

Ботанический журнал, 2019, T. 104, № 10, стр. 1574-1587
БИОЛОГИЯ ОПЫЛЕНИЯ JUNIPERUS EXCELSA И J. DELTOIDES (CUPRESSACEAE) НА ЮЖНОМ БЕРЕГУ КРЫМА
С. П. Корсакова 1, *, И. С. Саркина 1, Н. А. Багрикова 1, **
1 ФГБУН “Никитский ботанический сад – Национальный научный центр РАН”
298648 г. Ялта, пгт Никита, спуск Никитский, 52, Россия
* E-mail: korsakova2002@mail.ru
** E-mail: nbagrik@mail.ru
Поступила в редакцию 13.09.2019
После доработки 30.09.2019
Принята к публикации 08.10.2019
Аннотация
В результате мониторинговых исследований изменений продолжительности и сезонной ритмики пыления Juniperus excelsa и J. deltoides установлено, что амплитуда экотопической изменчивости начала лета пыльцы на Южном берегу Крыма коррелирует с изменениями температур зимне-весенних месяцев. В связи с ростом температур, в последние десятилетия начало пыления J. excelsa смещается на более ранние сроки со средней скоростью 7 дней за 10 лет, J. deltoides – 4 дня. Анализ многолетних изменений гидротермических показателей, лимитирующих формирование опылительной капли, показал, что неблагоприятные условия во время вылета пыльцы в Крыму складываются для J. excelsa и J. deltoides, в среднем, в трех-четырех годах из 10. Установлено, что в природном заповеднике “Мыс Мартьян” основными экологическими факторами, лимитирующими функционирование опылительной капли у J. excelsa являются температура воздуха и обилие осадков, у J. deltoides – температура и влажность воздуха, обилие осадков. Выявлено, что период 2001–2010 гг. не был благоприятным для образования полноценных семян у J. excelsa. При дальнейшем росте средних температур воздуха в зимне-весенние месяцы, существует вероятность временно́го сдвига сезона лета пыльцы у J. excelsa на период выпадения обильных осадков и усиления негативного воздействия низкой влажности воздуха на процесс опыления у J. deltoides.
Древовидные можжевельники Juniperus excelsa M. Bieb. (можжевельник высокий) и Juniperus deltoides R.P. Adams (можжевельник дельтовидный) являются аборигенными доминантами древесно-кустарникового яруса реликтовых высокоможжевеловых редколесий Южного берега Крыма (ЮБК) на северной границе ареала, сохранившимися в том числе в природном заповеднике “Мыс Мартьян” (Krainyuk, 2018; Plugatar et al., 2018). Эти виды характеризуются низкой жизнеспособностью семян, слабым естественным возобновлением в Крыму (Grigorov, 1979; Kalafat et al., 2013), включены в Красные книги Российской Федерации (2008), Республики Крым (2015) и Севастополя (2018).
Можжевеловые редколесья занимают наиболее сухие каменистые и малоплодородные участки, в основном на крутых склонах южной, юго-западной и юго-восточной экспозиций, поэтому имеют большое значение для облесения крутых и скалистых горных склонов (Ruguzov et al., 1994).
Основными причинами слабого семенного возобновления у J. excelsa и J. deltoides в условиях Крыма являются большой процент неопыленных семязачатков и близкородственное скрещивание, как следствие изреженности насаждений, а также воздействия неблагоприятных экологических условий в период пыления (Sklonnaya, 1984; Ruguzov et al., 1994; Ruguzova, 2006; Kalafat et al., 2013). Одной из причин “пустосемянности” также является обилие в период лета пыльцы атмосферных осадков и их кислотность (при рН < 6.2) (Grigorov, 1979, 1983; Garcia et al., 2002; Ruguzova, 2006; Ilnitskii et al., 2014). При подкислении секрета опылительной капли пыльцевые зерна не могут освободиться от экзины и не участвуют в половом процессе (Ruguzov et al., 1994). В зависимости от генотипа, места произрастания и гидротермических условий года, доля полноценных семян от свободного опыления в крымских популяциях у J. excelsa колеблется от 1.3 до 26.4% (Sklonnaya, 1984; Ruguzov et al., 1994), у J. deltoides – от 0.4 до 42.7% (Ruguzova, 2006).
Существует мнение, что в настоящее время количество жизнеспособных семян, которые продуцируются ежегодно, достаточно для поддержания крымских популяций можжевельников при неизменном состоянии условий окружающей среды (Ruguzova, 2006). Однако, при увеличении бессистемных вырубок, приводящих к изреженности, изолированности насаждений и сокращению их ареала, загрязнения воздуха кислотными оксидами и изменениях климата природные популяции этих видов могут уменьшаться. Поскольку растения можжевельника имеют длительные сроки жизни, результат низкой (или отсутствующей) регенерации будет заметен только через 25–50 лет (Ruguzova, 2012).
Изучение многолетних изменений средних величин и коэффициентов линейных трендов температурных индексов на территории ЮБК за 1930–2017 гг. показало однозначное увеличение средних и абсолютных температур воздуха, продолжительности и теплообеспеченности вегетационного периода (Plugatar et al., 2015; Korsakova, 2018a). Анализ временного хода средних годовых температур воздуха позволил выявить неоднородный характер потепления: с 1943 по 1965 гг. происходило потепление, которое сменилось небольшим похолоданием в 1967–1987 гг. Начиная с 1988 г. отмечается фаза наиболее интенсивного потепления, которое продолжается до сих пор. За последнюю четверть века скорость потепления возросла в 5–7 раз и составила 0.79°С за десятилетие, средний из абсолютных годовых минимумов увеличился более, чем на 1°С. Самые высокие темпы роста температур отмечены в летний сезон (Plugatar et al., 2015). В годовом ходе осадков прослеживается общая тенденция их увеличения в декабре–феврале и существенное снижение в апреле (Korsakova, 2018a).
При решении вопросов сохранения редких и исчезающих видов ретроспективный анализ динамики сроков цветения позволяет правильно определить функциональное состояние растений в конкретном местообитании в определенное время, оценить их реакцию на изменения термического режима и выявить уязвимость по отношению к изменению климата (Zlobin et al., 2013; Özkan et al., 2015). Изменения фенодат можно рассматривать как адаптацию между генетическими требованиями вида и экологическими условиями местообитания.
Цель работы: выявить особенности антэкологии J. excelsa и J. deltoides на фоне потепления климата и оценить воздействие климатических изменений на жизнеспособность семян древовидных можжевельников в условиях ЮБК.
МАТЕРИАЛЫ И МЕТОДЫ
Исследования проведены в 1974–2017 гг. в природном заповеднике “Мыс Мартьян”, расположенном в приморской зоне южного макросклона Крымских гор у мыса Мартьян (44°30'–44°31' с.ш., 34°15'–34°16' в.д.), которым заканчивается Никитский хребет – отрог Главной гряды, в створе высот от 0 до 240 м над ур. м. (Maslov et al., 2012; Plugatar et al., 2018). По агроклиматическому районированию (Vazhov, 1977) территория заповедника с площадью 1.2 км2 входит в Западный южнобережный субтропический район. Климат заповедника относится к субтропическому варианту средиземноморского типа с умеренно жарким засушливым летом, преобладанием осенне-зимних осадков и мягкой зимой с частыми оттепелями. Как и на всем Южном берегу, в заповеднике отсутствует период с устойчивыми среднесуточными температурами ниже 0°С. Средняя годовая температура воздуха между верхней (северной) и нижней (приморской) границами заповедника изменяется от 12.0 до 13.6°С. Самыми холодными месяцами года являются январь и февраль, средние температуры которых составляют от 3°С на северной границе до 4°С в приморской зоне. Самые высокие среднемесячные температуры воздуха наблюдаются в июле–августе (от 23 до 25°С). Среднегодовое количество осадков между верхней и нижней границами заповедника уменьшается от 600 до 490 мм (Plugatar et al., 2015, 2018).
В заповеднике J. excelsa является одним из основных лесообразующих видов, образует чистые можжевеловые и смешанные дубово-можжевеловые сообщества, относящиеся к классу Quercetea pubescentis Doing-Kraft ex Scamoni et Passarge 1959 (Mucina et al., 2016), а J. deltoides является характерным компонентом подлеска высокоможжевеловых лесов.
В ходе мониторинговых исследований изменений продолжительности и сезонной ритмики пыления J. excelsa и J. deltoides регистрировали начало, массовое наступление и конец фенофазы. Фенологические наблюдения проводили в течение всего года с интервалом в семь дней, а в период пыления – с интервалом в два-три дня, на постоянных объектах и маршруте, проходящем в центральной части заповедника, по методике И.Н. Бейдемана (1974). Началом фенологической фазы считался тот день, когда она была зафиксирована хотя бы на одном растении, окончанием – день, когда она завершилась у всех растений. Дата массового наступления фазы регистрировалась тогда, когда в нее вступило около 50% органов не менее чем у 50% особей. Наблюдения проводили за 20 модельными особями J. excelsa и 25 особями J. deltoides (10 женских и 15 мужских).
В работе использованы данные агрометеорологической станции “Никитский сад” (44°31' с.ш., 34°15' в.д., высота 208 м над ур. м.).
Статистическая обработка данных проведена с помощью программ MS Excel 2010 и Statistica10.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
На ЮБК вылет пыльцы у J. excelsa в зависимости от гидротермических условий года начинается в конце января – начале апреля, у J. deltoides – в начале апреля – середине мая. Ко времени вылета пыльцевых зерен из микроспорангия семязачаток у J. excelsa и J. deltoides хорошо развит, нуцеллус дифференцирован на базальную и апикальную зоны, интегумент образует довольно длинный микропилярный канал (Ruguzov et al., 1994; Ruguzova, 2006). Массовое пыление у J. excelsa наступает в среднем через 8–13 дней после начала фенофазы, у J. deltoides – через 5–6 дней и характеризуется значительной межгодовой вариабельностью (рис. 1).
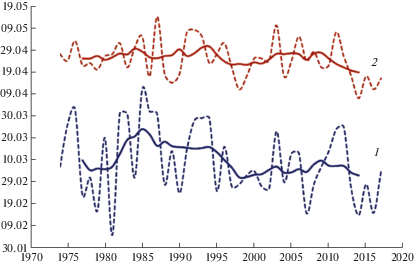
Рис. 1.
Межгодовая динамика сроков пыления Juniperus excelsa (1) и J. deltoides (2) в природном заповеднике “Мыс Мартьян” (сглаженная кривая (сплошная линия) получена 7-летним скользящим осреднением). По оси абсцисс – годы, по оси ординат – дата начала массового пыления.
Fig. 1. Interannual changes of the pollination terms in Juniperus excelsa (1) and J. deltoides (2) in the “Cape Martyan” Nature Reserve (smoothed curve (solid line) is obtained by 7-year moving averaging). X-axis – years, Y-axis – mass pollination start date.
Пыльца, эффективная для опыления, разносится в радиусе до 20 м, но основная ее масса оседает в радиусе 5 м. У J. excelsa только 11% семязачатков опыляются пыльцой других деревьев, а около 90% – воспринимают пыльцу этого же дерева (Sklonnaya, 1984). Это объясняется изреженностью популяций данного вида в Крыму и высокой (выше 70%) относительной влажностью воздуха в период пыления (табл. 1). В ненастную погоду радиус разлета пыльцы резко сокращается. Моросящий дождь удаляет из воздуха 99% пыльцы (Ruguzova, 2006). При изреженности насаждений и неблагоприятных погодных условиях большинство семязачатков остаются неопыленными.
Таблица 1.
Экологические условия и продолжительность лета пыльцы Juniperus excelsa в заповеднике “Мыс Мартьян” Table 1. Environmental conditions and the duration of the Juniperus excelsa pollen release in the “Cape Martyan” Nature Reserve
| Период, гг. Period, years |
Температура воздуха, °С Air temperature, °С |
Сумма
осадков, мм Precipitation, mm |
Относительная влажность воздуха, % Air humidity, % |
Продолжительность пыления, дни Duration of pollen release, days | ||
|---|---|---|---|---|---|---|
| Средняя Average | Средняя максимальная Average maximum | Средняя минимальная Average minimum | ||||
| 1974–2017 | 6.6 ± 2.5 | 10.3 ± 2.9 | 3.8 ± 2.2 | 40 ± 38 | 71 ± 6 | 26 ± 14 |
| 1981–1990 | 7.6 ± 3.0 | 11.2 ± 3.6 | 4.7 ± 2.4 | 34 ± 48 | 72 ± 7 | 22 ± 14 |
| 1991–2000 | 6.5 ± 2.3 | 10.0 ± 2.7 | 3.5 ± 1.9 | 29 ± 31 | 70 ± 6 | 24 ± 14 |
| 2001–2010 | 5.2 ± 1.3 | 8.9 ± 1.7 | 2.3 ± 1.1 | 70 ± 33 | 71 ± 4 | 36 ± 9 |
| 2011–2017 | 7.2 ± 1.2 | 10.9 ± 1.5 | 4.5 ± 1.2 | 37 ± 30 | 75 ± 5 | 24 ± 10 |
Вылет пыльцы происходит не одновременно. В пределах особи у J. excelsa он продолжается в среднем 3–7 дней, а в пределах популяции – 3–5 недель (табл. 1). При выпадении обильных осадков и снижении температур, длительность периода пыления может увеличиваться до 74 дней (1981 г.), при отсутствии осадков и высоких температурах (выше 10°С) – сокращаться до 6–11 дней. Пыльцевой режим в природных популяциях J. excelsa характеризуется продолжительным периодом массового пыления (в среднем 5–13 дней) и может достигать 25 суток (2007 г.).
Пыление единичных деревьев J. deltoides длится в среднем 3–4 дня, а в пределах ценопопуляции – 2–3 недели (табл. 2) с колебаниями от 8 до 37 дней. По сравнению с J. excelsa, продолжительность лета пыльцы J. deltoides в популяции на 10 дней меньше. Длительность массового пыления изменяется в диапазоне от 1 до 12 суток и в среднем составляет 6 дней.
Таблица 2.
Экологические условия и продолжительность лета пыльцы Juniperus deltoides в заповеднике “Мыс Мартьян” Table 2. Environmental conditions and the duration of the Juniperus deltoides pollen release in the “Cape Martyan” Nature Reserve
| Период, гг. Period, years |
Температура воздуха, °С Air temperature, °С |
Сумма осадков, мм Precipita-tion, mm |
Относительная влажность
воздуха, % Air humidity, % |
Продолжительность
пыления, дни Duration of pollen release, days |
||
|---|---|---|---|---|---|---|
| Средняя Average | Средняя максимальная Average maximum | Средняя минимальная Average minimum | ||||
| 1974–2017 | 12.8 ± 2.4 | 16.5 ± 2.6 | 9.5 ± 2.1 | 15 ± 16 | 65 ± 9 | 16 ± 7 |
| 1981–1990 | 12.7 ± 2.6 | 16.3 ± 2.9 | 9.4 ± 2.3 | 16 ± 16 | 64 ± 6 | 15 ± 3 |
| 1991–2000 | 14.1 ± 1.6 | 17.9 ± 1.8 | 10.7 ± 1.3 | 7 ± 9 | 64 ± 11 | 12 ± 6 |
| 2001–2010 | 12.6 ± 2.8 | 16.3 ± 2.7 | 9.4 ± 2.6 | 13 ± 18 | 63 ± 11 | 15 ± 5 |
| 2011–2017 | 12.9 ± 2.9 | 16.9 ± 3.3 | 9.5 ± 2.6 | 14 ± 10 | 66 ± 8 | 21 ± 8 |
Длительность пыления J. excelsa в 2001–2010 гг. превышала средние многолетние показатели почти в полтора раза (табл. 1). В этот временной интервал на ЮБК наблюдался рост температур воздуха в ноябре и в феврале (Korsakova, 2018a), что способствовало более раннему началу пыления. Лет пыльцы приобретал пролонгированный характер, температуры воздуха (средние, минимальные и максимальные) были на 1.4–1.5°С ниже, а количество осадков на 75% выше средних многолетних значений. Анализируя многолетнюю динамику длительности пыления и погодных условий в этот период, можно заключить, что период с 2001 по 2010 г. не был благоприятным для образования полноценных семян J. excelsa на мысе Мартьян.
В последние годы отмечена тенденция роста продолжительности пыления J. deltoides на фоне небольшого снижения температур. При этом сумма осадков не превышала средние многолетние показатели, а средние максимальные температуры воздуха в период пыления были выше 16°С (табл. 2). Это указывает на отсутствие ухудшений гидротермических условий для опыления в исследованный временной интервал, за исключением отдельных лет в период с 2001 по 2017 г., когда относительная влажность воздуха опускалась ниже критических значений для формирования опылительной капли. Данная тенденция позволяет выдвинуть предположение об усилении лимитирующей роли влажности воздуха в процессе опыления для J. deltoides при дальнейшем росте температур во время лета пыльцы.
Расчеты, проведенные по результатам многолетних исследований, показали, что прохождение фазы пыления J. excelsa ускоряется в среднем на два дня, J. deltoides – на один день с ростом температуры на 1°С, и на столько же затягивается при ее снижении. Пролонгации пыления на один день способствует выпадение от 3 до 5 мм осадков. Наибольшее влияние на продолжительность лета пыльцы J. excelsa и J. deltoides оказывает сумма осадков, выпавших в этот период, доля влияния которых, соответственно, составляет 59 и 24% (p < 0.001). Влияние температур на длительность пыления проявляется слабее (соответственно R2 = 0.12–0.13 и R2 = 0.13–0.19; p < 0.05). Влияние относительной влажности воздуха в связи с ее невысокой изменчивостью менее значимо (R2 = 0.03–0.10; p > 0.05).
Сопряженный анализ межгодовой изменчивости сроков лета пыльцы у можжевельников и средних месячных температур воздуха показал наличие статистически значимой зависимости начала пыления J. excelsa и J. deltoides от средних температур февраля, марта и апреля (табл. 3). Начало пыления J. excelsa наиболее связано с температурами февраля, а J. deltoides – марта (R2 = 0.51). Повышение температуры февраля на 1°С приводит к сдвигу на более ранние сроки (на 7 дней) начала пыления J. excelsa. При повышении температуры марта на 1°С вылет пыльцы у J. deltoides начинается раньше, в среднем, на 4 дня.
Таблица 3.
Результаты регрессионного анализа зависимости даты начала лета пыльцы (Y) Juniperus excelsa и J. deltoides от средних месячных температур воздуха (x, °С) Table 3. Results of regression analysis of the dependence of the pollen release start dates (Y) in Juniperus excelsa and J. deltoides on average monthly air temperatures (x, °С)
| Месяц Month |
Виды растений Plant species |
Уравнение линейной регрессии Equation of linear regression |
Коэффициент детерминации, R2 Determination coefficient, R2 |
|---|---|---|---|
| Февраль February |
J. excelsa | Y = 82.26 – 7.01х | 0.51* |
| J. deltoides | Y = 118.98 – 2.75х | 0.34* | |
| Март March |
J. excelsa | Y = 93.82 – 6.13х | 0.28* |
| J. deltoides | Y = 132.44 – 3.99х | 0.51* | |
| Апрель April |
J. excelsa | Y = 56.77 + 0.26х | 0.00 |
| J. deltoides | Y = 142.53 – 3.13х | 0.26* |
Феноклиматические показатели (среднемноголетняя дата, изменчивость и степень разброса дат) являются видовыми характеристиками. О степени изменчивости фенодат и устойчивости растений в меняющихся погодно-климатических условиях с достаточной достоверностью позволяет судить показатель среднего квадратического отклонения (σ), адекватно оценивающий вариабельность (пластичность) сезонных процессов и отражающий степень консерватизма растений (Shumik et al., 2016). Фенодаты начала пыления J. excelsa (σ = 15.2–19.5 дней) характеризуются значительной лабильностью, что согласно фенологическому критерию свидетельствует о высокой адаптации растений к условиям произрастания. Для J. deltoides (σ = 8.9–9.5 дней), у которого данная фаза репродуктивного цикла наступает во второй половине весны, этот показатель ниже (табл. 4). Стабильность фенофазы пыления также связана с вариабельностью температур зимне-весенних месяцев. Меньший коэффициент вариации температур месяца обусловливает бо́льшую стабильность фаз, которые проходят в эти месяцы. В связи с тем, что февраль и март на ЮБК характеризуются высокими колебаниями температур (коэффициент вариации в феврале составляет 53–61%, а в марте – 27–33%), сроки пыления J. excelsa менее стабильны. Коэффициент вариации температур воздуха в апреле значительно ниже (12–16%), что способствует меньшей разбалансировке дат начала вылета пыльцы J. deltoides.
Таблица 4.
Среднемесячная температура воздуха (°С), даты начала пыления Juniperus excelsa и J. deltoides за 1974–2017 гг. и наблюдаемые тенденции Table 4. Average monthly temperature (°C), pollination start dates in Juniperus excelsa and J. deltoides during 1974–2017, and observed trends
| Показатели Indicators |
Период, годы/Period, years | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1974–2017 | 1981–2017 | 2001–2017 | |||||||
| Среднее Average | σ | b | Среднее Average | σ | b | Среднее Average | σ | b | |
| Февраль, °C/February, °C | 3.3 | 2.0 | 0.4 | 3.3 | 1.9 | 0.6* | 3.8 | 2.0 | 0.8 |
| Март, °C/March, °C | 5.2 | 1.7 | 0.4 | 5.6 | 1.7 | 0.7* | 6.2 | 1.7 | 1.0 |
| Апрель, °C/April, °C | 10.3 | 1.6 | 0.2 | 10.4 | 1.5 | 0.3 | 10.6 | 1.3 | 0.8 |
| J. excelsa | 28.II | 19.3 | –4.5* | 28.II | 19.5 | –7.0* | 20.II | 15.2 | 3.6 |
| J. deltoides | 20.IV | 9.3 | –0.8 | 21.IV | 9.5 | –3.9* | 18.IV | 8.9 | –10.0* |
Коэффициенты линейного тренда (табл. 4) с 1981 по 2017 г. указывают на статистически значимую тенденцию смещения на более ранние сроки начала пыления J. excelsa со средней скоростью 7 дней за 10 лет, J. deltoides – 4 дня. После 2000 г. отмечено усиление темпов смещения для J. deltoides. В первых десятилетиях XXI века начало пыления J. excelsa отмечается в среднем на 8 дней раньше среднемноголетних показателей (1974–2017 гг.), J. deltoides – на 2 дня.
Выявленные особенности фенологических реакций древовидных можжевельников на изменения температур зимне-весенних месяцев дают основания для прогнозных оценок их поведения при дальнейшем потеплении климата. Учитывая прогнозируемый для ЮБК рост температуры воздуха зимних и весенних месяцев (Efimov et al, 2015; Korsakova, 2018b), в ближайшие 20–30 лет существует высокая вероятность временно́го сдвига вылета пыльцы у J. excelsa на сезон выпадения обильных осадков, что ухудшит условия для опыления и естественного возобновления. Вероятность ухудшений гидротермических условий в период пыления для J. deltoides в ближайшие десятилетия невысокая.
Исследованные виды способны возобновляться только семенным путем. J. excelsa представлен однодомными, а J. deltoides – двудомными особями. По данным исследователей, изучавших закономерности формирования семян у древовидных можжевельников (Sklonnaya, 1984; Ruguzov et al., 1994; Ruguzova, 2006), в Крыму интервал от закладки репродуктивных органов до опыления у J. excelsa составляет 7–8 месяцев, у J. deltoides – 10 месяцев. Между опылением и оплодотворением у J. excelsa проходит 3–4 месяца, у J. deltoides – 15 месяцев. Репродуктивный цикл (от закладки мужских и женских шишек до образования семян с дифференцированным зародышем) у J. excelsa длится 27 месяцев, у J. deltoides – 40.
Наиболее чувствительными к изменению экологических условий из всех фаз полового процесса у древовидных можжевельников, произрастающих в Крыму, являются процессы мейоза и опыления (Ruguzov et al., 1994). А.И. Ругузовой было выявлено, что асинхронность протекания мейоза в пределах популяции, дерева, микростробила и микроспорангия обеспечивает при любых погодных условиях образование достаточного количества жизнеспособной пыльцы для опыления семязачатков и этот фактор не лимитирует формирование полноценных семян у J. deltoides (Rugusova, 2006). Чаще всего цикл репродукции прерывается на стадии опыления, что обусловлено как местом произрастания, так и погодными условиями в период опыления. Экспериментальные и гистологические исследования показали, что для появления опылительной капли у J. excelsa и J. deltoides необходима не только физиологическая готовность нуцеллуса к процессу секреции, но и определенные гидротермические условия (Ruguzov et al., 1992). Опылительная капля формируется у J. excelsa при максимальной температуре воздуха не ниже 10°С (среднесуточной – не ниже 6.5°С) и относительной влажности не менее 60%, у J. deltoides – не ниже 16°С (среднесуточной – не ниже 12°С) и относительной влажности не менее 50% (Sklonnaya, 1984; Ruguzov et al., 1994; Ruguzova, 2006). По некоторым данным, при отсутствии опыления, в благоприятных гидротермических условиях опылительная капля может функционировать от 5 (Surso, Selivanova, 2016) до 14 (Ruguzova, 2006) суток.
В связи с этим, нами была проанализирована динамика основных гидротермических факторов среды (средних суточных температур и относительной влажности воздуха, суммы осадков), лимитирующих образование жизнеспособных семян в период пыления J. excelsa и J. deltoides на ЮБК (рис. 2–3).
Рис. 2.
Многолетняя динамика гидротермических условий в период массового пыления J. excelsa в природном заповеднике “Мыс Мартьян”. По оси абсцисс – годы, по оси ординат – факторы среды: а – температура воздуха, °С; б – относительная влажность воздуха, %; в – количество осадков, мм. Пунктирными линиями и стрелками выделены пороговые значения и зоны экологической толерантности: к температуре (а) и относительной влажности (б) воздуха для функционирования опылительной капли; количеству осадков (в), лимитирующих лет пыльцы.
Fig. 2. Long-term changes of hydrothermal environmental factors during a full pollen release season of J. excelsa in the “Cape Martyan” Nature Reserve. X-axis – years, Y-axis – environmental factors: a – air temperature, °C; b – air humidity, %; c – precipitation, mm. Dashed lines and arrows indicate thresholds and zones of environmental tolerance: to the air temperature (a) and relative humidity (b) for the functioning of the pollination drop; to the precipitation (c) limiting the pollen release.
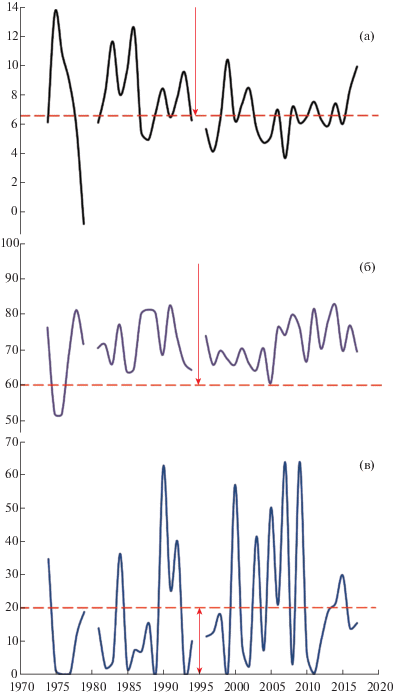
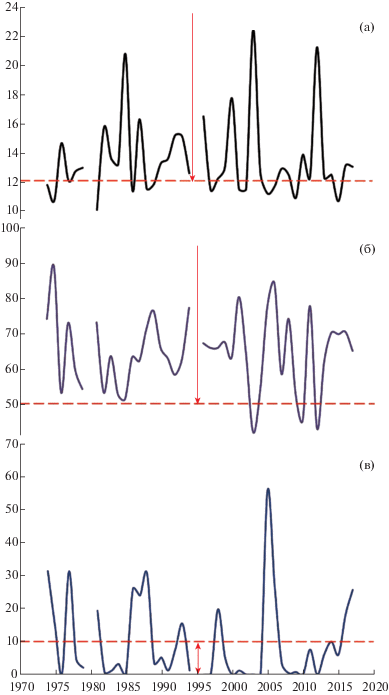
Рис. 3.
Многолетняя динамика гидротермических условий в период массового пыления J. deltoides в природном заповеднике “Мыс Мартьян”. По оси абсцисс – годы, по оси ординат – факторы среды: а – температура воздуха, °С, б – относительная влажность воздуха, %, в – количество осадков, мм. Пунктирными линиями и стрелками выделены пороговые значения и зоны экологической толерантности: к температуре (а) и относительной влажности (б) воздуха для функционирования опылительной капли; количеству осадков (в), лимитирующих лет пыльцы.
Fig. 3. Long-term changes of hydrothermal environmental factors during a full pollen release season of J. deltoides in the “Cape Martyan” Nature Reserve. X-axis – years, Y-axis – environmental factors: a – air temperature, °C; b – air humidity, %; c – precipitation, mm. Dashed lines and arrows indicate thresholds and zones of environmental tolerance: to the air temperature (a) and relative humidity (b) for the functioning of the pollination drop; to the precipitation (c) limiting the pollen release.
После 1977 г. в период пыления J. excelsa (рис. 2) значения относительной влажности воздуха находились в зоне оптимума. Основными экологическими факторами, лимитирующими процесс опыления для данного вида в период лета пыльцы, являлись среднесуточная температура воздуха и атмосферные осадки.
Анализ данных за период с 1974 по 2017 г. показал, что неблагоприятные температурные условия для формирования опылительной капли у можжевельника высокого, наблюдались в 38% лет (или три-четыре года из 10), обилие осадков в период пыления – в 26% лет (два-три года из 10). Наиболее неблагоприятные условия для опыления при сочетании этих двух факторов в период пыления отмечены в 17% лет, из них 10% лет (около 64%) приходились на 2001–2010 гг.
В период пыления J. deltoides (рис. 3) неблагоприятные для формирования опылительной капли температурные условия, а также обилие осадков наблюдались в среднем один раз в три года (в 31% лет). В XXI столетии, на фоне общего уменьшения количества осадков в апреле, во время пыления можжевельника дельтовидного на ЮБК выявлено снижение частоты их выпадения, что благоприятно для лета пыльцы. В качестве негативного фактора в период пыления, в 2001–2017 гг., при повышении температур отмечено снижение влажности воздуха ниже критических значений для формирования опылительной капли.
ЗАКЛЮЧЕНИЕ
Амплитуда экотопической изменчивости начала лета пыльцы J. excelsa и J. deltoides зависит от вариабельности температур зимне-весенних месяцев. Меньший коэффициент их вариации обусловливает бо́льшую стабильность фенофаз, которые проходят в эти месяцы. Основное влияние на продолжительность пыления оказывает количество выпавших в этот период осадков.
Основными факторами, лимитирующими функционирование опылительной капли у J. excelsa на ЮБК, являются температура воздуха и обилие осадков, у J. deltoides – температура, влажность воздуха и обилие осадков. Неблагоприятные гидротермические условия в период лета пыльцы складываются для данных видов можжевельников, в среднем, в трех-четырех годах из 10. Период с 2001 по 2010 г. не был благоприятным для образования полноценных семян у J. excelsa. При дальнейшем росте температур существует вероятность сдвига сезона лета пыльцы у J. excelsa на период выпадения обильных осадков, а также усиления негативного воздействия низкой влажности воздуха на процесс опыления у J. deltoides.
Список литературы
[Beideman] Бейдеман И.Н. 1974. Методика изучения фенологии растений и растительных сообществ. Новосибирск. 156 с.
Efimov V.V., Volodin E.M., Anisimov A.E. 2015. Modeling of the Black Sea region climate changes in the XXI century. – Phys. Oceanogr. 2: 3–13. https://doi.org/10.22449/1573-160X-2015-2-3-13
Garcia D., Zamora R., Gomez J.M., Hodar J.A. 2002. Annual variability in reproduction of Juniperus communis L. in a Mediterranean Mountain: relationship to seed prediation and weather. – Ecoscience. 9 (2): 251–255.
[Grigorov] Григоров А.Н. 1979. Семеношение и качество семян можжевельника высокого в Крыму. – Бюлл. Гос. Никитского бот. сада. 3 (40): 10–13.
[Grigorov] Григоров А.Н. 1983. Можжевельник высокий (Juniperus excelsa Bieb.) в Крыму (биоэкологические особенности, возобновление и охрана): Автореф. дис. … канд. биол. наук. Киев. 22 с.
[Ilnitskii et al.] Ильницкий О.А., Плугатарь Ю.В., Корсакова С.П. 2014. Проблема “кислотных осадков” на Южном берегу Крыма и их влияние на растительный мир. – Бюлл. Гос. Никитского бот. сада. 111: 7–21.
[Kalafat et al.] Калафат Л.А., Николаева А.В., Егорова А.В. 2013. Семенная продуктивность видов рода Juniperus L. в Крыму. – Промышленная ботаника. 13: 163–168.
[Korsakova] Корсакова С.П. 2018a. Анализ временной изменчивости характеристик термического режима на Южном берегу Крыма. – Бюлл. Гос. Никитского бот. сада. 128: 100–111. https://doi.org/10.25684/NBG.boolt.128.2018.13
[Korsakova] Корсакова С.П. 2018b. Оценка будущих изменений климата на Южном берегу Крыма. – Экосистемы. 15 (45): 151–165.
[Krainyuk] Крайнюк Е.С. 2018. Ботанические исследования в заповеднике “Мыс Мартьян”: 45-летние итоги. – Научн. зап. природ. запов. “Мыс Мартьян”. 9: 19–29. https://doi.org/10.25684/NBG.scnote.009.2018.02
[Krasnaya…] Красная книга города Севастополя. 2018. Калининград; Севастополь. 432 с.
[Krasnaya…] Красная книга Российской Федерации (растения и грибы). 2008. М. 855 с.
[Maslov et al.] Маслов И.И., Крайнюк Е.С., Саркина И.С., Хаустов А.А., Сергеенко А.Л. 2012. Сохранение биоты в природном заповеднике “Мыс Мартьян”. – Тр. Никитского бот. сада. 134: 15–32.
Mucina L., Bültmann H., Dierßen K., Theurillat J.-P., Raus T., Čarni A., Šumberová K., Willner W., Dengler J., García R.G., Chytrý M., Hájek M., Di Pietro R., Iakushenko D., Pallas J., Daniëls F.J.A., Bergmeier E., Santos Guerra A., Ermakov N., Valachovič M., Schaminée J.H.J., Lysenko T., Didukh Ya.P., Pignatti S., Rodwell J.S., Capelo J., Weber H.E., Solomeshch A., Dimopoulos P., Aguiar C., Hennekens S.M., Tichý L. 2016. Vegetation of Europe: hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. – Appl. Veget. Sci. 19 (51): 3–264. https://doi.org/10.1111/avsc.12257
Özkan K., Sentürk Ö., Mert A., Negiz M.G. 2015. Modeling and mapping potential distribution of Crimean juniper (Juniperus excelsa Bieb.) using correlative approaches. – J. Environ. Biol. 36 (1): 9‒15.
[Plugatar et al.] Плугатарь Ю.В., Багрикова Н.А., Белич Т.В., Костин С.Ю., Крайнюк Е.С., Маслов И.И., Садогурский С.Е., Садогурская С.А., Саркина И.С. 2018. Природный заповедник “Мыс Мартьян”. Cимферополь. 104 с.
[Plugatar et al.] Плугатарь Ю.В., Корсакова С.П., Ильницкий О.А. 2015. Экологический мониторинг Южного берега Крыма. Симферополь. 164 с.
[Red…] Красная книга Республики Крым: растения, водоросли и грибы. 2015. Симферополь. 480 с.
[Ruguzov et al.] Ругузов И.А., Склонная Л.У., Костина В.П. 1994. Цитоэмбриологические основы сохранения редких видов рода Juniperus L. в Крыму. – Укр. бот. журн. 51 (2/3): 211–217.
[Ruguzov et al.] Ругузов И.А., Склонная Л.У., Чеботарь А.А. 1992. Об опылительной капле у хвойных. – Бот. журн. 77 (12): 40–52.
[Ruguzova] Ругузова А.И. 2006. Биологические особенности можжевельника красного (Juniperus oxycedrus L.) в Крыму в связи с его охраной: Дис. … канд. биол. наук. Ялта. 163 с.
Ruguzova A.I. 2012. Seed productivity Juniperus hemispaerica C. Presl. Juniperus sabina L. and their preservation in the Crimea. – Науковий вісник Національного університету біоресурсів і природокористування України. Сер.: Лісівництво та декоративне садівництво. 171 (1): 294–299.
[Shumik et al.] Шумик Н.И., Заименко Н.В., Остапюк В.М. 2016. Сезонная ритмика интродуцированных растений как критерий их устойчивости и адаптации. – Бюлл. Бот. сада-института. 15: 96–98.
[Sklonnaya] Склонная Л.У. 1984. Процессы семенообразования и качество семян у Juniperus excelsa Bieb. и Cedrus deodara (D. Don) G. Don в Крыму: Дис. … канд. биол. Наук. Ялта. 142 с.
[Surso, Selivanova] Сурсо М.В., Селиванова Н.В. 2016. Опыление у можжевельника обыкновенного (Juniperus communis L.): механизм вовлечения пыльцы и влияние мужского гаметофита на развитие семязачатков и “шишкоягод”. – Лесной журн. 4: 40–53. https://doi.org/10.17238/issn0536-1036.2016.4.40
[Vazhov] Важов В.И. 1977. Агроклиматическое районирование Крыма. – Тр. Гос. Никитского бот. сада. 71: 92–120.
[Zlobin et al.] Злобин Ю.А., Скляр В.Г., Клименко А.А. 2013. Популяции редких видов растений: теоретические основы и методика изучения. Сумы. 439 с.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал