

Ботанический журнал, 2019, T. 104, № 1, стр. 12-40
ОЛИГОТРОФНЫЕ СФАГНОВЫЕ И СФАГНОВО-ЗЕЛЕНОМОШНЫЕ ЕЛЬНИКИ ЕВРОПЕЙСКОЙ РОССИИ И УРАЛА
И. Б. Кучеров 1, , *, С. А. Кутенков 2, ***
1 Ботанический институт им. В.Л. Комарова Российской академии наук
197376 Санкт-Петербург, ул. Проф. Попова, 2, Российская Федерация
2 Институт биологии Карельского научного центра РАН
185910 г. Петрозаводск, ул. Пушкинская, 11, Российская Федерация, Республика Карелия
* E-mail: atragene@mail.ru
*** E-mail: effort@krc.karelia.ru
Поступила в редакцию 01.03.2018
После доработки 09.12.2018
Принята к публикации 11.12.2018
Аннотация
На основе 260 геоботанических описаний, выполненных авторами в 1995–2017 гг., а также взятых из литературы, проведена доминантно-детерминантная классификация заболоченных олиготрофных ельников (из Picea abies s. l.) сфагновых и сфагново-зеленомошных Европейской России и Урала (с учетом отдельных описаний с севера Западной Сибири), в результате чего выделено 11 синтаксонов разного ранга. Ельники вороничные зеленомошно-сфагновые, ерниковые и голубичные сфагновые характерны для северной тайги, включая ее северную полосу (крайнесеверную тайгу), тогда как багульниковые сфагновые произрастают в средней тайге и южной полосе подзоны северной тайги, не проникая севернее. Ельники деренно-морошковые сфагновые выявлены лишь в низкогорьях Северно-Западной Карелии. Ельники черничные сфагновые, представленные различными вариантами и модификациями, распространены наиболее широко наряду с пушицевыми сфагновыми, развивающимися при подтоплении. В моховом ярусе лесов всех названных синтаксонов доминирует Sphagnum angustifolium в сопровождении S. russowii, иногда также S. fuscum.
Ельники сфагновые (в широком смысле, включая сфагново-зеленомошные) – наиболее важная, но все еще малоизученная группа ассоциаций заболоченных еловых (из Picea abies s. l.) лесов. Площадь, занятая ими, различается в разных регионах. В Мурманской обл. доля площади, занятой подобными лесами, составляет лишь 4.6% от общей площади еловых лесов (Melekhov, 1966). Но уже в северной тайге Карелии леса данной группы ассоциаций занимают 8% от всей лесопокрытой площади и 34% от общей площади ельников. В средней тайге Карелии аналогичные цифры составляют 12% и 26% (Gromtsev, 2008). На севере Русской равнины распространенность заболоченных ельников возрастает к востоку и северо-востоку. В северной тайге Архангельской обл. только на долю ельников “долгомошных” (сфагново-зеленомошных) приходится 30% от общей площади еловых лесов. В полосе крайнесеверной тайги эта величина снижается до 25%. Зато доля площади, занятой собственно ельниками сфагновыми, достигает 13% (Rysin, Savelieva, 2002). В крайнесеверной тайге Республики Коми на ельники долгомошные приходится 21% от общей площади еловых лесов, на сфагновые – 26% без учета ерниковых (с ярусом Betula nana) ельников, которые занимают еще 19% (Bobkova, Patov, 2006). В южной полосе северной тайги Коми ельники долгомошные произрастают на 46%, сфагновые – на 16% от общей площади еловых лесов (Bobkova, 2006a), в средней тайге, где заболоченность ниже, – соответственно на 37% и 12% (Bobkova, 2006b).
Независимо от занимаемой площади, для ельников сфагновых характерен высокий уровень разнообразия сообществ, что обусловлено различными сочетаниями условий переувлажнения и богатства почв, порождающими многообразие режимов заболачивания. Тем не менее, ценотическое разнообразие этих лесов в большинстве регионов выявлено недостаточно полно по сравнению с ельниками зеленомошными и даже травяными. Основная проблема при этом связана с необходимостью идентификации видов мохового яруса, которые в заболоченных лесах являются лучшими индикаторами экологических режимов. Часто наблюдаются “близнецовые” ассоциации, входящие в разные ряды заболачивания и различающиеся по доминирующим, но плохо распознаваемым в полевых условиях видам сфагновых мхов при высоком уровне сходства вышележащих ярусов. Как правило, такие ассоциации не распознаются и объединяются в один экологически гетерогенный синтаксон.
В настоящей статье авторы стремятся выявить ценотическое разнообразие ельников сфагновых и сфагново-зеленомошных, развитых в условиях олиготрофного заболачивания, с господством Sphagnum angustifolium и иных экологически близких ему ацидофильных видов мхов, а также географическое распространение этих сообществ в Европейской России, на Урале и на сопредельных территориях.
ИСПОЛЬЗОВАННЫЕ ДАННЫЕ И МЕТОДЫ
Нами использовано 260 геоботанических описаний олиготрофных сфагновых и сфагново-зеленомошных ельников таежной зоны Европейской России и Урала. Из общей выборки 194 описания сделаны авторами в экспедициях 1996–2017 гг. В их число входит и несколько описаний, выполненных С.А. Кутенковым на материковом юге Ямала, что расширяет наши представления об ареале соответствующих синтаксонов. Однако этих описаний явно недостаточно, чтобы упоминать в названии работы Западную Сибирь.
Авторские описания выполнялись по стандартной геоботанической методике (Ponyatovskaya, 1964; Shennikov, 1964; Dylis, 1974) с оценкой проективных покрытий (ПП) видов по ярусам на площади не менее 400 м2. В каждом из описаний делалась почвенная прикопка для измерения мощности подстилки и органолептической оценки гранулометрического состава почвы либо измерялась мощность торфяной залежи (Kucherov et al., 2010; Kucherov, Kutenkov, 2011, 2012; Kucherov, 2018).
Часть описаний из крайнесеверной и северной тайги Республики Коми (26) взята из рукописи докторской диссертации Ю.П. Юдина (Yudin, 1948), хранящейся в библиотеке БИН РАН. Содержание диссертации опубликовано после смерти ее автора (Yudin, 1954) без публикации исходных данных. Еще 40 описаний опубликованы в литературе (Regel, 1923; Soczava, 1927, 1930; Sambuk, 1932; Andreyev, 1935; Nekrasova, 1935; Blagoveshchenskii, 1936; Sokolova, 1936; Lyubimova, 1937; Korchagin, 1940, 1956; Nepomilueva, 1970; Katenin, 1972; Kolesnikov, 1985; Morozova, Korotkov, 1999).
Классификация растительности выполнена доминантно-детерминантным методом, предполагающим выделение синтаксонов по доминантам с последующим уточнением их объема по детерминантным группам экологически близких видов (Vasilevich, 1995; Vasilevich, Bibikova, 2004, et al.; Kucherov, Kutenkov, 2011, 2012; Kucherov, 2018). Однако, в отличие от предыдущих работ, мы не подтверждаем установленные группы по критерию Кокрена. Его применение часто приводит к разделению экологически близких видов по разным детерминантным группам, особенно если группы маловидовые, что типично при дифференциации флористически бедных сообществ. Разделение обусловлено наличием у видов индивидуальных “хвостов” распределений по градиентам ведущих факторов. В итоге группирование видов в условиях, оптимальных для их произрастания, оказывается не вполне корректным с геоботанической точки зрения.
К одной ассоциации отнесены описания с общим набором доминантов и детерминантов на всем протяжении ее ареала. У субассоциаций могут быть собственные доминанты и детерминанты в пределах части либо всего ареала ассоциации. Варианты могут выделяться только по детерминантам (Kucherov, 2018), модификации – по доминантам.
Выделенные в результате классификации 11 синтаксонов разного ранга сведены в фитоценотическую таблицу (табл. 1). В литературе очень мало подробных геоботанических данных об изучаемых сообществах, поэтому мы приводим проективное покрытие и постоянство видов в процентной шкале без генерализации. Названия синтаксонов даны согласно традиции школы В.Н. Сукачева (Sukachev, 1931). Названия в форме, принятой в работах по доминантно-детерминантной классификации (Vasilevich, Bibikova, 2004; Kucherov et al., 2010; Kucherov, 2018), приводятся как синонимы. В обоих случаях применен заимствованный из европейских работ принцип nomina conservanda: использованы традиционные краткие эпитеты “myrtillosum”, “uliginosum”, “cornosum” вместо более корректных, но громоздких “vacciniosum myrtilli”, “vaccinietosum uliginosi” и т.д. Приведенные в тексте данные об обилии видов в составе древостоя, подроста и подлеска также соответствуют фитоценотической таблице (табл. 1); в ней же содержатся сведения о покрытии и высоте выделенных ярусов.
Таблица 1.
Фитоценотическая характеристика олиготрофных сфагновых ельников севера Европейской России и сопредельных территорий. Table 1. Phytocoenotic record of oligotrophic peatmoss-spruce boreal forests of European Russia and adjacent areas
| Вид/Species | Ярус Layer |
Синтаксоны/Syntaxa | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | |||||||||||||
| Детерминантные виды / Determinants | |||||||||||||||||||||||
| Sphagnum capillifolium | d | 8 | 69 | + | 13 | 2 | 8 | 1 | 21 | 2 | 14 | 1 | 9 | 2 | 29 | 2 | 16 | 1 | 15 | ||||
| Cladonia arbuscula s. l. | d | 3 | 77 | 1 | 27 | + | 8 | + | 36 | + | 14 | + | 17 | + | 3 | + | 10 | ||||||
| C. rangiferina s. l. | d | 4 | 85 | 1 | 43 | + | 8 | + | 13 | + | 11 | + | 6 | + | 14 | + | 5 | ||||||
| Sphagnum fuscum | d | 13 | 69 | 5 | 40 | + | 17 | + | 20 | + | 10 | + | 9 | 1 | 6 | + | 5 | + | 5 | ||||
| Cladonia stellaris | d | 5 | 77 | 1 | 30 | + | 13 | + | 3 | ||||||||||||||
| C. gracilis subsp. elongata | d | + | 15 | + | 10 | + | 3 | ||||||||||||||||
| Vaccinium uliginosum | c | 18 | 100 | 14 | 93 | 12 | 100 | + | 40 | 10 | 97 | 2 | 74 | 2 | 75 | + | 27 | 1 | 23 | + | 38 | + | 20 |
| Empetrum nigrum s. l. | c | 17 | 100 | 9 | 93 | 1 | 67 | + | 100 | 6 | 100 | 3 | 83 | 3 | 50 | + | 27 | + | 29 | + | 30 | + | 15 |
| Ledum palustre | c | 6 | 54 | 6 | 77 | 4 | 92 | 7 | 54 | 16 | 100 | 17 | 92 | + | 32 | 1 | 17 | + | 30 | 1 | 35 | ||
| Betula czerepanovii | a2+b | + | 13 | + | 3 | ||||||||||||||||||
| Andromeda polifolia | c | + | 15 | 2 | 57 | + | 8 | + | 40 | + | 10 | + | 14 | + | 17 | + | 9 | + | 3 | + | 11 | + | 15 |
| Betula nana | b | 1 | 23 | 33 | 100 | 1 | 50 | + | 10 | + | 11 | + | 5 | + | 11 | + | 11 | + | 15 | ||||
| Oxycoccus palustris | c | + | 23 | 1 | 40 | 1 | 67 | + | 40 | + | 23 | 1 | 23 | + | 8 | + | 5 | + | 23 | + | 27 | 1 | 25 |
| Chamaedaphne calyculata | c | 1 | 17 | 2 | 75 | 1 | 18 | 1 | 49 | 1 | 25 | + | 9 | + | 14 | + | 32 | 1 | 55 | ||||
| Sphagnum magellanicum s. l. | d | 1 | 10 | 7 | 50 | 1 | 40 | + | 5 | + | 17 | 1 | 17 | 2 | 23 | 2 | 51 | 3 | 46 | 5 | 40 | ||
| Equisetum sylvaticum | c | + | 23 | 1 | 30 | + | 8 | + | 40 | 2 | 87 | 2 | 71 | 17 | 100 | 20 | 100 | 1 | 63 | + | 43 | 3 | 55 |
| Melampyrum pratense s. l. | c | + | 31 | + | 27 | + | 33 | + | 80 | 1 | 72 | + | 60 | + | 58 | + | 45 | + | 31 | + | 35 | + | 35 |
| Linnaea borealis | c | + | 15 | + | 3 | + | 8 | + | 100 | + | 36 | + | 26 | 1 | 33 | 1 | 41 | + | 31 | + | 41 | + | 15 |
| Lycopodium annotinum s. l. | c | 1 | 8 | + | 100 | + | 18 | + | 6 | + | 8 | + | 27 | + | 11 | + | 11 | + | 15 | ||||
| Maianthemum bifolium | c | + | 60 | + | 3 | + | 8 | + | 14 | + | 6 | + | 30 | ||||||||||
| Chamaepericlymenum suecicum | c | 4 | 80 | + | 5 | + | 9 | + | 8 | + | 5 | + | 16 | + | 5 | ||||||||
| Solidago virgaurea s. l. | c | + | 15 | + | 3 | + | 80 | + | 8 | + | 3 | + | 3 | + | 3 | + | 10 | ||||||
| Dactylorhiza maculata s. l. | c | + | 8 | + | 80 | + | 5 | + | 3 | + | 5 | + | 3 | + | 3 | + | 20 | ||||||
| Calluna vulgaris | c | 1 | 15 | + | 13 | + | 60 | + | 3 | + | 6 | ||||||||||||
| Molinia caerulea | c | + | 60 | + | 3 | ||||||||||||||||||
| Sphagnum girgensohnii | d | 3 | 13 | + | 8 | 7 | 60 | 1 | 8 | 1 | 14 | + | 9 | 2 | 23 | 3 | 19 | 8 | 35 | ||||
| Juniperus communis s. l. | b | 1 | 23 | + | 17 | 2 | 80 | 1 | 31 | + | 9 | + | 5 | + | 3 | + | 19 | + | 5 | ||||
| Avenella flexuosa s. l. | c | 1 | 46 | + | 17 | + | 8 | 1 | 80 | 1 | 74 | + | 43 | + | 25 | + | 14 | + | 9 | + | 30 | + | 10 |
| Listera cordata | c | + | 8 | + | 3 | + | 25 | + | 40 | + | 36 | + | 17 | + | 8 | + | 27 | + | 29 | + | 16 | + | 10 |
| Orthilia secunda | c | + | 15 | + | 10 | + | 17 | + | 40 | + | 26 | + | 43 | + | 25 | + | 36 | + | 14 | + | 27 | ||
| Dicranum scoparium | d | + | 15 | + | 17 | + | 8 | + | 40 | + | 26 | 1 | 40 | + | 8 | + | 32 | + | 23 | 1 | 46 | + | 20 |
| Sorbus aucuparia s. l. | a2+b | + | 8 | + | 17 | 3 | 100 | + | 23 | + | 20 | 1 | 17 | 1 | 45 | + | 23 | + | 35 | + | 20 | ||
| Trientalis europaea | c | 2 | 60 | + | 10 | + | 11 | + | 17 | 1 | 36 | + | 17 | + | 24 | + | 20 | ||||||
| Luzula pilosa | c | + | 3 | + | 36 | + | 23 | + | 8 | + | 32 | + | 6 | + | 16 | + | 15 | ||||||
| Dicranum polysetum | d | 1 | 23 | + | 3 | 1 | 46 | 1 | 43 | + | 33 | + | 23 | + | 31 | 1 | 41 | + | 10 | ||||
| Pinus sibirica | a1+2 | + | 3 | + | 5 | 1 | 3 | 1 | 5 | 1 | 6 | 2 | 30 | ||||||||||
| Eriophorum vaginatum | c | 2 | 40 | + | 33 | + | 40 | + | 28 | + | 34 | + | 8 | + | 27 | + | 31 | + | 49 | 9 | 100 | ||
| Константные виды / Constants | |||||||||||||||||||||||
| Picea abies s. l. | a1 | 26 | 100 | 26 | 97 | 23 | 100 | 33 | 100 | 33 | 97 | 31 | 100 | 32 | 100 | 33 | 100 | 35 | 97 | 37 | 100 | 24 | 100 |
| Betula pubescens s. l. | a1+2 | 10 | 92 | 4 | 50 | 7 | 83 | 4 | 40 | 9 | 97 | 8 | 94 | 7 | 83 | 16 | 95 | 12 | 97 | 7 | 84 | 14 | 95 |
| Pinus sylvestris | a1+2 | 1 | 31 | 3 | 43 | 3 | 67 | + | 20 | 3 | 46 | 6 | 66 | 6 | 75 | 3 | 41 | 3 | 34 | 8 | 73 | 2 | 50 |
| Picea abies s. l. | b | 7 | 100 | 5 | 80 | 6 | 100 | 8 | 100 | 9 | 95 | 6 | 91 | 7 | 92 | 10 | 100 | 10 | 97 | 11 | 100 | 6 | 90 |
| Betula pubescens s. l. | b | 2 | 92 | + | 33 | 3 | 100 | 2 | 100 | 2 | 79 | 2 | 91 | 2 | 58 | 1 | 64 | 1 | 63 | 1 | 62 | 1 | 85 |
| Vaccinium myrtillus | c | 8 | 92 | 6 | 73 | 4 | 67 | 18 | 100 | 14 | 100 | 25 | 100 | 14 | 100 | 11 | 100 | 18 | 97 | 26 | 97 | 6 | 90 |
| V. vitis-idaea | c | 4 | 62 | 3 | 73 | 4 | 92 | 3 | 100 | 9 | 100 | 8 | 100 | 10 | 92 | 5 | 95 | 4 | 83 | 10 | 100 | 3 | 75 |
| Carex globularis | c | 5 | 69 | 8 | 70 | 4 | 75 | 5 | 80 | 10 | 95 | 7 | 89 | 5 | 75 | 14 | 91 | 28 | 100 | 2 | 81 | 17 | 90 |
| Rubus chamaemorus | c | 1 | 38 | 16 | 93 | 16 | 100 | 25 | 100 | 8 | 79 | 12 | 86 | 14 | 83 | 12 | 95 | 10 | 97 | 10 | 70 | 17 | 85 |
| Pleurozium schreberi | d | 17 | 92 | 13 | 83 | 3 | 75 | 6 | 100 | 19 | 95 | 13 | 94 | 12 | 75 | 7 | 82 | 8 | 94 | 8 | 100 | 2 | 80 |
| Polytrichum commune | d | 8 | 77 | 5 | 77 | 12 | 50 | 14 | 60 | 18 | 97 | 12 | 83 | 18 | 92 | 11 | 95 | 13 | 94 | 10 | 65 | 8 | 70 |
| Sphagnum angustifolium | d | 11 | 46 | 31 | 77 | 45 | 100 | 57 | 100 | 17 | 51 | 21 | 60 | 7 | 33 | 34 | 73 | 47 | 77 | 43 | 95 | 30 | 65 |
| S. russowii | d | 4 | 46 | 13 | 63 | 18 | 75 | 12 | 100 | 19 | 85 | 22 | 94 | 19 | 58 | 13 | 73 | 9 | 54 | 14 | 70 | 11 | 45 |
| Hylocomium splendens | d | 5 | 54 | 5 | 37 | 3 | 67 | 1 | 80 | 13 | 87 | 8 | 91 | 11 | 83 | 6 | 86 | 3 | 66 | 7 | 76 | + | 25 |
| Aulacomnium palustre | d | + | 23 | + | 37 | + | 42 | 1 | 60 | + | 41 | + | 37 | 1 | 42 | + | 27 | + | 31 | + | 46 | + | 35 |
| Прочие виды / Companions | |||||||||||||||||||||||
| Larix sibirica | a1+2 | + | 23 | 1 | 13 | 1 | 23 | + | 11 | + | 9 | + | 9 | ||||||||||
| Populus tremula | a1+2 | + | 3 | + | 17 | + | 8 | + | 3 | ||||||||||||||
| Abies sibirica | a1+2 | 1 | 14 | + | 9 | ||||||||||||||||||
| Picea abies s. l. | a2 | 6 | 54 | 7 | 40 | + | 8 | 2 | 20 | 10 | 64 | 6 | 71 | 10 | 50 | 8 | 68 | 10 | 60 | 4 | 38 | 2 | 45 |
| Salix caprea | a2+b | + | 8 | + | 8 | + | 13 | + | 20 | + | 8 | + | 5 | + | 8 | + | 5 | ||||||
| Pinus sylvestris | b | + | 15 | + | 13 | + | 50 | + | 28 | + | 46 | 1 | 42 | + | 5 | + | 11 | + | 14 | + | 10 | ||
| Rosa acicularis | b | + | 8 | + | 3 | + | 8 | + | 18 | + | 6 | + | 9 | + | 3 | + | 11 | ||||||
| Salix phylicifolia | b | + | 8 | + | 23 | + | 20 | + | 10 | + | 6 | + | 3 | + | 5 | ||||||||
| Abies sibirica | b | + | 3 | + | 3 | 1 | 14 | 1 | 11 | + | 3 | + | 15 | ||||||||||
| Salix aurita | b | + | 40 | + | 17 | + | 6 | + | 8 | 3 | 10 | ||||||||||||
| Melampyrum sylvaticum s. l. | c | + | 3 | + | 20 | + | 5 | + | 3 | + | 9 | + | 6 | + | 8 | 1 | 30 | ||||||
| Carex paupercula | c | + | 20 | + | 3 | + | 8 | + | 9 | + | 9 | + | 11 | 1 | 15 | ||||||||
| Dryopteris carthusiana | c | + | 6 | + | 18 | + | 6 | + | 16 | + | 5 | ||||||||||||
| Carex pauciflora | c | 1 | 3 | 3 | 25 | + | 3 | + | 3 | + | 8 | 1 | 25 | ||||||||||
| Oxycoccus microcarpus | c | + | 27 | + | 8 | + | 10 | + | 3 | ||||||||||||||
| Carex canescens | c | 2 | 8 | + | 3 | + | 3 | + | 6 | + | 14 | + | 10 | ||||||||||
| Chamaenerion angustifolium | c | + | 20 | 1 | 10 | + | 9 | + | 5 | + | 6 | + | 5 | ||||||||||
| Equisetum palustre | c | 1 | 8 | + | 3 | 2 | 17 | + | 5 | + | 6 | + | 5 | ||||||||||
| Carex vaginata | c | + | 23 | + | 7 | + | 3 | + | 10 | ||||||||||||||
| C. nigra | c | + | 3 | + | 3 | + | 16 | ||||||||||||||||
| Polytrichum strictum | d | 1 | 23 | 1 | 33 | 1 | 42 | + | 41 | + | 20 | + | 9 | + | 11 | + | 11 | 1 | 25 | ||||
| Ptilium crista-castrensis | d | + | 10 | + | 17 | + | 20 | + | 23 | 1 | 29 | + | 25 | + | 27 | + | 14 | 1 | 14 | + | 10 | ||
| Sphagnum flexuosum | d | 4 | 7 | 12 | 28 | 8 | 29 | 6 | 17 | 11 | 23 | 8 | 23 | 10 | 30 | ||||||||
| Dicranum majus | d | + | 8 | + | 10 | + | 33 | + | 20 | + | 18 | 1 | 11 | + | 17 | + | 14 | + | 14 | + | 22 | ||
| Pohlia nutans s. l. | d | + | 15 | + | 7 | + | 50 | + | 13 | + | 9 | + | 8 | + | 9 | + | 11 | + | 22 | + | 10 | ||
| Sphagnum fallax | d | 3 | 10 | + | 8 | 3 | 40 | 1 | 13 | + | 3 | 1 | 9 | 5 | 29 | + | 5 | + | 5 | ||||
| Peltigera aphthosa | d | + | 31 | + | 7 | + | 8 | + | 18 | + | 9 | + | 8 | + | 3 | + | 15 | ||||||
| Nephroma arcticum | d | 2 | 31 | + | 27 | + | 5 | + | 6 | + | 3 | + | 3 | + | 15 | ||||||||
| Sphagnum centrale | d | + | 3 | + | 20 | + | 3 | 1 | 6 | + | 14 | + | 11 | + | 11 | 4 | 20 | ||||||
| Barbilophozia lycopodioides | d | + | 15 | + | 10 | + | 5 | + | 9 | + | 5 | + | 9 | + | 3 | + | 5 | ||||||
| Polytrichum juniperinum | d | + | 10 | + | 8 | + | 3 | + | 11 | + | 5 | + | 6 | + | 5 | ||||||||
| Sphagnum wulfianum | d | + | 3 | 2 | 3 | + | 9 | 1 | 14 | + | 10 | ||||||||||||
| Cladonia gracilis subsp. gracilis | d | 1 | 31 | + | 10 | + | 3 | + | 10 | ||||||||||||||
| Lophozia spp. | d | + | 3 | + | 5 | + | 6 | + | 5 | + | 11 | ||||||||||||
| Warnstorfia exannulata | d | + | 8 | + | 5 | + | 8 | + | 6 | + | 11 | ||||||||||||
| Straminergon stramineum | d | + | 7 | + | 3 | + | 5 | + | 8 | + | 15 | ||||||||||||
| Sphagnum warnstorfii | d | 2 | 17 | 3 | 8 | 1 | 3 | 3 | 5 | ||||||||||||||
| Dicranum fuscescens | d | + | 3 | + | 5 | + | 3 | + | 3 | + | 20 | ||||||||||||
| Sphagnum riparium | d | + | 3 | 2 | 20 | + | 3 | + | 3 | + | 3 | 3 | 10 | ||||||||||
| Warnstorfia fluitans | d | + | 3 | + | 20 | + | 9 | + | 8 | ||||||||||||||
| Cladonia deformis | d | + | 23 | + | 10 | ||||||||||||||||||
| Sphagnum lindbergii | d | + | 20 | + | 3 | + | 3 | ||||||||||||||||
| Средние сомкнутость крон/проективное покрытие (%) и высота (м) ярусов: Average canopy density/projective cover (%) and height (m) of layers: |
|||||||||||||||||||||||
| – 1-го яруса древостоя/1st tree layer | a1 | 0.4 | 15 | 0.3 | 10 | 0.4 | 12 | 0.4 | 15 | 0.4 | 15 | 0.5 | 16 | 0.5 | 17 | 0.5 | 17 | 0.5 | 17 | 0.5 | 19 | 0.4 | 16 |
| – 2-го яруса древостоя/2nd tree layer | a2 | 0.1 | 9 | 0.1 | 2.5 | – | – | <0.1 | 7 | 0.2 | 9.5 | 0.1 | 8 | 0.1 | 12 | 0.1 | 11 | 0.1 | 12 | <0.1 | 12 | 0.1 | 11 |
| – подроста и подлеска/ undergrowth and shrubs | b | 10 | 2.0 | 40 | 1.3 | 5 | 2.3 | 15 | 2.7 | 10 | 2.0 | 10 | 2.3 | 10 | 2.9 | 15 | 2.1 | 15 | 3.0 | 15 | 3.4 | 10 | 1.8 |
| – травяно-кустарничкового/dwarfshrubs and herbs | c | 60 | 65 | 50 | 65 | 65 | 75 | 75 | 65 | 60 | 50 | 65 | |||||||||||
| – мохового/mosses | d | 100 | 100 | 100 | 95 | 95 | 90 | 75 | 90 | 95 | 95 | 95 | |||||||||||
| Средний бонитет древостоя / Average stand quality class | V | Vb | Va | V | V | V | IV | IV | IV | IV | V | ||||||||||||
| Средняя мощность торфа, см / Average peat thickness, cm | 25 | 55 | 50 | 75 | 30 | 35 | 25 | 35 | 40 | 90 | 50 | ||||||||||||
| Число описаний/Number of relevés (Σ = 260) | 13 | 30 | 12 | 5 | 39 | 35 | 12 | 22 | 35 | 37 | 20 | ||||||||||||
Примечания / Notes. Синтаксоны / Syntaxa: 1 – Piceetum (P.) sphagno fusci-empetrosum; 2 – P. sphagno angustifolii-nanobetulosum; 3 – P. sphagno angustifolii-uliginosum; 4 – P. sphagno angustifolii-cornosum, 5–7 – P. sphagno angustifolii-ledosum: 5 – subass. uliginosum, 6 – subass. typicum, 7 – subass. equisetosum; 8–10 – P. sphagno angustifolii-myrtillosum: 8 – var. Equisetum sylvaticum, 9 – var. typica f. Carex globularis, 10 – var. typica f. typica; 11 – P. sphagno angustifolii-eriophorosum.
Ярусы / Layers: a1 – 1-й ярус древостоя / dominant tree layer, a2 – 2-й ярус древостоя / subordinate tree layer, b – подрост и подлесок / undergrowth and shrubs, c – травяно-кустарничковый / dwarfshrubs and herbs, d – моховой / epigeic mosses.
Для видов приводятся: в левой части колонок – среднее проективное покрытие, в правой – постоянство, при характеристике древостоя – средние сомкнутость крон и высота. Детерминантные группы выделены полужирной рамкой и серым фоном; виды в их пределах сгруппированы по ярусам, далее в порядке убывания встречаемости во всем массиве описаний таблицы. Для доминирующих видов значения покрытия и постоянства даны полужирным шрифтом. Символом “+” отмечено среднее проективное покрытие видов менее 0.5%. Исключены сопутствующие виды со встречаемостью менее 10% хотя бы в одном из синтаксонов, всходы деревьев и кустарников, эпифитные лишайники и мхи (Kucherov et al., 2010; Kucherov, Kutenkov, 2011, 2012).
Average cover % and constancy % are given for species in the left and the right sides of columns, respectively. Average canopy density and tree height are given for canopy layers in the same way. The determinant groups are marked by bold margins and the grey backgrounds; species within them are sorted by layer, then by descending occurrence within the whole relevé set of the table. The cover and constancy values are shown in bold for dominant species. Average species cover less than 0.5% is given as “+”. Species with occurrence less than 10% at least in one of the syntaxa are excluded from the table together with tree and shrub seedlings and epiphytic mosses and lichens (Kucherov et al., 2010; Kucherov, Kutenkov, 2011, 2012).
Номенклатура сосудистых растений дана по С.К. Черепанову (Cherepanov, 1995), мхов – по сводке М.С. Игнатова с соавторами (Ignatov et al., 2006), лишайников – по “Списку лихенофлоры России” (A checklist…, 2010). При отнесении всех европейских таежных популяций ели к единому комплексу Picea abies s. l. мы следуем П.П. Попову (Popov, 2005). Границы подзон таежной зоны приняты по Т.И. Исаченко и Е.М. Лавренко (Isachenko, Lavrenko, 1980; см. рисунки 1–3) с рядом последующих уточнений (Aleksandrova, Yurkovskaya, 1989; Safronova, Yurkovskaya, 2015). Вслед за геоботаниками петрозаводской и сыктывкарской школ (Yudin, 1948, 1954; Ramenskaya, 1983; Martynenko, 1999; Bobkova, Patov, 2006) мы признаем выделение крайнесеверной тайги в ранге особой полосы в северной части северотаежной подзоны. Этой полосе свойствен специфичный набор лесных синтаксонов, что было подтверждено на материале сосняков и лиственничников севера Европейской России (Kucherov, 2018). Отсутствие данных о границах полосы в Архангельской обл. мешает закартировать ее в масштабе Европейской России в целом.
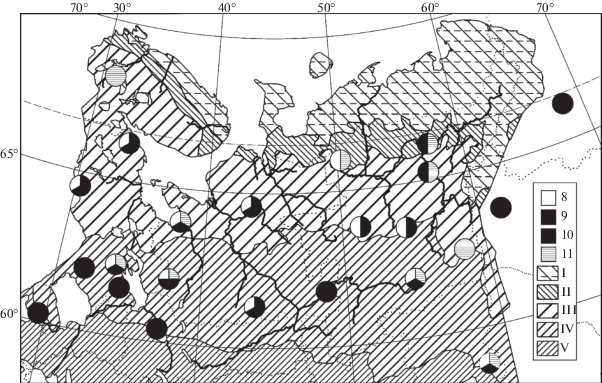
Рис. 1.
Распространение ельников вороничных зеленомошно-сфагновых (1), ерниковых (2), голубичных (3) и деренно-морошковых (4) сфагновых в Европейской России и на Урале (по данным имеющейся выборки описаний).
Зоны и подзоны: I – тундра, II – лесотундра и подгольцовые редколесья, III – северная, IV – средняя, V – южная тайга. Картооснова по Т.И. Исаченко и Е.М. Лавренко (Isachenko, Lavrenko, 1980), с объединением зональных выделов и их высотно-поясных аналогов. Полоса крайнесеверной тайги примерно соответствует северной части подзоны III, на Северо-Востоке Европейской России – также южной части зоны II; она не отражена на карте ввиду недостатка данных о ее границах. Местонахождения синтаксонов 2 и 3 на юге Ямала находятся за пределами картоосновы.
Fig. 1. Distribution of Piceetum (P.) sphagno fusci-empetrosum (1), P. sphagno angustifolii-nanobetulosum (2), P. sphagno angustifolii-uliginosum (3), and P. sphagno angustifolii-cornoso-chamaemorusum (4) in European Russia and the Urals (mapped according to the available relevé set).
Zones and subzones: I – tundra, II – forest-tundra and subalpine open woodlands, III – northern-boreal, IV – middle-boreal, V – southern-boreal subzones. The base map after T. I. Isachenko and E. M. Lavrenko (1980) is generalized by the association of zonal / subzonal units together with their altitudinal analogs. The northernmost-boreal belt approximately corresponds to the northern part of subzone III, also to the southern part of zone II in the North-East of European Russia; its mapping is yet impossible because of the insufficient knowledge of its ranges. The South Yamal findings of syntaxa 2 and 3 are located beyond the base map limits.
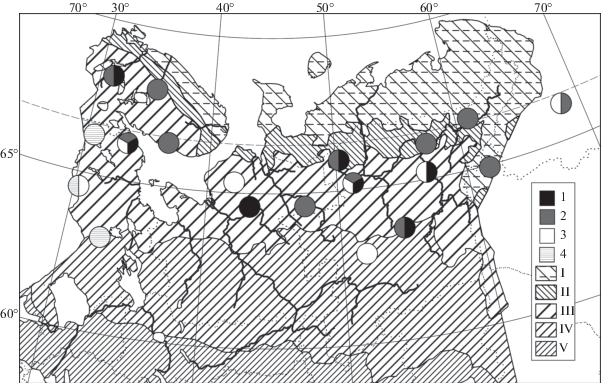
Рис. 2.
Распространение ельников багульниковых сфагновых (5–7) в Европейской России и на Урале (по данным имеющейся выборки описаний).
Fig. 2. Distribution of Piceetum sphagno angustifolii-ledosum (5–7) in European Russia and the Urals (mapped according to the available relevé set).
Субассоциации / Subassociations: 5 – uliginosum, 6 – typicum, 7 – equisetosum sylvaticae. Отображение зональности как на рис. 1 / Zonation units are the same as in Fig. 1.
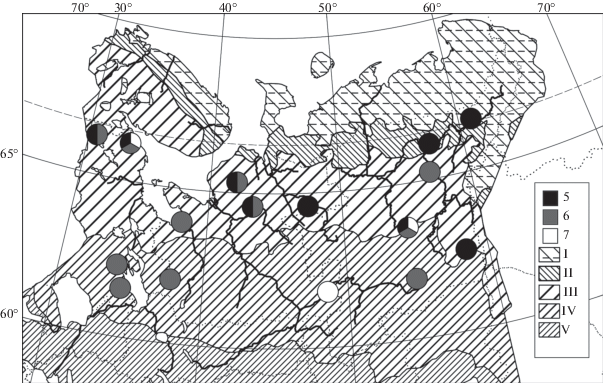
Рис. 3.
Распространение ельников черничных (8–10) и пушицевых (11) сфагновых в Европейской России и на Урале (по данным имеющейся выборки описаний).
Fig. 3. Distribution of Piceetum (P.) sphagno angustifolii-myrtillosum (8–10) and P. sphagno angustifolii-eriophorosum (11) in European Russia and the Urals (mapped according to the available relevé set). Подчиненные синтаксоны / Subordinate syntaxa: 8 – var. Equisetum sylvaticum, 9 – var. typica f. Carex globularis, 10 – var. typica f. typica. Местонахождение синтаксона 10 на юге Ямала находится за пределами картоосновы. Отображение зональности как на рис. 1. The South Yamal finding of syntaxon 10 is located beyond the base map limits. Zonation units are the same as in Fig. 1.
Для оценки климатических условий распространения синтаксонов в Европейской России и на Урале использованы суммы градусо-дней выше 10°C (превышений среднесуточных температур над базовой температурой в 10°C; GDD>10) по данным глобальной сети среднемноголетней (1983–2004) спутниковой метеосъемки NASA SSE (NASA…, 2006). На основе данных из упомянутой сети также рассчитаны индексы континентальности В. Конрада (K; Tukhanen, 1980) (табл. 2).
Таблица 2.
Амплитуды среднемноголетних (1983–2004) значений избранных климатических параметров и индексов (NASA…, 2006) в пределах ареалов синтаксонов олиготрофных сфагновых ельников. Table 2. Amplitudes of averaged (1983–2004) values of selected climatic parameters and indices (NASA…, 2006) within the ranges of oligotrophic peatmoss-spruce forest syntaxa
| Синтаксоны/Syntaxa | Метеопараметры/Climatic parameters | |
|---|---|---|
| GDD > 10, °C | K | |
| 1. Piceetum (P.) sphagno fusci-empetrosum | 208–451 | 32.2–44.1 |
| 2. P. sphagno angustifolii-nanobetulosum | 140–410 | 26.6–48.2 |
| 3. P. sphagno angustifolii-uliginosum | 140–476 | 34.4–48.2 |
| 4. P. sphagno angustifolii-cornoso-chamaemorosum | 285–424 | 35.1–36.2 |
| 5. P. sphagno angustifolii-ledosum subass. uliginosum | 245–451 | 34.4–45.9 |
| 6. Subass. typicum | 281–544 | 34.4–44.4 |
| 7. Subass. equisetosum | 281–573 | 34.4–44.1 |
| 8. P. sphagno angustifolii-myrtillosum var. Equisetum sylvaticum | 281–636 | 34.4–44.6 |
| 9. Var. typica f. Carex globularis | 281–636 | 35.4–46.6 |
| 10. Var. typica f. typica | 140–619 | 31.2–48.2 |
| 11. P. sphagno angustifolii-eriophorosum | 208–636 | 32.2–45.6 |
ОБЩАЯ ХАРАКТЕРИСТИКА ОЛИГОТРОФНЫХ ЕЛЬНИКОВ СФАГНОВЫХ И СФАГНОВО-ЗЕЛЕНОМОШНЫХ
Зонально-климатические ареалы сообществ олиготрофных ельников сфагновых (включая сфагново-зеленомошные) могут быть двух основных типов – крайнесеверо- и северотаежными либо северо- и среднетаежными. Отмечены и синтаксоны, приуроченные лишь к южной полосе подзоны северной тайги, а также азональные типы, которые, однако, исчезают в южной тайге вместе с другими синтаксонами из состава группы, уступая место мезотрофным ельникам. Лишь в океаническом климате Западной Фенноскандии олиготрофные сфагновые ельники подлинно азональны и проникают далеко на юг (Dierßen, 1996).
Во всех широтных полосах подзоны северной тайги рассматриваемые сообщества характерны для заболоченных водоразделов и окраин олигомезотрофных болот. Как в северной, так и в средней тайге они приурочены также к бессточным либо слабопроточным депрессиям, ложбинам стока, межсельговым и межозовым понижениям (Rysin, Savelieva, 2002), а в низкогорьях – к лощинам и шлейфам горных склонов.
Собственно ельники сфагновые, как правило, развиты на торфяно-глеевых либо торфяно-болотных верховых почвах (Sklyarov, Sharova, 1970; Zaboyeva, 1975; Medvedeva et al., 1980) по переувлажненной торфяной залежи средней либо значительной мощности, изолирующей корневые системы ели и большинства других видов растений от минеральных горизонтов почвы. Кислотность (pHH20) торфа в норвежских ельниках составляет 3.6–4.0 (Kielland-Lund, 1981), в Республике Коми – от 3.9 до 4.3–4.4 (Zaboyeva, 1975; Bobkova, 2006a), pHKCl в Карелии – от 3.2 до 4.2 (Medvedeva et al., 1980).
Ельники сфагново-зеленомошные растут на торфянисто-подзолисто-глеевых или торфянисто-глеевых почвах (Sklyarov, Sharova, 1970; Zaboyeva, 1975) по мелкой залежи, почти всегда на силикатах; pHH20 торфа с поверхности 4.1–4.5, в толще залежи – до 4.8 (Zaboyeva, 1975; Kielland-Lund, 1981; Bobkova, 2006a). Для них типичен двухкомпонентный нанорельеф из кочек и сфагновых ковров. В ельниках сфагновых по окраинам болот нанорельеф бывает и трехкомпонентным с участием обводненных западин. Для водораздельных и низкогорных ельников сфагновых более характерны ровные либо волнистые моховые ковры, копирующие неровности подстилающей поверхности.
Древостои низкобонитетные (V–Vb, реже IV), в большинстве случаев сомкнутость 1-го яруса не превышает 0.4, в крайнесеверной тайге – 0.3. В древостоях с бонитетом ниже V и при подтоплении много угнетенных деревьев и сухостоя. Во многих типах сообществ есть примесь березы (Betula pubescens s. l., в крайнесеверной тайге – B. czerepanovii) к ели. Подрост ели в крайнесеверной тайге и при подтоплении малочислен и угнетен, в иных случаях жизнеспособен и умеренно обилен. Для яруса кустарничков и трав (проективное покрытие, далее – ПП, 50–75%) характерно участие Vaccinium myrtillus, V. vitis-idaea, тундрово-болотных кустарничков-оксилофитов (V. uliginosum, Ledum palustre, Empetrum nigrum s. l.), Carex globularis, Rubus chamaemorus. В ряде синтаксонов в составе яруса присутствует также Equisetum sylvaticum. Соотношение обилий этих видов меняется от синтаксона к синтаксону. В моховом ярусе (ПП обычно 95–100%) доминирует Sphagnum angustifolium в сопровождении S. russowii, реже S. fuscum.
Ельники сфагновые с господством Sphagnum angustifolium и обилием болотных кустарничков произрастают в самых бедных условиях минерального питания из всех возможных для заболоченных еловых лесов, находясь на границе атмосферно-грунтового и атмосферного рядов питания. По мере накопления торфа и прекращения влияния грунтовых вод они переходят в сосняки кустарничково-сфагновые и безлесные верховые болота (Kutenkov, Kuznetsov, 2013; Kutenkov, 2015). В ельниках сфагново-зеленомошных на начальных этапах заболачивания S. angustifolium разделяет господство с Polytrichum commune, Pleurozium schreberi и (в меньшей степени) Hylocomium splendens. Зеленые мхи обычно растут на кочках и при стволах, а S. angustifolium формирует ковры.
ХАРАКТЕРИСТИКА ВЫДЕЛЕННЫХ СИНТАКСОНОВ
Piceetum (P.) sphagno fusci-empetrosum (Sphagno fusci-Empetro-P.) – ельник (е.) вороничный зеленомошно-сфагновый (табл. 1, № 1). Торфяная залежь средне- либо маломощная (5–50 см), нанорельеф не выражен. Сомкнутость 1-го яруса елового древостоя 0.3, средняя высота 15 м; Betula pubescens s. l. (0.1) формирует 2-й ярус высотой 6–10 м. Бонитет V. Подрост еловый. В ярусе кустарничков согосподствуют Empetrum hermaphroditum и Vaccinium uliginosum (ПП по 20%), им сопутствуют V. myrtillus (10%), V. vitis-idaea, Ledum palustre, Carex globularis (по 5%). Изредка в малом обилии встречается Rubus chamaemorus. В пестром напочвенном ковре сочетаются Sphagnum fuscum, S. angustifolium, S. capillifolium, Pleurozium schreberi (по 10–20%) и кустистые лишайники (Cladonia arbuscula s. l., C. rangiferina s. l., C. stellaris; по 5%). Sphagnum capillifolium и Cladonia arbuscula s. l. выступают детерминантами ассоциации; Sphagnum fuscum, Cladonia rangiferina s. l., C. stellaris и C. gracilis subsp. elongata – дифференциальные виды, объединяющие ассоциацию с ельниками ерниковыми в противовес прочим синтаксонам.
Сообщества данной ассоциации наиболее характерны для полосы крайнесеверной тайги. Они описаны в Мурманской обл. в долинно-лесном поясе предгорий Сальных тундр, а также в Республике Коми, где встречаются по Цилемской Печоре, рекам Пижме и Кожве на водораздельных участках, удаленных на 1–8 км от реки (P. fruticuloso-clado-hypno-sphagnosum, P. uligini-vaccinioso-sphagnosum (Yudin, 1948, 1954; Martynenko, 1999)). Реже подобные ельники заходят в южную полосу северной тайги, где отмечены как на заболоченных водоразделах в Пинежском заповеднике на севере Архангельской обл., так и по краю болот у ст. Лоухи на севере Карелии (P. empetroso-ledosum (Blagoveshchenskii, 1936)) и в окрестностях п. Нижний Одес на водоразделе Ижмы и Печоры (рис. 1).
Вариантом рассматриваемой ассоциации выступает P. callunoso-sphagnosum на торфяно-глеевых почвах водоразделов запада Беломорско-Кулойского плато с напочвенным ярусом сходного состава (но без Sphagnum capillifolium) и с доминированием (возможно, вследствие пожаров) Calluna vulgaris в ярусе кустарничков. Vaccinium uliginosum и Empetrum nigrum s. l. несколько уступают Calluna vulgaris по обилию. Встречаются Arctous alpina, Eriophorum vaginatum, Carex globularis; отмечено возобновление сосны (Leontiev, 1937; Rysin, Savelieva, 2002).
P. sphagno angustifolii-nanobetulosum (Sphagno angustifolii-Betulo nani-P.) – е. ерниковый сфагновый (табл. 1, № 2). Произрастает на заболоченных водоразделах и пологих склонах грив и увалов, в Лапландии и Северной Карелии также по окраинам болот и заболоченным берегам озер, на Терском берегу Белого моря – на аллювиальных речных террасах (Regel, 1923), а на Полярном Урале – в горных долинах (Soczava, 1927). Торф мощностью от 20–30 до 120–150 см залегает поверх песка либо суглинка; на Южном Ямале торф мерзлый. Нанорельеф на водоразделах волнистый, по окраинам болот – слабовыраженно кочковатый. Кочки, высотой 10–20 см, при этом занимают 70–75% площади ценоза, лишь в ельнике из бассейна р. Мезенской Пижмы – 40% (Korchagin, 1956). Древостой разрежен (0.3, реже 0.4), сильно варьирует по высоте 1-го яруса – от 5–8 м в крайнесеверной тайге до 17–22 м на южном пределе ареала ассоциации близ п. Нижний Одес в северной тайге Республики Коми. Средний бонитет Vб. Во 2-м ярусе слабо выражена примесь Betula czerepanovii. Подрост ели малочислен. Развит умеренно густой (ПП 30–40%) ярус подлеска из B. nana до 1 м высотой. Ф.В. Самбук (Sambuk, 1932) считает этот ярус тундровым по происхождению, а само сообщество ельника ерникового – результатом инкумбации ярусов. В травяно-кустарничковом ярусе согосподствуют Vaccinium uliginosum, Empetrum nigrum s. l. и Rubus chamaemorus (по 10–15%). Постоянны, но не столь обильны (по 5%) Vaccinium myrtillus, V. vitis-idaea, Ledum palustre и Carex globularis. В моховом ярусе преобладают Sphagnum angustifolium (35%) и S. russowii (15%), иногда с примесью S. fallax или S. flexuosum, на микроповышениях – Pleurozium schreberi (15%) и Polytrichum commune (10%). Постоянные, но малообильные мхи и лишайники из группы Sphagnum fuscum–Cladonia rangiferina (см. выше) отличают ассоциацию от ельников голубичных сфагновых, а Betula nana и Oxycoccus palustris – от вороничных сфагново-зеленомошных. Собственная детерминантная группа ассоциации маловидовая; ее формируют Andromeda polifolia и Betula czerepanovii.
Ельник ерниковый сфагновый – характерная ассоциация крайнесеверной тайги. В Мурманской обл. она отмечена в Лапландском заповеднике (Piceetum nano-betulosum (Nekrasova, 1935), Ловозерских горах (P. sphagnosum (Lyubimova, 1937)) и на Терском берегу Белого моря (P. sphagnosum (Regel, 1923)), а в Архангельской обл. – в северной части бассейна Мезени (Sambuk, 1932). Ю. Д. Цинзерлинг (Zinserling, 1932) приводит ее для Кольского п-ова и Северной Карелии в целом. В Республике Коми синтаксон многократно описан по Цилемской Печоре и ее притокам, в бассейне р. Усы (P. sphagno-betulosum (Sambuk, 1932), P. nano-betuloso-sphagnosum (Yudin, 1948, 1954)) и близ п. Сивая Маска (Katenin, 1972). Восточнее леса данной ассоциации известны с Малого Урала (Sphagnopiceetum nano-betulosum (Soczava, 1927)) и Южного Ямала. В собственно северную тайгу подобные ельники заходят на юг до района Лоухи-Кестеньгского тракта (P. fruticuloso-sphagnosum (Blagoveshchenskii, 1936), Sphagnetum fruticuloso-piceiosum (Sokolova, 1936)), р. Мезенской Пижмы (Korchagin, 1956), Средней Печоры близ д. Кипиево (P. nano-betuloso-sphagnosum, P. chamaemoroso-sphagnosum (Sambuk, 1932)) и Печоро-Ижемского водораздела (рис. 1).
P. sphagno angustifolii-uliginosum (Sphagno angustifolii-Uliginosi-P.) – е. голубичный сфагновый (табл. 1, № 3). В крайнесеверной тайге встречается на водоразделах либо (на Ямале) по склонам пойменных террас, южнее – по окраинам болот, реже в депрессиях на плоских вершинах завалуненных озов. Торф мощностью от 10 до 140 см поверх песчаных либо щебнистых отложений или скальных пород, на Ямале мерзлый. Нанорельеф не выражен, реже двухкомпонентный с кочками высотой 10–30 (редко до 100) см и господством сфагновых ковров (75–85% площади ценоза). Древостой сомкнутостью 0.3–0.7. Его высота возрастает от 9 до 18 м с уменьшением географической широты; бонитет при этом меняется от Vб до IV (табл. 1). Примесь Betula pubescens s. l. к Picea abies s. l. до 2 единиц по составу древостоя. Подрост березово-еловый, умеренно обильный. В травяно-кустарничковом ярусе согосподствуют Vaccinium uliginosum и Rubus chamaemorus (по 15%). Прочие виды болотных кустарничков, а также Vaccinium myrtillus, V. vitis-idaea и Carex globularis постоянны, но не обильны. Ярус мхов сформирован Sphagnum angustifolium (45%), S. russowii (20%) и Polytrichum commune (10%) с примесью S. magellanicum s. l. (5%). Последний вид вместе с Chamaedaphne calyculata составляет маловидовую группу дифференциальных видов синтаксона. Betula nana и Oxycoccus palustris отличают ельники голубичные сфагновые от деренно-морошковых (см. ниже).
В полосе крайнесеверной тайги ассоциация описана по р. Сояне на Беломорско-Кулойском плато (на известняках (Braslavskaya et al., 2017)), по левому притоку Печоры р. Цильме (P. uligini-vaccinioso-sphagnosum (Yudin, 1948, 1954)) и на Ямале по р. Полуй. В южной полосе северной тайги леса данного синтаксона отмечены в Финляндии (Heidelbeerbruch (Ruuhijärvi, 1960)), в Костомукшском заповеднике (Kutenkov, 2015), близ оз. Кумозеро и на южном берегу губы Чупа в Карелии, а в Республике Коми – по р. Чикишне (Yudin, 1948, 1954). В Коми ельники голубичные сфагновые проникают в среднюю тайгу, где известны на междуречье Симвы и Вишеры (Yudin, 1948, 1954) (рис. 1). О том, что они встречаются и в средней тайге Архангельской обл., пишет В. Г. Чертовской (Chertovskoi, 1978).
Замещающая ассоциация ельников голубичных сфагновых описана в среднегорьях Центральной Европы на высоте 800–1200 м над ур. м. под названием Vaccinio uliginosi-Piceetum Schubert 1972, она же Sphagno-Piceetum Zukrigl 1973. В системе синтаксонов школы Ж. Браун-Бланке ее относят к союзу Vaccinio uliginosi-Pinion sylvestris Passarge 1968 порядка Vaccinio-Piceetalia Br.-Bl. et al. 1939 класса Vaccinio-Piceetea Br.-Bl. et al. 1939. Ельники этой ассоциации растут по глубокой торфяной залежи в условиях охлаждающего влияния грунтовых вод, вследствие которого корневые системы всех видов растений сосредоточены в нескольких сантиметрах под поверхностью залежи. Древостой сомкнутостью 0.3–0.6 с примесью Betula carpatica. В приземных ярусах доминантами и одновременно детерминантами ассоциации являются Vaccinium uliginosum, V. myrtillus, Eriophorum vaginatum, а среди мхов – Sphagnum angustifolium s. l. (incl. S. fallax, S. flexuosum) и S. magellanicum s. l. одновременно с мезотрофным S. girgensohnii. Вероятно, олиготрофные и мезотрофные виды сфагнов дифференцированы по элементам нанорельефа; указано, что S. girgensohnii характерен для всех синтаксонов заболоченных ельников гор Средней Европы. К малообильным дифференциальным видам относятся Vaccinium vitis-idaea, Oxycoccus palustris, Melampyrum pratense, Bazzania trilobata, Sphagnum russowii, Polytrichum commune, а к доминирующим константам – Calamagrostis villosa, Molinia caerulea и Sphagnum capillifolium. Ассоциация известна от Западных Карпат и Рудных гор до Гарца (Chytrý et al., 2013) и Альп (Willner, Grabherr, 2007). В Чехии она изредка встречается и по окраинам болот на равнине (Chytrý et al., 2013).
Иной, но флористически близкий к предыдущему среднеевропейский синтаксон ельников голубичных сфагновых приводится из гор Шварцвальда под именем Bazzanio-Piceetum Br.-Bl. et al. 1939 vaccinietosum uliginosi Oberd. 1992. Он отнесен к подсоюзу Vaccinio-Piceenion Oberd. 1957 союза Vaccinio-Piceion Br.-Bl. et al. 1939 порядка Vaccinio-Piceetalia. Эти леса произрастают в прохладном субокеаническом климате на высоте 600–900 м над ур. м. в понижениях по краю верховых болот на олиготрофных торфяных и торфяно-глеевых почвах. Диагностические виды субассоциации – Vaccinium uliginosum, Eriophorum vaginatum, Sphagnum angustifolium, S. magellanicum s. l., Aulacomnium palustre, ассоциации в целом – Melampyrum pratense subsp. oligocladum и Bazzania trilobata. Постоянны Vaccinium vitis-idaea и (для ассоциации в целом) V. myrtillus, Sphagnum girgensohnii, Hylocomium splendens, Pleurozium schreberi, Rhytidiadelphus loreus. Встречаются Listera cordata, Oxycoccus palustris, Polytrichum commune. Ельники могут быть как высоко-, так и низкоствольными (Oberdorfer, 1992). Леса этой ассоциации по сравнению с предыдущей заболочены не так сильно. Тем не менее, обе ассоциации, возможно, подлежат включению в более низком ранге в состав единого синтаксона.
Голубичные ельники – единственный синтаксон олиготрофных сфагновых еловых лесов, для которого по данным литературы удалось выявить замещающие синтаксоны в лесном поясе гор Средней Европы. Видимо, это обусловлено широтой эколого-ценотической амплитуды Vaccinium uliginosum по градиенту океаничности-континентальности климата (Ellenberg et al., 1992). Для всех остальных синтаксонов олиготрофных сфагновых ельников замещающие их типы сообществ за пределами Европейского, Уральского и Западносибирского Севера не выявлены. Это коренным образом отличает данные сообщества от заболоченных ельников, развитых в условиях мезотрофного питания, которые распространены гораздо более широко.
P. sphagno angustifolii-cornoso-chamaemorusum (Sphagno angustifolii-Corno-P.) – е. деренно-морошковый сфагновый (табл. 1, № 4). Специфичная для Западной Карелии низкогорная северотаежная ассоциация. Ее участки описаны в Муезерском районе на склонах гор Воттоваара и Лысая близ оз. Сегозеро и в природном парке “Тулос”, а также на северо-западе Лоухского района в национальном парке “Паанаярви” (Kutenkov, 2009, 2015) (рис. 1). Как правило, сообщества развиваются в ложбинах водотоков и расщелинах пологих горных склонов при наличии умеренно обогащенного подтока сверху, реже вблизи болот. Мощность торфа 50–130 см поверх каменистой силикатной породы, песка либо суглинка; нанорельеф не выражен. Сомкнутость древостоя 0.3–0.5, высота 10–14, лишь иногда до 22 м. Бонитет IV–Va. Примесь березы к ели почти не выражена. Подрост ели умеренно обилен (ПП 10%); развит редкий подлесок из Juniperus communis s. l. и Sorbus aucuparia s. l. (по 2–3%). Доминанты травяно-кустарничкового яруса – Vaccinium myrtillus, Rubus chamaemorus (по 20–25%) и Chamaepericlymenum suecicum (10%); их спутниками выступают Vaccinium vitis-idaea, Carex globularis и Trientalis europaea (по 2–5%). Постоянны, но крайне малообильны Vaccinium uliginosum и Empetrum nigrum s. l. наряду с Linnaea borealis и Lycopodium annotinum. Не отмечен (возможно, случайно) Ledum palustre. В то же время нередки Calluna vulgaris и Molinia caerulea; в малом обилии появляется Equisetum sylvaticum. В моховом ковре Sphagnum angustifolium (60%) сопровождают не только S. russowii и Polytrichum commune (по 10–15%), но и S. girgensohnii (5%), что отражает различия микроэкотопов по условиям питания. Такое же покрытие у Pleurozium schreberi; прочие виды таежных зеленых мхов встречаются в малом обилии.
Синтаксон характеризует обширная группа детерминантов с преобладанием олигомезотрофных и мезотрофных таежно-лесных мезофитов (Lycopodium annotinum s. l., Maianthemum bifolium, Solidago virgaurea s. l.) и гигромезофитов (Sphagnum girgensohnii), сопровождающих Picea abies s. l. В состав группы входят и мезотрофные гигромезофиты – гипоарктический амфиокеанический Chamaepericlymenum suecicum и бореальный болотно-лесной Dactylorhiza maculata s. l., – а также растущие вместе на пустошах и болотах Центральной Европы субатлантические Calluna vulgaris (олиготрофный психромезофит) и Molinia caerulea (мезоэвтрофный гигромезофит). Дифференциальная группа в составе Equisetum sylvaticum, Melampyrum pratense s. l. (incl. subsp. alpestre) и Linnaea borealis сближает синтаксон с ельниками багульниковыми, морошково-черничными и пушицевыми сфагновыми, отличая от крайнесеверотаежных вороничных, ерниковых и голубичных. Только с ельниками морошково-черничными и пушицевыми деренно-морошковые объединяет маловидовая группа, состоящая из Sorbus aucuparia s. l. и Trientalis europaea, с морошково-черничными и багульниковыми – Orthilia secunda и Dicranum scoparium, а с мелкозалежными багульниковыми сфагново-зеленомошными голубичной субассоциации – Juniperus communis s. l., Avenella flexuosa и Listera cordata.
Леса данного синтаксона должны произрастать также в сопредельных с Карелией низкогорных районах Финляндии, но упоминаний о них в литературе мы не встретили. Флористически близкие ельники с незначительным обилием Chamaepericlymenum suecicum и Molinia caerulea, дважды описанные первым из авторов в предгорьях Сальных тундр и в приморском скальном комплексе в Керетской Карелии, отличались массовостью болотных кустарничков, в особенности Ledum palustre, в силу чего их описания включены в следующую ассоциацию.
P. sphagno angustifolii-ledosum (Sphagno angustifolii-Ledo-P.) – е. багульниковый сфагновый (табл. 1, № 5–7). Северотаежная ассоциация с иррадиацией как в среднюю, так и в крайнесеверную тайгу (рис. 2). Леса, относящиеся к ней, могут произрастать как на силикатных породах, так и на известняках. Сомкнутость древостоя 0.4–0.6 с учетом выраженного 2-го елового яруса; средняя высота 1-го яруса 15–17 м, средний бонитет IV–V. Примесь березы пушистой к ели до 2, сосны – до 1 единицы по составу древостоя. Еловый подрост жизнеспособен, умеренно обилен. В травяно-кустарничковом ярусе значима роль Ledum palustre в сочетании с Vaccinium myrtillus и Rubus chamaemorus; Trientalis europaea нетипичен. Выделяются три субассоциации, различающиеся по степени заболачивания, общему облику, набору доминантов и сопутствующих видов.
Subass. uliginosum – е. голубичный сфагново-зеленомошный (табл. 1, № 5). Субассоциация, переходная к ельникам голубичным и (отчасти) деренно-морошковым. Произрастает как по окраинам болот на водоразделах, так и в ложбинах и по покатостям коренных берегов рек, в долинах ручьев, в нижней части и при основании пологих горных склонов. Мощность торфяной залежи не более 50 см. Почвы торфянисто- и торфяно-подзолисто-глеевые (Sklyarov, Sharova, 1970; Zaboyeva, 1975; Bobkova, 2006a), различного гранулометрического состава. Нанорельеф от ровного до двух- либо трехкомпонентного (с западинами; 1–2% площади ценоза) кочковатого с преобладанием сфагновых ковров (60–80%); высота кочек 15–30 см. Отмечены и сообщества с нанорельефом из высоких (30 см) кочек и обводненных западин (до 20–40% площади ценоза, глубиной 10–20 см) и формированием ветровально-почвенного комплекса. Высота 1-го яруса древостоя варьирует от 8–9 м в крайнесеверной тайге до 15–17 м у южных границ северной (бонитет при этом меняется от Va до IV), 2-го, развитого в высокоствольных ельниках, – от 5 до 14 м. Средняя сомкнутость 1-го яруса 0.4, 2-го 0.2, средний бонитет V. В подлеске единичные кусты Juniperus communis s. l. Доминанты травяно-кустарничкового яруса – Vaccinium uliginosum, V. myrtillus, V. vitis-idaea, Rubus chamaemorus и Carex globularis (по 10–15%). На тиманских супесях корни Vaccinium uliginosum могут проникать на глубину свыше 1 м; здесь вид служит индикатором более высокой влажности нижних горизонтов профиля (Andreyev, 1935). В числе сопутствующих видов обильнее других Empetrum nigrum s. l., Ledum palustre (по 5–7%) и Equisetum sylvaticum (2%); последние два вида встречаются спорадически. Постоянен Melampyrum pratense s. l.; представители таежного мелкотравья (Linnaea borealis, Orthilia secunda, Luzula pilosa) редки. В ярусе мхов Sphagnum angustifolium, S. russowii и (в западинах) S. flexuosum (по 10–20%) сочетаются c Pleurozium schreberi, Hylocomium splendens и Polytrichum commune (по 15–20%). Дифференциальные виды, сближающие синтаксон с ельниками деренно-морошковыми и отличающие от “типичных” багульниковых – Juniperus communis s. l., Avenella flexuosa и Listera cordata.
Субассоциация обычна в южной полосе северной тайги от Карелии до Урала. В Карелии она известна из района ст. Лоухи (P. equisetoso-myrtillosum (Blagoveshchenskii, 1936)) и с южного берега губы Чупа, в Архангельской обл. – из Пинежского заповедника. В Республике Коми подобные леса описаны на Среднем Тимане (P. polytrichoso-chamaemorosum, на известняках (Andreyev, 1935)) и Печоро-Ижемском водоразделе, в районе Зеленоборского лесного стационара близ г. Вуктыл (P. myrtillosum (Bobkova, 2006a)) и в бассейне Илыча (P. baccoso-globulari-caricosum (Korchagin, 1940)). Крайне восточное местонахождение синтаксона в северной тайге сделано у подножия г. Паснёр на восточном макросклоне Северного Урала, в верховьях р. Няисманья (Sphagnopiceetum caricosum septentrionaliuralense (Soczava, 1930)). Леса этой субассоциации проникают и в крайнесеверную тайгу. В пределах последней в Карелии они известны из национального парка “Паанаярви”, в Архангельской обл. – из верховий р. Сояны (где растут также на известняках (Braslavskaya et al., 2017)), а в Республике Коми – по рекам Понью (Nepomilueva, 1970) и Косью (P. ledoso-sphagnosum (Yudin, 1948, 1954)) и близ п. Сивая Маска (Katenin, 1972). В средней тайге синтаксон не отмечен (рис. 2).
В финской системе классификации болотных местообитаний ельники с господством олиготрофных кустарничков по неглубокой (до 20 см) и, как правило, неоднородной залежи (с верхним слоем верхового и нижним переходного торфа) описаны как “thin-peated spruce heath forests” (Eurola et al., 1984).
Subass. typicum (табл. 1, № 6) наиболее характерна для окраин болот, реже для понижений на боровых террасах рек и в скальных ландшафтах, долин ручьев в заболоченных лесах; в крайнесеверной тайге отмечена на заболоченных водоразделах и в нижней части склонов низкогорий. Торф мощностью (5) 25–150 см. Почвы от торфянисто- и торфяно-подзолисто-глеевых до торфяно-болотных верховых (Sklyarov, Sharova, 1970; Zaboyeva, 1975; Bobkova, Patov, 2006) поверх песка, суглинка либо двучленных отложений, в карбонатных ландшафтах – иногда поверх плитчатого известняка. Нанорельеф не выражен либо (по окраинам болот) кочковатый с высотой кочек от 10–20 до 30–60 см; кочки и сфагновые ковры обычно представлены в равной пропорции; западины нетипичны. Сомкнутость 1-го яруса древостоя (0.1) 0.3–0.7, 2-го 0.1. Высота 1-го яруса 10–17 (22) м; средний бонитет V. В составе подроста есть примесь сосны к ели; в подлеске очень редко встречаются единичные кусты Betula nana. В ярусе кустарничков и трав согосподствуют Ledum palustre, Rubus chamaemorus и Vaccinium myrtillus (по 15–25%) в сопровождении V. vitis-idaea и Carex globularis (по 7–8%). Vaccinium uliginosum, Empetrum nigrum s. l. и Equisetum sylvaticum постоянны, но не обильны. Из таежного мелкотравья чаще встречается Orthilia secunda; Linnaea borealis и Luzula pilosa редки. Моховой покров из Sphagnum angustifolium, S. russowii (по 20%), Polytrichum commune и Pleurozium schreberi (по 10% за счет мелкозалежных описаний); в описаниях с западинами появляется Sphagnum flexuosum. Дифференциальные виды синтаксона, отличающие его от других субассоциаций, – растения верховых болот: Chamaedaphne calyculata и достаточно редкий Sphagnum magellanicum s. l.
Синтаксон обычен в южной полосе подзоны северной тайги от Финляндии (Zwergstrauchbruch (Ruuhijärvi, 1960)) до гор юга Северного Урала (Kolesnikov et al., 1973) с возможной дизъюнкцией ареала на правобережье Средней Печоры. В Керетской Карелии он отмечен на южном берегу губы Чупа и близ оз. Кумозеро, а в Кемской – на материке (P. sphagnosum, P. fontinale-sphagnosum (Rutkovskii, 1933)) и на беломорском острове Комостров. Из Костомукшского заповедника он описан, следуя системе Ж. Браун-Бланке, как Rubo chamaemori-Piceetum abietis K.-Lund 1962 pinetosum sylvestris O. Morozova & V. Korotkov 1999 (Morozova, Korotkov, 1999). В Архангельской обл. ельники данного синтаксона встречены близ г. Онега и на четвертичной террасе Пинеги в бывшем заказнике “Голубинский”, а в Республике Коми – в районе п. Нижний Одес. Отдельные находки этих лесов в крайнесеверной тайге известны в национальном парке “Паанаярви”, в верховьях р. Сояны (Braslavskaya et al., 2017) и на Цилемской Печоре (P. ledoso-sphagnosum (Yudin, 1948, 1954), P. polytrichoso-sphagnosum (Bobkova, Patov, 2006)).
Местами леса субассоциации встречаются и в средней тайге – в Медвежьегорском (Ledo-P. (Vasilevich, Bibikova, 2004)), Кондопожском (в заповеднике “Кивач”) и Прионежском районах Карелии, в южной части национального парка “Кенозерский” (Ledo-P. (Kucherov et al., 2010)), в бассейне Северной Двины и Нижней Вычегды на юго-востоке Архангельской обл. (P. fruticuloso-sphagnosum (Piavchenko, 1957)) и в Якшинском участке Печоро-Илычского заповедника (рис. 2). На северо-западе Карельского перешейка ельники багульниковые сфагновые, развитые здесь на перегнойно-торфяных почвах межсельговых понижений, выходят на границу средней и южной тайги (Dmitrieva, 1973).
Subass. equisetosum sylvaticae – е. хвощово-багульниковый зеленомошно-сфагновый (табл. 1, № 7). Синтаксон, переходный к ельникам хвощово-морошково-черничным (см. ниже), изредка встречается по окраинам болот, заболоченным берегам озер и долинам ручьев. Торфяная залежь малой или средней мощности (15–60 см) поверх песка, скальной породы либо (на Тимане) карбонатной супеси или суглинка. Нанорельеф не выражен или выражен слабо, высота кочек не более 10–25 см. Сомкнутость 1‑го яруса древостоя (0.2) 0.3–0.7, его высота составляет 10–21 (27) м; бонитет возрастает до IV. Сомкнутость 2-го яруса 0.1 при высоте 9–13 (17) м. В травяно-кустарничковом ярусе наряду с Ledum palustre, Vaccinium myrtillus и Rubus chamaemorus в число согосподствующих видов входит Equisetum sylvaticum (все по 15%). ПП Carex globularis, Vaccinium uliginosum и Empetrum nigrum s. l. снижается до 2–5%, а постоянство – до III–IV класса. Из видов мелкотравья Linnaea borealis встречается спорадически, Orthilia secunda редка, Luzula pilosa почти исчезает. Сравнительно с другими синтаксонами, ПП мохового яруса уменьшается до 75%. Господство в нем переходит к Sphagnum russowii, Polytrichum commune (по 20%), Hylocomium splendens и Pleurozium schreberi (по 10%). Sphagnum angustifolium редок, растет в примеси к S. russowii. Олиготрофные кустарнички (Vaccinium uliginosum, Empetrum nigrum s. l., Ledum palustre) дифференцируют синтаксон от ельников морошково-черничных сфагновых.
Субассоциация несколько раз описана в северной тайге Керетской Карелии по южному берегу губы Чупа, близ ст. Лоухи (P. myrtilloso-polytrichosum (Blagoveshchenskii, 1936)) и в районе бывшего Лоухи-Кестеньгского тракта (P. sphagnosum (Sokolova, 1936)). Она известна также на Среднем Тимане (P. chamaemoroso-equisetoso-sphagnosum и P. polytrichoso-equisetosum на суглинках, P. polytrichoso-myrtillosum на супесях (Andreyev, 1935)) и на водоразделе Ижмы и Печоры, а в средней тайге найдена на юго-востоке Архангельской обл. в окрестностях п. Урдома (рис. 2). В крайнесеверную тайгу ельники рассматриваемого синтаксона не проникают. Указаний на них в зарубежной литературе нами также не найдено.
P. sphagno angustifolii-myrtillosum (Sphagno angustifolii-Myrtillo-P.) – е. морошково-черничный сфагновый (табл. 1, № 8–10). Средне- и северотаежная ассоциация с единичными местонахождениями в крайнесеверной тайге. Сомкнутость 1-го яруса древостоя (0.2) 0.3–0.8, высота от 14 до 20–21 (23) м; бонитет древостоя обычно IV. Сомкнутость 2-го яруса древостоя 0.1. Всегда обилен еловый подрост (ПП 10%). В составе травяно-кустарничкового яруса господство в различных пропорциях разделяют Vaccinium myrtillus, Rubus chamaemorus, Carex globularis и Equisetum sylvaticum. В ярусе мхов всегда преобладает Sphagnum angustifolium (35–45%) в сопровождении S. russowii, Polytrichum commune и Pleurozium schreberi (по 10–15%); Hylocomium splendens не столь обилен (3–7%). Ассоциация дифференцируется негативно, по утрате диагностической роли олиготрофными кустарничками (Ledum palustre, Vaccinium uliginosum, Empetrum nigrum s. l.), которые встречаются намного реже, чем в ельниках багульниковых, и всегда в малом обилии. Позитивная дифференциация выражена слабее, по присутствию Sorbus aucuparia s. l. и Trientalis europaea. В то же время произрастание Equisetum sylvaticum и малообильных видов мелкотравья (Linnaea borealis, Orthilia secunda, Luzula pilosa), а также сопровождающих их мхов (Dicranum polysetum, D. scoparium) объединяет ельники морошково-черничные с багульниковыми.
Выделены два варианта, слабо разграниченные флористически, но хорошо различимые по доминантам травяно-кустарничкового яруса.
Var. Equisetum sylvaticum – е. хвощово-морошково-черничный сфагновый (табл. 1, № 8). Встречается в притеррасной части речных долин, по склонам долин ручьев, по краю болот у подножия склонов, в ложбинах на водораздельных плато. Обычно произрастает на силикатных почвообразующих породах, на карбонатных и сульфатных – намного реже, только если они перекрыты четвертичными отложениями. Мощность торфа от (5) 15–25 до 60–90 см. Почвы от торфянисто- и торфяно-глеевых (Kielland-Lund, 1981; Bobkova, 2006b) до торфяно-болотных (Sklyarov, Sharova, 1970; Zaboyeva, 1975) на песках, суглинках или двучленных отложениях. Нанорельеф из приствольных повышений высотой 15–35 (50) см и ковров с преобладанием последних (60–80% площади ценоза). При наиболее развитом нанорельефе в составе ковра появляется Sphagnum flexuosum (до 10%). Примесь березы к ели в 1-м ярусе древостоя – до 3 единиц по составу, сосны – незначительна. 2-й ярус также березово-еловый. В подлеске единичные кусты Sorbus aucuparia s. l. В ярусе трав и кустарничков доминирует Equisetum sylvaticum (20%) в сопровождении Vaccinium myrtillus, Rubus chamaemorus, Carex globularis (по 10–15%) и Vaccinium vitis-idaea (5%). Вариант дифференцируется негативно, по отсутствию Chamaedaphne calyculata и Sphagnum magellanicum s. l. С формальных позиций именно его стоило бы назвать “типичным”.
Ельники хвощового варианта нередки в южной полосе северной тайги. Здесь они описаны в Костомукшском заповеднике (Morozova, Korotkov, 1999), по Карельскому берегу Белого моря, близ г. Онега, в Пинежском заповеднике, на Среднем Тимане (P. sphagnoso-equisetosum (Andreyev, 1935)), Печоро-Ижемском водоразделе и в районе Печорского водохранилища (P. equisetoso-sphagnosum (Bobkova, 2006b)). В средней тайге леса этого синтаксона столь же обычны в Кондопожском (заповедник “Кивач”) и Пудожском (Усть-Река) районах Карелии, на левобережье р. Устьи на юге Архангельской обл., по р. Выми (P. globulari-caricoso-sphagnosum) и в верховьях Вычегды (P. chamaemoroso-equisetoso-sphagnosum (Yudin, 1948, 1954)). На Среднем Урале они найдены в горно-лесном поясе хребта Басеги, где ранее были известны лишь ассоциации мезотрофных заболоченных ельников (Balandin, Ladygin, 2002). В крайнесеверной тайге известна единственная находка на р. Тобыш (P. globulari-caricoso-sphagnosum (Yudin, 1948, 1954)) (рис. 3).
По данным литературы, ельники рассматриваемого варианта очень обычны в Скандинавии. Судя по согосподству Equisetum sylvaticum, Rubus chamaemorus и Vaccinium myrtillus при малом обилии болотных кустарничков, леса именно этого либо близкого к нему синтаксона описаны из Юго-Восточной Норвегии как Chamaemoro-Piceetum abietis typicum K.-Lund 1962 (союз Vaccinio-Piceion порядка Vaccinio-Piceetalia). В моховом покрове лесов этой субассоциации, как и ряда других синтаксонов ельников сфагновых Средней и Атлантической Европы (см. выше), Sphagnum angustifolium доминирует совместно со S. girgensohnii. Но это именно олиготрофный синтаксон, исходя из значений pHH20 торфяной залежи, равных 3.6–3.9 (Kielland-Lund, 1981). К. Дирсен (Dierßen, 1996) указывает на господство Sphagnum angustifolium при подчиненной роли S. girgensohnii для ассоциации Rubo chamaemori-Piceetum (K.-Lund 1962) в целом, но это противоречит данным автора синтаксона (Kielland-Lund, 1981).
Л. Пэльссон (Påhlsson, 1994) приводит заболоченные ельники Скандинавии, в которых согосподствуют Equisetum sylvaticum, Vaccinium myrtillus (а также V. uliginosum), Sphagnum angustifolium, S. girgensohnii и S. russowii, как Picea abies-Vaccinium spp.-Sphagnum spp.-typ. Это явно сводный синтаксон, в котором объединены как олиготрофные (нескольких разных ассоциаций), так и мезотрофные сфагновые ельники.
Var. typica (табл. 1, № 9–10) от предыдущего варианта отличают присутствие Chamaedaphne calyculata и Sphagnum magellanicum s. l. Подрост еловый с крайне незначительной примесью березы и сосны; подлесок не выражен. Вариант представлен двумя модификациями (в традиции школы Ж. Браун-Бланке – доминантными формами), тождественными флористически, но различающимися по общему облику, отчасти также по ареалу. В обеих из них Equisetum sylvaticum постоянен, но не обилен.
F. Carex globularis – е. круглоосоковый сфагновый (табл. 1, № 9). Сообщества, развитые на силикатных породах – на заболоченных водоразделах, в депрессиях коренных берегов рек и в болотистых речных долинах, при основании склонов озов, намного реже по окраинам безлесных болот. Торф мощностью 15–110 см; почвы торфянисто- и торфяно-глеевые (Sambuk, 1932; Sokolova, 1936; Yudin, 1948, 1954), формируются на песках, суглинках либо щебнистых отложениях. Нанорельеф кочковатый с возможным преобладанием по площади (до 60–80%) как кочек (высотой от 10–15 до 70–80 см), так и ковров, реже волнистый, повторяющий форму минерального дна. Как и в лесах предыдущего синтаксона, при наиболее дифференцированном нанорельефе, порождающем переувлажнение участков ковра, в формировании последних принимает участие Sphagnum flexuosum. Примесь березы к ели в 1-м ярусе древостоя составляет 2 единицы по составу. Характерен ковер обильной (ПП 30%) Carex globularis, создающей нежно-зеленый аспект. Ей сопутствуют Vaccinium myrtillus (20%) и Rubus chamaemorus (15%), в меньшей степени Vaccinium vitis-idaea (5%).
В северной тайге Карелии синтаксон известен из Костомукшского заповедника (Morozova, Korotkov, 1999; Kutenkov, 2015) и района бывшего Лоухи-Кестеньгского тракта (Sphagnetum globulari-caricoso-piceiosum (Sokolova, 1936)). В северной тайге Архангельской обл. он описан из окрестностей г. Онега и Пинежского заповедника (Carici globularis-P. (Kucherov et al., 2010)), а в Республике Коми – с Печоро-Ижемского водораздела и со Средней Печоры близ д. Кипиево (P. sphagno-caricosum (Sambuk, 1932)). На северо-востоке Коми леса данного синтаксона по р. Сыне проникают в крайнесеверную тайгу (P. globulari-caricoso-sphagnosum (Yudin, 1948, 1954)). На восточном макросклоне Северного Урала ельники сфагновые с Carex globularis найдены в верховьях р. Северной Сосьвы (Sphagnopiceetum caricosum septentrionaliuralense (Soczava, 1930)). В северной тайге Западной Сибири этот же синтаксон с доминированием Sphagnum russowii в моховом покрове описан в верхнем течении р. Таз как P. obovatae ledoso-sphagnosum var. caricosum globularis (Neshatayev et al., 2002). В средней тайге Карелии рассматриваемые ельники описаны из района бывшего природного парка “Хиисьярви” как Mustikka-corpi (Brandt, 1933), а также из Заонежской (заповедник “Кивач”) и Пудожской (национальный парк “Водлозерский”) Карелии. Ю. Д. Цинзерлинг (Zinserling, 1932) приводит эти леса (P. globulari-caricosum) как характерные для юга Карелии от Петрозаводска до границы с Ленинградской областью. Их встречаемость еще более возрастает на юге Архангельской обл. Здесь эти леса известны из северной части национального парка “Кенозерский” (где оказались “растворенными” в синтаксоне Sphagno girgensohnii-P. chamaemoretosum (Kucherov et al., 2010)) и из ряда местностей в бассейне р. Устьи. В средней тайге Республики Коми аналогичные ельники отмечены по р. Выми (P. globulari-caricoso-sphagnosum (Yudin, 1948, 1954)) и в Якшинском участке Печоро-Илычского заповедника, а на Среднем Урале – в горно-лесном поясе Басег. В Вологодской обл. (заказник “Ярбозерский бор” к северу от оз. Белое) ельники сфагновые с Carex globularis выходят на границу средней и южной тайги (рис. 3). В Финляндии подобные леса хорошо известны как из северной (Ruuhijärvi, 1960), так и из средней (Eurola, 1962) тайги под названием Carex globularis-Reisermoore.
В сводке Л. П. Рысина и Л. И. Савельевой (Rysin, Savelieva, 2002) ельники с C. globularis отнесены к ассоциации P. caricoso-sphagnosum (с преобладанием крупных видов осок) в группе ассоциаций ельников травяно-сфагновых, что ошибочно.
F. typica (табл. 1, № 10) по глубокой (до 2.6–3 м) торфяной залежи может встречаться не только на силикатных, но изредка и на карбонатных породах. На силикатах известны и мелкозалежные сообщества с мощностью торфа (15) 30–40 см. Ельники этого синтаксона произрастают по окраинам болот (безлесных либо облесенных сосной) и низким заболоченным берегам крупных озер, по ложбинам стока и понижениям речных террас, при основании склонов озов, очень редко (на Ямале) в затопляемых поймах. Почвы от торфянисто-подзолисто-глееватых иллювиально-железисто-гумусовых на песках (Bobkova, 2006a) до торфяно-болотных на суглинках, двучленных отложениях или на скальном основании. Нанорельеф обычно двухкомпонентный с кочками высотой 10–30 см и преобладанием ковров (70–85% площади ценоза), реже кочек (до 60%). Иногда наблюдается и трехкомпонентный нанорельеф с западинами (3–10%) и преобладанием ковров (50–70% площади ценоза); в этом случае высота кочек достигает 60–80, а глубина западин – 20–30 см. Реже нанорельеф не развит. В средней тайге древостой может достигать высоты 23–24 м и III класса бонитета. В 1-м ярусе к ели выражена примесь и березы, и сосны, в обоих случаях до 1 единицы по составу. 2‑й ярус ели выражен слабо. В травяно-кустарничковом ярусе согосподствуют Vaccinium myrtillus (25%), V. vitis idaea и Rubus chamaemorus (по 10%); Carex globularis не обильна. В конце лета, когда листья Vaccinium myrtillus облетают, ярус выглядит сквозистым. В моховом покрове – спорадическая примесь Sphagnum magellanicum s. l. (чаще на кочках; 3% ПП). S. flexuosum нехарактерен. Дифференциация от ельников пушицевых сфагновых – как по наличию на приствольных повышениях мезофитов (Orthilia secunda, Luzula pilosa, Dicranum polysetum, D. scoparium), исчезающих при подтоплении, так и негативная, по редкости Eriophorum vaginatum.
Синтаксон обычен в северной тайге Карелии (P. chamaemorosum (Цинзерлинг, 1932)), в том числе в Костомукшском заповеднике (Morozova, Korotkov, 1999; Kutenkov, 2015), природном парке “Тулос” и на беломорских островах Комостров и Чернецкий. В северной тайге Архангельской обл. олиготрофный ельник морошково-черничный описан в ложбине четвертичной террасы Пинеги в пределах бывшего заказника “Голубинский”. В Республике Коми сходные леса отмечены в районе Зеленоборского стационара (P. hylocomioso-sphagnosum (Bobkova, 2006a)) и на Среднем Тимане. В последнем из регионов описаны как P. polytrichoso-chamaemorosum на супесях, так и P. sphagnoso-chamaemorosum с согосподством Sphagnum angustifolium, S. girgensohnii, S. russowii и S. riparium, “вкрапленных друг в друга” в сфагновом ковре по залежи мощностью свыше 1 м (Andreyev, 1935: 29). В крайнесеверной тайге известна изолированная находка олиготрофного ельника морошково-черничного на Южном Ямале.
В средней тайге Карелии леса данного синтаксона встречаются почти повсеместно. Их описания сделаны в Суоярвском (Хиисьярви, Толвоярви), Кондопожском (Кивач), Прионежском (Деревянка) и Пудожском (национальный парк “Водлозерский”) районах. Столь же широко они распространены и в Финляндии, как в средней, так и в северной тайге (Moltebeerbrücher (Ruuhijärvi, 1960, Eurola, 1962), Rubus chamaemorus spruce mires (Eurola et al., 1984)). На юге Архангельской обл. рассматриваемые ельники найдены в южной части национального парка “Кенозерский”, где они ранее ошибочно рассматривались в составе ассоциации мезотрофных ельников черничных сфагновых Sphagno girgensohnii-P. (Kucherov et al., 2010). Они описаны также в бассейне р. Устьи и однократно – в окрестностях п. Урдома в бассейне Нижней Вычегды. Восточнее подобные ельники, видимо, не проникают. На Карельском перешейке (Шевелево, заказник “Озеро Щучье”) и в Вологодской обл. к северу от оз. Белое они выходят на границу южной тайги (рис. 3), но также не распространяются далее вглубь нее. Указания на ельники черничные сфагновые в южной тайге Европейской России (Rysin, Savelieva, 2002) должны относиться к мезотрофным сообществам со Sphagnum girgensohnii, зональный ареал которых более широк.
P. sphagno angustifolii-eriophorosum (Sphagno angustifolii-Eriophoro-P.) – е. пушицевый сфагновый (табл. 1, № 11). Формируется при подтоплении по окраинам болотных массивов, реже по склонам долин ручьев, на надпойменных террасах, а в крайнесеверной и (на правобережье Печоры) северной тайге – на заболоченных водоразделах, практически всегда на силикатных породах. Почвы от торфяно-глеевых (Korchagin, 1940) до торфяно-болотных на средних и тяжелых суглинках, очень редко на песках. Торфяная залежь мощностью 30–100 см. Нанорельеф кочковатый. Обычно он двухкомпонентный с преобладанием сфагновых ковров (50–90% площади сообщества) и кочками высотой 10–35 см, реже трехкомпонентный с топкими западинами (до 20%). Сомкнутость 1-го яруса древостоя варьирует от 0.2 до 0.7 при высоте (6) 12–22 м; формально средний бонитет IV, но большое число деревьев усыхает. Примесь березы пушистой к ели – до 4 единиц по составу 1-го яруса. Сомкнутость 2-го яруса 0.1; часто он отсутствует. Подрост ели малочислен и угнетен, также с примесью березы. В травяно-кустарничковом ярусе согосподствуют Rubus chamaemorus, Carex globularis и Eriophorum vaginatum (ПП по 10–15%) в сопровождении Vaccinium myrtillus, V. vitis-idaea и Equisetum sylvaticum (по 3–5%). Болотные кустарнички единичны, за исключением спорадически встречающейся Chamaedaphne calyculata (1%). Из таежного мелкотравья сохраняются лишь Trientalis europaea (способный расти и на приствольных повышениях, и на сфагновом ковре) и Linnaea borealis (на валеже), но и они достаточно редки. В топком моховом ковре к Sphagnum angustifolium (30%) примешивается S. flexuosum (10%), на кочках – S. russowii (10%), S. magellanicum s. l. и S. centrale (по 5%). Собственные детерминанты ассоциации – Eriophorum vaginatum и Pinus sibirica, образующая малочисленную, но постоянную примесь к Picea abies s. l. в древостое верхнепечорских и уральских сообществ. Последнее не случайно: по мере продвижения на восток в северной и средней тайге Западной Сибири ель в условиях олиготрофного заболачивания все более уступает место кедру (Krylov, 1961; Neshatayev et al., 2002), формируя древостои лишь мезотрофных заболоченных лесов (Rysin, Savelieva, 2002).
Ельники пушицевые сфагновые встречаются во всех подзонах тайги Европейской России. В полосе крайнесеверной тайги они описаны в Лапландском заповеднике, на Цилемской Печоре по р. Косме (P. chamaemoroso-sphagnosum), с междуречья Космы и Тобыша (P. eriophoroso-chamaemorosum) и по руч. Седьёль (P. globulari-caricoso-sphagnosum (Yudin, 1948, 1954)), а также в бассейне р. Усы (Сынянырд; P. chamaemoroso-sphagnosum (Sambuk, 1932)). Они отмечены также на севере Беломорско-Кулойского плато (Leontiev, 1937). В собственно северной тайге данные ельники известны с левобережья р. Онеги напротив г. Онега и в верховьях Илыча (Betuleto-Piceetum globulari-caricosum (Korchagin, 1940)). В. А. Мартыненко (Martynenko, 1999) считает подобные леса широко распространенными в северной тайге Республики Коми. В средней тайге они отмечены в заповеднике “Кивач”, в национальном парке “Кенозерский” (где ранее были включены в ассоциацию Ledo-P. (Kucherov et al., 2010)), в верховьях Вычегды (P. globulari-caricoso-sphagnosum (Yudin, 1948, 1954), P. caricoso-sphagnosum (Kolesnikov, 1985)) и Печоры и в низкогорьях Басег (рис. 3). Указания на то, что ареал ельников пушицевых сфагновых (P. eriophoroso-sphagnosum) ограничен лишь северной (в широком смысле) тайгой (Rysin, Savelieva, 2002), требуют коррекции.
ЗАКЛЮЧЕНИЕ
Всего в Европейской России и на Урале, а также на юге Ямала нами описано 7 ассоциаций олиготрофных сфагновых ельников с тремя субассоциациями, двумя вариантами и двумя модификациями. По своей зональной приуроченности три синтаксона (ельники вороничные, ерниковые и голубичные) характеризуются “центром тяжести” ареала в крайнесеверной тайге, еще три (деренно-морошковые, а также голубичная и хвощовая субассоциации ельников багульниковых) – в южной полосе северной тайги. У четырех синтаксонов (“типичные” ельники багульниковые и все варианты и модификации морошково-черничных) “центр тяжести” ареала охватывает и южную полосу северной тайги, и среднюю тайгу. Ельники пушицевые сфагновые в равной мере представлены во всех рассмотренных широтных выделах.
С точки зрения меридиональной зональности, один синтаксон (ельники деренно-морошковые) является выраженно “западным”, его ареал ограничен западом Карелии. Еще трем синтаксонам ельников (вороничным, багульниковым хвощовой субассоциации и “типичной” модификации морошково-черничных) свойственно умеренное западное тяготение: сообщества этих синтаксонов полностью либо преимущественно отсутствуют на правобережье Печоры и тем более на Урале. У ельников голубичных сфагновых ареал, напротив, расширяется по мере приближения к Уралу. Сообщества оставшихся шести синтаксонов сопоставимо часто встречаются по всему северу Европейской России от Лапландии и/или Карелии до Урала включительно. Два из них (ельники ерниковые и осоковая модификация морошково-черничных) отмечены и на севере Западной Сибири.
Заметим, что с учетом единичных местонахождений на периферии ареала (особенно изолированных ямальских находок) амплитуды толерантности сообществ большинства синтаксонов по отношению к факторам теплообеспеченности и континентальности климата в большинстве случаев оказываются значительно шире (табл. 2), чем следует из анализа основной части их ареалов. Лишь ареал ельников деренно-морошковых строго ограничен северной тайгой Западной Карелии. Следует, однако, более внимательно изучить возможное распространение ельников данной ассоциации в зарубежной Фенноскандии.
По своей топоэдафической приуроченности, ельники четырех синтаксонов (голубичные, багульниковые голубичной и хвощовой субассоциаций и “типичная” модификация морошково-черничных) могут произрастать не только на силикатных, но и на карбонатных породах, если последние перекрыты мощным слоем торфа. Леса всех прочих синтаксонов встречаются только на силикатных почвообразующих породах, включая случаи перекрытия коренных карбонатных либо сульфатных пород изолирующим слоем четвертичных силикатных отложений.
Список литературы
[A checklist…] Список лихенофлоры России. 2010. СПб. 194 с.
[Aleksandrova, Yurkovskaya] Александрова В.Д., Юрковская Т.К. 1989. Геоботаническое районирование Нечерноземья европейской части СССР. Л. 64 с.
[Andreyev] Андреев В.Н. 1935. Лесная растительность южного Тимана. – Тр. Полярной комиссии СОПС АН СССР. 24: 7–64.
[Balandin, Ladygin] Баландин С.В., Ладыгин И.В. 2002. Флора и растительность хребта Басеги (Средний Урал). Пермь. 191 с.
[Blagoveshchenskii] Благовещенский Г.А. 1936. Эволюция растительного покрова болотного массива “1007 км” у ст. Лоухи (Карелия). – Тр. БИН АН СССР. 3 (3): 141–232.
[Bobkova] Бобкова К.С. 2006a. Еловые леса северной подзоны тайги. – В кн.: Коренные еловые леса Севера: Биоразнообразие, структура, функции. СПб. С. 52–98.
[Bobkova] Бобкова К.С. 2006b. Еловые леса средней подзоны тайги. – В кн.: Коренные еловые леса Севера: Биоразнообразие, структура, функции. СПб. С. 99–158.
[Bobkova, Patov] Бобкова К.C., Патов А.И. 2006. Еловые леса крайнесеверной подзоны тайги (притундровой зоны). В кн.: Коренные еловые леса Севера: Биоразнообразие, структура, функции. СПб. С. 25–33.
Brandt A. 1933. Hiisjarven luonnonpuiston kasvillisuudesta. – Silva Fenn. 32: 1–108.
[Braslavskaya et al.] Браславская Т.Ю., Горячкин С.В., Кутенков С.А., Мамонтов В.Н., Попов С.Ю., Пучнина Л.В., Сидорова О.В., Торхов С.В. Чуракова Е.Ю. 2017. Флора и растительность Беломорско-Кулойского плато. Архангельск. 302 с.
[Cherepanov] Черепанов С.К. 1995. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). СПб. 991 с.
[Chertovskoi] Чертовской В.Г. 1978. Еловые леса европейской части СССР. М. 176 с.
Chytrý M., Zelený D., Návratilová J., Sádlo J. 2013. Boreokontinentální jehličnaté lesy. – In: Vegetace České republiky. T. 4: Lesní a křovinná vegetace. Praha. S. 380–432.
Dierßen K. 1996. Vegetation Nordeuropas. Stuttgart. 839 S.
[Dmitrieva] Дмитриева Е.В. 1973. Ельники северо-западной части Карельского перешейка. – Лесоведение. 2: 51–64.
[Dylis] Дылис Н.В. (ред.) 1974. Программа и методика биогеоценологических исследований. М. 404 с.
Ellenberg H., Weber H.E., Düll R., Wirth W., Werner W., Paulißen D. 1992. Zeigerwerte von Pflanzen in Mitteleuropa. 2 Aufl. – Scripta Geobot. 18: 1–258.
Eurola S. 1962. Über die regionale Einteilung der südfinnischen Moore. – Ann. Bot. Soc. Zool. Bot. Fenn. “Vanamo”. 33 (2): 1–243.
Eurola S., Hicks S., Kaakinen E. 1984. Key to Finnish mire types. – In: European Mires. L. 117 p.
[Gromtsev] Громцев А. Н. 2008. Основы ландшафтной экологии европейских таежных лесов России. Петрозаводск. 238 с.
Ignatov M.S., Afonina O.M., Ignatova E.A. 2006. Check-list of mosses of East Europe and North Asia. – Arctoa: Бриологический журнал. 15: 1–130.
[Isachenko, Lavrenko] Исаченко Т.И., Лавренко Е.М. 1980. Ботанико-географическое районирование. – В кн.: Растительность европейской части СССР. Л. С. 10–22.
[Katenin] Катенин А.Е. 1972. Растительность лесотундрового стационара. – В кн.: Почвы и растительность восточноевропейской лесотундры. Л. С. 118–259.
Kielland-Lund J. 1981. Die Waldgesellschaften SO Norwegens. – Phytocoenologia. 9 (1–2): 53–250.
[Kolesnikov] Колесников Б.П. 1985. Лесная растительность юго-восточной части бассейна Вычегды. Л. 216 с.
[Kolesnikov et al.] Колесников Б.П., Зубарева Р.С., Смологонов Е.П. 1973. Лесорастительные условия и типы леса Свердловской области. Свердловск. 176 с.
[Korchagin] Корчагин А.А. 1940. Растительность северной половины Печорско-Ылычского заповедника. – В кн.: Труды Печорско-Ылычского заповедника. Т. 2. С. 5–415.
[Korchagin] Корчагин А.А. 1956. Еловые леса западного Притиманья в бассейне р. Мезенской Пижмы. – Учен. записки Ленингр. гос. ун-та. Сер. Геогр. 11: 111–239.
[Krylov] Крылов Г.В. 1961. Леса Западной Сибири. М. 255 с.
[Kucherov] Кучеров И.Б. 2018. Эколого-ценотическое разнообразие светлохвойных лесов средней и северной тайги Европейской России: Автореф. дис. … докт. биол. наук. СПб. 46 с.
[Kucherov et al.] Кучеров И.Б., Разумовская А.В., Чуракова Е.Ю. 2010. Еловые леса национального парка “Кенозерский” (Архангельская область). – Бот. журн. 95 (9): 1268–1301.
[Kucherov, Kutenkov] Кучеров И.Б., Кутенков С.А. 2011. Травяно-сфагновые сосняки средней и северной тайги Европейской России. – Бот. журн. 96 (6): 733–763.
[Kucherov, Kutenkov] Кучеров И.Б., Кутенков С.А. 2012. Кустарничковые сфагново-зеленомошные и сфагновые сосняки средней и северной тайги Европейской России. – Тр. Карельск. Науч. Центра РАН. Сер. Биогеогр. 1: 16–32.
[Kutenkov] Кутенков С.А. 2008. Болотные и заболоченные леса. – В сб.: Скальные ландшафты Карельского побережья Белого моря: природные особенности, хозяйственное освоение, меры по сохранению. Петрозаводск. С. 41–51.
[Kutenkov] Кутенков С.А. 2015. Анализ растительности ельников чернично-сфагновых северотаежной подзоны Восточной Фенноскандии. – Тр. Гос. природн. заповедника “Костомукшский”. 1: 61–68.
[Kutenkov et al.] Кутенков С.А., Стойкина Н.В., Бойчук М.А. 2009. Болота и заболоченные земли. – В сб.: Природный комплекс горы Воттоваара: особенности, современное состояние, сохранение. Петрозаводск. С. 35–47.
[Kutenkov, Kuznetsov] Кутенков С.А., Кузнецов О.Л. 2013. Разнообразие и динамика заболоченных и болотных лесов Европейского Севера России. – В кн.: Разнообразие и динамика лесных экосистем России. Кн. 2. М. С. 152–206.
[Leontiev] Леонтьев А.М. 1937. Растительность Беломорско-Кулойской части Северного края. – Тр. БИН АН СССР. 3 (2): 81–222.
[Lyubimova] Любимова А.А. 1937. Растительность и почвы побережья оз. Ловозеро (Кольский полуостров). – Тр. БИН АН СССР. 3 (2): 345–489.
[Martynenko] Мартыненко В.А. 1999. Темнохвойные леса. – В кн.: Леса Республики Коми. М. С. 133–184.
[Medvedeva et al.] Медведева В.М., Корнилова Л.И., Вайнблат В.З. 1980. Основные типы заболоченных ельников Карелии. – В кн.: Болотно-лесные системы Карелии и их динамика. Л. С. 78–99.
[Melekhov] Мелехов И.С. 1966. Леса Мурманской области (Кольский полуостров). – В кн.: Леса СССР. Т. 1. С. 70–77.
[Morozova, Korotkov] Морозова О.В., Коротков В.Н. 1999. Классификация лесной растительности Костомукшского заповедника. – Заповедное дело. 5: 56–78.
NASA surface meteorology and solar energy. 2006. http://eosweb.larc.nasa.gov (Accessed 15.05.2006).
[Nekrasova] Некрасова Т.П. 1935. Очерк растительности Лапландского заповедника. – Тр. Ленингр. о-ва естествоиспыт. 64 (2): 239–272.
[Nepomilueva] Непомилуева Н.И. 1970. Кедр сибирский (Pinus sibirica Tour.) на северной границе ареала в Коми АССР. – Бот. журн. 55 (7): 1011–1025.
[Neshatayev et al.] Нешатаев В.Ю., Потокин А.Ф., Томаева И.Ф., Егоров А.А., Добрыш А.А., Чернядьева И.В., Потемкин А.Д. 2002. Растительность, флора и почвы Верхне-Тазовского государственного заповедника. СПб. 154 с.
Oberdorfer E. 1992. Süddeutsche Pflanzengesellschaften. Teil 4: Wälder und Gebüsche. 2 Aufl. Jena. A. Textband. 282 S. – B. Tabellenband. 580 S.
Påhlsson L. (ed.) 1994. Vegetationstyper i Norden. Köpenhavn. 627 s.
[Piavchenko] Пьявченко Н.И. 1957. Типы заболачивания лесов в бассейне Северной Двины. – Тр. Ин-та леса АН СССР. 36: 5–55.
[Ponyatovskaya] Понятовская В.М. 1964. Учет обилия и особенности размещения видов в естественных растительных сообществах. – В кн.: Полевая геоботаника. Т. 3. М., Л. С. 209–299.
[Popov] Попов П.П. 2005. Ель европейская и сибирская: структура, интерградация и дифференциация популяционных систем. Новосибирск. 231 с.
[Ramenskaya] Раменская М.Л. 1983. Анализ флоры Мурманской области и Карелии. Л. 203 с.
Regel K. 1923. Die Pflanzendecke der Halbinsel Kola. T. I. Lapponia Varsugae. – In: Mémoires de la Faculté des Sciences de L’Université de Lithuanie. Kaunas. 246 S.
[Rutkovskii] Рутковский В.И. 1933. Типы лесов Кемского края АКССР. – Тр. Ин-та изучения леса АН СССР. 1: 1–97.
Ruuhijärvi R. 1960. Über die regionale Einteilung der nordfinnischen Moore. – Ann. Bot. Soc. Zool. Bot. Fenn. “Vanamo”. 31 (1): 1–360.
[Rysin, Savelieva] Рысин Л.П., Савельева Л.И. 2002. Еловые леса России. М. 335 с.
[Safronova, Yurkovskaya] Сафронова И.Н., Юрковская Т.К. 2015. Зональные закономерности растительного покрова равнин Европейской России и их отображение на карте. – Бот. журн. 100 (11): 1121–1141.
[Sambuk] Самбук Ф.В. 1932. Печорские леса. – Тр. Бот. музея АН СССР. 24: 63–245.
[Shennikov] Шенников А.П. 1964. Введение в геоботанику. Л. 447 с.
[Sklyarov, Sharova] Скляров Г.А., Шарова А.С. 1970. Почвы лесов Европейского Севера. М. 272 с.
[Soczava] Сочава В.Б. 1927. Ботанический очерк лесов Полярного Урала от р. Нельки до р. Хулги. – Тр. Бот. музея АН СССР. 21: 1–78.
[Soczava] Сочава В.Б. 1930. К фитосоциологии темнохвойного леса. – Журн. Русск. бот. о-ва. 15 (1–2): 7–41.
[Sokolova] Соколова Л.А. 1936. Растительность района Лоухи-Кестеньгского тракта (Карелия). – Тр. БИН АН СССР. 3 (3): 241–306.
[Sukachev] Сукачев В.Н. 1931. Руководство к исследованию типов леса. 3-е изд. М. 328 с.
Tukhanen S. 1980. Climatic parameters and indices in plant geography. – Acta Phytogeogr. Suec. 67: 1–105.
[Vasilevich] Василевич В.И. 1995. Доминантно-флористический подход к выделению растительных ассоциаций. – Бот. журн. 80 (6): 28–39.
[Vasilevich, Bibikova] Василевич В.И., Бибикова Т.В. 2004. Сфагновые ельники Европейской России. – Бот. журн. 89 (5): 734–748.
Willner W., Grabherr G. (eds.) 2007. Die Wälder und Gebüsche Österreichs. Ein Bestimmungswerk mit Tabellen. Im 2 Bände. München. 608 S.
[Yudin] Юдин Ю.П. 1948. Темнохвойные леса Коми АССР (геоботаническая характеристика): Дис. … докт. биол. наук. Сыктывкар. 323 с.
[Yudin] Юдин Ю.П. 1954. Темнохвойные леса. – В сб.: Производительные силы Коми АССР. М.; Л. Т. 3. Вып. 1. С. 42–125.
[Zaboyeva] Забоева И.А. 1975. Почвы и земельные ресурсы Коми АССР. Сыктывкар. 344 с.
[Zinserling] Цинзерлинг Ю.Д. 1932. География растительного покрова северо-запада европейской части СССР. Л. 376 с.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал