

Биология внутренних вод, 2022, № 2, стр. 205-208
Первые результаты молекулярно-генетического анализа европейского вселенца Eurytemora velox (Crustacea, Calanoida)
Н. М. Сухих a, *, В. И. Лазарева b
a Зоологический институт Российской академии наук
Санкт-Петербург, Россия
b Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
пос. Борок, Некоузский р-н, Ярославская обл, Россия
* E-mail: Susikh1@mail.ru
Поступила в редакцию 17.06.2021
После доработки 26.06.2021
Принята к публикации 30.06.2021
- EDN: PQOGJG
- DOI: 10.31857/S0320965222020140
Аннотация
Представлены первые результаты анализа фрагментов четырех генов СО1, ITSs и 18SrRNA для популяций вида-вселенца Eurytemora velox (Lilljeborg, 1853) из Чебоксарского водохранилища и озера в г. Киев. Анализ участков генов СО1 и ITSs показал, что исследуемые популяции различаются на несколько нуклеотидных замен. Анализ двух последовательностей ДНК консервативного ядерного гена 18SrRNA ожидаемо не выявил различий между популяциями. Сравнение полученных последовательностей с таковыми из водоемов Польши (данные международной базы GenBank; https://www. ncbi.nlm.nih.gov/nuccore/MT146445) показало, что они группируются с последовательностями вида из Чебоксарского водохранилища. Вероятно, обнаруженная в водных объектах Польши E. velox происходит из Каспийского региона, откуда расселилась и в Чебоксарское водохранилище. Популяция из озера в г. Киев, предположительно, поднялась по р. Днепр из Черноморского региона. По-видимому, вид независимо расселился по крайней мере из двух регионов-доноров.
В последние десятилетия эвригалинная Eurytemora velox (Lilljeborg, 1853) активно расселяется в пресных и солоноватых водах Европы. Вид обнаружен фактически повсеместно в солоноватых водах низовьев рек Северного, Балтийского, Каспийского, Черного и Азовского морей, а также в пресных водах всех основных рек, впадающих в эти моря (Боруцкий и др., 1991; Gaviria, Forro, 2000). С каждым годом появляются новые данные, свидетельствующие о расширении ареала E. velox. В большинстве случаев факт вселения этого вида очевиден. Но иногда непонятно, находки E. velox – действительно результат недавнего вселения или следствие недостаточной изученности биоразнообразия региона. Подобное предположение о находках E. velox в регионах, близких к Черному морю, высказано Самчишиной (2000).
В результате быстрого распространения E. velox сложно определить нативный ареал вида. Считается, что исходная область его распространения – места древних морских трансгрессий бассейнов-предшественников Черного и Каспийского морей (Сабанеев, 1930). Предполагается, что источником расселения E. velox стал именно Каспийский регион (Самчишина, 2000), где вид пережил плейстоценовое оледенение. Также высказано мнение, что существует и некий Балтийский рефугиум (Литориновое море), из которого произошла балтийская популяция (Ekmann, 1907). Именно из Балтийского региона (пролив Эресунн) и описан вид. Одно из первых упоминаний о E. velox дано в работе Sars (1897). Указано, что вид встречается в устье р. Нева, Финском и Ботническом заливах, на Британских островах, в Голландии, Франции, Германии и Норвегии в пресных и в солоноватых водах, здесь же впервые описана каспийская популяция E. velox из лагун Северного Каспия – устья р. Волга (Sars, 1897). По сообщению Spandl (1926), в начале ХХ в. вид присутствовал в некоторых пресных водоемах Европы, среди них указана р. Волга (до г. Казань). По крайней мере, с 1960-х годов вид отмечают как обычный в прибрежье Рыбинского вдхр. (Верхняя Волга) (Рыбинское…, 1972). В монографии “Волга и ее жизнь” (1978) он представлен как обычный уже для всей реки, в том числе для участка будущего Чебоксарского вдхр., заполненного в 1981 г., по данным Уломского, E. velox регистрируют в водохранилищах р. Кама с конца 1950-х гг. (Lazareva, 2020). Таким образом, вид стали регулярно обнаруживать в реках Волга и Кама в середине ХХ в., когда начал формироваться Волжско-Камский каскад водохранилищ и интенсифицировались гидробиологические исследования новых искусственных водоемов. В эстуариях рек, впадающих в Азовское море, вид отмечен с начала ХХ в. (Зернов, 1901). В бассейне Черного моря он впервые найден в нижнем течении р. Днепр (Сабанеев, 1930). В бассейне среднего течения р. Днепр в окрестностях г. Киев вид-вселенец впервые отмечен в середине прошлого века (Травянко, Цееб, 1967). На сегодняшний день в бассейнах Азовского и Черного морей E. velox встречается почти повсеместно (Литвинова, Вежновец, 2018; Samchishina et al., 2020).
Несмотря на активное расширение ареала в столь короткие сроки, полноценных генетических исследований E. velox до сих не проводили. В международной базе данных GenBank (https:// www.ncbi.nlm.nih.gov/nuccore/MT146445 Дата обращения 01.06.2021 г.) обнаружено всего пять нуклеотидных последовательностей ДНК вида из водоемов Польши. Последовательности значатся, как неопубликованные, но дано название статьи “First report to potentially invasive copepod Eurytemora velox (Lilljeborg, 1853) based on molecular characterization”. В описании указано, что рачки найдены в р. Одер и проливе Дзивна, впадающих в Балтийское море. В последние годы вид в небольших количествах также появился в эстуариях североморских рек Франции (Сена) и Бельгии (Шельда), а также в притоках р. Шельда, где доля вселенца достигает ≤97% численности всех эуритемор (личные сообщения проф. Sami Souissi и проф. Tackx Michèle). В связи с этим нами проведен предварительный молекулярно-генетический анализ имеющегося материала E. velox. Исследованы представители вида из Чебоксарского вдхр. (бассейн р. Волги, впадающей в Каспийское море) и озера в г. Киев (бассейн р. Днепр, впадающей в Черное море). Предполагают, что обе эти популяции являются вселенцами (Samchishina et al., 2020). Источником расселения для украинской популяции, вероятно, служит Понто-Каспийский регион (опресненные участки Черного моря) (Самчишина, 2000; Samchishina et al., 2020). Чебоксарская популяция, скорее всего, близка к таковой из Северного Каспия и сформировалась из особей, поднявшихся вверх по р. Волге от ее устья, где вид обитал еще в конце XIX в. (Sars, 1897).
Нами получены пять последовательностей участков генов митохондриальной цитохром оксидазы 1 (СО1) длиной 687 п.о. (две для чебоксарской и три для киевской популяций), шесть последовательностей участков ядерных генов ITS1-ITS2 длиной 772 п.о. (ITSs) (по три для каждой популяции) и две последовательности участка ядерного гена 18SrRNA длиной 656 п.о. (по одной для каждой популяции). Все последовательности загружены в международную базу данных GenBank под номерами: MZ400499–MZ400504; MZ373318–MZ373322; MZ373302. В статье впервые публикуются результаты генетического анализа E. velox. Секвенирование ДНК выполнено в Закрытом акционерном обществе “Евроген”. В ходе исследований использована федеральная коллекция Зоологического института РАН.
Анализ участков генов СО1 и ITSs показал, что популяции из озера в г. Киев и Чебоксарского водохранилища имеют небольшие генетические различия – 0.8 и 0.1% нуклеотидных замен соответственно. Мы предположили, что примерно такой же уровень различий обнаружится между черноморской и каспийской популяциями. Подобные уровни генетических различий наблюдаются между морфотипами E. affinis (Poppe, 1880): балтийским, североморским и восточно-атлантическим (Gesche et al., 2011; Sukhikh et al., 2016). Анализ двух последовательностей ДНК консервативного гена 18SrRNA не выявил различий между популяциями E. velox из киевского озера и Чебоксарского вдхр.
Сравнение полученных данных с двумя имеющимися в открытом доступе GenBank последовательностями участка гена СО1 из Польши (МТ146446 и МТ146445) показало, что они группируются с эуритеморами из Чебоксарского водохранилища (рис. 1).
Рис. 1.
Медианная сеть гаплотипов Eurytemora velox (Lilljeborg, 1853), построенная по семи нуклеотидным последовательностям участка гена СО1. Поперечные черточки – количество мутаций между гаплотипами. Черный – гаплотипы из Польши (MT146446 и MT146445), серый – из Чебоксарского вдхр. (718, 719), белый – из г. Киев (712, 714, 715).
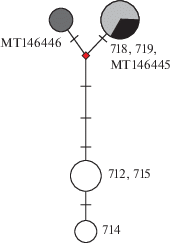
Выводы. Из-за близкого расположения Чебоксарского вдхр. к нативному местообитанию E. velox наиболее вероятно, что в водных объектах Польши обнаружена эуритемора, происходящая из Каспийского региона, а не наоборот. Популяция из озера в г. Киев отличается на несколько замен от каспийских и польских. Предположительно, это потомки особей, поднявшихся по р. Днепр из Черноморского региона, т.е. произошло независимое расселение вида, по крайней мере, из двух регионов-доноров. Дальнейшее исследование нативных популяций E. velox из Азово-Черноморского и Каспийского регионов, а также популяций вида из других регионов поможет более полно раскрыть картину происхождения вселенца и путей его расселения.
Список литературы
Боруцкий Е.В. Степанова Л.А., Кос М.С. 1991. Определитель Calanoida пресных вод СССР. Санкт-Петербург: Наука.
Волга и ее жизнь. 1978. Ленинград: Наука.
Зернов С.А. 1901. Планктон Азовского моря // Ежегодник Зоологического музея академии наук. Т. 6. С. 4.
Литвинова А.Г., Вежновец В.В. 2018. Распространение и показатели численности чужеродной каланоидной копеподы Eurytemora velox Lilljeborg, 1853 в Днепро-Бугском канале // Сахаровские чтения 2018 года: экологические проблемы XXI века: Матер. 18-й междунар. науч. конф. Минск, 17–18 мая 2018 г. Минск: Информационно-вычислительный центр Министерства финансов Республики Беларусь. С. 61.
Рыбинское водохранилище и его жизнь. 1972. Ленинград: Наука.
Сабанеев П.Л. 1930. Eurytemora velox (Lill.) у порожнистій частині Дніпра // Збір. Пр. Дніпр. біол. стан. 5.
Самчишина Л.В. 2000. Рецентное вселение олигогалиного вида Eurytemora velox (Lill.) (Copepoda, Calanoida) в Днепр и его крупные притоки // Экология моря. Вып. 52. С. 52.
Травянко В.С., Цееб Я.Я. 1967. Зоопланктон верхнего Днепра и водоeмов его поймы // Гидробиологический режим Днепра в условиях зарегулированного стока. Киев: Наукова думка. С. 74.
Ekmann S. 1907. Uber das Crustaceen plankton des Ekoln (Mälaren) und fiber verschiedene Kategorien von marinen Relikten in schwedischen Binnenseen // Zoologiska Studier.
Elton C. 1929. The Ecological Relationships of Certain Freshwater Copepods // Journal of Ecology. V. 17(2). P. 383. https://doi.org/10.2307/2256050
Gaviria S., Forro L. 2000. Morphological characterization of new populations of the copepod Eurytemora velox (Lilljeborg, 1853) (Calanoida, Temoridae) found in Austria and Hungary // Hydrobiologia. V. 438. P. 205. https://doi.org/10.1023/A:1004173704289
Lazareva V.I. 2020. Long-Term Changes in the Composition and Abundance of the Zooplankton Community in Kama River Reservoirs // Inland Water Biology. V. 13. № 2. P. 214. https://doi.org/10.1134/S199508292002025X
Samchyshyna L., Gromova Y., Zorina-Sakharova K. 2020. Recent distribution of Eurytemora velox (Lilljeborg, 1853) (Copepoda, Calanoida) in brackish and fresh waters of Ukraine // Crustaceana. V. 93. № 3-5. P. 275. https://doi.org/10.1163/15685403-00003992
Sars G.O. 1897. Temorella lacinulata (Fischer) // Ежегодник Зоологического музея Императорской академии наук. Т. 2. Санкт-Петербург. С. 66.
Spandl H. 1926. Copepoda. Ruderfusskrebse // P. Schulze Biologie der Tiere Deutschlands. V. 15. P. 1.
Sukhikh N.M., Souissi A., Souissi S. et al. 2016. Genetic and morphological heterogeneity among populations of Eurytemora affinis (Crustacea: Copepoda: Temoridae) in European waters // C.R. Biol. V. 339. P. 197. https://doi.org/10.1016/j.crvi.2016.03.004
Winkler G., Souissi S., Poux C., Castric V. 2011. Genetic heterogeneity among Eurytemora affinis populations in Western Europe // Mar. Biol. V. 158. Р. 1841. https://doi.org/10.1007/s00227-011-1696-5.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод