

Биология внутренних вод, 2022, № 4, стр. 342-365
Типология строения суспензориума костистых рыб (Teleostei) в связи с их питанием (обзор)
В. В. Махотин a, Е. С. Громова a, *
a Московский государственный университет
Москва, Россия
* E-mail: zhenya_s@inbox.ru
Поступила в редакцию 04.08.2021
После доработки 25.01.2022
Принята к публикации 11.02.2022
- EDN: HSVETY
- DOI: 10.31857/S0320965222040192
Аннотация
Приведен обзор особенностей устройства суспензориума у представителей Teleostei на дефинитивной стадии развития в зависимости от особенностей его функционирования при различных способах питания этой группы животных. В основе предлагаемой качественной классификации строения подвесочного аппарата рыб лежит укрепление или ослабление его конструкции. В обзоре дана характеристика разновидностей внутреннего кинетизма суспензориума видов Teleostei и описан механизм “внутренней аддукции”.
Устройство костномускульной системы черепа взаимосвязано с особенностями пищевого поведения Teleostei (Westneat, 1991, 1994), последнее имеет ведущее значение в формировании диеты (Wainwright, Richard, 1995). Авторы Wainwright (1991), Turingan (1994) придерживаются мнения, что функциональная морфология аппарата питания костистых рыб формирует характер их кормодобывания. В литературе исследования, описывающие вариацию строения составляющих черепа у широкого набора представителей, принадлежащих к различным таксонам Teleostei, в связи с действием этих структур в процессе питания и/или особенностями экологии добычи пищи, представляют собой нечастое явление (Wainwright, Richard, 1995; Westneat, 2004), особенно в отношении суспензориума. В публикациях подобной тематики для изучения обычно используют лишь выборочные таксоны Teleostei без сравнения с остальными представителями этой группы (Воскобойникова, 1985). Более распространены работы, в которых авторы приводят спектр данных о различиях морфологии устройства подразделений аппарата питания (но без суспензориума) ряда видов рыб без обсуждения функциональных причин обнаруженного разнообразия (Winterbottom, 1974a; Datovo, Vari, 2014; Кузищин и др., 2020). Суспензориум Teleostei представляет собой единую “костную щеку” рыбы и в основном cформирован элементами первичной верхней челюсти животного с включением некоторых покровных окостенений (Osse, 1969; Anker, 1974; Turingan, Wainwright, 1993). Часть составляющих его костей могут быть озублены (Sulak, 1977). В настоящей работе при описании суспензориума мы также будем использовать условный термин “подвесочный аппарат”. Впервые суспензориум появляется у новоперых рыб Neopterygii. Предпосылками для его формирования стали переориентация, укорочение и смещение вперед hyomandibulare относительно его положения, свойственного палеонискам Palaeonisciformes (Дзержинский, 2005). Как правило, подвесочный аппарат Teleostei, помимо сочленений с нейрокраниумом, формирует суставные поверхности еще: с жаберной крышкой, с гиоидом и с вторичной верхней и нижней челюстями. Количество исследований, описывающих морфологию суспензориума Teleostei, существенно превышает число публикаций, посвященных особенностям его действия (Ballintijn et al., 1972). В современной русскозычной литературе отсутствуют обзоры с анализом внутреннего кинетизма подвесочного аппарата у ряда видов Teleostei. Имеются отдельные исследования, дающие представления о подвижности составляющих суспензориума друг относительно друга лишь у одиночных таксонов указанной группы рыб (Liem, 1970; Ballintijn et al., 1972; Westneat, Wainwright, 1989 и др.).
Цель настоящего обзора – обобщение имеющейся информации по анатомии подвесочного аппарата у Teleostei и качественная оценка функционального значения выявленных особенностей спланхнокраниума, связанных с различными способами питания – всасыванием (Lauder, 1985), укусом (Barel, 1983) и тараном (Konow, Ferry-Graham, 2013) – у большого числа видов. Классификация вариантов взаимосвязи суспензориума с нейрокраниумом у Teleostei, разработанная ранее (Stensio, 1921, 1932; De Beer, 1937; Hofer, 1945; Дзержинский, 2005 и др), требует дополнения и обновления современными морфологическими данными, сведениями определяющими характер внутреннего кинетизма суспензориума у ряда видов костистых рыб.
При описании устройства черепа рассматриваются сочленения суспензориума с этмоидным, глазничным и слуховым отделами нейрокраниума. В работе использованы термины, применяемые в литературе при характеристике скелета (Osse, 1969; Гуртовой и др., 1976), соединительнотканных элементов (Lauder, 1980) и мышц Teleostei (Winterbottom, 1974a).
Общая характеристика конструкции суспензориума Teleostei
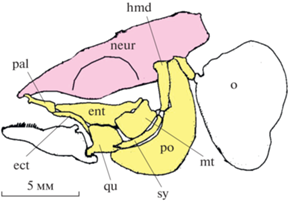
В суспензориуме большинства генерализованных форм Teleostei (таких как лососевые Salmonidae (Громова, Махотин, 2016), умбровые Umbridae (Wilson, Veilleux, 1982) (рис. 1), определенные сельдевые Clupeidae (Kirchhoff, 1958) самый ростральный элемент – palatinum, тогда как крыловидные кости (ectopterygoideum, entopterygoideum и metapterygoideum) занимают относительно него каудальную позицию (Kirchhoff, 1958; Osse, 1969; Wilson, Veilleux, 1982). Quadratum, находящийся в наиболее вентральной области “костной щеки” рыбы, участвует в формировании челюстного сустава сверху и сзади. У большинства форм Teleostei praeoperculum включено в состав суспензориума (Osse, 1969; Anker, 1974; Turingan, Wainwright, 1993), в связи с чем, смещения данной кости не зависят от движений жаберной крышки (Branch, 1966; Liem, 1970; Waltzek, Wainwright, 2003). Metapterygoideum своим задним краем контактирует с передней частью hyomandibulare, последняя, в свою очередь, соединена с ростральной кромкой prаeoperculum (Osse, 1969; Ballintijn et al., 1972). Обычно точная окклюзия элементов переднего отдела суспензориума и нижней челюсти у Teleostei отсутствует, за исключением лишь некоторых видов, таких как акантонус Acanthonus (Howes, 1992).
Рис. 1.
Генерализованный вариант устройства суспензориума умбры Umbra limi (по Wilson, Veilleux, 1982). ect – ectopterygoideum, ent – entopterygoideum, hmd – hyomandibulare, mt – metapterygoideum, neur – neurocranium, o – operculum, pal – palatinum, po – prаeoperculum, qu – quadratum, sy – symplecticum. Здесь и на рис. 2, 3, 5, 6, 9 масштаб на фрагментах рисунков взят из оригинальных публикаций, если авторы его указали.
У большинства Teleostei присутствует этмогиостилия: причленение (“подвеска”) суспензориума к слуховому отделу нейрокраниума и этмоиду (De Beer, 1937; Hofer, 1945; Дзержинский, 2005, 2013). У бóльшей доли представителей описываемой группы рыб в рамках этмогиостилии рalatinum – основной элемент, который выполняет крепление переднего конца суспензориума к ростральному (этмоидному) отделу нейрокраниума (Sulak, 1977; Olney et al., 1993; Konstantinidis, Johnson, 2016; Воскобойникова, Назаркин, 2017). У части видов в образовании этой связи также могут участвовать крыловидные кости (Grande, Poyato-Ariza, 2010), в частности у дистиходонтовых Distichodontidae (Vari, 1979), хоботнорыловых Mastacembelidae и чаудхуриевых Chaudhuriidae (Britz, 1996). У различных представителей Teleostei palatinum формирует одинарную, двойную или тройную связь с этмоидным отделом нейрокраниума. Наличие одного и того же числа точек причленения суспензориума к этмоидной части черепной коробки у примитивных и продвинутых видов Teleostei, по-видимому, является следствием различных факторов. Так, у некоторых таксонов группы присутствие единственного сустава palatinum с этмоидом может быть результатом слияния исходных двух подобных структур в качестве адаптации к противостоянию к повышенной нагрузке, прикладываемой к челюстному аппарату рыбы в процессе питания (Hofer, 1945). У других представителей потеря каудального из ростральных суставов суспензориума с черепной коробкой обусловлено чрезвычайным удлинением этмоидного отдела нейрокраниума (Norman, 1925). Кроме того, на развитие одного или двух указанных сочленений влияет положение нижней челюсти по отношению к черепной коробке. Морфологическое значение увеличения количества точек контакта palatinum с нейрокраниумом до конца не выяснено. Предположительно, это может обеспечивать рост способности переднего отдела черепа к поглощению давления, исходящего со стороны ротовой полости во время схватывания добычи и/или ее внутриротовой обработки, во избежание деформации или поломки скелетной конструкции головы (Hofer, 1945).
Среди Teleostei имеются примеры различных вариантов переориентации (относительно нейрокраниума и глазницы) суспензориума (рис. 2, рис. 3). Это изменение положения костей черепа животного может обусловить потерю прямого сочленения palatinum с нейрокраниумом. Так, у индоокеанского бомбиля Harpadon nehereus передний контакт суспензориума с черепной коробкой присутствует, однако, образован maxillare anterior (входит в состав суспензориума), с задним концом которого скреплена вышеупомянутая кость (рис. 3е) (Sulak, 1977). У фрактолема Анзорга Phractolaemus ansorgei palatinum не соединен с черепной коробкой, взамен имеется причленение к mesethmoideum посредством ectopterygoideum и quadratum (рис. 2е) (Grande, Poyato-Ariza, 2010). Переориентация подвесочного аппарата может проявляться в виде: 1) изменения соотношения величины и формы различных долей/отдельных костей суспензориума; 2) формирования в нем “окон”; 3) значительного увеличения или уменьшения его размера в целом; 4) изменения положения входящих в него элементов друг относительно друга; 5) изменения положения всего суспензориума на голове рыбы (рис. 2, рис. 3). Вариация морфологии подвесочного аппарата наблюдается на уровне таксонов Teleostei разного ранга. Значительная модификация конструкции суспензориума может быть связана со специализацией процесса кормодобывания у многих аностомовых Anostomidae, например, у сартора Sartor elongatus или петуланоса Petulanos plicatus (Sidlauskas, Vari, 2008), а также паразитических сомов ванделлии Vandellia и тридентопсиса Tridentopsis (Arratia, 1990). Вследствие формирования особого механизма выдвижения MJA у фрактолема Анзорга Phractolaemus ansorgei присутствует перестройка подвесочного аппарата, в связи с чем изменяется местоположение нескольких костей – palatinum, quadratum и ectopterygoideum (Grande, Poyato-Ariza, 2010).
Рис. 2.
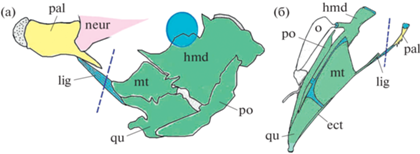
Разнообразие морфологии суспензориума у различных видов Teleostei: а – экстремальное видоизменение костей суспензориума у губана-обманщика Epibulus insidator (по: Westneat, Wainwright, 1989); б – крупный ростральный вырост hyomandibulare трихомиктериевого сома Trichomycterus sp. (по: Arratia, 1990); в – отсутствие entopterygoideum у тихоокеанской желеносой рыбы Ateleopus japonicus (по: Sasaki et al., 2006); г – огромное prаeoperculum малоротой макропинны Macropinna microstoma (по: Chapman, 1942); д – редукция entopterygoideum у вельветового сома Diplomystes (по: Fink, Fink, 1981); е – потеря symplecticum у фрактолема Анзорга Phractolaemus ansorgei (по: Grande, Poyato-Ariza, 2010); ж – слияние palatinum и ectopterygoideum в единый элемент у красивого щукоглава Luciocephalus pulcher (по: Liem, 1967). so – suboperculum, остальные обозначения, как на рис. 1. Целевые для просмотра элементы даны желтым. Здесь и на рис. 3, 5–7 голубой цвет — глазное яблоко ряда представителей Teleostei.

Рис. 3.
Окна (голубые) в конструкции суспензориума у (а) плоскоголового сома Pinirampus pirinampu (по: Howes, 1983b) и (б) обыкновенного илистого прыгуна Periophthalmus barbarus (по: Michel et al., 2014). Значительное удлинение переднего отдела суспензориума у (в) батизавра Bathysaurus (по: Sulak, 1977) и (г) трубкорыла Sternarchorhynchus chaoi (по: Santana, Vari, 2010), с увеличением степени наклона нижней ветви hyomandibulare в ростральном направлении у последнего. Удлинение суспензориума в направлении назад у (д) батимикропса Bathymicrops regis (по: Sulak, 1977) и (е) индоокеанского бомбиля Harpadon nehereus (по: Johnson et al., 1996). с pal – cartilago palatini, mx ant – maxillare anterior, остальные обозначения, как на рис. 1.

Таким образом, в рамках вариации морфологии подвесочного аппарата относительно его генерализованного устройства можно выделить две основные тенденции – укрепления и ослабления его структуры (рис. 4). Эти направления эволюционных преобразований находят проявление как в модификации конструкции самого суспензориума, так и в особенностях его контакта с черепной коробкой. Среди Teleostei, помимо видов со значительной степенью проявления ряда признаков этих двух крайних вышеупомянутых состояний, можно обнаружить широкий спектр представителей, чей подвесочный аппарат демонстрирует варианты промежуточной степени развития укрепления или ослабления его устройства.

Укрепление суспензориума
Укрепление суспензориума происходит путем увеличения внутренней прочности его конструкции и усиления фиксации суспензориума относительно нейрокраниума.
Причина наличия массивного суспензориума у определенных видов помацентровых Pomacentridae – поедание твердой пищи (Emery, 1973). Отсутствие внутренней подвижности характерно для прочно “скроенного” робустного суспензориума иглобрюховых Tetraodontidae (рис. 5а) и рыб-ежей Diodontidae, который способен выдерживать значительные нагрузки как при использовании дурофагии, так и при силовой прокачке воды посредством ротовой полости во время поведения “раздувания” (Winterbottom, 1974b; Wainwright et al., 1995).
Рис. 5.
Примеры укрепления суспензориума среди Teleostei: а – прочно “скроенный” суспензориум (круг – глазное яблоко) у острорылого иглобрюха Canthigaster (по: Tyler, 1980); б – увеличение роли суспензориума в формировании челюстного аппарата у макрорамфосодеса Macrorhamphosodes (по: Tyler, 1980); в – прирастание dermopalatinum перламутрового макрогнатуса Macrognathus pancalus (по: Britz, 1996) к этмоидного отделу нейрокраниума; г – робустное hyomandibulare утконосого угря Hoplunnis punctata (по: Eagderi, 2010). dpal – dermopalatinum, eth – ethmoideum, iop – interoperculum, mх – maxillare, pal-pt – palatopterygoideum, prmx – premaxillare, v – vomer. Остальные обозначения, как на рис. 1, 2. Целевые для просмотра элементы закрашены.

Увеличение прочности суспензориума у некоторых видов Teleostei достигается путем объединения его костей друг с другом. Так, у мормиропса Mormyrops deliciosus и камнеголова Petrocephalus bane entopterygoideum и ectopterygoideum сливаются между собой (Ridewood, 1904) для возможности применения крепкого укуса добычи. У части предствителей Teleostei имеет место увеличение размера костей “скелетной щеки” рыбы, поскольку основной вклад или его большую долю в укус добычи сверху осуществляет не вторичная, а первичная верхняя челюсть, т.е. суспензориум (Konstantinidis, Johnson, 2016). Доминирование роли первичной верхней челюсти в схватывании корма у Pisces обычно рассматривается в качестве примитивного признака (Дзержинский, 2005), если только это не представляет собой вторичное явление. Так, суспензориум играет ведущую роль в формировании челюстного аппарата в сравнении со сниженным значением MJA у макрорамфосодеса Macrorhamphosodes platycheilus (рис. 5б) (Tyler, 1980) и бородатого единорога Anacanthus barbatus (Winterbottom, 1974b). У индоокеанского бомбиля Harpadon nehereus (рис. 3е) и рыбы-ящерицы Saurida происходит включение части вторичной верхней челюсти в состав суспензориума. Их маленькое maxillare anterior неподвижно сочленяется с передним концом крупного palatinum, служа его озубленным продолжением и участвуя в процессе кусания добычи (Sulak, 1977).
У определенных видов Teleostei развивается увеличение прочности фиксации суспензориума относительно нейрокраниума. Эта модификация касается как передней (а), так и задней (б) областей их контакта:
а) У ряда форм впереди глазницы животного, каудальнее традиционной области контакта суспензориума с черепной коробкой посредством palatinum, развивается дополнительная точка крепления. Так, у бентопелагических особей тихоокеанской желеносой рыбы Ateleopus japonicus суспензориум формирует длинный, вертикально вверх ориентированный хрящевой отросток для прикрепления к латеральному предглазничному “крылу” нейрокраниума (рис. 2в) (Sasaki et al., 2006). У некоторых Teleostei элементы подвесочного аппарата включаются в состав черепной коробки. Так, у иглобрюховидных Tetraodontoidei в качестве адаптации к дурофагии palatinum объединяется с нейрокраниумом: у иглобрюховых Tetraodontidae оно прочно срастается с сошником и этмоидом с сохранением швов между тремя этими костями (Tyler, 1980). На palatinum располагается сочленовная фасетка для контакта с остальной долей суспензориума. У макрорамфосодеса Macrorhamphosodes platycheilus palatinum и ectopterygoideum неподвижно скреплены с vomer, таким образом, входя в состав нейрокраниума, в рамках адаптации к укреплению переднего отдела черепа – формированию монолитного длинного рыла. Протяженность последнего достигает до 49% длины тела рыбы (рис. 5б) (Tyler, 1980). По мнению некоторых авторов (Pietsch, 1978b), palatinum палочкохвоста Stylephorus chordatus неподвижно прирастает впереди к его черепной коробке. Dermopalatinum перламутрового макрогнатуса Macrognathus pancalus прочно скреплено с ростральным отделом нейрокраниума: оно соединяется швами с сошником и этмоидом (рис. 5в) (Britz, 1996).
б) Для кусания пищи у карповых Carapidae приспособлен мощный, с костными утолщениями hyomandibulare, который способен противостоять нагрузке, оказываемой на него со стороны сокращающихся порций АМ. Прочность причленения кости к нейрокраниуму достигается за счет ее большой ширины, которая превышает ее длину в вертикальной плоскости. В связи с этим, два сочлененных мыщелка hyomandibulare располагаются на значительном расстоянии друг от друга и на уровне переднего и заднего концов слухового отдела черепной коробки (Parmentier et al., 1998). Подобная конструкция данной кости также свойственна пегасу Pegasus: протяженность описываемого элемента суспензориума лишь ненамного меньше его высоты, а суставные головки hyomandibulare для присоединения к нейрокраниуму широко разнесены друг от друга (Pietsch, 1978a).
Потеря устойчивой передней точки причленения суспензориума к нейрокраниуму в результате обособления palatinum обеспечивает прочность сустава hyomandibulare с черепной коробкой у многих сомов Siluriformes (Adriaens, 2003). Последнее свойственно также угреобразным Anguilliformes. Так, у утконосых угрей Nettastomatidae ширина мощного робустного hyomandibulare – основного элемента, выполняющего крепление суспензориума к черепной коробке, – превосходит его длину (рис. 5г). Две сочленовные головки этой кости для причленения к нейрокраниуму разнесены далеко друг от друга и потому создают очень устойчивое и прочное соединение указанных частей черепа (Eagderi, 2010). Упрощение конструкции суспензориума за счет уменьшения числа составляющих его элементов является адаптацией к высоким нагрузкам на его поверхность, возникающих в ходе рытья в грунте и крепкого укуса добычи. Каудальная подвеска суспензориума позволяет освободить в голове рыбы пространство для размещения и увеличения объема в ходе функционирования гипертрофированного АМ. Так, суспензориум грязевого угря Pythonichthys macrurus фактически включает только две кости, составляющие монолитный робустный костный блок: мощное hyomandibulare, соединенное неподвижным швом с quadratum (Eagderi, 2010). Гипертрофия широкого робустного hyomandibulare (ширина кости равна ¾ ее длины) анабаса Anabas testudineus с чрезвычайным развитием верхнего отдела данной кости (Dutta, 1975), по-видимому, связана с присутствием у этой рыбы наземной локомоции. Жаберные крышки животного принимают активное участие в опоре на субстрат и крепятся к hyomandibulare. Поэтому на суше последняя кость анабаса фактически принимает на себя всю массу тела рыбы, в связи с чем ей требуется надежная поддержка на нейрокраниуме.
Ослабление суспензориума: области его внутренней подвижности
У части видов Teleostei в рамках суспензориума формируются области внутренней подвижности. Это происходит из-за конструкций, находящихся между костями: 1) областей, заполненных хрящевой и/или соединительной тканью; 2) суставов или 3) связок (Gosline, 1951; Winterbottom, 1974b). В отдельных случаях, происходит значительное пространственное отделение элементов суспензориума друг от друга (Howes, Fumihito, 1991). Формирование вышеуказанных особенностей конструкции “костной щеки” дает в результате ослабленное соединение компонентов подвесочного аппарата. Появлению внутреннего кинетизма (см. далее) также способствует наложение одних элементов суспензориума на другие и расположение составляющих его скелетных единиц в разных плоскостях.
1) Наиболее распространенным из вышеперечисленных вариантов соединения отдельных элементов в рамках обладающего внутренней подвижностью суспензориума является наличие в его пределах гнущихся хрящевых областей в составе синхондрозов (Liem, 1970; Vial, Ojeda, 1992). Так, увеличение протяженности суспензориума гоноринха Gonorynchus greyi в продольном направлении служит причиной потери непосредственного контакта составляющих его костей и, вследствие этого, возможности их смещения друг относительно друга из-за широких областей между ними, заполненных хрящевой тканью (рис. 6а) (Grande, Poyato-Ariza, 2010). Palatinum барабулевых Mullidae (Gosline, 1984) соединено с entopterygoideum гнущейся зоной хряща.
Рис. 6.
Зоны подвижности в пределах суспензориума у Teleostei: а – хрящевые области у гоноринха Gonorynchus greyi (по: Grande, 1999); б – высокоподвижные суставы у обыкновенной длиннорылой рыбы-бабочки Forcipiger longirostris (по: Konow, Ferry-Graham, 2013); в – связки у сома-ванделлии Vandellia cirrhosa (по: Arratia, 1990). Области внутренней подвижности между передним, срединным и задним отделами суспензориума (закрашены желтым, зеленым или голубым) показаны синей пунктирной линией. lig – ligamentum, остальные обозначения, как на рис. 1, 5. Здесь и на рис. 7, 9 хрящ обозначен точками и связки черточками на голубом фоне.

2) Высокоподвижные суставы, приближающиеся по своему строению к шаровидным, развиваются между элементами суспензориума лишь у немногих представителей Teleostei, в частности у губана-обманщика Epibulus (Delsman, 1925), пескарки Callionymus (Kayser, 1962) и сома-коряжки Bunocephalus (Shafland, 1968). Наличие хорошо дифференцированных суставов обеспечивают присутствие внутреннего гиперкинетизма суспензориума у губана-обманщика Epibulus insidator; при этом монолитная структура его подвесочного аппарата почти целиком теряется (рис. 2а) (Delsman, 1925). У длиннорылых видов рыб-бабочек Chaetodontidae приобретение суставов внутри суспензориума увеличивает направленное вперед выдвижение верхней и нижней челюстей (рис. 6б) (Konow, Ferry-Graham, 2013). Так, слабое сочленение palatinum c остальным суспензориумом присутствует у просяной рыбы-бабочки Chaetodon miliaris (Motta, 1982).
3) У части представителей Teleostei кости в суспензориуме могут быть скреплены связками с отсутствием швов. Так, у коричневого илиофа Ilyophis brunneus тонкое ectopterygoideum соединено с quadratum посредством lig. pterygoideo-quadratum (Eagderi et al., 2016). У некоторых сомов Siluriformes, в частности у лягушкоротого сома Chaca еntopterygoideum связкой скреплено с metapterygoideum, либо с metapterygoideum и hyomandibulare, в зависимости от вида рода (Brown, Ferraris, 1988). У кольчужных Loricariidae и астроблеповых Astroblepidae сомов, а также паразитических сомов охмакантуса Ochmacanthus и ванделлии Vandellia metapterygoideum и quadratum соединены связкой и небольшим количеством хряща (рис. 6в) (Arratia, 1990). У удильщика Brachionichthys hirsutus palatinum и ectopterygoideum разнесены друг от друга и связаны лишь посредством тяжа соединительной ткани (Pietsch, 1981).
4) Некоторые авторы описывают у определенных видов Teleostei изоляцию (полное отделение) palatinum от суспензориума без костной связи между ними. Как правило, при этом между данной костью и каудальным отделом подвесочного аппарата все же сохраняется контакт посредством соединительной ткани. В случае каждого конкретного вида Teleostei критерии правомерности выделения этого варианта конструкции – оценка протяженности свободного промежутка между palatinum и задней частью суспензориума – определены исследователями по их собственному усмотрению (Winterbottom, 1974b; Gosline, 1975; Tyler, 1980; Fink, 1985; Adriaens, 2003; Diogo, Vandewalle, 2003; Huysentruyt, Adriaens, 2005).
В качестве наиболее характерного примера указанного типа строения подвесочного аппарата можно указать небно-максиллярный механизм для управления максиллярным усиком у сомообразных Siluriformes (рис. 7а) (Gosline, 1975; Howes, Fumihito, 1991; Adriaens, 2003; Huysentruyt, Adriaens, 2005). В ходе его образования palatinum переходит к функционированию в составе MJA, а суспензориум утрачивает свое традиционное ростральное место причленения к нейрокраниуму (Gosline, 1975). У определенных продвинутых таксонов группы, например перистоусых Mochokidae (Arratia, 1992) и панцирниковых Callichthyidae (Huysentruyt, Adriaens, 2005) сомов, обособленное palatinum формирует самостоятельный, но слабый сустав с ectoethmoideum (Sarmento-Soares, Porto, 2006). У более примитивных сомообразных palatinum прилегает к ectoethmoideum. Способ прикрепления к этмоиду определяет характер подвижности palatinum относительно черепной коробки: кость может выполнять переднезадние смещения (“скольжение”) или повороты (передним концом латерально, а задним медиально) (Gosline, 1975). Как правило, palatinum сомов Siluriformes контактирует длинным тяжем соединительной ткани/связкой с каудальным отделом суспензориума; кроме того, часто подобные мягкие, протяженные массивные структуры соединяют крыловидные кости с этмоидом (Gosline, 1975; Arratia, 1990, 1992; Diogo, Chardon, 1998; Huysentruyt, Adriaens, 2005). Так, у перистоусых сомов Mochokidae palatinum соединено длинной связкой с metapterygoideum, принадлежащим основной доле суспензориума. У большинства видов данного семейства в составе этой связки присутствует сильно редуцированный передний птеригоидный элемент (Arratia, 1992). Следует отметить, что у некоторых специализированных сомов, таких как амфилиевые Amphiliidae и многие багариеподобные сомы Sisoroidea, palatinum сочленяется лишь с нейрокраниумом и больше не соединяется связкой с остальным суспензориумом. Следовательно, эта кость приобретает еще большую свободу самостоятельных движений (Diogo, Vandewalle, 2003). У единороговых Monacanthidae рудиментарное palatinum отделяется от суспензориума, утрачивает cвое каудальное прочное скрепление с его костями и становится частью вторичной верхней челюсти – монолита maxillare и premaxillare, перемещаясь вместе с последним. Palatinum cоединен с передним отделом суспензориума либо при помощи связки, либо подвижным, обособленным кусочком хряща. Ростральный конец суспензориума самостоятельно причленяется к парасфеноиду посредством скользящего сочленения entopterygoideum и/или ectopterygoideum. Основную поддержку и контроль подвижности переднего отдела суспензориума относительно нейрокраниума осуществляют ААР и RAP (Winterbottom, 1974а; Tyler, 1980). Обособленное от остальной части суспензориума palatinum, с которой оно связано лишь соединительной тканью, свойственно таким продвинутым представителям стомиеобразных Stomiiformes, как аристомиас Aristostomias, малакост Malacosteus, пахистомиас Pachystomias и фотостомиас Photostomias (рис. 7б). У эустомиаса Eustomias изолированный от каудального отдела суспензориума скелетный сегмент формируют ectopterygoideum и palatinum. Этот ростральный фрагмент “костной щеки” животного контактирует с ее задней частью лишь связкой и перемещается вместе с maxillare, а не с остальной долей суспензориума (Fink, 1985). Наличие внутренней подвижности в пределах подвесочного аппарата у вышеуказанных форм помогает им в процессе проглатывания сверхкрупной добычи увеличивать пространство своей ротовой полости с последующим упругим возвращением ее в исходное (нерастянутое) положение (подобие механизма кинетизма черепа некоторых змей Serpentes (Дзержинский, 2005)). Также обширное отделение palatinum от крыловидных костей суспензориума свойственно некоторым производным видам губановых Labridae, в частности центролабрусу Centrolabrus (Westneat, 1993).
Рис. 7.
Изоляция palatinum от каудальной доли суспензориума: а – астроблепового сома Astroblepus (в рамках небно-максиллярного механизма) (по Arratia, 1990); б – фотостомии Photostomias guernei (по: Fink, 1985). Области внутренней подвижности суспензориума показаны синей пунктирной линией. Обозначения, как на рис. 1, 6.
У видов Teleostei со значительно обособленным подвижным palatinum данная кость может приобретать самостоятельное мускульное обслуживание. Так, у обыкновенного илистого прыгуна Periophthalmus barbarus для этой цели дифференцируется отдельное подразделение ААР (Michel et al., 2014). Изолированным palatinum сомообразных Siluriformes управляет m. extensor tentaculi, который является подразделением ААР: его сокращение вызывает протракцию максиллярного усика (Gosline, 1975; Adriaens, 2003). У продвинутых представителей отряда, таких как анциструс Ancistrus triradiatus, присутствуют два m. extensor tentaculi – латеральный и медиальный (Geerinckx et al., 2007).
Снижение прочности соединения суспензориума с нейрокраниумом
У определенных форм Teleostei области контакта суспензориума с черепной коробкой отличаются слабой структурой, с отсутствием выраженных мыщелков, что обеспечивает наличие более значительной степени подвижности того или иного отдела суспензориума относительно нейрокраниума. При этом прикрепление элементов суспензориума к черепной коробке осуществляется при помощи: соединительной ткани (Liem, 1970; Gosline, 1975; Howes, 1983b; Albert, Crampton, 2009), плоского сустава (Громова, Махотин, 2020) или вовсе отсутствует (Regan, 1924; Pietsch, 1978b; Eagderi, 2010). У большинства видов присутствует ослабление контакта переднего, а не заднего, отдела подвесочного аппарата с нейрокраниумом (Konow, Ferry-Graham, 2013). Так, ослабленное присоединение редуцированной ростральной части суспензориума к черепной коробке имеется у многих сомов Siluriformes. Например, миниатюрный entopterygoideum плоскоголового сома Pinirampus pirinampu находится внутри связки, прикрепляющейся к ectoethmoideum, тогда как крошечный ectopterygoideum включен в состав второй связки, оканчивающейся на palatinum (Howes, 1983b). У коричневого илиофа Ilyophis brunneus lig. pterygoideo-ethmoideum соединяет тоненький ectopterygoideum с этмоидным отделом нейрокраниума (Eagderi et al., 2016). Palatinum контактирует с черепной коробкой при помощи связки, без образования сустава у нандовых Nandidae, например, у рыбы-лист Monocirrhus polyacanthus (Liem, 1970), а также у некоторых длиннорылых видов рыб-бабочек Chaetodontidae (Konow, Ferry-Graham, 2013).
Описываемое ослабление области контакта суспензориума с нейрокраниумом может формироваться по нескольким причинам. Подобные особенности конструкции черепа развиваются как у представителей с отсутствием силового укуса жертвы, так и у хищников, заглатывающих чрезвычайно крупную добычу. Данные черты анатомии позволяют определенным рыбам-бабочкам Chaetodontidae использовать их подвижные передние челюсти для сбора корма с разнообразного и сложного по рельефу субстрата (Konow, Ferry-Graham, 2013). Финк (Fink, 1985) сообщает о наличии лишь слабого контакта миниатюрного palatinum с этмоидным отделом нейрокраниума у многих представителей стомиеобразных Stomiiformes, что вероятно помогает им заглатывать сверхкрупную добычу (см. выше).
Классификация вариантов соединения суспензориума с черепной коробкой у Teleostei
В течение многих лет исследователей интересовал вопрос взаимосвязи челюстного аппарата с нейрокраниумом у различных групп позвоночных Vertebrata (Goodrich, 1930; Clack, 1992; Depew, Simpson, 2006), в том числе у рыб Pisces (Gregory, 1904; Stensio, 1921; Moy-Thomas, 1939; Wilga, Ferry, 2015). Среди наиболее значимых русскоязычных работ недавнего времени можно отметить работы Ф.Я. Дзержинского (Дзержинский, 2005; Дзержинский и др., 2013), а также переводные руководства – Т. Ромера, Т. Парсонса (1992) и Р. Кэрролла (1992). На основании изучения ряда источников литературы, рассмотренных (см. выше) в сочетании с собственными экспериментальными данными (Gromova, Makhotin, 2018 и др.; Махотин, Громова, 2019; Громова, Махотин, 2020), в данной работе описаны варианты соединения суспензориума с черепной коробкой у представителей Teleostei. Однa из немногочисленных работ, посвященных сравнительному анализу строения “подвески” суспензориума к нейрокраниуму у некоторых видов данной группы – это исследование Хофера (Hofer, 1945). Однако, на настоящий момент оно недостаточно полно, содержит определенные ошибки и неточности. Ниже авторы приводят ряд дополнительных сведений по этой теме и дают скорректированную версию классификации вариантов контакта подвесочного аппарата с черепной коробкой, встречающихся среди Teleostei.
Дебир (De Beer, 1937) полагал, что исходно у примитивных Teleostei небноквадратный хрящ соединяется с нейрокраниумом в четырех точках. Сочленения образовывали hyomandibulare, ростральный конец небного отдела суспензориума, а также пара отростков (pr. basalis и pr. oticus), контактирующих с заглазничной областью черепной коробки (рис. 8). В ходе эволюции группы широкое распространение получило более продвинутое состояние – этмогиостилия, в результате развития которой происходила редукция processus basalis и processus oticus. Некоторые авторы (Stensio, 1932) полагают, что на месте последнего у современных Teleostei окостеневает metapterygoideum, в связи с чем разновидности pr. metapterygoideus рассматривают в качестве рудиментов pr. oticus. Одной из причин формирования этмогиостилии было пространственное смещение ротовой полости в голове рыбы в ростральном направлении, в сравнении с ее каудальным положением, присутствующим у палеонисков Palaeonisciformes (Hofer, 1945). Этот процесс потребовал развития наружного каркаса переднего отдела ротовой полости. У исследованных нами рыб – семги Salmo salar (Громова, Махотин, 2016), белого толстолобика Hypophthalmichthys molitrix (Махотин, Громова, 2019) и рубиново-пепельной рыбы-попугая Scarus rubroviolaceus (Громова, Махотин, 2020) – характер подвески суспензориума к нейрокраниуму можно описать как этмогиостилию. Между представителями Teleostei с крайними проявлениями сильной и слабой степени поддержки точек контакта суспензориума с нейрокраниумом в рамках этмогиостилии существует ряд многочисленных промежуточных морфологических форм. Следует отметить, что помимо этмогиостилии, формирование иных вариантов подвески суспензориума к черепной коробке (гиостилии, этмостилии, орбитоэтмогиостилии и аутоэтмогиостилии, см. далее) – это результат специализации аппарата питания обладающих им представителей Teleostei.
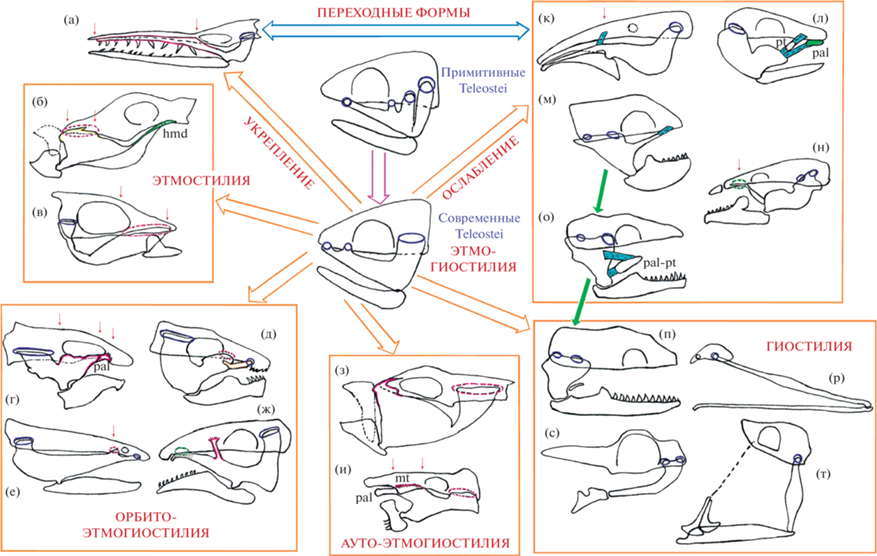
Рис. 8.
Схема типов контакта (“подвески”) суспензориума с нейрокраниумом у Teleostei. а – южноокеанский кинжалозуб Anotopterus vorax (по: Воскобойникова, Назаркин, 2017); б – карибский пуховик Canthigaster rostrata (по: Tyler, 1980); в – малоротая макропинна Macropinna microstoma (по: Chapman, 1942); г – большеголовый угорь Macrocephenchelys brachialis (по: Robins, Robins, 1971); д – угрехвостый сом Plotosus lineatus (по: Oliveira et al., 2001); е – темный болотный угорь Оphisternon aenigmaticum (по: Britz, 1996); ж – аптеронотус Бонапарта Apteronotus bonapartii (по: Hilton, Fernandes, 2017); з – рыбы-ежи Diodontidae (по: Tyler, 1980); и – анциструс Ancistrus triradiatus (по: Howes, 1983a; Arratia, 1990); к – трубкорыл Sternarchorhynchus (по: Santana, Vari, 2010); л – сом Diplomystes chilensis; м – курносый угорь Simenchelys parasitica (по: Eagderi et al., 2016); н – нагаихтис Nagaichthys filipes (по: Britz, Kottelat, 2003); о – грязевой угорь Pythonichthys macrurus (по: Eagderi, 2010); п – анархиас Anarchias allardicei (по: Mehta, Wainwright, 2007); р – большерот Eurypharynx pelicanoides (по: Eagderi, 2010); с – пегас Pagasus (по: Pietsch, 1978a); т – палочкохвост Stylephorus chordatus (по: Pietsch, 1978b). Усиление областей контакта суспензориума с нейрокраниумом: а – укрепленная этмогиостилия; б, в – этмостилия; г, ж – орбитоэтмогиостилия; з, и – аутоэтмогиостилия. Ослабление точек контакта суспензориума с нейрокраниумом: к, о – ослабленная этмогиостилия; п, р, с, т – гиостилия. Синие круги и овалы — подвижные сочленовные мыщелки/поверхности для контакта суспензориума с нейрокраниумом умеренной устойчивости; розовый цвет – скелетные элементы и/или области малоподвижного контакта суспензориума с нейрокраниумом, зеленый – скелетные элементы и/или области слабого контакта суспензориума с нейрокраниумом. Красные вертикальные стрелки указывают на важные области присоединения переднего отдела первичной верхней челюсти к черепной коробке, а также их протяженность в переднезаднем направлении, салатовые стрелки – вероятные анатомические стадии развития гиостилии в группе угрей Anguilliformes. Связки окрашены голубым цветом.
Тенденция к образованию гиостилии (рис. 8)
У части видов Teleostei в рамках этмогиостилии наблюдается ослабление передней области крепления суспензориума к нейрокраниуму, приводящее к формированию “функциональной гиостилии” (рис. 8к, 8л). В своей максимальной степени развития эта тенденция, вероятно, ведет к развитию классической (морфологической) гиостилии – подвеске челюстей посредством одного hyomandibulare, поскольку причленение небного отдела суспензориума к нейрокраниуму подвергается редукции. Среди спектра представителей группы можно найти примеры, представляющие собой разные стадии на пути формирования выше указанной конструкции.
Так, у некоторых представителей Teleostei передний отдел суспензориума, как и его точка прикрепления к черепной коробке, хоть и присутствуют, но подвергаются значительной редукции, как это имеет место у рыбы-ножа Orthosternarchus tamandua. У этого вида ростральный конец суспензориума представлен лишь тонким и грацильным стебельком небного хряща, который контактирует с этмоидом (Hilton, 2007). По-видимому, сужение и сильная редукция переднего отдела суспензориума (исчезновение palatinum и ectopterygoideum) вызвана необходимостью сокращения площади поперечного сечения начального отдела рыла с целью формирования удлиненного, конусообразного черепа для использования механизма всасывания корма по типу “пипетки” или “груши” (Marrero, Winemillar, 1993; Hilton, Fernandes, 2017). Подобная тенденция достигает еще большей степени проявления у трубкорылых сестринских представителей родов стернархоринхус Sternarchorhynchus и платиростернархус Platyurosternarchus в чрезвычайном удлинении и сужении entopterygoideum, длина которого превышает длину остальной части суспензориума, без какой-либо выраженной фасетки для контакта с нейрокраниумом (Santana, Vari, 2010). У этих двух видов трубкорылов суспензориум прикреплен к нейрокраниуму лишь в области глазницы при помощи lig. pterygo-craniale, связывающего entopterygoideum (либо entopterygoideum и quadratum) с orbitosphenoideum, тогда как более ростральная точка контакта суспензориума с черепной коробкой теряется (рис. 8к) (Santana, Vari, 2010). Задний птеригоид малайского зубастого сома Belodontichthys dinema соединен с передней частью черепной коробки лишь при помощи тонкой связки, включающей в себя крошечный передний птеригоид. В связи с этим суспензориум фактически утрачивает область прочной фиксации своей передней доли относительно рострального отдела нейрокраниума (Howes, Fumihito, 1991).
Среди угреобразных Anguilliformes обнаружены примеры “постепенного перехода” от этмогиостилии к классической гиостилии по мере роста силы укуса добычи и необходимости увеличения прочности конструкции суспензориума.
Так, у садового угря Гилберта Ariosoma gilberti присутствует этмогиостилия (Eagderi, 2010).
Далее, суспензориум курносого угря Simenchelys parasitica (рис. 8м) имеет упрощенное строение, консолидирован и уже утрачивает свой передний отдел. Вместо отсутствующих palatinum, metapterygoideum, symplecticum и entopterygoideum, тонкий костный стебелек palatopterygoideum соединен посредством узкого тяжа соединительной ткани с этмоидной частью черепной коробки. Меньшее количество составных элементов в суспензориуме снижает вероятность их смещения друг относительно друга под воздействием высокой нагрузки, которая возникает при силовом укусе добычи. Область контакта hyomandibulare с нейрокраниумом у курносого угря малоподвижна и обладает повышенной прочностью за счет значительного увеличения протяженности этой зоны соединения, а также широкого разнесения между собой двух сочленовных головок hyomandibulare. Редукция передних костей суспензориума связана с необходимостью увеличения свободного пространства для размещения гипертрофированной челюстной мускулатуры, прежде всего разросшегося АМ (Eagderi et al., 2016).
Подобная тенденция находит большее развитие в случае грязевого угря Pythonichthys macrurus (рис. 8о): у данной формы маленький рalatopterygoideum лишь при помощи двух связок подвижно соединен с крупным робустным задним отделом суспензориума – монолитным блоком скрепленных прочным швом hyomandibulare и quadratum. Таким образом, у грязевого угря имеется потеря скелетного контакта передней части суспензориума с черепной коробкой (Eagderi, 2010). То же состояние ковергентно развивается у продвинутых видов чаудхуриевых Chaudhuriidae, например нагаихтиса Nagaichthys filipes (рис. 8н), который придерживается схожего способа кормодобывания (Britz, Kottelat, 2003).
И наконец, пример классической гиостилии наблюдается среди мурен Muraenidae – анархиаса Anarchias allardicei (рис. 8п) и гимноторакса Gymnothorax prasinus. У этих представителей palatopterygoideum почти полностью исчезает и сохраняется лишь в виде тонкой редуцированной косточки, лежащей у переднего края огромного hyomandibulare, широко причленяющегося к черепной коробке (Mehta, Wainwright, 2007). В связи со значительным уменьшением размеров quadratum и praeoperculum hyomandibulare является, по сути, единственным хорошо развитым элементом в суспензориуме. Подобное устройство суспензориума также свойственно орегонскому слитножабернику Synaphobranchus oregoni (Robins, 1971). Формирование специализированной скелетной конструкции черепа у мурен обусловлено наличием уникальной системы фарингеального транспорта добычи к пищеводу при помощи перехватывающего ее укуса аппарата глоточных челюстей, а также вентрокаудальной ориентацией крупной порции А3 AM (Eagderi, 2010).
Причиной тенденции к развитию гиостилического варианта конструкции суспензориума может выступать необходимость увеличения амплитуды медиолатеральных смещений суспензориума с целью более эффективного изменения объема ротовой полости при всасывании и/или использовании внутреннего гидравлического механизма (см. ниже). При этом подвижная костная составляющая переднего отдела суспензориума сокращает свой размер (редуцируется). Возникающий благодаря этому рост соединительнотканного компонента в области прикрепления к нейрокраниуму рострального отдела суспензориума рыбы обеспечивает возможность отклонений данного, по сути, слабо зафиксированного относительно черепной коробки костного подразделения. Сустав hyomandibulare с нейрокраниумом, наоборот, усилен. Так, у бычка кремерии Kraemeria лишь тонкий стерженек palatinum + pterygoideum связывает с этмоидом крупный каудальный блок костей суспензориума (Gosline, 1955).
Гиостилическая “подвеска” (контакт суспензориума с нейрокраниумом только посредством hyomandibulare) суспензориума к нейрокраниуму в классическом варианте формируется у палочкохвоста Stylephorus chordatus (рис. 8т). У этого вида передняя доля суспензориума редуцируется, а palatinum включается в состав черепной коробки. Hyomandibulare сочленяется с нейрокраниумом лишь одной головкой. Благодаря такому варианту строения узкий суспензориум рыбы приобретает значительную свободу движений относительно черепной коробки. Фактически суспензориум представляет собой костный “стержень”, включающий сильно редуцированные элементы, вследствие утраты его роли в увеличении в латеральном направлении объема ротовой полости. Расширение последней происходит в результате растяжения соединительной ткани ее боковых стенок (Regan, 1924; Pietsch, 1978b). Некоторые авторы предполагают также наличие области внутренней подвижности в суспензориуме данной рыбы (Olney et al., 1993). Следует отметить, что гиостилия палочкохвоста – это результат специализации его аппарата питания вследствие использования уникального механизма кормодобывания – высокообъемного всасывания, (Pietsch, 1978a) и представляет собой вторичное явление.
По данным Нельсона (Nelson, 2006), мешкоротовые Saccopharyngidae – это наиболее анатомически видоизмененныe представители из всех видов позвоночных Vertebrata. Причина формирования гиостилии у мешкорота Saccopharynx ampullaceus и большерота Eurypharynx pelicanoides (рис. 8р) также может быть связана с особым способом питания – “заглатыванием”, при котором происходит сильное растяжение боковых стенок ротовой полости (Tchernavin, 1947; Nielsen et al., 1989; Eagderi, 2010). Гиостилия у этих форм дает возможность выполнять чрезвычайно высоко амплитудное расширение внутреннего пространства головы рыбы. По своей морфологии суспензориум мешкорота и большерота напоминает таковой палочкохвоста: представляет собой очень длинный (в сравнении с величиной нейрокраниума) грацильный костный “стержень”, причленяющийся одной сочленовной головкой к черепной коробке. Задний конец суспензориума в области челюстного сустава расположен на уровне первого десятка позвонков. Передний отдел суспензориума утрачен (Eagderi, 2010). Некоторые авторы предполагают, что в хрящевых областях центральной области суспензориума большерота (Bertin, 1934; Nielsen et al., 1989) и мешкорота (Tchernavin, 1947) присутствует значительная степень подвижности, благодаря которой имеется возможность сгибов его поверхности, необходимая для увеличения способности расширения пространства ротовой полости этих специализированных форм.
Формирование гиостилии в той или иной степени сопровождается упрощением конструкции суспензориума путем потери по крайней мере нескольких составляющих его окостенений. У некоторых представителей, в частности пегаса Pegasus (рис. 8с), при этом исчезают более половины костей суспензориума – palatinum, ectopterygoideum, entopterygoideum и metapterygoideum (Pietsch, 1978a). За счет этого достигается увеличение прочности конструкции аппарата питания, которая необходима для использования тарана челюстей: поддержания возможности эффективной протракции гипертрофированного максиллярного механизма (Дзержинский, 2005).
Таким образом, конвергентное устройство черепа (гиостилия) развивается у представителей трех неродственных таксонов (угреобразных Anguilliformes, опахообразных Lampriformes и колюшкообразных Gasterosteiformes), использующих разные способы начального захвата добычи – всасывание, заглатывание и таран челюстей (Pietsch, 1978a; Lauder, Liem, 1981; Longo et al., 2016).
Укрепление “подвески” суспензориума (рис. 8)
Уменьшение степени подвижности между нейрокраниумом и суспензориумом в рамках этмогиостилии часто происходит при повышенных нагрузках на последний. Это может иметь место, например, у тех видов Teleostei, которые потребляют пищу с твердыми покровами или измельчают ее в ходе внутриротовой обработки, а также у некоторых рыбоядных хищников. Так, протяженность зоны соединения огромного робустного palatinum к длинному предглазничному отделу черепной коробки у южноокеанского кинжалозуба Anotopterus vorax по площади в >5 раз превышает участок контакта hyomandibulare с нейрокраниумом у данной рыбы (рис. 8а) (Воскобойникова, Назаркин, 2017). Описываемый вариант подвески суспензориума свидетельствует, что у этой формы именно передний, а не задний, отдел черепа принимает на себя основную нагрузку во время укусов добычи.
Этмостилия. У некоторых специализированных видов Teleostei возможен следующий вариант изменения исходной этмогиостилической конструкции – значительное доминирование передней (этмоидной), а не каудальной, области подвески суспензориума. Подобное имеет место у карибского пуховика Canthigaster rostrata (рис. 8б) и паучеглазого иглобрюха Canthigaster amboinensis, у которых крупное робустное palatinum чрезвычайно широко и прочно прикрепляется к сошнику, тогда как маленькое, сильно редуцированное hyomandibulare фактически утрачивает значимую роль в соединении с нейрокраниумом (Tyler, 1980). Мы обозначили данное состояние в качестве “этмостилии” (доминирование широкой передней малоподвижной зоны крепления первичной верхней челюсти). По-видимому, причиной его формирования является наличие повышенных нагрузок, прилагаемых к переднему отделу суспензориума рыбы во время использования разгрызающего укуса (Turingan, 1994; Mehta, 2009) в рамках дурофагии. Также усиление прочности малоподвижной обширной области прикрепления робустной ростральной доли суспензориума к этмоидной части нейрокраниума свойственно малоротой макропинне Macropinna microstoma (рис. 8в) (Chapman, 1942).
Орбитоэтмогиостилия. У хищных ножетелок Magosternarchus sp. и Sternarchella orthos, использующих силовой укус добычи, помимо традиционного причленения переднего конца суспензориума к этмоиду, возникает дополнительное малоподвижное место крепления – entopterygoideum посредством костного отростка к frontale в районе глазницы (Lundberg et al., 1996). Фактически, на примере этих представителей прослеживается тенденция к формированию среди Teleostei орбитоэтмогиостилии (развитие третьей дополнительной точки крепления суспензориума к орбитальному отделу нейрокраниума). У серебряной араваны Osteoglossum bicirrhosum также присутствует ее разновидность: наличие базиптеригоидного сустава у этого вида повышает устойчивость его суспензориума относительно нейрокраниума (Громова и др., 2017), что необходимо для выполнения внутриротовой обработки добычи – скобления (raking) (Sanford, Lauder, 1989). В более специализованном состоянии на пути формирования данного варианта конструкции черепа (орбитоэтмогиостилии), в частности, у ножетелки аптеронотуса Бонапарта Apteronotus bonapartii (рис. 8ж), в образовании областей контакта суспензориума с нейрокраниумом роль наиболее ростральной точки их соединения снижается в пользу усиления развития глазничной области “подвески” – отростка entopterygoideum к orbitosphenoideum. Ростральный конец суспензориума редуцирован в тонкий хрящевой стебелек pars autopalatinum, соприкасающийся с ethmoideum ventralis (Hilton, Fernandes, 2017). Доминирование присоединения робустного суспензориума к черепной коробке в области глазницы посредством хорошо дифференцированного вертикального отростка entopterygoideum присутствует у гимнота Gymnotus carapo (Albert et al., 2005). В целом, если рассматривать передний отдел черепа у хвостоперых ножетелок Apteronotidae, то основной поверхностью соприкосновения с добычей сверху, во время ее схватывания охотником, является нижняя сторона нейрокраниума, поскольку maxillare и premaxillare развиты слабо (Albert, Crampton, 2009). Таким образом, признаки орбитоэтмогиостилии можно обнаружить у форм, использующих и укус, и всасывание.
Черты развития орбитоэтмогиостилии присутствуют у угрехвостого сома Plotosus lineatus (рис. 8д), по-видимому, вследствие широкого использования укуса не только во время питания, но и во время турнирных боев между самцами. Суспензориум этой рыбы, по сравнению с другими сомообразными, обладает большей прочностью конструкции: palatinum формирует более крепкое соединение с крыловидными костями посредством коротких соединительнотканных волокон. Помимо сустава palatinum с ectoethmoideum, у угрехвостого сома имеется прочное прикрепление metapterygoideum к orbitosphenoideum и ectoethmoideum. Помимо этого, entopterygoideum образует сустав с vomer (Oliveira et al., 2001). У большеголового угря Macrocephenchelys brachialis (рис. 8г) хорошо развитый pterygoideum очень широко и прочно сочленяется с нейрокраниумом в области сошника и парасфеноида (в районе глазницы), а остатки palatinum неподвижно прирастают к vomer. Присутствует обширная область соединения hyomandibulare с черепной коробкой. Суспензориум весьма прочный и монолитный: pterygoideum, quadratum и hyomandibulare скреплены между собой взаимопроникающими костными отростками (Robins, Robins, 1971).
Наиболее близким к амфистилии следует считать вариант контакта суспензориума с нейрокраниумом (орбитоэтмогиостилии) у слитножаберниковых Synbranchidae, в частности, у темного болотного угря Ophisternon aenigmaticum (рис. 8е) и слитножаберника Synbranchus marmoratus. У них формируется значительное удлинение заглазничного отдела нейрокраниума, а предглазничный отдел остается коротким. Вследствие этой модификации черепа пространство для размещения на нем крупного АМ возрастает, а разрез рта животного значительно увеличивается, что повышает эффективность укуса добычи. Для укрепления конструкции аппарата питания позади глаза рыбы суспензориум формирует третью область сочленения с черепной коробкой – между ectopterygoideum и frontale (помимо суставов palatinum с этмоидом и hyomandibulare со слуховым отделом) (Gosline, 1983; Britz, 1996).
Аутоэтмогиостилия. У рыб-ежей Diodontidae в связи с прочным и малоподвижным прикреплением переднего и заднего отделов суспензориума к нейрокраниуму проявляется тенденция к образованию аутоэтмогиостилии (рис. 8з). У этой группы рыб palatinum включается в состав черепной коробки, а снизу прочно срастается с ectopterygoideum и entopterygoideum. В отличие от иглобрюховых Tetraodontidae, hyomandibulare развито хорошо и имеет крепкое причленение к нейрокраниуму (Tyler, 1980). Малоподвижное прикрепление суспензориума к черепной коробке обеспечивает прочную опору робустному MJA и надежное место начала для мощного АМ (Turingan, 1994). Таким образом, рыбы-ежи демонстрируют более специализированную, по сравнению с иглобрюховыми, стадию приспособления их висцерального аппарата к использованию дурофагии. Для некоторых мормировых Mormyridae свойственно наличие акинетичного черепа (суспензориума) (Hofer, 1945). Так, для мормиропса Mormyrops deliciosus и камнеголова Рetrocephalus bane характерно слияние palatinum обеих сторон головы с сошником, вследствие чего передний отдел суспензориума прикрепляется к нейрокраниуму неподвижно. В связи с прочным причленением hyomandibulare монолитного суспензориума к черепной коробке у обоих видов (Ridewood, 1904), в строении их висцерального аппарата можно отметить присутствие признаков аутоэтмогиостилии. Формирование данной специализации устройства черепа обусловлено использованием животными силового укуса добычи.
Тенденцию к формированию аутоэтмогиостилии (малоподвижное, прочное крепление суспензориума к нейрокраниуму спереди и сзади) можно заметить в группе кольчужных сомов Loricariidaе (рис. 8и) (Howes, 1983a; Schaefer, Provenzano, 2008). “Костная щека” этих форм испытывает большие нагрузки в процессе присасывания животного к субстрату. Так, у анциструса Ancistrus triradiatus присутствует значительное увеличение прочности рострального и каудального соединений суспензориума с нейрокраниумом: между ними формируются три области прямого контакта. У этого вида, в сравнении с другими не столь уплощенными сомами, palatinum демонстрирует малоподвижное прикрепление к metapterygoideum (коротким, а не длинным, соединительнотканным промежутком), которое находится в составе монолитной конструкции первичной верхней челюсти. Metapterygoideum рыбы самостоятельным прочным суставом надежно сочленяется с ectoethmoideum (Schaefer, 1990, 1991; Diogo, Chardon, 1998). Часть авторов полагает, что между metapterygoideum и ectoethmoideum кольчужных сомов формируется неподвижный шов (Arratia, 1990). Крупное palatinum анциструса причленяется к ectoethmoideum, а также соединено с mesethmoideum связкой. Авторы подчеркивают, что hyomandibulare надежно и малоподвижно скреплено швом с pteroticum (Diogo, Vandewalle, 2003; Geerinckx et al., 2007). Из-за отсутствия у многих кольчужных сомов LAP, их суспензориум имеет весьма сокращенную амплитуду перемещений. Обширная степень его подвижности уменьшала бы способность этих рыб к адгезии к субстрату.
Кинетизм суспензориума. Механизм “внутренней аддукции”
Подвижность суспензориума относительно черепной коробки как целого у большинства Teleostei можно охарактеризовать как плеврокинетизм – перемещение латеромедиально “костных щек” рыбы в поперечной плоскости (Иорданский, 1990). Особенности мобильности суспензориума относительно нейрокраниума формирует характер прикрепления “костной щеки” рыбы к черепной коробке, в частности структура задействованных связок и/или сочленений (см. выше). У многих Teleostei мыщелки/суставы, при помощи которых суспензориум осуществляет переднее (этмоидное) и заднее причленения к черепной коробке, отличаются по своему строению, а также могут находиться не на одной оси (быть не соостны) (Otten, 1983). Характер смещений суспензориума во многом определяют движения hyomandibulare, которое, как правило, имеет один (Branch, 1966; Pietsch, 1978b; Hilton, Fernandes, 2017) или два (Pietsch, 1978a; Grande, Poyato-Ariza, 2010; Odani, Imamura, 2011) сочленовных мыщелка для связи с нейрокраниумом (Grande et al., 2013). Последние могут, однако, варьировать по форме, размеру и/или находиться в разных плоскостях вследствие изгиба тела или рельефа hyomandibulare. Присутствие единственного мыщелка предоставляет кости бóльшую свободу поворотов относительно черепной коробки. Также важную роль играют конструкция, размер, масса и характер прикрепления обслуживающих суспензориум мускулов. В связи с вышеуказанным, подвижность суспензориума относительно нейрокраниума и других элементов черепа характеризуется сложной функцией, которая включает в себя несколько независимых переменных.
Анализ ряда литературных источников, описывающих морфологию первичной верхней челюсти Teleostei, позволил выделить, помимо мобильности их “костной щеки” как целого, присутствие внутренней подвижности одних составляющих суспензориума относительно других (см. выше). Исходная монолитная структура “костной щеки” животного делится на два или более функционально независимых “компонента” (Громова, Махотин, 2020). Кинетизм этих компонентов в основном включает в себя их смещения латеромедиально, которые в некоторых случаях могут дополняться протракцией (с обратной ретракцией). В результате такого строения черепа положение челюстного сустава рыбы может быть жестко не закреплено, и он способен в некоторой степени совершать перемещения вовнутрь–наружу и/или вперед–назад относительно нейрокраниума (Gosline, 1973). В формировании внутреннего кинетизма суспензориума также имеет значение соотношение размеров скрепленных друг с другом компонентов и величина области их соединения. Так, связь тонкого грацильного элемента с крупным робустным костным блоком морфологически предполагает возможность прогибов в узкой зоне их контакта (Gosline, 1963). Кроме того, на характер внутренней подвижности суспензориума Teleostei влияют особенности его соединения с нейрокраниумом – устройство задействованных связок и/или сочленений (Громова, Махотин, 2020).
Латеромедиальный кинетизм. Во многих случаях у Teleostei присутствует латеромедиальный кинетизм между передним (palatinum и/или entopterygoideum/ectopterygoideym) и задним отделами суспензориума (остальные кости). Двухкомпонентный суспензориум с одной зоной внутренней подвижности описываемой категории имеется у бычковых Gobiidae (Gosline, 1955), песчаного копальщика Crystallodytes cookei (Gosline, 1963), удильщиков тетрабрахиума Tetrabrachium ocellatum, клоуна-сангвиника Antennarius sanguineus (Pietsch, 1981), рогатого фонаря Centrophryne spinulosa (Pietsch, 1972) и некоторых хоботнорыловидных Mastacembeloidei (Travers, 1984; Britz, 1996). Трехкомпонентным вариантом конструкции первичной верхней челюсти обладает королевский спинорог Balistes vetula (рис. 9а), который имеет тонкое “Т”-образное palatinum и большой центральный костный блок, включающий ecto-, ento-, metapterygoideum, quadratum, symplecticum, скрепленный широким хрящевым промежутком с hyomandibulare. У спинороговых Balistidae palatinum подвижно сочленено с ectopterygoideum, а с нейрокраниумом – полоской соединительной ткани (Hofer, 1937). Благодаря такому устройству срединный скелетный блок способен осуществлять смещения относительно hyomandibulare в медиолатеральном направлении, повышая эффективность схватывания добычи путем всасывания. Всасывание представляет собой процесс попеременного расширения и сжатия внутренних камер головы животного для затягивания корма в рот вместе с небольшим количеством окружающей ее воды (Carroll et al., 2004). Многие авторы (Barel, 1983; Lauder, 1985; Gibb, Ferry-Graham, 2005) отмечают важность возможности расширения полостей черепа Teleostei для повышения эффективности процесса их всасывания, однако, часто упускают из виду значимость предшествующего всасыванию сжатия этих пространств. Расширение ротовой полости будет тем эффективнее, чем меньше был еe изначальный объем (Barel, 1983). В основе всасывания у королевского спинорога Balistes vetula лежит процесс приведения медиально подвижного центрального отдела суспензориума за счет раннего начала сокращения мощного RAP в процессе изначального схватывания добычи (Wainwright, Turingan, 1993). Сходную возможность отклонения костей суспензориума вовнутрь пространства черепа во время всасывания у рыб-игл Syngnathidae отмечает Бранч (Branch, 1966): так, symplecticum обыкновенной морской иглы Syngnathus acus, будучи соединенное с нейрокраниумом связкой, поворачивается медиально при сокращении оканчивающегося на нем ААР, тем самым сокращая объем полости рыла животного (рис. 9б). Использование описываемого механизма “внутренней аддукции” сегментов суспензориума для увеличения эффективности всасывания также свойственно австралийскому звездочету Leptoscopus macropygus и рыбе-топорику Sternoptyx diaphana (рис. 9в) (Weitzman, 1974). Подобное приспособление присуще и некоторым харациновым Characidae, таким как брикон Brycon, у которого имеется мостик “ectopterygoideum-entopterygoideum”, действующий как планка, соединяющая два подвижных отдела суспензориума – передний (palatinum) и задний (основная несущая кость – hyomandibulare) (Gosline, 1973). Ковергентно схожая анатомия свойственна перкофовой рыбе Matsubaraea fusiforme (Odani, Imamura, 2011). Подразделение суспензориума на три подвижных относительно друг друга cегмента, два из которых palatinum и hyomandibulare, присутствует у гиреллы Girella laevifrons (Vial, Ojeda, 1992) и обыкновенного илистого прыгуна Periophthalmus barbarus (рис. 3б) (Michel et al., 2014). Нами показано, что у некоторых видов Teleostei механизм внутренней аддукции является этапом работы внутреннего гидравлического механизма – интенсификации процесса попеременной смены объемов в камерах головы рыбы в процессе питания, в частности, у белого толстолобика Hypophthalmichthys molitrix (Махотин, Громова, 2019) и рубиново-пепельной рыбы-попугая Scarus rubroviolaceus (Громова, Махотин, 2019).
Рис. 9.
Внутренняя подвижность – присутствие механизма “внутренней аддукции”: а, б — в трехкомпонентном суспензориуме оранжевополосого спинорога Baliatapus undulatus (по: Tyler, 1980) и обыкновенной морской иглы Syngnathus acus (по: Branch, 1966) соответственно; в — в двухкомпонентном суспензориуме рыбы-топорика Sternoptyx diaphana (по: Weitzman, 1974). Пунктирная линия — область подвижности в рамках суспензориума. Обозначения, как на рис. 1, 6.

Переднезадние смещения. Выдвижение суспензориума отмечено у видов Teleostei, у которых palatinum отделено от entopterygoideum и ectopterygoideum. Это губан-обманщик Epibulus insidiator (Westneat, Wainwright, 1989), желтая длиннорылая рыба-бабочка Forcipiger flavissimus (Ferry-Graham et al., 2001a), рыба-лист Monocirrhus polyacanthus (Liem, 1970) и цихлиды Petenia splendida и Caquetaia spectabile (Waltzek, Wainwright, 2003). Внутренняя подвижность в суспензориуме у обыкновенной длиннорылой рыбы-бабочки Forcipiger longirostris лежит в основе возможности его протракции и, таким образом, выдвижения вперед челюстного сустава, а вместе с ним и всего MJA. Cочленения присутствуют между palatinum и entopterygoideum (поддерживается lig. palato-entopterygoideum) и между hyomandibulare и остальным комплексом костей (рис. 6б). Поворот вперед нижней ветви hyomandibulare становится возможен благодаря наличию одинарной головки, соединяющей данный элемент с нейрокраниумом. Смещение рострально (на 25°) центрального скелетного комплекса суспензориума происходит во время его аддукции посредством сокращения ААР и вызывает протракцию челюстного сустава и palatinum (Ferry-Graham et al., 2001b). Подобный механизм внутреннего кинетизма трехкомпонентного суспензориума присутствует и у некоторых цихлид Cichlidae – петении Petenia splendida и кагуеты Caquetaia spectabile. Поворот hyomandibulare у первого из этих видов достигает 24°, а у второго – 19°. Palatinum контактирует с птеригоидами при помощи гибкой полоски хряща. Смещение центрального костного блока суспензориума в суставе с hyomandibulare посредством сокращения ААР помогает выдвижению челюстного сустава этих двух видов (Waltzek, Wainwright, 2003). У определенных видов Teleostei самостоятельные сдвиги prаeoperculum относительно других составляющих первичной верхней челюсти могут вызвать протракцию отдельных ее костей (Liem, 1970). У некоторых представителей группы praeoperculum, обычно включенное в состав суспензориума, приобретает самостоятельную подвижность. Так, у керчака Leptocottus возможность подобных смещений возникает в рамках развития механизма защиты от врагов. Praeoperculum данного вида формирует два подвижных шарнирных сустава с остальным суспензориумом. Подвижность кости обеспечивается посредством сокращения специальной части LAP, которая образует обособленное место окончания на praeoperculum. В результате функционирования данного мускульного подразделения praeoperculum отклоняется вперед и вверх, выставляя расположенный на нем крупный костный шип в сторону животного-агрессора (Gosline, 1989). Кроме того, существованию протракции суспензориума у Teleostei способствует редукция некоторых или всех составляющих его рострального отдела – palatinum, ento-, ectopterygoideum. Это сокращение размеров указанных костей предоставляет пространство для поворота суспензориума вперед (Westneat, 1991). Так, значительная редукция palatinum и entopterygoideum у гнатодолуса Gnathodolus bidens, способствующая значительному уменьшению величины передней части суспензориума, обеспечивает возможность скольжения рострального отдела последнего вперед-назад по двум глубоким продольным желобкам сошника в процессе открывания и закрывания нижней челюсти рыбы (Sidlauskas, Vari, 2008).
ЗАКЛЮЧЕНИЕ
Строение суспензориума Teleostei представлено широким разнообразием как конструкции в целом, так и отдельных ее составляющих. В настоящей работе была прослежена взаимосвязь между функциональной морфологией подвесочного аппарата и экологией питания рыб. Продемонстрировано, что особенности строения суспензориума влияют на эффективность ловли корма и тем самым могут определять способы использования пищевых ресурсов в окружающей среде обитания животного. Признаки строения челюстей можно использовать в качестве индикаторов экологии питания исследуемого представителя ихтиофауны. На основе анализа литературных данных скорректирована систематизация вариантов взаимодействия суспензориума с нейрокраниумом у Teleostei. Исходное состояние для развития ряда специализированных видов устройства черепа у рыб этой группы – это этмогиостилия. Приведены примеры таксонов, представляющих собой разные морфологические “стадии” на пути формирования выделенных нами вариантов взаимодействия суспензориума и нейрокраниума у Teleostei.
Перемещения элементов суспензориума относительно нейрокраниума имеют место преимущественно в поперечной и/или парасагиттальной плоскостях. Выделенные нами варианты подвижности черепа Teleostei – кинетизм элементов в рамках конструкции суспензориума и мобильное прикрепление последнего к нейрокраниуму – могут присутствовать по отдельности, либо сочетаться в одном и том же механизме головы животного и быть представлены в разной степени. Для рыб, применяющих укус кинетизм черепа способствует гибкой подгонке челюстей рыбы к проглатываемому пищевому объекту. Описываемая подвижность может служить им также для лучшего удержания вырывающейся добычи в челюстях хищника. Внутренняя мобильность суспензориума повышает его манипуляционные возможности. Для всасывающих видов Teleostei развитие механизма “внутренней аддукции” одних отделов подвесочного аппарата относительно других способствует успешному захвату добычи.
Список литературы
Воскобойникова О.С. 1985. Морфология и основные направления эволюции висцерального скелета нототениевых рыб (Nototheniidae): Дис. … канд. биол. наук. Cанкт-Петербург: Зоол. ин-т АН СССР.
Воскобойникова О.С., Назаркин М.В. 2017. Остеология южноокеанского кинжалозуба Anotopterus vorax и положение рода Anotopterus в отряде Aulopifomes // Вопр. ихтиологии. Т. 57. № 1. С. 4. https://doi.org/10.7868/80042875217010167
Громова Е.С., Дзержинский Ф.Я., Махотин В.В. 2017. Морфофункциональные особенности висцерального аппарата серебряной араваны Osteoglossum bicirrhosum (Osteoglossidae) // Вопр. ихтиологии. Т. 57. № 4. С. 379. https://doi.org/10.7868/S0042875217040038
Громова Е.С., Махотин В.В. 2016. Функциональная морфология висцерального аппарата семги Salmo salar (Salmonidae) // Вопр. ихтиологии. Т. 56. № 4. С. 410. https://doi.org/10.7868/S0042875216040068
Громова Е.С., Махотин В.В. 2019. Детали строения и функционирования аппарата глоточных челюстей рубиново-пепельной рыбы-попугая Scarus rubroviolaceus (Scaridae) // Вопр. ихтиологии. Т. 59. № 6. С. 1. https://doi.org/10.1134/S0042875219060031
Громова Е.С., Махотин В.В. 2020. Морфофункциональные особенности висцерального аппарата рубиново-пепельной рыбы-попугая Scarus rubroviolaceus (Scaridae) // Вопр. ихтиологии. Т. 60. № 2. P. 137. https://doi.org/10.31857/S0042875220010075
Гуртовой Н.Н., Матвеев Б.С., Дзержинский Ф.Я. 1976. Практическая зоотомия позвоночных (низшие хордовые, бесчелюстные, рыбы) // Учеб. пособие для биол. специальностей ун-ов. Москва: Высш. школа.
Дзержинский Ф.Я. 2005. Сравнительная анатомия позвоночных животных. Москва: Аспект Пресс.
Дзержинский Ф.Я., Васильев Б.Д., Малахов В.В. 2013. Зоология позвоночных. Москва: Академия.
Иорданский Н.Н. 1990. Эволюция комплексных адаптаций. Челюстной аппарат амфибий и рептилий. Москва: Наука.
Кузищин К.В., Груздева М.А., Филенко В.А. и др. 2020. Сельдь-черноспинка Alosa kessleri kessleri (Grimm, 1887) из р. Ахтубы нижневолжского бассейна: биологические и морфологические особенности // Биология внутр. вод. № 1. С. 67. (Kuzishchin K.V., Gruzdeva M.A., Filenko V.A. et al. 2020. Caspian anadromous shad Alosa kessleri kessleri (Grimm, 1887) from the Akhtuba river, lower Volga River basin: biological and morphological features // Inland Water Biol. V. 13. P. 79. https://doi.org/10.1134/S1995082920010125) https://doi.org/10.31857/S0320965220010143
Кэрролл Р. 1992. Палеонтология и эволюция позвоночных. Москва: Мир. Т. 1. С. 279. (Carroll R.L. 1988. Vertebrate paleontology and evolution // New York. W.H. Freeman and Company. P. 698).
Махотин В.В., Громова Е.С. 2019. Детали строения скелета, мышц и соединительнотканных элементов головы белого толстолобика Hypophthalmichthys molitrix (Cyprinidae) в связи с особенностями функционирования его висцерального аппарата // Вопр. ихтиологии. Т. 59. № 1. Р. 3. https://doi.org/10.1134/S0042875219010053
Ромер А., Парсонс Т. 1992. Анатомия позвоночных. Т. 1. Москва: Мир. (Romer A.S., Parsons Th.S. 1992. The vertebrate body. Saunders College Publish. P. 624.)
Adriaens D. 2003. Feeding mechanisms in catfishes // Catfishes. Sci. Publ. P. 221.
Albert J.S., Crampton W.G.R. 2009. A new species of electric knifefish, genus Compsaraia (Gymnotiformes: Apteronotidae) from the Amazon River, with extreme sexual dimorphism in snout and jaw length // System. Biodiv. V. 7. № 1. P. 81. https://doi.org/10.1017/S1477200008002934
Albert J.S., Crampton W.G.R., Thorsen D.H., Lovejoy N.R. 2005. Phylogenetic systematics and historical biogeography of the Neotropical electric fish Gymnotus (Teleostei: Gymnotidae) // System. Biodiv. V. 2. № 4. P. 375. https://doi.org/10.1017/S1477200004001574
Anker G.Ch. 1974. Morphology and kinetics of the head of the stickleback, Gasterosteus aculeatus // Trans. Zool. Soc. Lond. V. 32. P. 311.
Arratia G. 1990. Development and diversity of the suspensorium of trichomycterids and comparison with loricarioids (Teleostei: Siluriformes) // J. Morph. V. 205. P. 193. https://doi.org/10.1002/jmor.1052050208
Arratia G. 1992. Development and variation of the suspensorium of primitive catfishes (Teleostei: Ostariophysi) and their phylogenetic relationships // Bonn. Zool. Monograph. V. 32. P. 1.
Ballintijn C.M., Burg A., Egberink B.P. 1972. An electromyographic study of the adductor mandibulae complex of a free-swimming carp (Cyprinus carpio L.) during feeding // J. Exp. Biol. V. 57. P. 261.
Barel C.D.N. 1983. Towards a constructional morphology of cichlid fishes (Teleostei, Perciformes) // Nether. J. Zool. V. 33. № 4. P. 357.
Bertin L. 1934. Les poissons Apodes appartenant au sous-ordre des lyomeres // Dana Rep. V. 3. P. 1.
Branch G.M. 1966. The feeding mechanism of Syngnathus acus Linnaeus // Zool. Afric. V. 2. № 1. P. 69.
Britz R. 1996. Ontogeny of the ethmoidal region and hyopalatine arch in Macrognathus pancalus (Percomorpha, Mastacembeloidei), with critical remarks on Mastacembeloid inter- and intrarelationships // Am. Mus. Novit. № 3181. P. 1.
Britz R., Kottelat M. 2003. Descriptive osteology of the family Chaudhuriidae (Teleostei, Synbranchiformes, Mastacembeloidei), with a discussion of its relationships // Amer. Mus. Novit. № 3418. P. 1. https://doi.org/10.1206/0003-0082(2003)418<0001:DO-OTFC>2.0.CO;2
Brown B.A., Ferraris C.J. 1988. Comparative osteology of the asian catfish family Chacidae, with the description of a new species from Burma // Am. Mus. Novit. № 2907. P. 1.
Carroll A.M., Wainwright P.C., Huskey S.H. et al. 2004. Morphology predicts suction feeding performance in centrarchid fishes // J. Exp. Biol. V. 207. P. 3873. https://doi.org/10.1242/jeb.01227
Chapman W.McL. 1942. The osteology and relationship of the bathypelagic fish Macropinna microstoma, with notes on its visceral anatomy // Ann. Magaz. Nat. Hist. Ser. 11. V. 9. P. 272.
Clack J.A. 1992. The stapes of Acanthostega gunnari and the role of the stapes in early tetrapods // The evolutionary biology of hearing. New York: Springer-Verlag. P. 405.
Datovo A., Vari R.P. 2014. The adductor mandibulae muscle complex in lower teleostean fishes (Osteichthyes: Actinopterygii): comparative anatomy, synonymy, and phylogenetic implications // Zool. J. Linn. Soc. V. 171. P. 554. https://doi.org/10.1111/zoj12142
De Beer G.R. 1937. The development of the vertebrate skull. Oxford: Oxford Univ. Press.
Delsman H.C. 1925. Fishes with protrusile mouths // Treubia. V. 6. № 2. P. 98.
Depew M.J., Simpson C.A. 2006. 21st century neontology and the comparative development of the vertebrate skull // Develop. Dynamics. V. 235. P. 1256. https://doi.org/10.1002/dvdy.20796
Diogo R., Chardon M. 1998. Morphofunctional and comparative analysis of the suspensorium in catfish // Proceedings of the 5th Benelux Congress of Zoology. V. 235. P. 61. https://doi.org/10.1002/dvdy.20796
Diogo R., Vandewalle P. 2003. Catfishes as a case study for discussions on general evolution: the importance of functional uncouplings in morphological macroevolution // Europ. J. Morph. V. 41. № 3/4. P. 139. https://doi.org/10.1080/09243860500043596
Dutta H.M. 1975. The suspensorium of Ctenopoma acutirostre: a comparative functional analysis with Anabas testudineus // J. Morph. V. 146. P. 457.
Eagderi S. 2010. Structural diversity in the cranial musculoskeletal system in Anguilliformes: an evolutionary-morphological study: Diss. Doc. Sci. Biol. P. 1–157.
Eagderi S., Christiaens J., Boone M. et al. 2016. Functional morphology of the feeding apparatus in Simenchelys parasitica (Simenchelyinae: Synaphobranchidae), an alleged parasitic eel // Copeia. № 2. P. 421. https://doi.org/10.1643/CI-15-329
Emery A.R. 1973. Comparative ecology and functional osteology of fourteen species of damselfish (Pisces: Pomacentridae) at Alligator reef, Florida keys // Bull. Marine Sci. V. 23. № 3. P. 649.
Ferry-Graham L.A., Wainwright, P.C., Hulsey C.D., Bellwood D.R. 2001a. Evolution and mechanics of long jaws in butterflyfishes (Family Chaetodontidae) // J. Morph. V. 248. P. 120. https://doi.org/10.1002/jmor.1024
Ferry-Graham L.A., Wainwright, P.C., Bellwood D.R. 2001b. Prey capture in long-jawed butterflyfishes (Chaetodontidae): the functional basis of novel feeding habits // J. Exp. Mar. Biol. Ecol. V. 256. P. 167. https://doi.org/10.1016/S0022-0981(00)00312-9
Fink W.L. 1985. Phylogenetic interrelationships of the stomiid fishes (Teleostei: Stomiiformes) // Misc. Publ. Mus. Zool. Univ. Michigan. № 171. P. 1.
Fink S.V., Fink W.L. 1981. Interrelationships of the Ostariophysan fishes (Teleostei) // Zool. J. Linn. Soc. V. 72. № 4. P. 297. https://doi.org/10.1111/j.1096-3642.1981.tb01575.x
Geerinckx T., Brunain M., Herrel A. et al. 2007. A head with a suckermouth: a functional-morphological study of the head of the suckermouth armoured catfish Ancistrus cf. triradiatus (Loricariidae, Siluriformes) // Belg. J. Zool. V. 137. № 1. P. 47.
Gibb A.C., Ferry-Graham L. 2005. Cranial movements during suction feeding in teleost fishes: are they modified to enhance suction production? // Zoology. V. 108. P. 141. https://doi.org/10.1016/j.zool.2005.03.004
Goodrich E.S. 1930. Studies on the structure and development of vertebrates // London: Macmil. Co.
Gosline W.A. 1951. The osteology and classification of the ophichthid eels of the Hawaiian Islands // Pacif. Sci. V. 5. № 4. P. 298.
Gosline W.A. 1955. The osteology and relationships of certain gobioid fishes, with particular reference to the genera Kraemeria and Microdesmus // Pacif. Sci. V. 9. № 2. P. 158.
Gosline W.A. 1963. Notes on the osteology and systematic position of Hypoptychus dybowskii Steindachner and other elongate perciform fishes // Pacif. Sci. V. 17. № 1. P. 90.
Gosline W.A. 1973. Considerations regarding the phylogeny of cypriniform fishes, with special reference to structures associated with feeding // Copeia. V. 1973. № 4. P. 761.
Gosline W.A. 1975. The palatine- maxillary mechanism in catfishes, with comment on the evolution and zoogeography of modern Siluroids // Occ. Pap. Calif. Acad. Sci. V. 120. P. 1.
Gosline W.A. 1983. The relationships of the Mastacembelid and Synbranchid fishes // Japan. J. Icht. V. 29. № 4. P. 323.
Gosline W.A. 1984. Structure, function, and ecology in the goatfishes (Family Mullidae) // Pacif. Sci. V. 38. № 4. P. 312.
Gosline W.A. 1989. Movable head armature in higher teleostean fishes // Envir. Biol. Fish. V. 24. № 1. P. 75. https://doi.org/10.1007/BF00001612
Grande T., Poyato-Ariza F.J. 2010. Reassessment and comparative morphology of the gonorynchiform head skeleton // Gonorynchiformes and Ostariophysan relationships. A comprehensive review. Ser. Teleost. // Fish Biol. P. 1.
Grande T., Borden W.C., Smith W.L. 2013. Limits and relationships of Paracanthopterygii: A molecular framework for evaluating past morphological hypotheses // Global Diversity and Evolution. P. 385.
Gregory W.K. 1904. The relations of the anterior visceral arches to the chondrocranium // Biol. Bull. V. 7. № 1. P. 55.
Gromova E.S., Makhotin V.V. 2018. Maxillary apparatus in feeding of the silver carp Hypophthalmichthys molitrix (Cyprinidae) // J. Ichthyol. V. 58. № 6. P. 857. https://doi.org/10.1134/S0032945218060036
Hilton E.J. 2007. Redescription of Orthosternarchus tamandua (Boulenger, 1898) (Gymnotiformes, Apteronotidae), with reviews of its ecology, electric organ discharges, external morphology, osteology, and phylogenetic affinities // Proc. Acad. Nat. Sci. Phil. V. 156. P. 1. https://doi.org/10.1635/0097-3157(2007)156[1:ROO-TBG]2.0.CO;2
Hilton E.J., Fernandes C.C. 2017. Identity of “Apteronotus” bonapartii (Castelnau, 1855), a sexually dimorphic South American knifefish from the Amazon, with notes on its cranial osteology and on the taxonomic status of “Apteronotus” apurensis Fernandez-Yepez, 1968 (Gymnotiformes, Apteronotidae) // Proc. Acad. Nat. Sci. Phil. V. 165. P. 91. https://doi.org/10.1635/053.165.0109
Hofer H. 1937. Der Schadel von Tetrodon und Balistes, nebst allgemeinen Bemerkungen uber die Funktion des Palatoquadratbogens der Knochenfische // Diss. Philosoph. Fakult. Univ. Wien.
Hofer H. 1945. Zur Kenntnis der Suspensionsformen des Kieferbogens und deren Zusammenhange mit dem Bau des knochernen Gaumens und mit der Kinetik des Schadels bei den Knochenfischen // Zool. Jahrb. Anat. Ontog. V. 69. P. 321.
Howes G.J. 1983a. The cranial muscles of the loricarioid catfishes, their homologies and value as taxonomic characters (Teleostei: Siluroidei) // Bull. Br. Mus. Nat. Hist. Zool. V. 45. P. 309.
Howes G.J. 1983b. Problems in catfish anatomy and phylogeny exemplified by the Neotropical Hypophthalmidae (Teleostei: Siluroidei) // Bull. Brit. Mus. Nat. Hist. Zool. V. 45. P. 1.
Howes G.J. 1992. Notes on the anatomy and classification of ophidiiform fishes with particular reference to the abyssal genus Acanthonus Gunther, 1878 // Bull. Br. Mus. Nat. Hist. Zool. V. 58. № 2. P. 95.
Howes G.J., Fumihito A. 1991. Cranial anatomy and phylogeny of the South- east asian catfish genus Belodontichthys // Bull. Br. Mus. Nat. Hist. Zool. V. 57. № 2. P. 133.
Huysentruyt F., Adriaens D. 2005. Descriptive osteology of Corydoras aeneus (Siluriformes: Callichthyidae) // Cybium. V. 29. № 3. P. 261.
Kayser H. 1962. Vergleichende Untersuchung uber Vorstreckmechanismen der Oberkiefer bei Fischen. Der Bau und die Funktion des Kiefer- und Kiemenapparates von Knochenfischen der Gattung Ammodytes und Callionymus // Zool. Beitr. N.F. V. 7. P. 321.
Kirchhoff H. 1958. Funktionell – anatomische Untersuchung des Visceralapparates von Clupea harengus L. // Zool. Jahrb. Abteil. Anat. Ontog. Tier. Band 76. Heft 4. S. 462.
Konow N., Ferry-Graham L.A. 2013. Functional morphology of butterflyfishes // The biology of butterflyfishes. Taylor, Francis Group. Boca Raton: CRC Press. P. 19.
Konstantinidis P., Johnson G.D. 2016. Osteology of the telescopefishes of the genus Gigantura (Brauer, 1901), Teleostei: Aulopiformes // Zool. J. Linn. Soc. V. 179. № 2. P. 338. https://doi.org/10.1111/zoj.12469
Lauder G.V. 1980. Evolution of the feeding mechanism in primitive actinopterygian fishes: a functional anatomical analysis of Polypterus, Lepisosteus and Amia // J. Morphol. V. 163. P. 283. https://doi.org/10.1002/jmor.1051630305
Lauder G.V. 1985. Aquatic feeding in lower vertebrates // Functional vertebrate morphology. Cambridge: Harvard Univ. Press. P. 210–229.
Lauder G.V., Liem K.F. 1981. Prey capture by Luciocephalus pulcher: implications for models of jaw protrusion in teleost fishes // Envir. Biol. Fish. V. 6. № 3/4. P. 257. https://doi.org/10.1007/BF00005755
Liem K.F. 1970. Comparative functional anatomy of the Nandidae (Pisces: Teleostei) // Field. Zool. Ser. V. 56. P. 1.
Longo S.J., McGee M.D., Oufiero C.E. et al. 2016. Body ram, not suction, is the primary axis of suction-feeding diversity in spiny-rayed fishes // J. Exp. Biol. V. 219. P. 119. https://doi.org/10.1242/jeb.129015
Lundberg J.G., Fernandes C.C., Albert J.S., Garcia M. 1996. Magosternarchus, a new genus with two new species of electric fishes (Gymnotiformes: Apteronotidae) from the Amazon River basin, South America // Copeia. V. 1996. № 3. P. 657. https://doi.org/10.2307/1447530
Marrero C., Winemillar K.O. 1993. Tube-snouted gymnotiform and mormyriform fishes: convergence of a specialized foraging mode in teleosts // Envir. Biol. Fish. V. 38. P. 299. https://doi.org/10.1007/BF00007523
Mehta R.S. 2009. Ecomorphology of the moray bite: relationship between dietary extremes and morphological diversity // Physiol. Biochem. Zool. V. 82. № 1. P. 90. https://doi.org/10.1086/594381
Mehta R.S., Wainwright P.C. 2007. Biting releases constraints on moray eel feeding kinematics // J. Exp. Biol. V. 210. P. 495. https://doi.org/10.1242/jeb.02663
Michel K.B., Adriaens D., Aerts P. et al. 2014. Functional anatomy and kinematics of the oral jaw system during terrestrial feeding in Periophthalmus barbarus // J. Morph. V. 275. P. 1145. https://doi.org/10.1002/jmor.20291
Motta P.J. 1982. Functional morphology of the head of the inertial suction feeding butterflyfish, Chaetodon miliaris (Perciformes, Chaetodontidae) // J. Morph. V. 174. P. 283.
Moy-Thomas J.A. 1939. Palaeozoic Fishes. New York: Chemical Pub.
Nelson J.S. 2006. Fishes of the world. New Jersey: Wiley.
Nielsen J.G., Bertelsen E., Jespersen A. 1989. The Biology of Eurypharynx pelecanoides (Pisces, Eurypharyngidae) // Acta Zool. Stock. V. 70. № 3. P. 187. https://doi.org/10.1111/j.1463-6395.1989.tb01069.x
Norman J.R. 1925. Two new fishes from Tonkin, with notes on the Siluroid genera Glyptosternum, Exostoma, etc // Ann. Magaz. Nat. Hist. V. 15. № 89. P. 570. https://doi.org/10.1080/00222932508633249
Odani K., Imamura H. 2011. New phylogenetic proposal for the family Leptoscopidae (Perciformes: Trachinoidei) // Bull. Fish. Sci. Hokkaido Univ. V. 62. № 2/3. P. 49.
Oliveira C., Diogo R., Vandewalle P., Chardon M. 2001. Osteology and myology of the cephalic region and pectoral girdle of Plotosus lineatus, with comments on Plotosidae (Teleostei: Siluriformes) autapomorphies // J. Fish Biol. V. 59. P. 243. https://doi.org/10.1111/j.1095-8649.2001.tb00128.x
Olney J.E., Johnson G.D., Baldwin C.C. 1993. Phylogeny of lampridiform fishes // Bull. Mar. Sci. V. 32. № 1. P. 137.
Osse J.W.M. 1969. Functional morphology of the head of the perch (Perca fluviatilis L.): an electromyographic study // Neth. J. Zool. V. 19. P. 289.
Otten E. 1983. The jaw mechanism during growth of a generalized Haplochromis species: H. elegans Trewavas 1933 (Pisces, Cichlidae) // Neth. J. Zool. V. 33. № 1. P. 55.
Parmentier E., Chardon M., Poulicek M. et al. 1998. Morphology of the buccal apparatus and related structures in four species of Carapidae // Aust. J. Zool. V. 46. P. 391. https://doi.org/10.1071/ZO97035
Pietsch T.W. 1972. A review of the monotypic deep-sea anglerfish family Centrophrynidae: taxonomy, distribution and osteology // Copeia. V. 1972. № 1. P. 17. https://doi.org/10.2307/1442779
Pietsch T.W. 1978a. Evolutionary relationships of the sea moths (Teleostei: Pegasidae) with a classification of gasterosteiform families // Copeia. V. 1978. № 3. P. 517. https://doi.org/10.2307/1443620
Pietsch T.W. 1978b. The feeding mechanism of Stylephorus chordatus (Teleostei: Lampridiformes): functional and ecological implications // Copeia. V. 1978. № 2. P. 255. https://doi.org/10.2307/1443560
Pietsch T.W. 1981. The osteology and relationships of the anglerfish genus Tetrabrachium with comments on lophiiform classification // Fish. Bull. V. 79. № 3. P. 387.
Regan C.T. 1924. The morphology of a rare oceanic fish, Stylophorus chordatus, Shaw; based on specimens collected in the Atlantic by the “Dana” expeditions 1920–1922 // Proc. Royal Soc. Lond. Ser. B. V. 96. № 674. P. 193.
Ridewood W.G. 1904. On the cranial osteology of the fishes of the families Mormyridae, Notopteridae and Hyodontidae // Zool. J. Linn. Soc. V. 29. № 190. P. 188.
Robins C.H. 1971. The comparative morphology of the synaphobranchid eels of the straits of Florida // Proc. Acad. Nat. Sci. Phil. V. 123. № 7. P. 153.
Robins C.H., Robins C.R. 1971. Osteology and relationships of the eel family Macrocephenchelyidae // Proc. Acad. Nat. Sci. Phil. V. 123. № 6. P. 127.
Sanford C.P., Lauder G.V. 1989. Functional morphology of the “tongue-bite” in the osteoglossomorph fish Notopterus // J. Morphol. V. 202. P. 379. https://doi.org/10.1002/jmor.1052020307
Santana C.D., Vari R.P. 2010. Electric fishes of the genus Sternarchorhynchus (Teleostei, Ostariophysi, Gymnotiformes); phylogenetic and revisionary studies // Zool. J. Linn. Soc. V. 159. P. 223. https://doi.org/10.1111/j.1096-3642.2009.00588.x
Sarmento-Soares L.M., Porto M. 2006. Comparative anatomy of the cheek muscles within the Centromochlinae subfamily (Ostariophysi, Siluriformes, Auchenipteridae) // J. Morph. V. 267. P. 187. https://doi.org/10.1002/jmor.10395
Sasaki K., Tanaka Y., Takata Y. 2006. Cranial morphology of Ateleopus japonicus (Ateleopodidae: Ateleopodiformes), with a discussion on metamorphic mouth migration and lampridiform affinities // Ichthyol. Res. V. 53. P. 254. https://doi.org/10.1007/s10228-006-0343-5
Schaefer S.A. 1990. Anatomy and relationships of the scoloplacid catfishes // Proc. Acad. Nat. Sci. Phil. V. 142. P. 167.
Schaefer S.A. 1991. Phylogenetic analysis of the loricariid subfamily Hypoptopomatinae, (Pisces: Siluroidei: Loricariidae), with comments on generic diagnoses and geographical distribution // Zool. J. Linn. Soc. V. 102. P. 1. https://doi.org/10.1111/j.1096-3642.1991.tb01535.x
Schaefer S.A., Provenzano F. 2008. The Lithogeninae (Siluriformes, Loricariidae): anatomy, interrelationships, and description of a new species // Amer. Mus. Novit. № 3637. P. 1. https://doi.org/10.1206/625.1
Shafland J.L. 1968. Functional and anatomical convergence in body form and feeding behavior in three diverse species of freshwater bottom dwelling vertebrates // PhD. Thes. Univ. Chicago. P. 183.
Sidlauskas B.L., Vari R.P. 2008. Phylogenetic relationships within the South American fish family Anostomidae (Teleostei, Ostariophysi, Characiformes) // Zool. J. Linn. Soc. V. 154. P. 70. https://doi.org/10.1111/j.1096-3642.2008.00407.x
Stensio E.A. 1921. Triassic fishes from Spitzbergen // Vienna. A. Holzhausen. P. 43.
Stensio E.A. 1932. Triassic fishes from East Greenland collected by the Danish expeditions in 1929–1931 // Meddel. Gronland. V. 83. № 3. P. 1.
Sulak K.J. 1977. The systematics and biology of Bathypterois (Pisces, Chlorophthalmidae) with a revised classification of benthic myctophiform fishes // Galathea Rep. V. 14. P. 49.
Tchernavin V.V. 1947. Six specimens of Lyomeri in the British Museum (with notes on the skeleton of Lyomeri) // Zool. J. Linn. Soc. V. 41. № 279. P. 287. https://doi.org/10.1111/j.1096-3642.1940.tb02078.x
Travers R.A. 1984. A review of the Mastacembeloidei, a suborder of synbranchiform teleost fishes. Part 1. Anatomical descriptions // Bull. Brit. Mus. Nat. Hist. Zool. V. 46. № 1. P. 1.
Turingan R.G. 1994. Ecomorphological relationships among Caribbean tetraodontiform fishes // J. Zool. Lond. V. 233. P. 493. https://doi.org/10.1111/j.1469-7998.1994.tb05279.x
Turingan R.G., Wainwright P.C. 1993. Morphological and functional bases of durophagy in the queen triggerfish, Balistes vetula (Pisces, Tetraodontiformes) // J. Morph. V. 215. P. 101. https://doi.org/10.1002/jmor.1052150202
Tyler J.C. 1980. Osteology, phylogeny, and higher classification of the fishes of the order Plectognathi (Tetraodontiformes) // United States. Nat. Mar. Fish. Serv. V. 434. P. 1.
Vari R.P. 1979. Anatomy, relationships and classification of the families Citharinidae and Distichodontidae (Pisces, Characoidea) // Bull. Br. Mus. Nat. Hist. Zool. V. 36. № 2. P. 261.
Vial C.I., Ojeda F.P. 1992. Comparative analysis of the head morphology of Pacific temperate kyphosid fishes: a morpho-functional approach to prey-capture mechanisms // Rev. Chil. Hist. Nat. V. 65. P. 471.
Wainwright P.C. 1991. Ecomorphology: experimental functional anatomy for ecological problems // Amer. Zool. V. 31. P. 680. https://doi.org/10.1093/icb/31.4.680
Wainwright P.C., Richard B.A. 1995. Predicting patterns of prey use from morphology of fishes // Envir. Biol. Fish. V. 44. P. 97. https://doi.org/10.1007/BF00005909
Wainwright P.C., Turingan R.G. 1993. Coupled versus uncoupled functional systems: motor plasticity in the queen triggerfish Balistes vetula // J. Exp. Biol. V. 180. P. 290. https://doi.org/10.1242/jeb.180.1.209
Wainwright P.C., Turingan R.G., Brainerd E.L. 1995. Functional morphology of pufferfish inflation: mechanism of the buccal pump // Copeia. V. 1995. № 3. P. 614. https://doi.org/10.2307/1446758
Waltzek T.B., Wainwright P.C. 2003. Functional morphology of extreme jaw protrusion in neotropical cichlids // J. Morph. V. 257. P. 96. https://doi.org/10.1002/jmor.10111
Weitzman S.H. 1974. Osteology and evolutionary relationships of the Sternoptychidae with a new classification of stomiatoid families // Bull. Amer. Mus. Nat. Hist. V. 153. № 3. P. 327.
Westneat M.W. 1991. Linkage biomechanics and evolution of the jaw protrusion mechanism of the sling-jaw wrasse, Epibulus insidiator // J. Exp. Biol. V. 159. P. 165. https://doi.org/10.1242/jeb.159.1.165
Westneat M.W. 1993. Phylogenetic relationships of the tribe Cheilinini (Labridae: Perciformes) // Bull. Mar. Sci. V. 52. № 1. P. 351.
Westneat M.W. 1994. Transmission of force and velocity in the feeding mechanisms of labrid fishes (Teleostei, Perciformes) // Zoomorph. V. 114. P. 103. https://doi.org/10.1007/BF00396643
Westneat M.W. 2004. Evolution of levers and linkages in the feeding mechanisms of fishes // Integr. Comp. Biol. V. 44. P. 378. https://doi.org/10.1093/icb/44.5.378
Westneat M.W., Wainwright P.C. 1989. The feeding mechanism of the sling-jaw wrasse Epibulus insidiator (Labridae; Teleostei): evolution of a novel functional system // J. Morph. V. 202. P. 129. https://doi.org/10.1002/jmor.1052020202
Wilga C.A.D., Ferry L.A. 2015. Functional anatomy and biomechanics of feeding in Elasmobranchs // Physiology of elasmobranch fishes: structure and interaction with environment. Acad. Press Elsevier. P. 153. https://doi.org/10.1016/B978-0-12-801289-5.00004-3
Winterbottom R. 1974a. A descriptive synonymy of the striated muscles of the Teleostei // Proc. Acad. Nat. Sci. Phil. V. 125. P. 225.
Winterbottom R. 1974b. The familial phylogeny of the Tetraodontiformes (Acanthopteqgii: Pisces) as evidenced by their comparative myology // Smiths. Contrib. Zool. V. 155. P. l.
Wilson M.V.H., Veilleux P. 1982. Comparative osteology and relationships of the Umbridae (Pisces: Salmoniformes) // Zool. J. Linn. Soc. V. 76. P. 321. https://doi.org/10.1111/j.1096-3642.1982.tb02186.x
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод