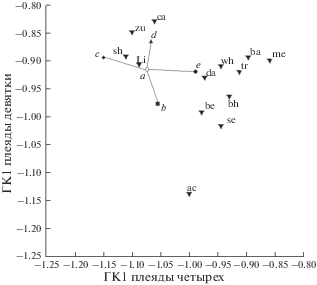
Таблица 3.
Собственные значения и весовые вклады признаков в собственные векторы главных компонент корреляционных плеяд признаков танских морфотипов
| Признак | ГК1ось X | ГК1оcь Y |
|---|---|---|
| B1 | – | 0.352 |
| B2 | – | 0.365 |
| B3 | – | 0.375 |
| B4 | – | 0.365 |
| HS1 | – | 0.304 |
| HS2 | – | 0.255 |
| Hm | – | 0.377 |
| Pop | – | 0.222 |
| Op | – | 0.348 |
| lоp | 0.478 | – |
| Pmx | 0.543 | – |
| Mx | 0.468 | – |
| De | 0.508 | – |
| Собственные значения | 3.210 | 6.490 |
| Доля объясненной дисперсии, % | 80.169 | 72.080 |
Примечание. Обозначения см. рис. 3.
Пищевые предпочтения экологических форм усачей оз. Тана, описаны в ряде работ (Nagelkerke et al., 1994; Sibbing at al., 1998; Nagelkerke, Sibbing, 2000; Sibbing, Nagelkerke, 2001; De Graaf, 2003; Nagelkerke, Rossberg, 2014). Генерализованные особи, как отмечено выше, всеядны. Относительно всеядны и особи морфотипа Lip (Li), хотя значительную долю их рациона составляют бентос и донные беспозвоночные, извлекаемые гипертрофированными губами из расщелин на каменистых участках дна. Особи морфотипа zurkis (zu) – поедатели высшей водной растительности; особи carplike (ca) питаются двустворчатыми моллюсками. Основная пища особей морфотипа shorthead (sh) – зоопланктон и взрослые насекомые. Если на рис. 6 к оси абсцисс мысленно провести вертикаль при значении ГК1 “плеяды четырех”, равном ~ (–1.03), перечисленные морфотипы окажутся слева от этой воображаемой линии. Справа от нее расположились графические символы так называемых “большеротых” (“large-mouthed”) морфотипов усачей оз. Тана (Mina at al., 2001). К их числу относятся рыбоядные хищники acute (ac), bigmouth big eye (be), dark (da), white hunch (wh), bigmouth small eye (se), bighead (bh), trout (tr), bigmouth mini eye (me), а также питающиеся детритом и донными личинками насекомых особи морфотипа barbel (ba). Уровень внешнего сходства с генерализованной особью позволяет разделить морфотипы усачей оз. Тана на две группы (рис. 2). Если в габитусе ca, zu, Li, sh, wh, tr, ba, da и me черты сходства с GF проглядываются вполне отчетливо, то облик bh, be, se и ac выразительно “иной”. Вероятно, отмечая различия, сопоставимые с таковыми различиями родов карповых рыб Евразии (Nagelkerke et al., 1994), авторы работы имели в виду уровень дивергенции этих четырех морфотипов.
Из рис. 6 видно, что особи группы “с” дивергируют в направлении морфотипа sh, особи “d” – в направлении морфотипов са и zu. Радиация групп “e” и “b” направлена к большеротым усачам: “e” – в сторону форм, сохраняющих внешнее сходство с генерализованной, “b” – в сторону “иных”, точнее – к морфотипам be и ac. Последнее дает основания утверждать, что даже сравнительно мелкие особи группы “b” (табл. 1) едва ли могут быть усачами специализированных морфотипов, недостаточно крупными для их безошибочного определения, ибо даже по жестким критериям Мины с соавт. (2011) be и ac надежно определяются при длине 10 см. Вместе с тем, как отмечено выше, в рамках гипотезы о полифилетическом происхождении танского флока они могут оказаться представителями другой линии GF, давшей начало морфотипам группы “иных” – всем, или некоторым.
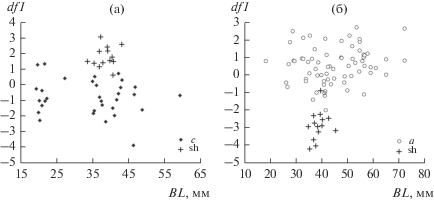
Следует отметить, что по оси ординат графические символы группы “с” и морфотипа sh расположены почти на одном уровне (рис. 6), т.е. они различаются исключительно морфологией челюстей, при отсутствии различий по другим рассматриваемым признакам. Учитывая пищевые предпочтения морфотипа sh, можно предположить, что особи группы “с” еще более, чем морфотип sh, привержены в своем питании зоопланктону и насекомым. Возможно и другое объяснение. Распределения особей групп “а”, “с” и морфотипа sh во всех трех возможных сочетаниях представляют собой гантелевидные структуры (рис. 5б, рис. 7). Это позволяет предположить, что совокупности особей sh и “с” – разные стадии дивергенции от исходной формы “а” в одном направлении.
Рис. 7.
Результаты дискриминации особей морфотипа sh и группы “с” (а), sh и группы “a” (б) GF усачей комплекса B. intermedius из оз. Тана.
Примерно на одном уровне значений оси абсцисс расположены и символы групп “а” и “е” (рис. 6). Подобно sh и “с”, они различаются почти исключительно морфологией челюстей. Мысленно проведя от символа “a” прямую, параллельную оси абсцисс, в сторону уменьшения значений ГК1 плеяды челюстной дуги, можно убедиться, что эта воображаемая линия весьма близка к линии регрессии распределения символов “а”, “е” da, wh, tr и me. Значит, при переходе от всеядных генерализованных особей к хищникам-засадчикам (da и wh) и далее к хищникам-преследователям (tr и me) меняется лишь морфология челюстей, при весьма близких значениях признаков, прямо с питанием не связанных. Это соответствует гипотезе об определяющей роли разделения пищевых ресурсов в формировании комплекса форм крупных африканских усачей в оз. Тана (Sibbing at al., 1998; Sibbing, Nagelkerke, 2001).
При анализе рис. 6 возникает вопрос о соотношении остеологических и внешнеморфологических различий между особями сравниваемых совокупностей. Специализированные морфотипы, некогда описанные на основании различий внешних признаков, хорошо различаются и по пропорциям черепа (Mina et al., 1993, 1996, 1998; Мина и др., 2011, 2013). Дивергирующие группы GF, выявленные в настоящем исследовании, по пропорциям черепа на рис. 6 различаются не меньше, а в ряде сочетаний – больше, чем морфотипы. Так, по оси абсцисс различия между группами “c” и “e” сопоставимы с различиями морфотипов da и me, по оси ординат различия между группами “d” и “b” примерно соответствуют различиям морфотипов ba и se. По диагностическим же признакам внешнего строения каждая из особей, слагающих группы GF, отнесена к одному морфотипу – intermedius, в качестве которого, как указывалось выше, в оз. Тана описана генерализованная форма. Объяснение этому несоответствию дает представление о диверсификации генерализованных особей, как о первой стадии формирования комплекса экологических форм усачей: в процессе формирования различий между формами “сперва происходит расхождение по пропорциям черепа, а уже затем по внешнему виду…”, со временем “…. различия могут увеличиваться под действием отбора, способствующего все большему развитию у специализированной формы признаков, отличающих ее от генерализованной формы” (Мина и др., 2016, с. 408). Кроме того, логично предположить, что группы GF, радиирующие в разные направления, предпочитают разные биотопы. Соответственно, в разных участках озера анализируемые уловы состоят из представителей преимущественно одной радиирующей формы в сочетании с представителями исходной формы “а”. Морфологический ряд смешанной таким образом совокупности непрерывен даже при анализе остеологических признаков (5б–5г), и неудивительно, что при сортировке таких уловов особи казались неразличимыми по признакам внешнего строения.
Следует отметить, что в первых публикациях современного периода изучения флока форм крупных усачей в оз. Тана, начатого работами Нагелькерке с соавт. (Nagelkerke et al., 1994) и Мины с соавт. (Mina et al., 1996) исследователи склонялись к идее его монофилетического происхождения. Этому, в частности, способствовала кажущаяся уникальность танского комплекса. Ситуация изменилась, когда флоки форм усачей обнаружили в других водоемах Эфиопии (Голубцов, 2010). В более поздней работе Мины с соавт. (2011) обсуждаются варианты уже полифилетического происхождения комплекса танских усачей. Со временем полифилетическое происхождение некоторых речных флоков подтвердили генетические исследования (Levin et al., 2019, 2020). Опубликованы данные, свидетельствующие о возможности полифилетического происхождении и танского флока усачей (Beshera, Harris, 2014; Levin et al., 2020).
В связи c этим необходимо подчеркнуть, что результаты настоящего исследования, основанного на анализе фенетических признаков, не свидетельствуют в пользу одной из гипотез. Заманчиво предположить, что, например, группа “e”, которая на рис. 6 радиирует в направлении морфотипов wh, tr, ba, da, me, некогда дала начало если не всем пяти этим морфотипам, то одному или нескольким из них. В то же время, очевидно, что это не более чем предположение, правдоподобное, но не доказанное (во всяком случае, на данном этапе исследований). То же можно сказать и в отношении трех других дериватов формы “а”. Результаты исследования дают основания достаточно уверенно утверждать, что совокупность генерализованных особей, обитающих в исследуемый период в озере, фенетически гетерогенна. Она подразделяется на группы, в разной степени обособленные, и отношения некоторых пар этих групп соответствуют “гантелевидным структурам”, характерным для процессов формообразования в условиях симпатрии. Вместе с тем, из факта фенетической радиации особей GF в оз. Тана, наблюдаемой в настоящее время, не следует, что именно эта совокупность генерализованных особей является исходной для каждого или для части танских морфотипов. Обоснованно судить о том, дала начало тем или иным морфотипам озера данная совокупность генерализованных особей, или же одна (или несколько) из других возможных групп-основателей в процессе множественного вселения в оз. Тана разных линий GF, позволит лишь продолжение целенаправленных генетических исследований.
Выводы. Проведенное исследование показало, что особи генерализованной формы крупных африканских усачей, обитающие в оз. Тана, в пространстве рассматриваемых признаков распределены неравномерно, образуя группы, в разной степени разобщeнные между собой. Некоторые из выявленных групп различаются без перекрывания, отношения других можно определить, как “гантелевидные структуры”, характерные для процессов формообразования в условиях симпатрии. Отношения выявленных групп можно рассматривать как свидетельство разнонаправленной фенетической радиации, источник которой – одна из групп. В процессе формирования различий между экологическими формами (морфотипами) усачей комплекса Barbus intermedius вначале происходит расхождение по пропорциям черепа, затем – по признакам внешней морфологии. Результаты исследования хорошо согласуются с гипотезой о близости генерализованной формы к форме предковой, давшей начало многообразию комплекса B. intermedius.
Список литературы
Голубцов А.С. 2010. “Пучки видов” рыб в реках и озерах: симпатрическая дивергенция в фаунистически обедненных рыбных сообществах как особый модус эволюции // Актуальные проблемы современной ихтиологии. К 100-летию Г.В. Никольского. Москва: Товарищество науч. изданий КМК. С. 96.
Животовский Л.А. 1984. Интеграция полигенных систем в популяциях: проблемы анализа комплекса признаков. Москва: Наука.
Левин Б.А. 2003. Структура локальных популяционных систем у рыб рода Barbus // Матер. ІІ міжнар. наук. конф. “Біорізноманіття та роль зооценозу в природних і антропогенних екосистемах”. Днепропетровск: Изд-во Днепропетровского национального ун-та. С. 53.
Кожара А.В., Маврин А.С., Мироновский А.Н. 2020. Структура фенетического разнообразия и систематика красноперки Scardinius erythrophtalmus водоемов и водотоков России и сопредельных стран. // Биология внутр. вод. № 6. С. 538. https://doi.org/10.31857/S0320965220060121
Крылов А.В., Зелалем В., Прокин А.А. 2020а. Качественный состав и количественные характеристики зоопланктона прибрежья оз. Тана (Эфиопия) в конце сухого сезона // Биология внутр. вод. № 2. С. 137. https://doi.org/10.31857/S0320965220020096
Крылов А.В., Зелалем В., Прокин А.А., Шкиль Ф.Н. 2020б. Зоопланктон прибрежной зоны оз. Тана в начале сухого сезона // Биология внутр. вод. № 5. С. 477. https://doi.org/10.31857/S0320965220050058
Майр Э. 1971. Принципы зоологической систематики. Москва: Мир. (Mayr E. 1969. Principles of Systematic Zoology. New York: McGraw-Hill.)
Мина М.В. 1986. Микроэволюция рыб: эволюционные аспекты фенетического разнообразия. Москва: Наука.
Мина М.В., Левин Б.А., Мироновский А.Н. 2005. О возможностях использования в морфометрических исследованиях рыб оценок признаков, полученных разными операторами // Вопр. ихтиологии. Т. 45. № 3. С. 331.
Мина М.В., Мироновский А.Н., Капитанова Д.В. 2011. Фенетические отношения и вероятные пути морфологической диверсификации африканских усачей комплекса Barbus intermedius из озера Тана (Эфиопия) // Вопр. ихтиологии. Т. 51. № 2. С. 149.
Мина М.В., Дзержинский К.Ф., Мироновский А.Н., Капитанова Д.В. 2013. Фенетическое разнообразие крупных усачей Barbus intermedius complex sensu Banister из прибрежной зоны Бахардарского залива (озеро Тана, Эфиопия) // Вопр. ихтиологии. Т. 53. № 6. С. 627. https://doi.org/10.7868/S0042875213050081
Мина М.В., Мироновский А.Н., Дгебуадзе Ю.Ю. 2016. Полиморфизм по пропорциям черепа у крупных африканских усачей Barbus intermedius sensu Banister, 1973 (Cyprinidae) из озер Аваса и Лангано (Рифтовая долина, Эфиопия) // Вопр. ихтиологии. Т. 56. № 4. С. 403. https://doi.org/10.7868/S0042875216040081
Мироновский А.Н. 2006. Факторы, обуславливающие сопоставимость данных, полученных путем оценки пластических признаков рыб // Вопр. ихтиологии. Т. 46. № 2. С. 240.
Мироновский А.Н. 2017. Место генерализованной формы в структуре фенетического разнообразия крупных африканских усачей комплекса Barbus intermedius. Метод корреляционных плеяд // Вопр. ихтиологии. Т. 57. № 4. С. 393. https://doi.org/10.7868/S0042875217040105
Banister K.E. 1973. A revision of the large Barbus (Pisces, Cyprinidae) of East and Central Africa. Pt. II // Bull. Brit. Mus. Nat. Hist. Zool. V. 26.
Beshera K.A., Harris P.M. 2014. Mitochondrial DNA phylogeography of the Labeobarbus intermedius complex (Pisces, Cyprinidae) from Ethiopia // J. Fish. Biol. V. 85. P. 228. https://doi.org/10.1111/jfb.12408
Bini G. 1940. I pesci del Lago Tana // Missione di Studio al Lago Tana. V. 3. № 2. P. 137.
Boulenger G.A. 1902. Descriptions of new fishes from the collection made by Mr. E. Degen in Abyssinia // Ann. Mag. Nat. Hist. Ser. 7. V. 60. P. 421.
Brunelli G. 1940. Le mutazioni del genere Barbus del lago Tana // Missione di Studio al Lago Tana. V. 3. № 2. P. 207.
De Graaf M. 2003. Lake Tana’s piscivorous Barbus (Cyprinidae, Ethiopia): Ecology Evolution, Exploitation. Doctoral thesis, Experimental Zoology Group, Wageningen University, P.O. Box 338, 6700 AH Wageningen, The Netherlands.
De Graaf M., Machiels M., Wudneh T., Sibbing F.A. 2004. Declining stocks of Lake Tana’s endemic Barbus species flock (Pisces, Cyprinidae): natural variation or human impact? // Biol. Conservation. V. 116. P. 277. https://doi.org/10.1016/S0006-3207(03)00198-8
Dejen E., Anteneh W., Vijverberg J. 2017. The decline of The Lake Tana (Ethiopia) fisheries: Causes and possible solutions // Land Degrad. Dev. V. 28, P. 1842. https://doi.org/10.1002/ldr.2730
Kondrashov A.S., Mina M.V. 1986. Sympatric speciation: when is it possible? // Biological J. Linnean Society. V. 27. № 3. P. 201.
Levin B.A., Casal-López M., Simonov E. et al. 2019. Adaptive radiation of barbs of the genus Labeobarbus (Cyprinidae) in the East African river // Freshwater Biol. V. 64. Iss. 10. P. 1721. https://doi.org/10.1111/fwb.13364
Levin B.A., Simonov E., Dgebuadze Yu.Yu. et al. 2020. In the rivers: Multiple adaptive radiations of cyprinid fishes (Labeobarbus) in Ethiopian Highlands // Scientific Reports. 10:7192 P. 1. https://doi.org/10.1038/s41598-020-64350-4
Mina M.V., Mironovsky A.N., Dgebuadze Yu.Yu. 1993. Allometrie et divergence entre les barbeaux du lac Tana (Ethiopie) // Cahiers d’Ethologie. V. 13. P. 219.
Mina M.V., Mironovsky A.N., Dgebuadze Yu.Yu. 1996. Lake Tana large barbs: phenetics, growth and diversification // J. Fish Biol. V. 48. P. 383.
Mina M.V., Mironovsky A.N., Golubtsov A.S., Dgebuadze Yu.Yu. 1998. ‘Barbus’ intermedius species flock in Lake Tana (Ethiopia). II. Morphological diversity of “large barbs” from Lake Tana and neighbouring areas: homoplasies or synapomorphies? // Ital. J. Zool. V. 65. Suppl. P. 9.
Mina M.V., Mironovsky A.N., Golani D. 2001. Consequences and modes of morphological diversification of East African and Eurasian barbins (genera Barbus, Varicorhinus and Capoeta) with particular reference to Barbus intermedius complex // Environ. Biol. Fish. V. 61. P. 241.
Nagelkerke L.A.J., Sibbing F.A. 2000. The large barbs (Barbus spp., Cyprinidae, Teleostei) of Lake Tana (Ethiopia), with a description of a new species, Barbus osseensis // Netherl. J. Zool. V. 50. P. 179.
Nagelkerke L.A.J., Rossberg A.G. 2014. Trophic niche-space imaging, using resource and consumer traits // Theoretical Ecology. V. 7. № 4. P.423. https://doi.org/10.1007/s12080-014-0229-5
Nagelkerke L.A.J., Sibbing F.A., Osse J.W.M. 1995. Morphological Divergence during Growth in the Large Barbs (Barbus spp.) of Lake Tana, Ethiopia // Neth. J. Zool. V. 45. P. 431.
Nagelkerke L.A.J., Sibbing F.A., Van den Boogaart J.G.M. et al. 1994. The barbs (Barbus spp.) of Lake Tana: a forgotten species flock? // Environ. Biol. Fish. V. 39. P. 1.
Rüppell E. 1836. Neuer Nachtrag von Beschreibungen und Abbildungen neuer Fische im Nil entdeckt // Abhandlungen aus dem Gebiete der beschreibenden Naturgeschichte. Bd 2. № 1. Frankfurt am Main: Mus. Senckenberg. P. 1.
Rohlf F.J. 1998. NTSYS-pc: numerical taxonomy and multivariate analysis system. Version 2.02. New York: Exeter Software Press.
Sibbing F.A., Nagelkerke L.A.J. 2001.Resource partitioning by Lake Tana barbs predicted from fish morphometrics and prey characteristics // Reviews in fish Biology and Fisheries. V. 10(4). P. 393.
Sibbing F.A., Nagelkerke L.A.J., Stet R.J.M., Osse J.W.M. 1998. Speciation of endemic Lake Tana barbs (Cyprinidae, Ethiopia) driven by trophic resource partitioning; a molecular and ecomorphological approach // Aquat. Ecol. V. 32. P. 217.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод