

Биология внутренних вод, 2022, № 6, стр. 654-663
Центрические диатомовые водоросли (Bacillariophyta, Coscinodiscaceae) планктона Чебоксарского водохранилища
С. И. Генкал a, *, А. Г. Охапкин b, Е. Л. Воденеева b, c
a Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
пос. Борок, Некоузский р-н, Ярославская обл, Россия
b Нижегородский государственный университет им. Н.И. Лобачевского,
Институт биологии и биомедицины
Нижний Новгород, Россия
c Нижегородский филиал “Всероссийский научно-исследовательский институт рыбного хозяйства и океанографии”
Нижний Новгород, Россия
* E-mail: genkal@ibiw.ru
Поступила в редакцию 14.04.2022
После доработки 07.06.2022
Принята к публикации 15.06.2022
- EDN: EAQSGM
- DOI: 10.31857/S0320965222060031
Аннотация
В ходе изучения современных материалов (2013–2020 гг.) по фитопланктону Чебоксарского водохранилища с помощью сканирующей электронной микроскопии было выявлено всего 29 видов и разновидностей центрических диатомовых водорослей из 28 родов: Actinocyclus – 1, Aulacoseira – 5, Conticribra – 1, Cyclostephanos – 1, Cyclotella – 6, Discostella – 1, Melosira – 1, Skeletonema – 1, Stephanodiscus – 8, Thalassiosira – 3. Зафиксированы три вида новых для альгофлоры водохранилища: Aulacoseira pusilla, Cyclotella cryptica и Stephanodiscus lacustris. Уточнено систематическое положение ряда таксонов из родов Aulacoseira, Cyclotella, Stephanodiscus и Thalassiosira и на основе проведенной ревизии представлен скорректированный видовой состав Coscinodiscaceae – 35 таксонов. Охарактеризованы роль центрических диатомовых водорослей в фитопланктоне водохранилища и тенденции изменений состава и обилия этой группы за период 1970–2020 гг. в условиях потепления климата и усиления инвазионных процессов.
ВВЕДЕНИЕ
Чебоксарское водохранилище образовано в декабре 1980 г. Диатомовые водоросли относили к доминирующей группе фитопланктона водоема и его основных притоков в период его заполнения (25–62% численности и 49–89% биомассы водорослей), что наблюдали и в незарегулированной Волге (Кузьмин, Охапкин, 1975; Юлова, 1982; Охапкин, 1984). Среди доминантов отмечены представители Coscinodiscaceae из родов Cyclotella, Melosira, Sceletonema, Stephanodiscus (Охапкин, Генкал, 1989). Обобщенные данные по водорослям, в том числе центрическим диатомовым, по трассе строящегося Чебоксарского водохранилища приведены Г.В. Кузьминым (1978) – 51 таксон видового и внутривидового ранга (Melosira – 18, Cyclotella – 14, Stephanodiscus – 14, Coscinodiscus – 2, Thalassiosira – 3). По результатам изучения фитопланктона на 16 станциях основной акватории водохранилища и системы его боковой приточности (1985 год: май, июль, октябрь) выявлено 25 таксонов Coscinodiscaceae (Cyclostephanos – 1, Cyclotella – 3, Melosira – 7, Sceletonema – 2, Stephanodiscus – 9, Thalassiosira – 3), в том числе девять новых для флоры водоема (Охапкин, Генкал, 1989).
В первом атласе по Bacillariophyta планктона р. Волги (материалы 1970–1989 гг.) для водохранилища опубликованы краткие диагнозы и электронные иллюстрации 31 вида и разновидности из 8 родов: Actinocyclus – 2, Aulacosira – 4, Cyclostephanos – 1, Cyclotella – 5, Melosira – 1, Sceletonema – 2, Stephanodiscus – 11, Thalassiosira – 4 (Генкал, 1992). Позднее, на основе изучения материалов 1981–1990 гг., выявлено 35 видов, подвидов, разновидностей и форм из 8 родов: Attheya – 1, Cyclostephanos – 1, Cyclotella – 5, Melosira – 13, Rhizosolenia – 1, Sceletonema – 2, Stephanodiscus – 9, Thalassiosira – 3 (Охапкин, 1994). В 2000 г. для каскада волжских водохранилищ по литературным данным и результатам собственных исследований (1953–1999 гг.) приведен перечень водорослей, в том числе и диатомовых (Bacillariophyta, Coscinodiscaceae), в котором для Чебоксарского водохранилища указаны 37 видов и разновидностей (Acanthoceras – 1, Actinocyclus – 1, Aulacoseira – 5, Cyclostephanos – 1, Cyclotella – 8, Melosira – 1, Rhizosolenia – 1, Skeletonema – 2, Stephanodiscus – 10, Thalassiosira – 7 (Корнева, Генкал, 2000). Почти аналогичный список опубликован и в последней обобщающей работе по волжским водохранилищам на основе оригинальных и литературных данных и современной таксономии (1976–2005 гг.) – 38 таксонов видового и внутривидового ранга: Acanthoceras – 1, Actinocyclus – 1, Aulacoseira – 6, Cyclostephanos – 1, Cyclotella – 8, Melosira – 1, Rhizosolenia – 1, Skeletonema – 2, Stephanodiscus – 11, Thalassiosira – 6 (Корнева, 2015). Количественные показатели фитопланктона водоема и диатомовых в его составе обсуждаются в целом ряде работ (Охапкин, 1994); Корнева, 2015), при этом Coscinodiscaceae по-прежнему доминируют в его альгоценозах.
Цель работы – на основе данных многолетних исследований первых десятилетий ХХI в. охарактеризовать современный состав, показатели обилия центрических диатомовых водорослей, оценить динамику и роль этой группы в формировании планктонных фитоценозов водохранилища и выявить тенденции их изменений за пятидесятилетний период.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Материалом для работы послужили пробы фитопланктона, собранные в 2000–2020 гг. на 87 станциях Чебоксарского водохранилища и устьевых участках его крупных притоков (Ока, Узола, Кудьма, Cура, Ветлуга, Керженец и др.), из них в верхнем речном районе (до г. Нижнего Новгорода) – 22 станции, среднем речном (до устья р. Сура) – 37, озерном – 28. Отбор проб из фотического слоя воды, их качественный и количественный анализ проводили стандартными методами (Методика…, 1975) в местах нереста и нагула рыб при осуществлении рыбохозяйственного мониторинга (Государственное задание Всероссийского научно-исследовательского института рыбного хозяйства и океанографии). Материалы 2013–2020 гг. изучены с применением сканирующего электронного микроскопа (СЭМ) JSM-6510LV. Освобождение створок диатомей от органических веществ проводили методом холодного сжигания (Балонов, 1975). Для характеристики тенденций изменения количественных показателей альгоценозов привлечены данные предыдущих исследований (1969–1990 гг. (Охапкин, 1994)). Проведен стандартный статистический анализ полученных данных.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
В исследованном материале обнаружено 29 видов и разновидностей центрических диатомовых водорослей, в том числе новые для альгофлоры Чебоксарского водохранилища. Ниже приведены их краткие диагнозы и оригинальные иллюстрации.
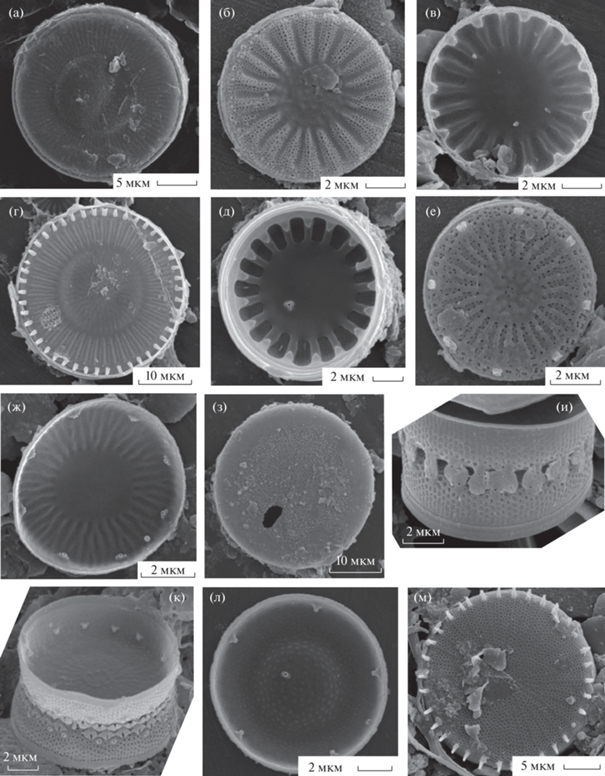
Actinocyclus normanii (Gregory ex Greville) Hustedt (рис. 1а). Створки диаметром 13–16.6 мкм, высотой 9.3–11.5 мкм, рядов ареол 16–20 в 10 мкм, ареол в ряду 18–22 в 10 мкм.
Рис. 1.
Электронные микрофотографии створок (СЭМ): а – Actinocyclus normanii, б – Aulacoseira ambigua, в – A. granulata, г – A. islandica, д – A. pusilla, е – A. subarctica, ж – Conticribra weissflogii, з, и – Cyclostephanos dubius, к, л – C. invisitatus, м – Cyclotella atomus var. atomus, н – C. atomus var. gracilis, о – C. cryptic. а–з, к – наружная поверхность створок; и, л–о – внутренняя.

Aulacoseira ambigua (Grunow) Simonsen (рис. 1б). Створки диаметром 13–28.7 мкм, ареол в ряду 10–14 в 10 мкм.
A. granulata (Ehrenberg) Simonsen (рис. 1в). Створки диаметром 8.2–30.5 мкм, высотой 11.4–43.3 мкм, рядов ареол 4–14 в 10 мкм, ареол в ряду 4–14 в 10 мкм.
A. islandica (O. Müller) Simonsen (рис. 1г). Створки диаметром 9.2–21.2 мкм, высотой 8.7–12.5 мкм, рядов ареол 12–16 в 10 мкм, ареол в ряду 10–20 в 10 мкм.
*A. pusilla (F. Meister) Tuji et Houki (рис. 1д). Створки диаметром 5.2–6.9 мкм, высотой 2.4–5.2 мкм, рядов ареол 25–30 в 10 мкм, ареол в ряду 30–40 в 10 мкм.
A. subarctica (O. Müller) Hawortrh (рис. 1е). Створки диаметром 6–6.6 мкм, высотой 6.8–11.3 мкм, рядов ареол 20 в 10 мкм, ареол в ряду 20–25 в 10 мкм.
Conticribra weissflogii (Grunow) K. Stachura-Suchoples et D.M. Williams (рис. 1ж). Створка диаметром 15.6 мкм, краевых выростов 14 в 10 мкм.
Cyclostephanos dubius (Fricke) Round (рис. 1з, 1и). Створки диаметром 9.7–10 мкм, штрихов 10–14 в 10 мкм.
C. invisitatus (Hohn et Hellerman) Theriot, Stoermer et Häkansson (рис. 1к, 1л). Створки диаметром 8.3–12 мкм, штрихов 15–20 в 10 мкм.
Cyclotella atomus Hustedt var. atomus (рис. 1м). Створки диаметром 5.0–6.4 мкм, штрихов 12–15 в 10 мкм.
C. atomus var. gracilis Genkal et Kiss (рис. 1н). Створки диаметром 5.0–6.4 мкм, штрихов 16–20 в 10 мкм.
*C. cryptica Reimann, Lewin et Guillard (рис. 1о). Створка диаметром 5.9 мкм, штрихов 8 в 10 мкм.
C. distinguenda Hustedt (рис. 2а). Створки диаметром 11.6–18.7 мкм, штрихов 10–18 в 10 мкм.
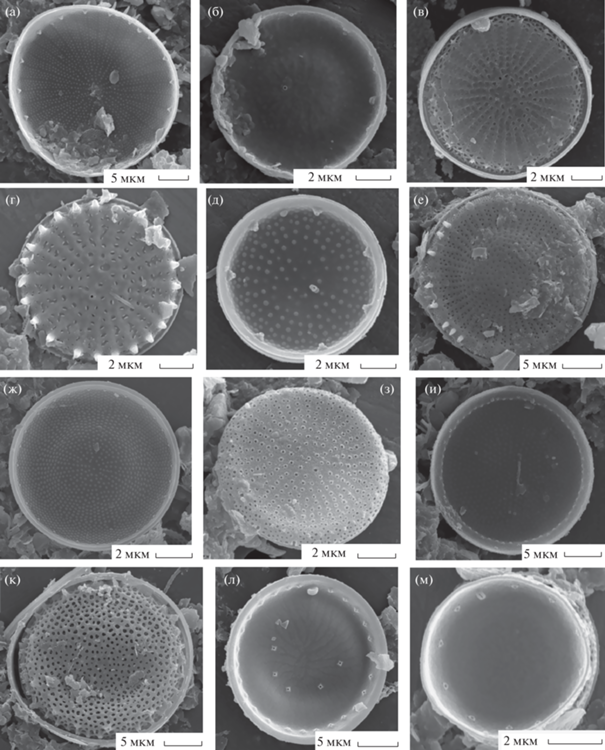
Рис. 2.
Электронные микрофотографии створок (СЭМ): а – Cyclotella distinguenda, б, в – C. meduanae, г, д – C. meneghiniana, е, ж – Discostella pseudostelligera, з – Melosira varians, и – Skeletonema subsalsum, к – Stephanodiscus binderanus, л – S. delicatus, м – S. hantzschii. а, б, г, е, з, и, м – наружная поверхность створок; в, д, ж, к, л – внутренняя.
C. meduanae Germain emend. Genkal (рис. 2б, 2в). Створки диаметром 6.6–7.7 мкм, штрихов 10–12 в 10 мкм.
C. meneghiniana Kützing (рис. 2г, 2д). Створки диаметром 8.3–23.4 мкм, штрихов 8–9 в 10 мкм.
Discostella pseudostelligera (Hustedt) Houk et Klee (рис. 2е, 2ж). Створки диаметром 3.6-6.8 мкм, штрихов 25–35 в 10 мкм.
Melosira varians Agardh (рис. 2з). Створки диаметром 11.8–27 мкм, высотой 11–14.3 мкм.
Skeletonema subsalsum (Cleve-Euler) Bethge (рис. 2и). Створки диаметром 4.8–9.5 мкм, высотой 1.5–1.7 мкм.
Stephanodiscus binderanus (Kützing) Krieger (рис. 2к). Створки диаметром 4.8–9.5 мкм, высотой штрихов 15 в 10 мкм.
S. delicatus Genkal (рис. 2л). Створки диаметром 8.8–9 мкм, штрихов 15 в 10 мкм.
S. hantzschii Grunow (рис. 2м, 3а). Створки диаметром 9.7–20 мкм, штрихов 6–9 в 10 мкм.
*S. lacustris Klee et Houk (рис. 3б). Створки диаметром 4.2–7.2 мкм, штрихов 17–25 в 10 мкм.
Рис. 3.
Электронные микрофотографии створок (СЭМ): а – Stephanodiscus hantzschii, б – S. lacustris, в – S. makarovae, г, д – S. minutulus, е, ж – S. neoastraeae, з – S. triporus, и – Thalassiosira faurii, к, л – T. lacustris, м – T. pseudonana. а, б, д, ж, и, л, м – внутренняя поверхность створок; в, г, е, з, к – наружная.
S. makarovae Genkal (рис. 3в). Створки диаметром 6.4–7.7 мкм, штрихов 20–25 в 10 мкм.
S. minutulus (Kützing) Cleve et Möller (рис. 3г, 3д). Створки диаметром 7.1–8.9 мкм, штрихов 12–16 в 10 мкм.
S. neoastraea Häkansson et Hickel emend. Casper, Scheffler et Augsten (рис. 3е, 3ж). Створки диаметром 12.7–28.5 мкм, штрихов 8–10 в 10 мкм.
S. triporus Genkal et Kuzmin emend. Genkal, K.T. Kiss et Ács (рис. 3з). Створки диаметром 5.0–8.3 мкм, штрихов 15–20 в 10 мкм.
Thalassiosira faurii (Gasse) Hasle emend. Genkal (рис. 3и). Створки диаметром 17.7–19.3 мкм, краевых выростов 12–14 в 10 мкм.
T. lacustris (Grunow) Hasle (рис. 3к, 3л). Створки диаметром 18.7–33.6 мкм, краевых выростов 6–8 в 10 мкм.
T. pseudonana Hasle et Heimdal (рис. 3м). Створки диаметром 4.1–5.6 мкм, на створке 9–11 краевых выростов.
В последнем опубликованном списке приводится 38 видов, разновидностей и форм центрических диатомовых водорослей (Корнева, 2015), в настоящем исследовании. Обнаружено отсутствие следующих таксонов: Acanthoceras zachariasii (Brun) Simonsen, Aulacoseira distans (Ehrenberg) Simonsen, A. muzzanensis (Meister) Krammer, Cyclotella bodanica Eulenstein ex Grunow, C. planctonica Brunnthaler, C. radiosa (Grunow) Lemmerman, Rhizosolenia longiseta Zacharias, Skeletonema potamos (Weber) Hasle, Stephanodiscus invisitatus f. hakanssoniae Genkal et Kiss, S. triporus var. volgensis Genkal, S. cf. skabitschevskyi Popovskaya, Thalassiosira incerta Makarova.
Acanthoceras zachariasii и Rhizosolenia longiseta имеют нежную структуру панциря, и возможно, что по этой причине, после подготовки водорослей к электронной микроскопии в процессе изучения препаратов в СЭМ, их фиксация не удалась, кроме того они не отмечены при количественной обработке проб. Aulacoseira distans относится к ископаемым видам (Houk et al., 2017), в водоемах и водотоках России по данным электронной микроскопии не зафиксирована (Генкал и др., 2020). Идентифицированная в р. Волга с помощью световой микроскопии A. distans по результатам электронно-микроскопического изучения, по нашему мнению, относится к другому сходному низкопанцирному виду – A. subarctica.
Ряд видов Coscinodiscaceae сведен в синонимику: Aulacoseira muzzanensis к A. granulata (Genkal, Trifonova, 2020), Cyclotella planctonica к С. schroeteri (Schröter) Lemmermann (Houk et al., 2017), которая по данным электронной микроскопии не выявлена в водных объектах России (Генкал и др., 2020), C. radiosa – к Handmannia comta (Ehrenberg) Kociolek et Khursevich (Генкал и др., 2020), Stephanodiscus triporus var. volgensis – к типовой разновидности (Kiss et al., 2013; Генкал и др., 2020), а S. cf. skabitschevskyi – к Cyclostephanos dubius (Генкал и др., 2020). Cyclotella bodanica переведена в другой род Handmannia – H. bodanica (Eulenstein ex Grunow) Kociolek et Khursevich (Khursevich, Kociolek, 2012; Генкал и др., 2020). По современным представлениям, у Stephanodiscus invisitatus рядность штрихов варьирует от двухрядных до трехрядных (Генкал и др., 2020), поэтому форму с трехрядными штрихами (S. invisitatus f. hananssoniae Genkal et Kiss) не учитывали в качестве самостоятельной таксономической единицы.
Для трассы строящегося Чебоксарского водохранилища указаны два сходных по морфологии вида Cyclotella stelligera и C. pseudostelligera и в качестве одного из типичных представителей планктона р. Волга опубликована трансмиссионная электронная микрофотография последнего (Кузьмин, 1978, рис. 41: 12). Позднее для р. Волга был приведен только один вид C. stelligera, а C. pseudostelligera указан как синоним первого, а на ТЭМ и СЭМ иллюстрациях показан последний (Генкал, 1992: табл. XVII, 3–6; XVIII, XIX; XX, 1–5). Вероятно, именно по этой причине в последующих публикациях (Генкал и др., 2020) в списке по водохранилищу приводится С. stelligera. Отсутствие в них иллюстраций этого вида и его находок при изучении нашего многолетнего материала из разных участков водоема требует подтверждения присутствия С. stelligera. В работе (Корнева, 2015) также приводится Thalassiosira weissflogii (Grunow) Fryxell et Hasle, которая переведена в род Conticribra – C. weissflogii (Grunow) K. Stachura-Suchoples et D.M. Williams (Генкал и др., 2020).
Ниже приводится список Coscinodiscaceae фитопланктона Чебоксарского водохранилища c учетом результатов нашего изучения и новых данных по их таксономии: Actinocyclus normanii, Acanthoceras zachariasii, Aulacoseira ambigua, A. granulata, A. islandica, A. pusilla, A. subarctica, Conticribra weissflogii, Cyclostephanos dubius, C. invisitatus, Cyclotella atomus var. atomus, C. atomus var. gracilis, C. cryptica, C. distinguenda, C. meduanae, C. meneghiniana, Discostella pseudostelligera, Handmannia bodanica, H. comta, M. varians, Rhizosolenia longiseta, Skeletonema potamos, S. subsalsum, Stephanodiscus binderanus, S. delicatus, S. hantzschii, S. lacustris, S. makarovae, S. minutulus, S. neoastraea, S. volgensis Genkal et Korneva, Thalassiosira faurii, T. incerta, T. lacustris, T. pseudonana (Kobayasi et al., 1985; Theriot et al., 1987a, 1987b; Kiss, 1988; Krammer, Lange-Bertalot, 1991; Casper et al. 1992; Håkansson, 2002; Houk, Klee, 2004; Stachura-Suchoples, Williams, 2009; Houk et al, 2010, 2014, 2017; Kiss et al., 2012; Генкал и др., 2020).
В современный период диатомовые водоросли (в основном центрические) по-прежнему относятся к доминирующей группе водорослей в планктонных альгоценозах водохранилища (табл. 1). После аномально жаркого 2010 г. продуктивность фитопланктона и диатомовых водорослей в его составе по сравнению с первым десятилетием ХХI в. сильно выросла. Так, общие показатели биомассы в озерном районе водохранилища возросли в пять раз, в верхнем речном – в девять, тогда как показатели численности в каждом из этих районов водоема увеличились в >25 раз. Биомасса диатомовых водорослей, в том числе и центрических, также выросла, но менее заметно (в 5–6 раз в озерном и 3–4 раза в верхнем речном). Меньшие изменения общих показателей численности (рост в 1.8 раз) и биомассы (в 2 раза) фитопланктона отмечены в районе от устья р. Ока до устья р. Сура, где обилие диатомовых также увеличилось в 2–3 раза. Резкий рост общей численности фитопланктона на фоне менее значительных изменений обилия диатомей в летнем фитопланктоне водоема, по-видимому, – следствие усиления процессов “цветения” воды в Горьковском водохранилище и в озерном районе Чебоксарского. При активизации роста альгоценозов планктона после аномально жаркого 2010 г. роль диатомей, и среди них центрических, в летних сообществах в общей численности заметно снизилась в верхнем речном (в 6.0–7.3 раза) и общей биомассе в 1.5–1.8 раз) и озерном районах (в 6.7–7.5 и 1.3–1.6 раз соответственно) водоема. При этом, центрические диатомеи уменьшили относительное обилие и в общих его показателях для всего фитопланктона, и для таксоценоза Bacillariophyta.
Таблица 1.
Доля (%) диатомовых водорослей и Centrophyceae в численности (N) и биомассе (B) летнего фитопланктона в разных районах водохранилища до и после аномально жаркого 2010 г.
| Показатели обилия | ВР | Р | О | УО | УКУ | УС | УК | УВ | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | I | II | I | II | I | II | II | II | II | II | |
| Nдиат/Nобщ | 17.3 | 2.9 | 9.9 | 18.2 | 22.9 | 3.4 | 29.8 | 45.9 | 11.4 | 13.5 | 23.7 | 6.9 |
| Nцентр/Nобщ | 12.4 | 1.7 | 9.3 | 17.3 | 15.0 | 2.0 | 29.3 | 45.4 | 10.8 | 13.1 | 21.1 | 6.3 |
| Nцентр /Nдиат | 71.4 | 59.0 | 94.4 | 95.1 | 65.8 | 58.7 | 98.1 | 98.9 | 94.3 | 97.4 | 92.0 | 91.4 |
| Bдиат/Bобщ | 60.4 | 33.7 | 71.8 | 80.2 | 62.3 | 40.1 | 80.1 | 85.8 | 72.8 | 32.2 | 93.3 | 72.2 |
| Bцентр/Bобщ | 39.6 | 26.0 | 66.1 | 78.0 | 33.8 | 26.6 | 78.9 | 84.4 | 69.1 | 31.5 | 92.0 | 70.7 |
| Bцентр/Bдиат | 65.5 | 77.1 | 92.1 | 97.3 | 54.2 | 66.2 | 98.5 | 98.3 | 94.9 | 97.8 | 98.6 | 97.9 |
Примечание. Районы водохранилища: ВР – верхний речной; Р – речной; О – озерный; УО – устье р. Ока; УКУ – устье р. Кудьма; УС – устье р. Сура; УК – устье р. Керженец; УВ – устье р. Ветлуга. I – 2000–2010 гг.; II – 2011–2020 гг. Подстрочные сокращения: диат. – диатомовые водоросли, центр. – центрические. Даны средние значения за исследованные периоды.
В речном отделе диатомовые, в том числе и центрические водоросли, до 2010 г. достигали 9–10% численности и 66–72% биомассы фитопланктона, в следующем десятилетии относительная их численность возросла почти в 2 раза, относительная биомасса существенно (увеличение лишь в 1.1–1.2 раза) не изменилась. При этом средний ценотический объем клетки диатомовых, судя по отношению биомассы к их численности, в 1.43 раза стал меньше, чем в первую половину периода наблюдений. Роль центрических водорослей в формировании численности таксоценоза диатомовых водорослей в зоне воздействия вод Горьковского водохранилища (до устья р. Окa) снизилась на ~10% во второй половине периода наблюдений (с 71 до 59%), однако их относительная биомасса выросла с 66 до 77%. В речном районе доля этой группы в общей биомассе фитопланктона достигала максимальных значений для водоема (78% во второй половине ряда наблюдений). Их роль в организации таксоценоза диатомей оставалась максимальной в зоне воздействия окских вод (92–97%) и несколько ниже в районах до устья р. Окa (66–77%) и в озерном (54–66%). Таким образом, в этих районах примерно треть численности и треть биомассы диатомовых водорослей образовано пеннатными формами (Asterionella, Diatoma, Synedra и др.) Максимальные численность этой группы (473.0 и 439.0 млн кл./л в верхнем речном и озерном районах соответственно) и биомасса (30.0–31.0 г/м3) отмечали в речном и озерном участках, их значения свойственны эвтрофно-гипертрофным водным объектам.
По сравнению с периодом до создания водохранилища (1969–1980 гг.), на участке до устья р. Окa интегральная биомасса фитопланктона после 2010 г. выросла в 1.5 раза, доля диатомовых в ее составе незначительно снизилась (табл. 2). Уровень развития планктонных альгоценозов от устья р. Окa до впадения р. Сурa в современный период лишь в 1.2 раза ниже, чем в 1969–1980 гг., когда этот участок р. Волги подвергался самому мощному влиянию загрязнения и эвтрофирования. Роль диатомовых (в основном центрических) в этом районе за пятьдесят лет наблюдений не изменилась, оставаясь максимально высокой (80–83%) из всех охарактеризованных временных интервалов в результате следствием воздействия окских вод на этот район р. Волги. Более существенные изменения общей продуктивности фитопланктона и относительной биомассы Bacillariophyta отмечены ниже устья р. Сурa (озерный район), где интегральные показатели вегетации летних фитоценозов и доля диатомей в них, по сравнению с незарегулированным состоянием, снизились в 3.1 и 2.2 раза соответственно. После периода резкой депрессии биомассы планктонных водорослей, вследствие перестройки структуры сообществ на лимнофильные, здесь во вторую декаду XXI в. вновь отмечается подъем вегетации фитопланктона, но в летних ценозах диатомовые уже уступают преимущество другим группам водорослей, в основном, цианопрокариотам.
Таблица 2.
Средняя биомасса (г/м3), летнего фитопланктона (над чертой) и доля (%) диатомовых водорослей в ней (под чертой) за разные периоды наблюдений
| Период наблюдений, гг. | Плотина Горьковской ГЭС – устье р. Ока | Устье р. Ока – устье р. Сура |
Устье р. Сурa – Чебоксары |
|---|---|---|---|
| 1969–1980 | $\frac{{2.81}}{{40}}$ | $\frac{{9.35}}{{83}}$ | $\frac{{11.35}}{{88}}$ |
| 1981–1990 | $\frac{{2.04}}{{36}}$ | $\frac{{6.55}}{{76}}$ | $\frac{{2.72}}{{58}}$ |
| 2000–2010 | $\frac{{0.48}}{{60}}$ | $\frac{{3.86}}{{72}}$ | $\frac{{0.77}}{{62}}$ |
| 2011–2020 | $\frac{{4.15}}{{34}}$ | $\frac{{7.82}}{{80}}$ | $\frac{{3.69}}{{40}}$ |
В устьевом районе р. Окa динамика развития фитопланктона и, в частности, диатомовых водорослей, в XXI в. более стабильна, чем на основной акватории водохранилища (рис. 4 ). Количественные показатели альгоценозов здесь свойственны мезотрофно-эвтрофным водным объектам, а значение диатомовых, в том числе и центрических водорослей, в общих величинах обилия фитопланктона максимально из всех выделенных районов водоема. В альгоценозах планктона других притоков при определяющей роли центрических диатомовых водорослей (>90–99% обилия диатомей) их вклад в общие характеристики вегетации фитопланктона в разных реках колебался от 6.3 (р. Ветлуга) до 21.1% (р. Керженец) (по численности) и от 31.5% (р. Сура) до 92.0% (р. Керженец) (по биомассе) и был минимален в устье р. Сурa. В р. Керженец их роль была более заметна, а в р. Сурa – диатомовые и центрические (1/3 треть биомассы и 13.1–13.5% численности) уступали другим группам водорослей. Таким образом, летом наблюдали постепенное снижение значимости (вклада) диатомовых водорослей и в их составе центрических в структуре количественных показателей фитопланктона. Эти изменения менее заметны в верховьях водохранилища и более значимы в озерном расширении. Кроме того, отмечены низкие значения относительной численности диатомовых водорослей, включая и центрические, в устьях притоков. По-видимому, это связано с возрастанием роли мелкоклеточных фракций (в основном, цианопрокариот), особенно в устье р. Ветлугa, или других групп (зеленых, динофитовых, эвгленовых) в р. Сурa.
Постепенное измельчание клеток водорослей планктона в современный период существования водохранилища, особенно сильно выраженное в верхнем речном районе (в 3.2 раза) и озерном (в >5 раз), в том числе и диатомовых (в 1.43 раза в речном, 1.32 раза в озерном) по сравнению с первым десятилетием века, – еще один показатель мощных перестроек структуры фитопланктона водоема при росте его трофического статуса с мезотрофного уровня в 2000–2010 гг. до мезотрофно-эвтрофного и эвтрофного на современном этапе. Уменьшение размерных характеристик водорослей по многолетним данным (2001–2019 гг.) отмечено и в Волгоградском водохранилище (Шашуловская и др., 2021).
Среди доминирующих компонентов альгоценозов по-прежнему лидируют виды родов Stephanodiscus, Aulacoseira, Melosira и Cyclotella. Средние и максимальные значения их численности и биомассы после 2010 г. зарегистрированы в речном районе водохранилища от устья р. Окa до устья р. Сурa. Например, биомасса Aulacoseira granulata в первые годы существования водоема достигала в летние пики ее развития 2.9–5.6 г/м3, а в современный период – до 11.1 г/м3. У Aulacoseira ambigua максимальные значения обилия в 1980-х гг. обычно были 0.4–0.6 г/м3, единично – до 5.8 г/м3, сейчас – до 18.1 г/м3. В то же время, биомасса Aulacoseira subarctica остается примерно на уровне 1980-х годов (до 0.3–0.4 г/м3 реже более, до 0.45 г/м3 в современный период). Melosira varians в летних альгоценозах водоема в первое десятилетие после зарегулирования стока (≤0.9 г/м3) не играла ведущей роли в летних фитоценозах. На современном этапе сукцессии фитопланктона она, по-видимому, усиливает свои позиции (≤3.1 г/м3 в речном и ≤2.75 г/м3 в озерном), иногда достигая массового развития (в устье р. Керженец – ≤32.6 г/м3). Максимальное обилие важнейшего компонента фитопланктона Stephanodiscus hantzschii в сравнительном аспекте изменилось незначительно (с 2.3–5.7 и до 4.0–5.4 г/м3), у Cyclotella meneghiniana выросло с 1.3–14.7 до 18.2 г/м3. Другие виды рода Stephanodiscus (S. invisitatus, S. minutulus, S. neoastraea и др.) либо мало изменили уровень максимальной продуктивности, либо наблюдалась тенденция к его снижению.
Среди инвазивных видов в современный период наиболее заметен Actinocyclus normannii (до 2.31 г/м3 верхнем речном участке, 1.1 г/м3 в устье р. Ока, 0.87 г/м3 в р. Кудьма и 0.49 г/м3 в р. Сура). Thalassiosira incerta на основной акватории водоема встречалась с незначительной биомассой, развиваясь в р. Кудьме (до 2.69 г/м3) и р. Керженец (до 1.33 г/м3). Skeletonema subsalsum после периода подъема обилия в 1980-х гг. (7.4 г/м3) стабилизировала развитие на уровне 0.12–0.16 г/м3 в верхнем речном и озерном районах водоема и до 0.10–0.19 г/м3 в устьях его притоков. Кроме перечисленных планктонных видов, в системе боковой приточности левобережья отмечен солоновато-водный бентосный Plagiotropis lepidoptera (Gregory) Kuntze (Кулизин и др., 2021).
Таким образом, на фоне макроклиматических изменений в бассейне р. Волга во второй декаде XXI в. можно отметить мощную сукцессию в альгоценозах Чебоксарского водохранилища, заключающуюся в масштабных изменениях общей динамики фитопланктона и развития диатомовых водорослей (включая центрические). В доминирующем комплексе диатомей отмечено усиление позиций представителей рода Aulacoseira, Melosira varians, Cyclotella meneghiniana, стабилизация Stephanodiscus hantzschii и тенденция к снижению роли других видов этого рода. При этом изменялось соотношение показателей обилия и относительного участия диатомовых в структуре фитопланктона, развитие и укрепление позиций одних инвазивных видов (Actinocyclus, Thalassiosira), относительная стабилизация других (Skeletonema) и появление новых (Plagiotropis и др.).
Выводы. Современный состав Coscinodiscaceae Чебоксарского водохранилища включает 29 видов и разновидностей родов Aulacoseira, Cyclotella и Stephanodiscus, три из них – новые для альгофлоры водоема. Уточнены систематическое положение ряда таксонов и видовой состав этой группы диатомей. В первые два десятилетия ХХI в. в водохранилище наблюдается резкое возрастание продуктивности фитопланктона летом с мезотрофного уровня в 2000–2010 гг. до мезотрофно-эвтрофного и эвтрофного на современном этапе, и заметное уменьшение относительной роли диатомовых (и среди них центрических) в формировании показателей его обилия. Так, установленные в последней четверти ХХ в. относительные численность (25–62%) и биомасса (40–90%) диатомовых летнего фитопланктона водохранилища в современный период его существования заметно снизились (соответственно 3–18 и 34–80%), что косвенно свидетельствует об усилении после 2010 г. процессов “цветения воды” цианопрокариотами и перестройке размерной структуры сообществ. На фоне существенных макроклиматических изменений в бассейне р. Волгa наблюдается тенденция к исчезновению ряда представителей центрических диатомовых водорослей из летних альгоценозов водохранилища и обеднению его состава, а также появление новых, не встречавшихся ранее видов. Эти изменения вызваны продолжающимся эвтрофированием, влияние которого заметно увеличилось после аномально жаркого и маловодного 2010 г.
Список литературы
Балонов И.М. 1975. Подготовка водорослей к электронной микроскопии // Методика изучения биогеоценозов. Москва: Наука. С. 87.
Генкал C.И. 1992. Атлас диатомовых водорослей планктона реки Волги. Cанкт-Петербург: Гидрометеоиздат.
Генкал C.И., Куликовский М.C., Кузнецова И.В. 2020. Современные пресноводные центрические диатомовые водоросли России. Ярославль: Филигрань.
Корнева Л.Г. 2015. Фитопланктон водохранилищ бассейна Волги. Кострома: Костромской печатный дом.
Корнева Л.Г., Генкал С.И. 2000. Таксономический состав и эколого-географическая характеристика фитопланктона волжских водохранилищ // Каталог растений и животных водоемов бассейна Волги. Ярославль: Изд-во ЯГТУ. С. 5.
Кузьмин Г.В. 1978. Водоросли // Волга и ее жизнь. Ленинград: Наука. С. 299.
Кузьмин Г.В., Охапкин А.Г. 1975. Фитопланктон р. Волги на трассе строительства Чебоксарского водохранилища и прогноз его альгологического режима // Антропогенные факторы в жизни водоемов. Ленинград: Наука. С. 32.
Кулизин П.В., Воденеева Е.Л., Охапкин А.Г. 2021. Фитопланктон некоторых рек юга подзоны хвойно-широколиственных лесов бассейна Средней Волги в многолетнем аспекте // Самарск. науч. вестник. Т. 10. № 2. С. 45. https://doi.org/10.17816/snv2021102106
Методика изучения биогеоценозов внутренних водоемов. 1975. Москва: Наука.
Охапкин А.Г. 1994. Фитопланктон Чебоксарского водохранилища. Тольятти: Самарский науч. центр. РАН.
Охапкин А.Г., Генкал C.И. 1989. Центрические диатомовые водоросли в планктоне Чебоксарского водохранилища // Биология внутр. вод: Информ. бюл. Ленинград. № 83. С. 12.
Шашуловская Е.А., Мосияш С.А., Далечина И.Н. 2021. Многолетние изменения основных показателей трофического состояния крупного равнинного водохранилища под влиянием климатической трансформации и сукцессионных процессов // Биология внутр. вод. № 6. P. 547. https://doi.org/https://doi.org/10.31857/S0320965221060164
Юлова Г.А. 1982. Фитопланктон р. Волги от Городца до Чебоксар: Автореф. дис. … канд. биол. наук. Ленинград: Ботан. ин-ут РАН.
Casper S.J., Scheffler W., Håkansson H. 1992. Stephanodiscus neoastraea Håkansson et Hickel (Bacillariophyta, Centrales) in norddeutschen Seen und Flüssen // Arch. Protistenkd. V. 142 (3–4). S. 193. https://doi.org/10.1016/S0003-9365(11)80084-9
Genkal S.I., Trifonova I.S. 2020. Morphology and taxonomy of Aulacoseira muzzanensis (Bacillariophyta) // Новости систематики низших растений. Т. 54. № 2. P. 355. https://doi.org/10.31111/nsnr/2020.54.2.355
Håkansson H. 2002. A compilation and evaluation of species in the general Stephanodiscus, Cyclostephanos and Cyclotella with anew genus in the family Stephanodiscaceae // Diatom Res. V. 17 (1). P. 1. https://doi.org/10.1080/0269249X.2002.9705534
Houk V., Klee R. 2004. The stelligeroid taxa of the genus Cyclotella (Kützing) Brébisson (Bacillariophyceae) and their transfer into the genus Discostella gen.nov. // Diatom Res. V. 19 (2). P. 204. https://doi.org/10.1080/0269249X.2004.9705871
Houk V., Klee R., Tanaka H. 2010. Atlas of freshwater centric diatoms with a brief key and descriptions. Part III. Stephanodiscaceae. A. Cyclotella, Tertiarius, Discostella // Fottea. Suppl. V. 10. P. 1.
Houk V., Klee R., Tanaka H. 2014. Atlas of freshwater centric diatoms with a brief key and descriptions. Part IV. Stephanodiscaceae B. Stephanodiscus, Cyclostephanos, Pliocaenicus, Hemistepanos, Stephanocostis, Mesodictyon & Spaticribra // Fottea. Suppl. V. 14. P. 1.
Houk V., Klee R., Tanaka H. 2017. Atlas of freshwater centric diatoms with a brief key and descriptions.Second emended edition of Part I and II. Melosiraceae, Orthoseraceae, Paraliaceae and Aulacoseiraceae // Fottea. Suppl. 17. P. 1.
Khursevich G., Kociolek J.P. 2012. A preliminary, worldwide inventory of the extinct, freshwater fossil diatoms from the orders Thalassiosirales, Stephanodiscales, Paraliales, Aulacoseirales, Melosirales, Coscinodiscales, and Biddulphiales // Nova Hedwigia. V. 141. P. 315.
Kiss K.T. 1988. The Morphology and Taxonomy of Stephanodiscus invistatus Hohn et Hellerman (Bacillariophyceae) // Arh. Protistenkd. V. 135. P. 187.
Kiss K.T., Genkal S.I., Ector L. et al. 2013. Morphology, taxonomy and distribution of Stephanodiscus triporus (Bacillariophyceae) and related taxa // Eur. J. Phycol. V. 48 (4). P. 363. https://doi.org/10.1080/09670262.2013.843204
Kiss K.T., R. Klee, L. Ector, É. Ács. 2012. Centric diatoms of large rivers and tributaries in Hungary: morphology and biogeographic distribution // Acta Bot. Croat. V. 71 (2). P. 311.
Kobayasi H., Kobayashi H., Masahiko I. 1985. Fine structure and taxonomy of the small and tiny Stephanodiscus (Bacillariophyceae) species in Japan. 3. Co-occurence of Stephanodiscus minutulus (Kütz.) Round and S.parvus Stoerm. et Håk. // Jap. Phycol. V. 33 (4). P. 293.
Krammer K., Lange-Bertalot H. 1991. Bacillariophyceae. Teil 3: Centrales, Fragilariaceae, Eunotiaceae // Süsswasserflora von Mitteleuropa. Stuttgart: Gustav Fischer Verlag. Bd 2. H. 3.
Theriot E., Håkansson H., Kociolek J.P., Round F.E., Stoermer E.F. 1987a. Validation of the centric diatom genus name Cyclostephanos // Br. Phycol. J. V. 22 (4). P. 345. https://doi.org/10.1080/00071618700650411
Theriot E., Stoermer E.F., Håkansson H. 1987b. Taxonomic interoretation of the rimoportula of freshwater genera in the centric diatom family Thalassiosiraceae // Diatom Res. V. 2 (2). P. 251. https://doi.org/10.1080/0269249X.1987.9705003
Stachura-Suchoples K., Williams D.M. 2009. Description of Conticribra tricircularis, a new genus and species of Thalassiosirales, with a discussion on its relationship to other continuous cribra species of Thalassiosira Cleve (Bacillariophyta) and freshwater origin // Eur. J. Phycology. V. 44 (4). P. 477.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод