

Ботанический журнал, 2019, T. 104, № 2, стр. 226-253
ЕЛОВЫЕ ЛЕСА СЕВЕРО-ЗАПАДНОГО ПРИЛАДОЖЬЯ
М. А. Макарова *
Ботанический институт им. В.Л. Комарова РАН
197376 Санкт-Петербург, ул. проф. Попова д. 2, Россия
* E-mail: MMakarova@binran.ru
Поступила в редакцию 17.10.2018
После доработки 09.12.2018
Принята к публикации 11.12.2018
Аннотация
В статье приводится характеристика еловых лесов Северо-Западного Приладожья (Балтийский кристаллический щит). На основе эколого-фитоценотической классификации автором было выделено 3 группы ассоциаций (зеленомошные, травяные и сфагновые), 6 ассоциаций и 6 вариантов ассоциаций еловых лесов. Преобладают ельники чернично- и кислично-зеленомошные. Ельники с неморальными видами встречаются на участках неосвоенных озерных террас и по подножиям склонов сельг. На основе крупномасштабной карты растительности 2004 г. и данных дистанционного зондирования 2017 г. показаны изменения распространения еловых массивов.
Еловые леса – формация Северо-Запада Европейской России, начавшая свое развитие еще до наступления суббореального послеледникового периода (Nitsenko, 1960). Пыльца Picea зарегистрирована около 16500 лет назад на юго-западе Валдайской возвышенности и в районе Угличской возвышенности (Kozharinov et al., 2010). В Псковской, Ленинградской и Вологодской областях максимум развития ели приходится на период 5500 лет назад, в Карелии – 4500 (Elina, 1981). В последние 500 лет ареал ели уменьшился: на северной границе происходит заболачивание и ель отступает к югу, в то же время вдоль южной границы наблюдается усыхание ели (Kozharinov et al., 2010). В Ленинградской области в середине 1950-х гг. площадь еловых и мелколиственно-еловых лесов составляла около трети всей лесной территории (Nitsenko, 1960), к 2010 г. площадь ельников немного снизилась и составляла 27% (1236.3 тыс. га) лесного фонда Ленинградской обл. (Obzor, 2010).
Климатические изменения, стихийные явления (пожары, нашествия вредителей, ветровалы), вырубки ведут к изменению пространственной и внутриценотической структуры лесов. В связи с чем геоботанические исследования еловых лесов представляются важными и необходимыми.
РАЙОН ИССЛЕДОВАНИЙ
Полевые исследования проходили в 1997–1998 и 2003–2004 годы в Приозерском р-не Ленинградской обл. вблизи пос. Кузнечное и дер. Березово, в окрестностях Приладожской учебно-научной базы СПбГУ и в северо-западной части проектируемой ООПТ “Кузнечное”. Ключевой участок располагается в сельгово-ложбинном ландшафтном районе Северо-Западное Приладожье, на южной окраине Балтийского кристаллического щита (Isachenko, Reznikov, 2003). Согласно геоботаническому районированию территория относится к Восточно-европейской провинции Евразиатской хвойнолесной (таежной) области, южнотаежной зоне, геоботаническому району шхерного побережья Ладожского озера (Abramova, Kozlova, 1957). Характерен грядово-ложбинный рельеф, представленный сочетанием вытянутых гранитных гряд-сельг и межсельговых понижений. Вершины и привершинные участки гранитных сельг заняты сосняками лишайниковыми и кустарничково-зеленомошными. На склонах сельг произрастают еловые, мелколиственные, сосново-мелколиственные чернично-зеленомошные, чернично-травяные и травяные леса, местами неморальнотравные. К узким межсельговым котловинам приурочены травяно-сфагновые болота. На глинистых озерных террасах отмечаются ельники кисличные с неморальным разнотравьем, папоротниковые, осиновые и сероольхово-березовые травяно-кисличные, кислично-неморальнотравные и снытевые леса (Makarova, 2008). В пределах ключевого участка плакоры отсутствуют. Дренированные озерные глинистые террасы наиболее близки по своим ландшафтно-экологическим условиям к плакорам, но большинство из них было освоено в прошлом под сельскохозяйственные угодья, в настоящее время часть этих участков находится под сенокосными лугами, часть заброшена и активно зарастает мелколиственными породами с последующим восстановлением до мелколиственно-еловых и еловых лесов (Makarova, 2009).
МАТЕРИАЛЫ И МЕТОДЫ
В 2004 г. автором был изучен ключевой участок площадью 35 км2, на который составлена крупномасштабная карта растительности М 1:25000 (Makarova, 2010). При обследовании и картографировании растительности ключевого участка было сделано около 500 геоботанических описаний, из них 90 описаний еловых лесов. Закладывались пробные площади размером 400 м2, в них проводилось описание растительности по ярусам, включающие характеристики состава древостоя, сомкнутости, высоты и диаметра деревьев, подроста, подлеска, состава и проективного покрытия видов травяно-кустарничкового и мохово-лишайникового ярусов. Критерием для отнесения зеленомошных и травяных лесов к ельнику служила формула древостоя, в которой доля ели принималась от 5 и выше (табл. 1), в сфагновых – от 4 и выше (табл. 2). Учитывались также выявленные антропогенные воздействия; для ельников это были главным образом новые или старые выборочные рубки. Описания еловых лесов с проведенными выборочными рубками (с сомкнутостью древостоя менее 0.3) были использованы при разработке легенды к карте, так как в пределах ключевого участка они встречаются достаточно часто.
Таблица 1.
Геоботанические описания еловых зеленомошных и еловых травяных лесов (Северо-Западное Приладожье). Table 1. Releves of spruce green-moss and spruce herbaceous forests (North-Western Ladoga region)
| Группы ассоциаций Groups of associations |
Piceeta hylocomiosa | Piceeta herbosa | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ассоциации/Associations | 1. Piceetum myrtilloso- hylocomiosum | 2. Piceetum myrtilloso-oxalidoso-hylocomiosum | 3. Piceetum oxalidoso- hylocomiosum | 4. | |||||||||||||||||||||||
| Варианты/Variants | 2a | 3a | 3b | 4a | 4b | ||||||||||||||||||||||
| № описания/№ releve | 1 | 2' | 3' | 4 | 5 | 6 | 7 | 8 | 9 | 10' | 11 | 12 | 13 | 14' | 15' | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 |
| Год Year |
2004 | 1991 | 1996 | 1997 | 1997 | 1997 | 2004 | 1997 | 1997 | 1991 | 2003 | 2004 | 2004 | 1991 | 2001 | 1997 | 2004 | 2004 | 2004 | 1997 | 2004 | 1997 | 1997 | 1997 | 1997 | 2004 | 2004 |
| Сомкнутость крон: Density of crown: |
0.3 | 0.6 | 0.5 | 0.4 | 0.4 | 0.4 | 0.6 | 0.4 | 0.4 | 0.6 | 0.3 | 0.6 | 0.6 | 0.6 | 0.6 | 0.4 | 0.6 | 0.3 | 0.7 | 0.4 | 0.4 | 0.5 | 0.4 | 0.5 | 0.3 | 0.5 | 0.5 |
| Древостой 1-го полога Stand of the 1st canopy |
. | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Picea abies | 6 | 8 | 9 | 7 | 8 | 7 | 10 | 6 | 6 | 9 | 7 | 10 | 10 | 5 | 6 | 6 | 5 | 9 | 6 | 10 | 8 | 8 | 7 | 6 | 6 | 9 | 10 |
| Высота, м Height, m |
22–30 | 22–27 | 20–27 | 18–22 | 20 | 20 | 25 | 25 | 20 | 24–30 | 20 | 23–26 | 29 | 20–27 | 20–29 | 22 | 28–30 | 28 | 22 | 20 | 25–30 | 20 | 20 | 21 | 24 | 20–22 | 24–30 |
| Диаметр, см Diameter, cm |
15–40 | 18–31 | 20–32 | . | . | . | 30–50 | 20–45 | . | 34– 63 | 15–25 | 30–50 | 20–50 | 15–37 | 20–53 | . | 30 | 10–70 | 20–25 | . | 20–40 | . | . | . | 50 | 10–50 | 20, 50 |
| Pinus sylvestris | 3 | + | + | 1 | 1 | 1 | + | . | 1 | 1 | 3 | + | . | 1 | 2 | . | 4 | 1 | 2 | . | 2 | . | 2 | 1 | . | . | . |
| Высота, м Height, m |
20–28 | 19–25 | . | 16 | 18–20 | 19–20 | 24 | . | 18–20 | 20 | 20 | 25 | 29 | 22–25 | 21–25 | . | 25–28 | 26 | 25–26 | . | 20–23 | . | 20 | 21 | . | . | . |
| Диаметр, см Diameter, cm |
20–30 | 16–22 | . | . | . | . | 35 | . | . | 41 | . | 35 | 20–50 | 22–41 | 26–48 | . | 30–40 | 30–50 | 30–60 | . | 20 | . | . | . | . | . | . |
| Betula pendula+ B.pubescens | 1 | 1 | 1 | 2 | 1 | 1 | . | 4 | 2 | . | + | + | . | 2 | 2 | 2 | + | . | 2 | . | + | 1 | + | 3 | 3 | + | + |
| Высота, м Height, m |
20 | 23–28 | 18–24 | 20–22 | 19 | 19 | . | 22 | 19 | . | 14 | 22 | . | 17–25 | 19–27 | 18 | . | . | 24 | . | 20 | 19–20 | . | . | 20 | 18 | . |
| Диаметр, см Diameter, cm |
20 | 16–22 | 16–30 | . | . | . | . | . | . | . | . | 20–40 | . | 20–25 | 20–55 | . | . | . | 25–30 | . | 15–20 | . | . | . | 15–20 | 10–20 | . |
| Populus tremula | + | 1 | + | . | . | 1 | . | + | 1 | . | + | . | + | 2 | + | 2 | 1 | + | + | . | . | 1 | 1 | . | . | 1 | + |
| Высота, м Height, m |
. | 20–25 | 19– 21 | . | . | . | . | 20 | 20 | . | 14 | . | . | 22–27 | 20–23 | 18 | 25–27 | 27 | 25–26 | . | . | 19 | . | . | . | 20–25 | . |
| Диаметр, см Diameter, cm |
. | 12–25 | 24–48 | . | . | . | . | . | . | . | . | . | . | 37–44 | 25–26 | . | 20–30 | 50 | 40 | . | . | . | . | . | . | 40 | . |
| Древостой 2–го полога Stand of the 2nd canopy | . | 0.2 | . | . | . | . | . | . | . | . | 0.1 | . | . | . | . | . | . | . | 0.5 | . | 0.1 | . | . | . | . | . | . |
| Picea abies | . | 10 | + | . | . | + | . | . | . | + | 7 | . | . | + | + | . | . | . | 10 | . | 10 | . | . | . | . | . | . |
| Высота, м Height, m |
. | 10–16 | 9–16 | . | . | 12 | . | . | . | 9–16 | 12 | . | . | 10–17 | 10–17 | . | . | . | 14–18 | . | 10–15 | . | . | . | . | . | . |
| Диаметр, см Diameter, cm |
. | 8–16 | 8–15 | . | . | . | . | . | . | 10–27 | . | . | . | 13–20 | 11–25 | . | . | . | 10 | . | 10 | . | . | . | . | . | . |
| Betula pendula+B.pubescens | . | + | + | . | . | . | . | . | . | . | . | . | . | + | + | . | . | . | . | . | . | . | . | . | . | . | . |
| Высота, м Height, m |
. | 8 | 8–16 | . | . | . | . | . | . | . | . | . | . | 10–16 | 6–17 | . | . | . | . | . | . | . | . | . | . | . | . |
| № описания/№ releve | 1 | 2' | 3' | 4 | 5 | 6 | 7 | 8 | 9 | 10' | 11 | 12 | 13 | 14' | 15' | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 |
| Диаметр, см Diameter, cm |
. | 13 | 8–12 | . | . | . | . | . | . | . | . | . | . | 9–17 | 9–28 | . | . | . | . | . | . | . | . | . | . | . | . |
| Alnus incana | . | + | . | . | . | . | . | . | + | + | 1 | . | . | . | + | . | . | . | . | . | . | + | . | . | 1 | . | . |
| Высота, м Height, m |
. | 13 | . | . | . | . | . | . | 7 | 8–10 | 9–13 | . | . | . | 6–8 | . | . | . | . | . | . | 15 | . | . | 17 | . | . |
| Диаметр, см Diameter, cm |
. | 14 | . | . | . | . | . | . | . | 9–14 | . | . | . | . | 11–17 | . | . | . | . | . | . | . | . | . | 10 | . | . |
| Sorbus aucuparia | . | . | . | . | . | . | . | . | . | + | 2 | . | + | + | + | + | . | . | . | + | . | + | + | . | . | . | . |
| Высота, м Height, m |
. | . | . | . | . | . | . | . | . | 10 | 6 | . | 11 | . | 6–13 | 11 | . | . | . | 6 | . | 7 | . | . | . | . | . |
| Диаметр, см Diameter, cm |
. | . | . | . | . | . | . | . | . | 9 | . | . | 4–10 | . | 6–15 | . | . | . | . | . | . | . | . | . | . | . | . |
| Подрост, % Regrowth, % |
5 | 2 | 2 | 5 | 10 | 7 | 7 | 2 | 10 | 5 | 4 | 1 | 5 | 7 | 3 | 7 | 3 | 8 | + | 5 | 2 | 5 | 5 | 5 | 15 | + | + |
| Средняя высота, м Average height, m |
1 | 0.8 | 1.3 | 1 | 1 | 1 | 2 | 1 | 2 | 1 | 2 | 2 | . | 1 | 1.5 | 1.5 | 1.2 | 3 | 1 | 1 | 1.2 | 3 | 1.5 | 1.5 | 4.5 | . | 1 |
| Жизненность подроста ели Spruce regrowth vitality |
2 | . | . | . | . | . | . | . | . | 2-3 | . | 2-3 | . | . | . | . | 2-3 | 3 | . | . | 2-3 | . | 2-3 | . | . | . | . |
| Picea abies | 3 | 2 | + | 2 | 5 | 5 | 1 | + | 1 | 1 | 2 | + | 3 | + | 2 | 1 | 1 | 3 | . | 2 | 1 | 1 | 2 | 1 | 2 | + | + |
| Acer platanoides | . | . | . | . | . | . | 5 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Betula pendula+B.pubescens | . | . | . | . | . | . | . | . | + | . | + | . | . | + | . | . | . | . | . | . | . | 1 | + | . | . | . | . |
| Populus tremula | . | . | . | 1 | 1 | 1 | . | 1 | 5 | . | + | . | . | + | + | 1 | . | . | . | + | . | 1 | 1 | 1 | 1 | . | . |
| Sorbus aucuparia | 2 | + | + | 2 | 5 | 1 | 1 | 2 | 5 | 3 | 1 | 1 | 2 | + | 3 | 3 | 2 | 5 | + | 3 | 1 | 1 | 2 | 2 | 3 | . | . |
| Alnus incana | . | . | . | . | . | . | . | . | 1 | 1 | 1 | . | . | 5 | + | 2 | . | . | . | . | . | 1 | + | 2 | 7 | . | . |
| Padus avium | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | 1 | . | 2 | . | . |
| Кустарники, % Shrubs, % |
2 | . | . | 5 | . | 1 | 1 | 1 | 1 | 0.1 | 1 | 0.1 | 5 | 2 | . | + | 5 | 5 | . | . | 1 | 1 | 1 | 1 | 1 | . | . |
| Средняя высота, м Average height, m |
1 | . | . | 0.5 | . | 1 | . | 1 | 0.5 | . | 0.5 | . | . | 1.5 | . | 1.5 | . | . | . | . | . | 0.5 | 1.7 | 0.4 | 1 | . | . |
| Juniperus communis | 2 | . | + | 5 | . | 1 | . | 1 | 1 | + | + | . | . | 2 | + | + | . | . | . | . | . | . | . | . | + | . | . |
| Rubus idaeus | . | . | . | . | . | . | . | + | . | . | + | + | 1 | . | 1 | . | . | 3 | . | . | 1 | + | + | . | 1 | . | . |
| Lonicera xylosteum | . | . | . | . | . | . | + | . | . | . | . | . | 1 | . | + | + | 2 | 2 | . | . | . | + | + | . | + | . | . |
| Viburnum opulus | . | . | . | . | . | . | . | . | . | . | . | . | 1 | + | . | . | 1 | . | . | . | . | + | . | . | . | . | . |
| Daphne mezereum | . | . | . | . | . | . | . | . | . | + | 1 | . | 1 | . | . | . | 2 | . | . | . | . | . | . | . | . | . | . |
| Frangula alnus | . | . | . | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | 1 | + | . | . |
| № описания/№ releve | 1 | 2' | 3' | 4 | 5 | 6 | 7 | 8 | 9 | 10' | 11 | 12 | 13 | 14' | 15' | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 |
| Травяно-кустарничковый ярус, % Herb-dwarfshrub layer, % |
7 | 12 | 15 | 12 | 10 | 5 | 20 | 25 | 20 | 35 | 10 | 35 | 70 | 25 | 35 | 45 | 35 | 70 | 20 | 10 | 30 | 40 | 30 | 25 | 30 | 50 | 60 |
| Vaccinium vitis-idaea | 2 | 2 | 3 | 4 | 2 | + | 1 | 5 | 1 | 3 | 5 | 3 | 1 | 1 | 2 | 2 | 5 | 6 | 1 | + | + | + | 3 | 4 | 1 | 3 | + |
| Vaccinium myrtillus | 4 | 10 | 5 | 3 | 5 | 1 | 5 | 7 | 5 | 5 | 7 | 10 | 15 | 7 | 5 | 5 | 12 | 20 | 1 | 1 | 5 | 0 | 1 | 0 | 1 | 1 | 1 |
| Calamagrostis arundinacea | . | 1 | 3 | 2 | + | + | 2 | 7 | 5 | 3 | 2 | + | 5 | 5 | 25 | 25 | 20 | 20 | . | 1 | 1 | . | 6 | + | . | . | . |
| Maianthemum bifolium | 1 | 2 | + | . | 2 | + | 3 | 1 | 2 | 2 | 3 | 1 | 5 | 5 | 4 | 5 | 1 | 12 | . | 1 | 5 | 3 | 5 | 3 | 5 | 3 | 1 |
| Trientalis europaea | + | 1 | + | 1 | + | 1 | + | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 1 | 3 | 1 | 5 | . | + | 1 | 1 | 1 | 1 | 1 | 1 | + |
| Melampyrum pratense | . | 1 | . | 1 | . | 1 | + | 3 | 1 | 1 | + | . | + | 2 | + | 3 | . | 3 | . | + | . | . | . | 1 | . | . | . |
| Avenella flexuosa | + | . | 3 | 1 | 4 | 1 | . | 3 | . | 1 | + | + | 3 | 5 | 3 | 1 | . | + | + | 1 | 10 | + | 1 | + | + | . | . |
| Orthilia secunda | . | . | + | + | . | . | . | + | + | 2 | + | 2 | . | 5 | + | . | . | . | + | + | . | + | + | 1 | + | 2 | 3 |
| Luzula pilosa | + | 1 | + | 1 | + | 1 | + | 1 | 1 | + | + | . | 1 | . | 1 | . | . | 3 | 2 | + | 1 | 3 | 1 | 1 | . | . | . |
| Solidago virgaurea | . | 1 | . | + | + | + | + | + | 3 | 1 | + | . | + | 1 | + | 1 | + | . | 1 | + | . | + | + | 1 | + | . | . |
| Polypodium vulgare | . | . | + | + | . | + | . | 1 | + | + | + | . | . | . | + | + | . | . | . | + | . | . | . | . | . | . | . |
| Pyrola rotundifolia | . | . | . | + | . | . | . | . | + | 1 | . | . | + | 2 | . | + | . | . | . | . | . | . | . | 2 | . | . | . |
| Carex globularis | . | . | + | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | + | + | . |
| Linnaea borealis | . | 2 | . | . | . | . | 1 | . | . | . | . | . | . | . | 2 | . | . | . | . | 1 | + | . | . | . | . | . | . |
| Goodyera repens | . | 2 | . | + | . | + | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . |
| Lycopodium annotinum | . | 1 | . | . | . | . | + | + | . | 1 | . | . | . | . | . | . | . | . | . | + | + | + | + | + | + | . | 1 |
| Huperzia selago | . | . | . | . | . | . | . | + | . | 1 | . | . | . | . | + | 1 | . | . | . | . | . | . | + | + | + | . | . |
| Veronica officinalis | . | . | . | . | . | . | . | . | . | + | . | . | + | 2 | + | 1 | . | . | . | + | + | 1 | + | + | . | . | . |
| Pteridium aquilinum | . | . | . | . | . | . | . | . | . | . | . | . | . | 3 | . | . | . | 3 | . | . | . | . | 3 | . | . | . | . |
| Oxalis acetosella | . | . | . | . | + | + | 5 | 3 | 2 | 15 | 7 | 25 | 30 | 10 | 20 | 25 | 15 | 30 | 10 | 6 | 15 | 25 | 7 | 5 | 7 | 3 | 7 |
| Rubus saxatilis | . | 1 | . | + | . | + | 2 | 1 | 2 | 3 | + | . | 3 | 3 | 2 | 5 | . | 5 | 1 | 1 | 3 | . | 5 | 1 | 1 | 2 | . |
| Convallaria majalis | . | 1 | . | 1 | . | + | . | + | 2 | 1 | + | . | 3 | 5 | + | 5 | 1 | . | . | 1 | . | + | + | + | + | . | . |
| Carex digitata | . | . | . | . | . | + | + | 1 | 1 | 3 | . | 1 | 3 | 2 | 5 | 5 | . | 1 | . | + | 3 | 2 | + | . | . | . | . |
| Fragaria vesca | . | . | . | . | . | . | + | + | 7 | + | + | 1 | 1 | 3 | 2 | 5 | + | 4 | 2 | . | + | 3 | . | + | + | . | . |
| Hepatica nobilis | . | . | . | . | . | + | + | 2 | 5 | 2 | 2 | 2 | 20 | . | 5 | 5 | 3 | 10 | . | + | 2 | 5 | . | . | . | . | . |
| Dryopteris filix-mas | . | . | . | . | . | . | + | . | + | . | . | . | 3 | . | + | 1 | 1 | 2 | . | + | . | + | + | . | . | . | . |
| Lathyrus vernus | . | . | . | . | . | + | . | . | . | . | . | . | + | . | + | + | 3 | . | . | . | . | . | . | . | . | . | . |
| Paris quadrifolia | . | . | . | . | . | . | . | . | . | + | . | + | . | . | + | . | + | . | . | . | . | + | . | . | . | . | . |
| Actaea spicata | . | . | . | . | . | . | . | . | . | . | . | . | + | . | + | . | . | . | 1 | . | . | + | . | . | . | . | . |
| Milium effusum | . | . | . | . | . | . | . | + | . | . | . | . | 5 | . | + | . | . | . | . | . | + | 5 | . | . | . | . | . |
| Aegopodium podagraria | . | . | . | . | . | . | . | . | . | + | + | . | . | 7 | . | + | . | . | . | . | . | + | . | . | . | . | . |
| Stellaria holostea | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . | . | 3 | . | . | . | . | . |
| Circaea alpina | . | . | . | . | . | . | . | . | . | . | . | . | + | . | + | . | . | . | . | . | . | . | . | . | . | . | . |
| Hieracium murorum | . | . | . | . | . | . | . | + | . | . | . | . | . | 1 | . | + | . | . | . | . | . | . | . | . | . | . | . |
| Veronica chamaedrys | . | . | . | . | . | . | . | . | 1 | + | . | . | + | 1 | + | 1 | + | 3 | . | + | . | + | . | + | . | . | . |
| Geranium sylvaticum | . | . | . | . | . | . | . | + | . | . | . | . | + | + | + | + | . | . | . | . | . | + | . | 2 | . | . | . |
| Angelica sylvestris | . | . | . | . | . | . | . | . | . | 2 | + | . | . | 1 | . | + | + | . | . | . | . | + | . | + | 2 | . | . |
| Thelypteris phegopteris | . | . | . | . | . | . | . | . | . | . | . | . | 3 | . | + | . | . | . | . | + | . | . | 2 | + | . | . | . |
| Viola canina | . | . | . | . | . | . | . | + | + | . | . | . | . | 1 | + | . | . | . | . | . | . | + | . | . | . | . | . |
| Viola riviniana | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | 1 | . | . | . | . | 1 | . | . | + | . | . |
| Campanula persicifolia | . | . | . | . | . | . | . | . | 1 | . | . | . | . | + | + | + | . | . | + | . | . | + | . | . | . | . | . |
| Equisetum sylvaticum | . | . | . | . | . | . | + | . | . | . | 1 | + | 1 | 2 | + | . | . | . | . | . | . | 2 | 1 | 5 | 20 | 20 | 5 |
| Dryopteris carthusiana | . | . | + | . | + | . | + | + | . | 1 | 2 | 1 | + | . | 5 | 3 | . | . | 2 | + | + | 2 | 7 | 5 | 20 | 5 | 10 |
| Gymnocarpium dryopteris | . | . | . | . | . | . | 1 | . | . | 1 | + | 1 | 3 | . | 20 | 2 | . | . | . | 1 | . | 2 | 5 | 1 | 7 | 8 | 1 |
| Deschampsia cespitosa | . | . | . | . | . | . | . | . | . | . | 1 | . | . | 1 | . | 2 | . | + | . | . | + | 1 | 1 | 7 | 3 | + | 2 |
| Viola palustris | . | . | . | . | . | . | . | . | . | . | . | 2 | . | . | . | . | . | . | . | . | + | 1 | 2 | + | + | 5 | 8 |
| Athyrium filix-femina | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | + | 3 | + | 10 | 20 |
| Dryopteris expansa | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | 5 | 7 |
| Мохово-лишайниковый ярус, % Mosses-lichen layer, % |
95 | 90 | 95 | 65 | 85 | 40 | 50 | 65 | 40 | 70 | 7 | 90 | 50 | 35 | 60 | 50 | 20 | 45 | 40 | 60 | 80 | 25 | 60 | 60 | 15 | 95 | 45 |
| Pleurozium schreberi | 40 | 30 | 20 | 45 | 30 | 25 | 20 | 10 | 10 | 25 | 1 | 25 | 15 | 5 | 10 | 10 | 20 | 15 | 2 | 30 | 30 | 2 | 10 | + | + | . | . |
| Dicranum polysetum | 20 | 30 | 35 | 5 | 25 | 7 | 15 | 10 | 10 | 20 | 1 | 25 | 10 | 10 | 7 | 3 | + | 5 | 2 | 5 | 25 | 2 | 5 | 3 | 5 | + | . |
| Hylocomium splendens | 30 | 25 | 20 | 5 | 25 | 2 | 5 | 30 | 5 | 20 | 1 | 30 | 10 | 5 | 30 | 7 | . | 5 | 30 | 20 | . | . | . | 3 | + | . | . |
| Rhytidiadelphus triquetrus | . | . | . | 1 | 5 | 2 | 10 | 10 | 10 | 5 | 1 | 3 | 15 | 10 | 5 | 25 | + | 20 | 2 | 1 | 20 | 10 | 7 | 15 | + | . | . |
| Rhodobryum roseum | . | . | . | . | + | . | . | . | + | . | . | 5 | . | . | + | 3 | . | . | 2 | 1 | . | + | . | . | . | . | . |
| Plagiomnium elatum | . | . | . | . | . | 1 | . | 2 | + | . | 1 | . | . | 1 | 2 | 1 | . | . | 2 | + | . | 7 | + | 3 | 2 | . | 20 |
| Polytrichum commune | 1 | . | 1 | . | 1 | + | 1 | . | . | . | . | + | . | + | + | . | . | . | . | 1 | . | 3 | 5 | + | + | . | 1 |
| Sphagnum girgensohnii | 2 | . | 20 | . | . | . | 2 | . | . | . | 1 | + | . | . | 5 | . | . | . | . | 1 | 2 | . | 25 | 30 | 5 | 50 | 15 |
| Sphagnum centrale | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 10 | 5 |
| Rhizomnium pseudopunctatum | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 35 | . |
| Ptilium crista-castrensis | . | 3 | . | . | + | . | . | + | . | . | . | + | . | . | + | . | . | . | . | + | . | . | + | . | . | . | . |
| Ptilidium ciliare | . | . | + | . | . | + | . | + | 1 | . | . | . | . | . | . | . | . | . | . | + | . | + | . | . | . | . | . |
| Peltigera aphthosa | . | 2 | . | . | . | + | . | + | 2 | . | . | . | . | . | + | 1 | . | . | . | . | . | . | . | . | . | . | . |
| Видовое богатствоSpecies richness:травяно-кустарничковый ярусherb-dwarfshrub layer | 6 | 13 | 11 | 15 | 10 | 18 | 19 | 26 | 22 | 28 | 31 | 17 | 30 | 38 | 37 | 35 | 16 | 18 | 12 | 24 | 20 | 41 | 28 | 37 | 27 | 15 | 14 |
| мохово-лишайниковый ярус, % mosses-lichen layer, % |
8 | 5 | 9 | 5 | 7 | 10 | 6 | 10 | 9 | 4 | 6 | 11 | 4 | 7 | 14 | 8 | 3 | 4 | 6 | 11 | 5 | 10 | 8 | 10 | 8 | 4 | 5 |
| № описания/№ releve | 1 | 2' | 3' | 4 | 5 | 6 | 7 | 8 | 9 | 10' | 11 | 12 | 13 | 14' | 15' | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 |
Примечание
В таблицах 1, 2 соотношение пород древостоя (по числу особей) дано по 10-бальной шкале. Для подроста, кустарникового, травяно-кустарничкового и мохово-лишайникового ярусов приведено проективное покрытие видов (%). Ассоциации и варианты ассоциаций: 4 – ельники травяно-папоротниковые; 2а – ельники вейниково-чернично-кислично-зеленомошные с неморальными видами; 3a – ельники кислично-зеленомошные с неморальнотравными видами; 3b – ельники кислично-папоротниково-моховые; 4a – ельники щитовниково-хвощевые; 4b – ельники кочедыжниковые. Описания со знаком “'” были выполнены Г. А. Исаченко. Возраст древостоя: 3' – ели 40–80 лет (результаты бурения 1996 г.); 14' – ели 40–80 лет, сосны 70 лет (1991 г.); 15' – ели и сосны 40–80, реже 80–120 лет (2001 г.).
Note
In tables 1, 2 the proportion of tree species (by the number of individuals) is given on a 10-point scale. The projective cover (%) of species are provided for regrowth, shrub, grass-undershrub and moss-lichen layers. Associations and variants of associations: 4 – Piceetum herboso-dryopteridosum; 2a – Piceetum calamagrostidosum arundinaceosum myrtilloso-oxalidoso-hylocomiosum nemoroso-herbosum; 3a – Piceetum oxalidoso-hylocomiosum nemoroso-herbosum; 3b – Piceetum oxalidoso-dryopteridoso-hylocomiosum; 4a – Piceetum dryopteridoso-equisetosum; 4b – Piceetum athyridosum. Releves with the sign “'” were made by the G. A. Isachenko. Age of stand: 3' – spruce 40–80 years (the results of the drilling stand 1996); 14' – spruce 40–80 years, pine 70 years (1991); 15' – spruce and pine 40–80, rarely 80–120 years (2001).
Ниже приводятся малообильные виды, встреченные в 1–3 описаниях (первая цифра – номер описания, в скобках – проективное покрытие вида).
Species, which were found in 1–3 releves with small abundance, are given below (the first digit is the releve number, in parentheses is the projective cover of the species).
Кустарники / Shrubs: Ribes nigrum 7(+); Ribes rubrum 13(1); Salix caprea 14'(2); Salix cinerea 11(+). Травянистые растения/ Herbaceous plants: Agrostis tenuis 14'(1), 16(+); Alchemilla vulgaris s. l. 14'(+); Anthriscus sylvestris 22(1), 24(+); Calamagrostis canescens 23(1), 26(2); Campanula glomerata 14'(+); C. rotundifolia 8(+), 14'(1); Carex canescens 23(+), 25(+); C. pallescens 14'(1), 25(+); Cerastium holosteoides 15'(+); Cirsium heterophyllum 22(+); Crepis paludosa 22(+), 25(2); Equisetum pratense 11(+); Galium boreale 14'(1); G. album 16(+); G. palustre 11(+); G. uliginosum 24(+); Geum rivale 11(+), 22(+), 24(3); Hypericum maculatum 11(+), 14'(+), 24(+); Hypopitys monotropa 6(+); Listera ovata 10'(+); Melica nutans 16(+); Moehringia trinervia 21(+); Platanthera bifolia 4(+), 14'(+); Poa nemoralis 23(+), 24(+); Poa palustris 25(+); Polygonatum odoratum 18(1); Potentilla erecta 9(+), 14'(1), 24(1); Prunella vulgaris 11(+), 22(+), 24(1); Pyrola minor 8(+); Ranunculus auricomus 14'(1); Ranunculus fallax s. l. 11(+), 16(+), 22(+); Ranunculus repens 11(+), 22(+), 24(+); Stellaria graminea 16(+); S. nemorum 13(+); Succisa pratensis 14'(1), 24(+); Thyselium palustre 11(+); Trifolium medium 14'(+); Trollius europaeus 22(+); Vicia sepium 22(+). Мхи / Mosses: Aulacomnium palustre 1(1), 15'(+), 24(+); Climacium dendroides 22(+), 24(2); Dicranum scoparium 6(3), 20(1), 22(2); Pohlia nutans 12(+); Sciuro-hypnum oedipodium 21(3); Sphagnum angustifolium 1(+), 27(1); Sphagnum russowii 27(3); Barbilophozia barbata 12(1); Plagiochila asplenioides 12(2). Лишайники / Lichens: Cladonia alpestris 15'(+); Cladonia sylvatica 1(+), 3'(+), 8(+), 15'(+); Cetraria islandica 3'(+).
Таблица 2.
Геоботанические описания еловых сфагновых лесов (Северо-Западное Приладожье). Table 2. Releves of spruce sphagnous forests (North-Western Ladoga region)
| Ассоциации/Associations | 5. Piceetum myrtilloso-sphagnosum | 6. Piceetum herboso-sphagnosum | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Варианты/Variants | 6a | |||||||||
| Год/Year | 2004 | 1997 | 2004 | 2004 | 2004 | 1993 | 2004 | 2004 | 2004 | 1993 |
| № описания/№ releve | 28 | 29 | 30 | 31 | 32 | 33' | 34 | 35 | 36 | 37' |
| Сомкнутость крон: Density of crown: |
0.4 | 0.3 | 0.6 | 0.7 | 0.7 | 0.5 | 0.4 | 0.3 | 0.6 | 0.5 |
| Древостой 1-го полога Stand of the 1st canopy |
. | . | . | . | . | . | . | . | . | . |
| Picea abies | 6 | 5 | 7 | 10 | 9 | 10 | 4 | 4 | 5 | 6 |
| Высота, м/Height, m | 20–23 | 18 | 25 | 30 | 20–25 | 17–26 | 12 | 10–16 | 10–15 | 7–13 |
| Диаметр, см/Diameter, cm | 30–45 | 20 | 30–50 | 15–40 | 10–30 | 20–44 | 15 | 10–15 | 10–20 | 12–28 |
| Pinus sylvestris | + | 2 | 1 | + | 1 | + | 3 | 2 | + | 1 |
| Высота, м/Height, m | 18 | 18–20 | 22–23 | . | 22 | 18–23 | 15 | 15–17 | 18 | 14–21 |
| Диаметр, см/Diameter, cm | 20 | 20 | 25 | . | 20–30 | 24–40 | 20–35 | 20–25 | 15–20 | 20–44 |
| Betula pubescens | 2 | 2 | 2 | . | + | + | 3 | 4 | 5 | 3 |
| Высота, м/Height, m | 17–19 | 18–20 | 20–22 | . | 20 | 15–18 | 12 | 18 | . | 9–20 |
| Диаметр, см/Diameter, cm | 15–25 | . | 15–20 | . | 19–25 | 16–20 | 8–12 | 10–20 | . | 12–25 |
| Populus tremula | 2 | 1 | . | . | + | + | . | + | . | . |
| Высота, м/Height, m | 20 | 19 | . | . | . | 20–22 | . | 18–19 | . | . |
| Диаметр, см/Diameter, cm | 30–35 | . | . | . | . | 44 | . | 30 | . | . |
| Древостой 2-го полога Stand of the 2nd canopy |
. | . | . | . | . | . | . | . | . | 0.4 |
| Picea abies | + | . | + | . | . | + | . | . | . | 1 |
| Высота, м/Height, m | 17 | 10 | 17 | . | . | 8–15 | . | . | . | 6 |
| Диаметр, см/Diameter, cm | 15–20 | . | 15 | . | . | 8–16 | . | . | . | 8 |
| Betula pubescens | . | . | . | . | + | + | . | . | . | + |
| Высота, м/Height, m | . | . | . | . | 16–17 | 11 | . | . | . | 7 |
| Диаметр, см/Diameter, cm | . | . | . | . | 15 | 12 | . | . | . | 8 |
| Подрост, % Regrowth, % |
2 | 10 | 5 | 7 | 5 | 1 | 10 | 1 | . | 5 |
| Средняя высота, м Average height, m |
1.5 | 1.5 | 1.2 | 1.2 | . | 2 | 2.5 | 1.5 | . | 1,5 |
| Жизненность подроста ели Spruce regrowth vitality |
2 | . | 2–3 | . | 2–4 | . | 3 | . | . | . |
| Picea abies | 1 | 2 | 5 | 5 | 5 | 1 | 10 | 1 | . | 4 |
| Betula pubescens | . | . | . | 1 | . | . | . | + | . | + |
| Populus tremula | . | 3 | . | . | . | + | . | . | . | . |
| Sorbus aucuparia | 1 | 5 | . | 2 | . | + | . | . | . | + |
| Кустарники, % Shrubs, % |
. | 1 | . | . | + | . | 1 | . | . | 10 |
| Средняя высота, м Average height, m |
. | . | . | . | . | . | . | . | . | 3 |
| Salix aurita | . | . | . | . | + | . | 1 | . | . | 5 |
| Травяно-кустарничковый ярус, % Herb-dwarfshrub layer, % |
15 | 15 | 12 | 10 | 20 | 20 | 10 | 20 | 10 | 20 |
| Vaccinium myrtillus | 5 | 5 | 10 | 3 | 15 | 15 | + | 1 | 2 | 1 |
| Vaccinium vitis-idaea | 3 | 3 | 2 | 3 | 5 | 3 | 3 | 3 | 4 | + |
| Trientalis europaea | 2 | + | . | 1 | 3 | 1 | . | + | . | + |
| Maianthemum bifolium | 5 | 1 | 1 | 3 | 5 | 1 | . | . | + | . |
| Dryopteris carthusiana | + | + | 1 | . | 2 | . | . | . | + | + |
| Orthilia secunda | . | 2 | 3 | . | . | . | . | + | + | . |
| Carex globularis | . | 1 | . | 1 | 1 | 3 | 1 | . | 1 | 1 |
| Equisetum sylvaticum | . | . | + | . | 2 | + | 5 | 10 | 5 | 10 |
| Comarum palustre | . | . | . | . | . | . | 3 | 6 | 2 | 8 |
| Calamagrostis canescens | . | . | . | . | . | . | . | 2 | + | 2 |
| Equisetum fluviatile | . | . | . | . | . | . | . | . | . | 1 |
| Viola palustris | 3 | . | . | . | . | . | . | . | . | + |
| Athyrium filix-femina | + | . | . | . | . | . | . | . | + | . |
| Gymnocarpium dryopteris | + | . | . | . | . | . | . | . | + | . |
| Calamagrostis arundinacea | 2 | 3 | 2 | . | . | . | . | . | . | . |
| Мохово-лишайниковый ярус, % Mosses-lichen layer, % |
50 | 50 | 90 | 100 | 90 | 85 | 100 | 100 | 100 | 100 |
| Pleurozium schreberi | 3 | 10 | 5 | + | 3 | 5 | . | . | 5 | . |
| Dicranum polysetum | 2 | 10 | 5 | + | 3 | 5 | . | . | . | . |
| Hylocomium splendens | 2 | + | 5 | . | 3 | 1 | . | . | . | . |
| Rhytidiadelphus triquetrus | 3 | . | . | . | . | + | . | . | . | . |
| Polytrichum commune | 10 | 5 | 5 | 5 | 10 | 5 | 5 | 10 | 5 | 10 |
| Sphagnum girgensohnii | 30 | 25 | 70 | 95 | 70 | 70 | 80 | 90 | 60 | 70 |
| Sphagnum centrale | . | . | . | . | . | . | 8 | . | 20 | 10 |
| Sphagnum angustifolium | . | . | . | . | . | . | 5 | + | 5 | . |
| Sphagnum russowii | . | . | . | . | . | . | . | . | 5 | . |
| Sphagnum riparium | . | . | . | . | 1 | . | . | . | . | 5 |
| Sphagnum fimbriatum | . | . | . | . | . | . | . | . | . | 5 |
| Sphagnum cuspidatum | . | . | . | . | . | . | 2 | . | . | . |
| Видовое богатство:Species richness:травяно-кустарничковый ярусherb-dwarfshrub layer | 11 | 12 | 11 | 5 | 8 | 8 | 5 | 7 | 12 | 13 |
| Мохово-лишайниковый ярус, % Mosses-lichen layer, % |
6 | 5 | 5 | 4 | 5 | 6 | 2 | 2 | 4 | 2 |
Примечание
В таблице соотношение пород древостоя (по числу особей) дано по 10-бальной шкале. Для подроста, кустарникового, травяно-кустарничкового и мохово-лишайникового ярусов приведено проективное покрытие видов (%). Варианты ассоциаций: 6a – сосново-березово-еловые сабельниково-хвощево-сфагновые. Описания со знаком “'” были выполнены Г. А. Исаченко. Возраст древостоя: 33'– ели 40, 65, 85, 128 лет, сосны 78 лет; 37' – ели 33, 55, 80 лет, березы 69 лет, сосны 82 года (данные бурения 1993 г).
Note
In tables the proportion of tree species (by the number of individuals) is given on a 10-point scale. The projective cover (%) of species are provide for regrowth, shrub, herb-undershrub and moss-lichen layers. Variants of associations: 6a – Pineto-betulo-piceetum comaroso-equisetoso-sphagnosum. Releves with the sign “'” were made by the G. A. Isachenko. Age of stand: 33' – spruce 40, 65, 85, 128 years, pine 78 years; 37' – spruce 33, 55, 80 years, birch 69 years, pine 82 years, (the results of the drilling stand 1993.
Ниже приводятся малообильные виды, встреченные в 1–2 описаниях (первая цифра – номер описания, в скобках – проективное покрытие вида). Species, which were found in 1–3 releves with small abundance, are given below (the first digit is the revele number, in parentheses is the projective cover of the species).
Кустарники / Shrubs: Salix cinerea 32(+); Frangula alnus, Sorbus aucuparia, Salix caprea, S. phylicifolia и S. myrsinifolia 37'(+). Травянистые растения / Herbaceous plants: Avenella flexuosa 29(3), 33'(3); Carex acuta 36(+); Deschampsia cespitosa 37'(+); Dryopteris filix-mas 28(+); Hepatica nobilis 30(+); Juniperus communis 29(+); Luzula pilosa 29(1), 32(1); Melampyrum pratense 29(1); Oxalis acetosella 30(2), 37'(+); Platanthera bifolia 33'(+); Pteridium aquilinum 28(+), 29(+); Rubus saxatilis 30(+); Scirpus sylvaticus 37'(1); Vaccinium uliginosum 30(1).
ХАРАКТЕРИСТИКА ЕЛОВЫХ ЛЕСОВ
Типичные местоположения еловых лесов Северо-Западного Приладожья – склоны сельг, пологие внутрисельговые равнины, заболоченные межсельговые ложбины и озерные глинистые террасы (Makarova, 2011). На основе эколого-фитоценотической классификации автором было выделено 3 группы ассоциаций (зеленомошные, травяные и сфагновые), 6 ассоциаций и 6 вариантов ассоциаций еловых лесов.
ЕЛОВЫЕ ЗЕЛЕНОМОШНЫЕ ЛЕСА
Ельники зеленомошные типичны для таежной зоны, произрастают на различных по механическому составу почвах со средним или богатым минеральным питанием и умеренным увлажнением. По схеме типов леса (Fedorchuk et al., 2005) они относятся к малонарушенным еловым нормально дренированным лесам. В группе зеленомошных еловых лесов в Северо-Западном Приладожье выделяются ельники чернично-зеленомошные, ельники чернично-кислично-зеленомошные и ельники кислично-зеленомошные.
1. Ельники чернично-зеленомошные (табл. 1, оп. 1–6) занимают местообитания со средними показателями богатства почв и увлажнения – в основном слабоволнистые внутрисельговые равнинные участки, реже пологие склоны или террасированные ступени сельг. В примеси к ели обычны сосна и береза, реже осина. Сомкнутость древостоя 0.3–0.6. Ельники разновозрастные: от 50–60 до 90–100 лет. Высота древостоя колеблется в диапазоне 18–30 м, в среднем – 22 м. Диаметр стволов ели 15–40 см, до 50 см, сосны и березы – до 20–30 см. Во втором пологе местами отмечается ель (9–16 м выс.), реже – невысокие деревья ольхи серой или рябины. В подросте – ель с обилием до 5%, а также рябина, реже осина. Высота подроста ели не превышает 1 м. В кустарниковом ярусе редко отмечаются небольшие группы можжевельника (Juniperus communis)11. Травяно-кустарничковый ярус разрежен (общее проективное покрытие 5–15%), состоит из Vaccinium myrtillus, V. vitis-idaea, Maianthemum bifolium, Trientalis europaeus, Luzula pilosa. Изредка на камнях растут Polypodium vulgare, Linnaea borealis. В небольшом обилии была встречена Goodyera repens. Иногда произрастают такие виды, как Convallaria majalis, Rubus saxatilis, Lathyrus vernus. В составе травяно-кустарничкового яруса отмечено от 6 до 18 видов (общее количество – 25). Моховой покров слагают Hylocomium splendens, Dicranum polysetum, Pleurozium schreberi; другие виды мхов единичны. Приводимые здесь описания имеют группу видов относительно богатых почв и близки к ельникам черничникам богатым, выделяемым В. И. Василевичем (Vasilevich, 2004b).
Ельники чернично-зеленомошные описаны для всей таежной зоны (Korchagin, Senyaninova-Korchagina, 1957; Rysin, Saveleva, 2002), преобладают в среднетаежных еловых лесах (Morozova et al., 2008). Ельники чернично-зеленомошные характерны для Ленинградской обл. (Smirnova, 1928; Sokolov, 1929; Nitsenko, 1959, 1960; Dmitrieva, 1973; Vasilevich, 1983, 2004b; Volkova et al., 1999, 2003, 2007, 2017; Bibikova et al., 2002; Fedorchuk et al. 2005) и Карелии (Uskov, 1930; Zinserling, 1932; Vilikaynen, 1957, 1971; Kucherov et al., 2006). В травяно-кустарничковом ярусе этих лесов преобладает черника и отсутствует или встречается в небольшом количестве кислица и другие виды богатых почв (Vasilevich, 2015). Для Скандинавии приводится близкий по флористическому составу тип сообществ Picea abies – Vaccinium myrtillus (Pählsson, 1994). В Норвегии в ельниках черничных в травяно-кустарничковом ярусе черника, брусника и Avenella flexuosa доминируют вместе с Majanthemum bifolium, Trientalis europaeus, Orthilia secunda, Solidago virgaurea, Chamaepericlymenum suecicum, на более освещенных открытых местах может полностью доминировать луговик (Rekdal, Larson, 2005). Наиболее северные местонахождения ельников чернично-зеленомошных отмечены в Карелии вблизи границы северной и средней тайги (Yurkovskaya, Payanskaya-Gvozdeva, 1993; Morozova, Korotkov, 1999). На южном пределе распространения, близкие по составу ельники чернично-зеленомошные (асс. Eu-Piceetum abietis, вар. Quercus robur) описаны в Южном Нечерноземье и отнесены к интразональному типу растительности (Bulokhov, So-lomeshch, 2003). Основное отличие от ельников Северо-Западного Приладожья – участие дуба в составе древостоя.
2. Ельники чернично-кислично-зеленомошные (табл. 1, оп. 7–12), занимают слабоволнистые равнины, пологие склоны и подножия сельг. Сомкнутость древостоя 0.3–0.6. Ельники разновозрастные: средней спелости и приспевающие, возраст деревьев 50–100 лет. При высоте елей 19–20 м, диаметр стволов составляет 15–25 см. Ели высотой 23–27 (30) м имеют диаметр 30–50 см. Кроме ели в первом пологе часто присутствуют сосна, береза, осина. Во втором пологе изредка присутствует ель высотой до 12 м, рябина – 6 м и ольха серая – 7–13 м. В подросте (1–2 м выс.) с небольшим обилием встречаются ель, осина, рябина, береза, изредка клен. В кустарниковом ярусе эпизодически присутствуют Juniperus communis, Rubus idaeus, Lonicera xylosteum, Viburnum opulus, Frangula alnus, Daphne mezereum. Травяно-кустарничковый ярус средней сомкнутости (10–35%). В нем преобладают Vaccinium myrtillus и Oxalis acetosella, отмечаются другие бореальные виды: Calamagrostis arundinacea, Maianthemum bifolium, Trientalis europaeus, Avenella flexuosa, Solidago virgaurea, Dryopteris carthusiana, Gymnocarpium dryopteris, Carex digitata, Fragaria vesca, Equisetum sylvaticum. Местами встречается Hepa-tica nobilis. Проективное покрытие мохового покрова 40–90%, доминируют Hylocomium splendens, Dicranum polysetum, Pleurozium schreberi, Rhytidiadelphus triquetrus. В небольшом обилии присутствуют Plagiomnium elatum, Rhodobryum roseum.
В результате выборочных рубок на месте чернично-кислично-зеленомошных ельников образуются разреженные (сомкнутость 0.2) еловые древостои чернично-вейниково-кислично-зеленомошные с луговиком. Из-за осветления разрастаются Calamagrostis arundinacea (покрытие до 20–25%) и Avenella flexuosa (до 15%), происходит усиление доли участия подроста и подлеска до 20% (ель, береза, рябина). Ниценко (Nitsenko, 1960) описал кислично-вейниковые ельники, появляющиеся в результате осветления полога (при изреживании древостоя и усилении доли мелколиственных пород).
Существуют разные мнения относительно ельников с высоким покрытием черники и кислицы. Одно из них: к промежуточным сообществам между кисличниками и черничниками отнесены те ельники кислично-черничные, в которых преобладают бореальные, и практически не отмечены дубравные виды (Nitsenko, 1960; Dmitrieva, 1973). Похожие ельники кислично-черничные с Maianthemum bifolium, Rhytidiadelphus triquetrus приводятся для Южной Карелии (Vilikaynen, 1957), для северного побережья Финского залива (Bibikova et al., 2002). Другого мнения придерживается ряд авторов. С. Я. Соколов (Sokolov, 1929) рассматривал асс. Piceetum oxalidoso-myrtillosum в рамках типа леса Piceeta oxalidosa. А.П. Ильинским (Il’insky, 1921) была описана чернично-кисличная рамень в окрестностях Петергофа. Флористически ельники чернично-кисличные сходны с “типичными” кисличниками (Vasilevich, Bibikova, 2004b), для них характерно присутствие видов относительно более богатых субстратов (Alnus incana, Fragaria vesca, Rubus idaeus, Geranium sylvaticum, Lathyrus vernus), отсутствующих в ельниках черничниках (Vasilevich, 2004b; Kucherov, 2010). Для Северо-Западного Приладожья приводятся ельники кисличные с большим покрытием черники, сходные с нашими описаниями, в них преобладают бореальные, но характерны и неморальные виды (Fedorchuk et al., 1974; 2005).
Ельники чернично-вейниково-кислично-зеленомошные с неморальнотравными видами характерны для прогреваемых склонов южной, юго-западной и юго-восточной экспозиций и подножий сельг (табл. 1, оп. 13–18). Эти леса разновозрастные (возраст елей 40–90 лет), местами приспевающие и спелые (100–130 лет). Древостой слагает ель высотой от 20-22 до 28–30 м с диаметром стволов 10–50 см (изредка отмечались старовозрастные ели диаметром до 70 см), а также сосна (22–29 м выс., 20–50 см диам.), осина (18–27 м выс., 20–50 см диам.) и береза (17–27 м выс., от 20–24 до 55 см диам.). Сомкнутость древостоя 0.3–0.6. В подросте обычны ель, осина, ольха серая, рябина. В кустарниковом ярусе эпизодически встречаются Juniperus communis, Lonicera xylosteum, Daphne mezereum, Viburnum opulus, Frangula alnus, Rubus idaeus.
Этот тип ельников отличается, прежде всего, большим числом видов травяно-кустарничкового яруса (общее число видов – 38), а также усилением роли Calamagrostis arundinacea и неморального разнотравья (Hepatica nobilis, Dryopteris filix-mas, Lathyrus vernus, Aegopodium podagraria). Кроме того, появляются виды более влажных и богатых субстратов – Angelica sylvestris, Geranium sylvaticum, Hieracium murorum, Circaea alpina. Моховой покров характеризуется средней сомкнутостью (20–60%), преобладают Rhytidiadelphus triquetrus, Pleurozium schreberi, Hylocomium splendens, Dicranum polysetum. В небольшом обилии отмечен Plagiomnium elatum.
Выделение ельников неморальнотравных возможно в случае, когда суммарное покрытие неморальных видов достигает 10–15% (Vasilevich, Bibikova, 2004b). В нашем случае, имея суммарное покрытие для группы неморального разнотравья в пределах 10–25%, ассоциация ельников неморальнотравных не выделялась как самостоятельная, так как имелись другие виды-доминанты (кислица, вейник, черника). Как указывает В.И. Виликайнен (Vilikaynen, 1957), “ельники с неморальными элементами представляют выдвинутые на север “форпосты” южнотаежной группировки, но широкого распространения в Карелии они не имеют, чаще отмечаются в Южной Карелии совместно с осиной”. Сходные сообщества были описаны ранее Ю.Д. Цинзерлингом (Zinserling, 1932) как ельники с травами широколиственных лесов или Piceetum nemoriherbosum. Еловые и сосново-еловые неморальнотравно-зеленомошные леса с черникой, вейником, с мощным (до 70%) травяно-кустарничковым ярусом из видов неморальной и еловой свит приводятся для склонов, террас и межсельговых понижений Северного Приладожья (район Лахденпохья – Питкяранта) (Ipatov et al., 1998).
В описаниях (табл. 1, оп. 13–18) выделены 9 диагностических видов асс. Rhodobryo-Piceetum typicum Morozova et al. subass. nov. hoc locо: Picea abies, Athyrium filix-femina, Circaea alpina, Daphne mezereum, Gymnocarpium dryopteris, Oxalis acetosella, Phegopteris connectilis, Pyrola rotundifolia, Rhodobryum roseum. Согласно О.В. Морозовой и др. (Morozova et al., 2017) высока константность Maianthemum bifolium, Oxalis acetosella, Trientalis europaea, Actaea spicata, и отмечены Aegopodium podagraria, Lathyrus vernus, Paris quadrifolia, Stellaria holostea; из чего можно сделать вывод, что ельники чернично-вейниково-кислично-зеленомошные с неморальнотравными видами близки к асс. Rhodobryo-Piceetum typicum (ельникам неморальнотравным).
3. Ельники кислично-зеленомошные (табл. 1, оп. 19–21) встречаются в Северо-Западном Приладожье на пологих склонах и слабоволнистых ступенях сельг. Почвы достаточно богаты за счет подтока вод с верхних частей склонов и переноса и накопления делювиальных отложений. Местоположения дренированные. В древостое преобладает ель, присутствуют береза, осина, сосна. Сомкнутость древостоев – 0.4–0.7. Возраст елей 40–80 лет. Высота елей 20–22 м, диаметр стволов 20–25 см. Высота берез 19–24 м, диаметр до 30 см; высота осин 19–26 м, диаметр до 40 см; высота сосен 19–26 м, диаметр 30–40 см (единично были встречены старые сосны с диаметром до 60 см). Во втором пологе местами отмечается ель высотой 14–18 м, ольха серая – 15 м, рябина до 6–7 м. Подрост слабо развит. В кустарниковом ярусе изредка встречаются Rubus idaeus, Lonicera xylosteum, Viburnum opulus. В травяно-кустарничковом ярусе обычны Vaccinium myrtillus, V. vitis-idaea, Avenella flexuosa, Dryopteris carthusiana, Luzula pilosa, Solidago virgaurea, Maianthemum bifolium. Моховой покров, имеющий покрытие 40–60%, представлен видами Hylocomium splendens, Rhytidiadelphus triquetrus, Plagiomnium elatum, Rhodobryum roseum, Dicranum polysetum, Pleurozium schreberi.
В результате выборочных рубок на месте ельников кислично-зеленомошных образуются разреженные ельники мелкотравно-кислично-зеленомошные. С увеличением освещенности связано разрастание Avenella flexuosa, Maianthemum bifolium, Carex digitata. В травяно-кустарничковом покрове участвуют также Vaccinium myrtillus, Luzula pilosa, Dryopteris carthusiana, Rubus saxatilis, Hepatica nobilis. Из мхов обильны Pleurozium schreberi, Rhytidiadelphus triquetrus, Dicranum polysetum, небольшие куртинки образуют Sphagnum girgensohnii, Sciuro-hypnum oedipodium.
Были описаны два варианта. Первый – ельники кислично-зеленомошные с неморальнотравными видами (табл. 1, оп. 22) встречаются на пологих склонах сельг и отличаются от типичных кисличников наличием группы неморального и субнеморального разнотравья (Actaea spicata, Aegopodium podagraria, Dryopteris filix-mas, Hepatica nobilis, Stellaria holostea).
Сходные по флористическому составу ельники с большим обилием и участием неморальных видов (асс. Piceetum herboso-oxalidosum), в целом, являются типичными для южной тайги европейской части России (Rysin, Saveleva, 2002). Ранее для исследованного района отмечались ельники дубравно-травянисто-кисличные (Denisenkov, 1995). Для северо-западной части Карельского перешейка Е.В. Дмитриева (Dmitrieva, 1973) в пределах ельников зеленомошников выделяла ельники кисличники, внутри которых различала типичные (Piceetum oxalidosum) и ельники кисличники, обогащенные неморальным разнотравьем (Piceetum nemoroso-oxalidosum). Для восточной части Европейской России приводится асс. Melica nutantis–Piceetum abietis K.–Lund 1981 (Zaugolnova et al., 2009), близкая по своему флористическому составу к описанной в Северо-Западном Приладожье.
Второй вариант – ельники кислично-папоротниково-моховые (табл. 1, оп. 23–24) имеют переходный характер между кислично-зеленомошными и травяно-папоротниковыми еловыми лесами. Они развиваются на небольших внутрисельговых террасированных ступенях и слабовогнутых участках, в условиях временного переувлажнения. Древостой чаще смешанный, помимо ели обычна береза, в меньшей степени – осина и сосна. Высота деревьев 20–21 м, сомкнутость крон 0.3–0.5. В подросте обильны ель, ольха серая, рябина, реже осина и береза. Кустарниковый ярус слабо развит, но постоянно присутствует Frangula alnus. В травяно-кустарничковом ярусе примерно с равным обилием участвуют кислица, черника, брусника, костяника, майник, вейник. В небольшом количестве есть Equisetum sylvaticum, Dryopteris carthusiana, Gymnocarpium dryopteris, Deschampsia cespitosa. В моховом ярусе наряду с зелеными мхами (Rhytidiadelphus triquetrus, Pleurozium schreberi, Hylocomium splendens, Dicranum polysetum), становится заметна роль Sphagnum girgensohnii, появляется Polytrichum commune.
Сходные по составу сообщества выделял Ниценко (Nitsenko, 1960) как кислично-папоротниковые ельники, свойственные понижениям и ложбинам, часто с участием Stellaria nemorum. Ельники кислично-папоротниковые (Piceetum oxalidoso-filicosum) на относительно обедненных почвах с проточным увлажнением приводятся для северо-запада Карельского перешейка (Dmitrieva, 1973). На северном побережье Финского залива в районе пос. Комарово описаны ельники кислично-папоротниковые с участием неморального разнотравья (Bibikova et al., 2002). Для осушенных местоположений приводятся ельники кислично-папоротниковые с участием Athyrium filix-femina, Dryopteris expansa, Gymnocarpium dryopteris и группой разнотравья, в том числе высокотравья: Filipendula ulmaria, Cirsium oleraceum, Angelica sylvestris (Fedorchuk et al., 2005).
Ельники кисличные характерны для южнотаежной подзоны и повышают производительность к югу (Rysin, Saveleva, 2002). Близкие по флористическому составу описания ельников кислично-зеленомошных приводятся для всего Северо-Запада России (Il’insky, 1921; Smirnova, 1928; Sukachev, 1931; Zinserling, 1932; Nitsenko, 1960; Rysin, 1979; Vasilevich, 1983; Vasilevich, Bibikova, 2004b; Kucherov et al., 2006; Volkova et al., 1999, 2001, 2007). Для ассоциации свойственно доминирование Oxalis acetosella и преобладание бореальных видов, характерно участие неморальных видов (Vasilevich, 1983). Типичные для Карельского перешейка ельники кислично-зеленомошные довольно редко отмечаются в Псковской и Новгородской областях, переходя южнее в “безмоховые” кисличники (Vasilevich, Bibikova, 2004b).
ЕЛОВЫЕ ТРАВЯНЫЕ ЛЕСА
В.Н. Сукачев (Sukachev, 1931) выделял группу ельников болотно-травяных или ельников-логов (Piceetum fontinale). Описание травяных ельников приводится Ю.Д. Цинзерлингом (Zinserling, 1932): “… травяной покров густой, высокий и обычно разнообразный. Встречается по дну речек и ручьев, по логам, где имеется сток воды…”. В Северо-Западном Приладожье в сходных условиях (узкие лентообразные ложбины, нижние вогнутые части склонов сельг, богатые суглинистые подножия склонов с проточным режимом питания, изредка тальвеги) встречаются ельники травяно-папоротниковые. Моховой покров в этих сообществах очень неравномерный: сфагновые мхи перемежаются с куртинами зеленых мхов.
4. Ельники травяно-папоротниковые занимают вогнутые участки слабоволнистых глинистых озерных террас, подножия склонов сельг, внутрисельговые ложбины, представлены следующими вариантами ассоциации: щитовниково-хвощевыми, кочедыжниковыми и таволгово-папоротниковыми. В древостое преобладают крупные спелые и перестойные ели, в примеси участвуют береза, изредка осина, сосна или ольха серая. Высота елей варьирует от 20–24 до 30 м, диаметр – 10–50 см. Сомкнутость – 0.3–0.5. Еловый подрост высотой 1–5 м встречается постоянно. Кустарниковый ярус практически не развит.
Первый вариант – еловые с ольхой серой щитовниково-хвощевые леса (табл. 1, оп. 25), произрастают в более дренированных условиях. Травяно-кустарничковый ярус средней сомкнутости, но разнообразен по составу (до 27 видов). В нем преобладают Dryopteris carthusiana, Equisetum sylvaticum; другие виды гигромезофитного разнотравья (Calamagrostis canescens, Viola palustris, Deschampsia cespitosa) встречаются в меньшем обилии. Кислица и группа мезофитов, указывающая на более дренированные условия (Fragaria vesca, Convallaria majalis, Lycopodium annotinum, Gymnocarpium dryopteris, Carex pallescens), имеют незначительное покрытие. Моховой покров слабо развит, на повышениях отмечаются зеленые мхи (Dicranum polysetum, Pleurozium schreberi, Hylocomium splendens), а в понижениях появляется Sphagnum girgensohnii.
Для северо-запада Карельского перешейка Е. В. Дмитриева (Dmitrieva, 1973) описала ельник папоротниково-кисличный с Dryopteris carthusiana и Equisetum sylvaticum и стадию его заболачивания – ельник хвощево-папоротниково-сфагновый со Sphagnum apiculatum H. Lindb. (= S. fallax (Klinggr.) Klinggr. mend. Isov.). В.И. Василевич и Т.В. Бибикова (Vasilevich, Bibikova, 2004b) рассмаривают еловые папоротниковые леса внутри ассоциации ельников кисличников (асс. Oxalido–Piceetum) и выделяют сходную по составу субассоциацию dryopteridetosum carthusianae.
Второй вариант – еловые кочедыжниковые леса (табл. 1, оп. 26–27). В травяно-кустарничковом ярусе доминирует кочедыжник женский (Athyrium filix-femina), местами хвощ лесной. Постоянны, но менее обильны другие виды папоротников (Dryopteris carthusiana, D. expansa, Gymnocarpium dryopteris), кислица, ортилия (Orthilia secunda), некоторые гигромезофиты Viola palustris, Deschampsia cespitosa, Calamagrostis canescens, Crepis paludosa. В моховом покрове обильны Plagiomnium elatum, Rhizomnium pseudopunctatum, Sphagnum girgensohnii, S. centrale.
Для Карельского перешейка отмечена сходная по составу асс. Piceetum mixto-herbosum с большим участием Athyrium filix-femina и других гигромезофитов (Sokolov, 1929). В Скандинавии описан тип Picea abies–Dryopteris ssp. с крупными папоротниками: Athyrium filix-femina, Dryopteris expansa, D. carthusiana (Pählsson, 1994). Для северо-запада Европейской России асс. Athyrio–Piceetum выделена в группе ельников кисличных (Vasilevich, Bibikova, 2004b).
В Северо-Западном Приладожье на озерных глинистых террасах с более застойным увлажнением фрагментарно встречаются ельники таволговые. По схеме типов леса они относятся к еловым таволговым слабодренированным лесам (Fedorchuk et al., 2005). В прошлом большая часть этих ельников была вырублена, а освобожденные территории осушены и использовались под сельскохозяйственные угодья. В пределах исследованной территории были встречены небольшие участки ельников таволгово-папоротниковых. Древостой в них разрежен выборочной рубкой (сомкнутость 0.1), высотой 22–30 м, диаметром стволов до 50 см. Единично отмечены сосна, береза. Есть ели во втором пологе высотой до 10 м. Травяной ярус сомкнутый, проективное покрытие до 70%. Доминируют таволга (Filipendula ulmaria) и папоротники (Athyrium filix-femina, Dryopteris expansa, Gymnocarpium dryopteris). Значительно участие кислицы, майника, видов мезогигрофитного и гигрофитного разнотравья (Equisetum sylvaticum, Viola palustris, Crepis paludosa, Caltha palustris, Chrysosplenium alternifolium, Cirsium oleraceum). Моховой ярус слагают Sphagnum girgensohnii, S. russowii, S. centrale, Brachythecium rivulare, менее обильны Calliergon cordifolium, Rhytidiadelphus triquetrus, Plagiomnium elatum.
Ельники, близкие с описанными выше, приводятся в литературе как ельники приручейные (Vilikaynen, 1957; Mirin, 2003), папоротниково-таволговые или Piceetum filicoso-filipendulosum (Korchagin, Senyaninova-Korchagina, 1957), таволговые (Nitsenko, 1960; Vasilevich, 2004а; Kucherov et al., 2010) и папоротниковые (Volkova et al., 1999; Volkova, 2017; Bibikova et al., 2002). Ельник лог (Piceetum fontinale), выделенный Цинзерлингом (Zinserling, 1932) в группе травяных ельников, имеет сходный характер растительного покрова: на кочках растут бореальные виды, в понижениях – Filipendula ulmaria, Crepis paludosa, Athyrium filix-femina и другие папоротники. В заболоченных долинах ручьев Южной Карелии произрастают близкие по флористическому составу ельники сероольховые кочедыжниково-хвощево-таволговые сфагново-зеленомошные (Kucherov et al., 2006). В Норвегии ельники крупнотравные и крупнопапоротниковые встречаются в сходных местообитаниях – в нижней части склонов и вдоль ручьев, с хорошо развитым мощным почвенным слоем, проточным увлажнением и доступом питательных веществ. Это богатые видами леса с высокой продуктивностью древостоев и травяного яруса. В ельниках крупнопапоротниковых преобладают Athyrium filix-femina, Dryopteris expansa, Oreopteris limbosperma, в крупнотравных они играют гораздо меньшую роль, в них велика роль таволги, Aconitum lycoctonum, Cirsium heterophyllum, Geum rivale и др. (Rekdal, Larson, 2005).
ЕЛОВЫЕ СФАГНОВЫЕ ЛЕСА
Ельники сфагновые – типичная ассоциация для Северо-Запада европейской части России. В Северо-Западном Приладожье описаны две ассоциации еловых сфагновых лесов: чернично-сфагновые и низкобонитетные травяно-сфагновые. В зависимости от степени дренированности и проточности местообитаний в межсельговых понижениях формируются либо еловые леса, либо еловые болота. Ельники сфагновой группы занимают обычно слабовогнутые участки или обводненные понижения с торфянистыми, чаще торфяно-глеевыми почвами, мощность торфа составляет от 10 до 30 см, а в межсельговых ложбинах с более глубоким профилем и по ложбинам стока изредка встречаются еловые травяно-сфагновые болота, которые являются следующей стадией заболачивания еловых лесов (Makarova, Galanina, 2012).
5. Еловые чернично-сфагновые леса (табл. 2, оп. 28–33') встречаются во внутрисельговых слабовогнутых ложбинах с застойным увлажнением и на окультуренных в прошлом слабопроточных озерных террасах. В древостое кроме ели участвует сосна, береза и в меньшей степени осина. Сомкнутость 0.3–0.7, возраст елей от 50–80 до 100 лет (отдельные деревья до 130 лет – оп. 33'). Ель от 17 до 26 м (изредка 30 м) высотой, от 10 до 50 см в диаметре; береза – 17–22 м выс., 15–25 см диам.; сосна – 18–23 м выс., 20–40 см диам. Ель активна и во втором ярусе (8–17 м выс., 8–20 см диам.), и в подросте (1–5%, 0.4–1.5 м выс.). В подлеске обычна рябина, изредка отмечается подрост березы и осины. В кустарниковом ярусе единичны Salix aurita, S. cinerea, Juniperus communis. В травяно-кустарничковом ярусе преобладает Vaccinium myrtillus, в меньшем количестве – Vaccinium vitis-idaea, Carex globularis, Maianthemum bifolium, Trientalis europaea. В моховом покрове доминирует Sphagnum girgensohnii, содоминирует Polytrichum commune, постоянны, но малообильны Hylocomium splendens, Dicranum polysetum, Pleurozium schreberi.
Еловые чернично-сфагновые леса характерны для различных районов Ленинградской обл. (Smirnova, 1928; Nitsenko, 1960; Dyrenkov et al., 1969; Vasilevich, 1983; Volkova et al., 1999, 2007), Карелии (Uskov, 1930; Vilikaynen, 1957, 1971; Gromtsev, 2000; Kutenkov, 2006, 2011). В Финляндии такие сообщества относят к еловым болотам (Eurola et all., 1994).
В ельниках чернично-сфагновых после прореживания выборочными рубками разрастается Carex globularis (покрытие 10% при общем проективном покрытии 20%) и формируется чернично-осочково-сфагновый покров. Это связано с сильным осветлением древостоя ели (сомкнутость до 0.1). Ельники с большим участием Carex globularis были описаны как асс. Piceetum caricoso-sphagnosum (Sukachev, 1931; Zinserling, 1932).
6. Еловые травяно-сфагновые леса представлены низкобонитетными лесами, переходными к болотному типу растительности, которые изредка встречаются в неглубоких межсельговых понижениях Северо-Западного Приладожья.
Был описан вариант ассоциации – сосново-березово-еловые сабельниково-хвощево-сфагновые леса (табл. 2, оп. 34–37'). Они занимают заболачивающиеся глинистые ложбины и слабопроточные понижения (с торфом 10–25 см) между небольшими выположенными сельгами. Древостой характеризуется смешанным составом: ель, береза, в меньшей степени сосна. В зависимости от условий дренажа меняется бонитет ели от III до V класса, преобладают IV–V классы. Возраст елей 40–90 лет. На более дренированных участках высота ели достигает 16–18 м при диаметре стволов 20–25 см, в условиях ухудшения дренажа высота ели – 7–16 м, диаметр – 10–20 (до 28) см. Березы 9–20 м выс., 8–25 см диам. Сосны более мощные: 14–18 (до 21) м выс., 15–35 (до 44) см диам. Ель в подросте растет группами, высотой от 0.5 до 3 м. Из кустарников местами отмечаются ивы (Salix aurita, S. myrsinifolia, S. phylicifolia, S. cinerea) с покрытием 2–10%. Характерными и достаточно обильными видами травяно-кустарничкового яруса являются Comarum palustre, Equisetum sylvaticum, Calamagrostis canescens. Менее обильны Vaccinium vitis-idea, V. myrtillus. Также встречаются Dryopteris carthusiana, Athyrium filix-femina, Viola palustris, Carex nigra, Scirpus sylvaticus. В моховом покрове преобладают Sphagnum girgensohnii, S. centrale, Polytrichum commune, изредка в небольшом обилии отмечаются Sphagnum angustifolium, S. cuspidatum, S. fimbriatum, S. riparium, S. russowii.
В Ленинградской обл. описаны хвощево-сфагновые, осоково-сфагновые и травяно-сфагновые ельники (Smirnova, 1928; Nitsenko, 1960; Dmitrieva, 1973; Vasilevich, 1983; Mirin, 2003; Fedorchuk et al., 2005). В заповеднике “Кивач” в заболоченных межсельговых понижениях с торфяной залежью 40–200 см описаны ельники высоковейниково-хвощевые сфагновые (Kucherov et al., 2006). Цинзерлинг (Zinserling, 1932) выделял травянисто-сфагновый ельник (Piceeto sphagnoso-herbosum) с елью низкого бонитета на средних по богатству почвах. Это крайняя ассоциация в ряду заболоченных еловых лесов. При дальнейшем заболачивании рост ели бывает угнетен, и она начинает вытесняться сосной (Sukachev, 1931), что также было отмечено в заболоченных межсельговых ложбинах Приладожья (Makarova, Galanina, 2012).
СОВРЕМЕННОЕ СОСТОЯНИЕ ЕЛОВЫХ ЛЕСОВ
В настоящей работе приводится карта распространения еловых лесов по состоянию на 2004 г. (рис. 1а), созданная на основе геоботанической карты (Makarova, 2010), и карта распространения еловых лесов по состоянию на 2017 г. (рис. 1б), составленная на основе спутниковых снимков Sentinel-2. За последнее десятилетие исследованная территория была сильно вырублена − сплошными рубками была пройдена почти половина ельников исследованной территории. На картах полосатой штриховкой показаны вырубки, произведенные в течение последних 10 лет, клетчатой штриховкой – вырубки старше 10 лет.
Рис. 1.
Карта распределения еловых лесов на ключевом участке:
(а) по данным геоботанической карты (2004 г.); (б) – по данным дистанционного зондирования (Sentinel-2, 05-2017 г.)
Условные обозначения: 1 – еловые леса; 2 – новые вырубки (1–9 летней давности); 3 – старые вырубки (10–20 летней давности); 4 – гранитодобывающие карьеры; 5 – озера; 6 – дороги.
Fig. 1. The map of the distribution of spruce forests on the key-plot:
(а) according to the data of the geobotanical map (2004); (б) – according to remote sensing data (Sentinel-2, 05‑2017).
Legend: 1 – spruce forests; 2 – new cuttings (1–9 years old); 3 – old cuttings (10–20 years old); 4 – granite quarries; 5 – lakes; 6 – roads.
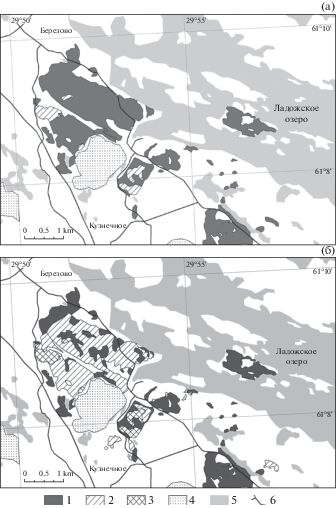
По данным 2004 г. на исследованном ключевом участке площадью 35 км2 лесами было покрыто 22.3 км2. Из них 5.5 км2 занимали ельники: чернично-зеленомошные (64.3%), кислично-зеленомошные (22%), сфагновые (9.4%), травяно-папоротниковые (2.6%), чернично-вейниково-кислично-зеленомошные с неморальнотравными видами (1.7%). Последние два типа встречаются редко, в местах, освоенных ранее под пашни и сенокосы – на озерных террасах и подножиях склонов с дерново-глеевыми глинистыми почвами. После 1940-х гг. сельскохозяйственые работы на многих террасах прекратились, и, на них началось постепенное восстановление еловых лесов, представленное сейчас разными возрастными стадиями: от закустаренных лугов и мелколесий до сосново-мелколиственно-еловых лесов.
По результатам спутниковых снимков Sentinel-2 (05-2017) были выявлены площади новых сплошных рубок, а также площади сохранившихся еловых лесов, проведено сравнение полученных данных с картой 2004 г., вычислены площади сохранившихся ельников. Площади вырубок в 2004 г. составляли 0.4 км2, в 2017 г. – 2.7 км2. Площадь сохранившихся еловых лесов составила 3.1 км2. Летом 2018 г. автором была проведена выборочная полевая верификация результатов дешифрирования проведенных в последние годы сплошных рубок еловых лесов. Верификация проводилась для следующих категорий отдешифрированных вырубок: 1) новые вырубки – без древесного и кустарникового ярусов; 2) вырубки 5–10-летние – с разреженным березовым подростом; 3) вырубки 10–20-летние – с густым березовым молодняком. Выборочная верификация карты 2017 г. показала, что 96% выделенных контуров вырубок были отдешифрированы правильно. Ошибочно отнесены к вырубкам неглубокие узкие ложбины с безлесной заболоченной растительностью.
О ЕЛЬНИКАХ НЕМОРАЛЬНОТРАВНЫХ
Подзональное положение Северо-Западного Приладожья трактовалось по-разному: территория отнесена к средней тайге (Geobotanicheskoe, 1989); к северному рубежу южной тайги (Abramova, Kozlova, 1957; Borisova, 1957; Nitsenko, 1958, 1964; Semenova-Tian-Shanskaya, 1960; Yurkovskaya, Payanskaya-Gvozdeva, 1993); к южной тайге (Hämet-Ahti, 1981). В северо-западном Приладожье кроме среднетаежных, большую роль начинают играть и южнотаежные леса, в том числе и ельники сложные с липой (Kamelin, 2017). На карте восстановленной растительности Карелии пограничная с ключевым участком территория, занята среднетаежными сосновыми лишайниковыми лесами и южнотаежными еловыми лесами с участием неморальных элементов (Yurkovskaya, Elina 2009). По мнению В.П. Денисенкова (Denisenkov, 1995) еловые леса являются коренной формацией в данном районе и по своему видовому составу и производительности относятся к южнотаежным. Критериями для отнесения территории к подзоне южной тайги является: преобладание на суглинистых почвах ельников кисличных, на богатых почвах – ельников неморальнотравных, богатый травяной покров в еловых, березовых и осиновых лесах (Nitsenko, 1958). Согласно указанным критериям, исследованную территорию в Северо-Западном Приладожье можно отнести к подзоне южной тайги. На ключевом участке площадь еловых лесов составляет около 25% от всех лесов, из них четвертую часть занимают ельники кислично-зеленомошные. Также достаточно богат неморальными видами травяной покров в березовых и осиновых лесах (Makarova, 2008, 2009). Распространение ельников кисличников и ельников с неморальными видами вдоль северо-западного побережья Ладожского озера вероятно связано с воздействием самой Ладоги (более мягким климатом в прибрежной части озера по сравнению Карельским перешейком) и с повышенным плодородием суглинистых почв озерных террас.
Согласно Г.А. Елиной (Elina, 1981), на сельгах Приладожья и Заонежья сохранились рефугиумы липы, вяза, дуба, клена, произраставших в атлантическое время (4.9–7.7 тыс. лет назад). Выщелачивание и оподзоливание почв в голоцене являлись одной из причин сокращения ареалов и исчезновения из состава лесов таежной зоны видов неморального флористического комплекса (Grosset, 1967). Но еще в прошлом столетии I. Hiitonen (1946, 1962) отмечал значительное участие более южных видов в прибрежных районах Северо-Западного Приладожья.
По данным мониторинга пробных площадей исследованной территории, проведенных в 1952, 1974, 1991 годы, отмечалось небольшое увеличение доли неморального разнотравья в напочвенном покрове сосновых лесов склонов сельг и мелколиственных лесов озерных террас Северо-Западного Приладожья (Isachenko, Penin, 1995). В еловых лесах такой положительной динамики не выявлено, что вполне естественно, так как Picea abies является активным конкурентом и преобразователем среды. В.Г. Карпов (Karpov, 1973) указывал на то, что ель (а именно, ее корневая система) вытесняет травянистые неморальные виды с местообитаний, где процесс оподзоливания и ухудшения азотного баланса почв еще не достигли своего критического значения для дубравных растений.
Таким образом, участие неморальных и субнеморальных видов в кустарниковом и травяном ярусах в лесах Северо-Западного Приладожья связано с: 1) историческими причинами (внутрисельговые ложбины служили рефугиумами дубравной свиты); 2) более мягким климатом вблизи Ладожского озера (теплая зима, прохладное и дождливое лето); 3) эдафическими особенностями (почвы склонов сельг обогащены органо-минеральными продуктами выветривания, почвы озерных террас сложены ленточными глинами); 4) особенностями рельефа (наличие прогреваемых склонов южной экспозиции); 5) лесообразующей породой (в еловых лесах доля неморального разнотравья будет ниже, чем в мелколиственных и сосновых).
ЗАКЛЮЧЕНИЕ
Приводимая в статье характеристика еловых лесов свидетельствует о том, что ельники ключевого участка относятся к среднетаежным и южнотаежным типам. Доля неморальных и субнеморальных видов невелика, но, тем не менее, ельники с неморальнотравными элементами являются характерной чертой Северо-Западного Приладожья.
В настоящее время еловых лесов на дренированных террасах в пределах ключевого участка не сохранилось, так как в прошлом они были освоены под сельскохозяйственные угодья. После 1940-х годов большинство сельскохозяйственных угодий было заброшено, сейчас они зарастают лесом, находясь на разных стадиях восстановления. При условии отсутствия антропогенных воздействий на террасах уже через 50 лет могут восстановиться ельники, т.к. в настоящее время в ряде мест описаны заключительные стадии – елово-мелколиственные и сосново-мелколиственно-еловые леса.
Напротив, за последнее десятилетие площади еловых массивов на полого-волнистых песчано-валунных и суглинистых террасах ключевого участка сократились почти вдвое за счет массовых сплошных рубок. К ним относится большая часть еловых чернично-зеленомошных, чернично-сфагновых и кислично-зеленомошных лесов. Наиболее сильно пострадала территория, расположенная к северо-западу от ООПТ “Кузнечное”.
Список литературы
[Abramova, Kozlova] Абрамова Т.Г., Козлова Г.И. 1957. Геоботанические районы Северного Приладожья и Карельского перешейка. – Вестник ЛГУ. Сер. Геол., геогр. 24: 152–170.
[Bibikova et al.] Бибикова Т.В., Волкова Е.А., Храмцов В.Н. 2002. Характеристика растительности. – В кн.: Комаровский берег – комплексный памятник природы. СПб. С. 49–66.
[Borisova] Борисова З.В. 1957. О границе между средней и южной тайгой на востоке Ленинградской области. – Бот. журн. 42(8): 1277–1282.
[Bulokhov, Solomeshch] Булохов А.Д., Соломещ А.И. 2003. Эколого-флористическая классификация лесов Южного Нечерноземья России. Брянск. 359 с.
[Cherepanov] Черепанов С.К. 1995. Сосудистые растения России и сопредельных государств. СПб. 992 с.
[Denisenkov] Денисенков В.П. 1995. Современное состояние растительности Приладожской учебно-научной станции. – В кн.: Длительные изменения и современное состояние ландшафтов Приладожья. СПб. С. 27–36.
[Dmitrieva] Дмитриева Е.В. 1973. Ельники северо-западной части Карельского перешейка. – Лесоведение. 2: 51–64.
[Dyrenkov et al.] Дыренков С.А., Адашевская О.Р., Федорчук В.Н. 1969. Ельники Вепсовской возвышенности. – Сб. науч.-иссл. работ по лес. хоз. 12: 102–122.
[Elina] Елина Г.А. 1981. Принципы и методы реконструкции и картирования растительности голоцена. Л. 159 с.
Eurola S., Huttunen A., Kukko-oja K. 1994. Suokasvillisuusopas. – Oulanka Reports. 13. 81 p.
[Fedorchuk et al.] Федорчук В.И., Дыренков С.А., Чертов О.Г., Мельницкая Г.Б., Рябинин Б.Н. 1974. Опыт применения комбинированного метода выделения лесотипологических единиц в северной части Карельского перешейка. – Экология. 6: 49–56.
[Fedorchuk et al.] Федорчук В.Н., Нешатаев В.Ю., Кузнецова М.Л. 2005. Лесные экосистемы Северо-Западных районов России. Типология, динамика, хозяйственные особенности. СПб. 382 с.
[Geobotanicheskoe] Геоботаническое районирование Нечерноземья европейской части РСФСР. 1989. Л. 64 с.
[Gromtsev] Громцев А.Н. 2000. Ландшафтная экология таежных лесов: теоретические и прикладные аспекты. Петрозаводск: КарНЦ РАН. 144 с.
[Grosset] Гроссет Г.Э. 1967. Пути и время миграции лесных крымско-кавказских видов на территорию Русской равнины и последующие изменения их ареалов в связи с эволюцией ландшафтов. – Бюлл. МОИП. 72 (5): 47–76.
Hämet-Ahti L. 1981. The boreal zone and its biotic subdivision. – Fennia. 159 (1): 69–75.
Hiitonen I. 1946. Karjalan kannas kasvien vaellustienä lajien nykylevinneisyyden valossa. – Ann. Bot. Soc. Zool.-Bot. Fenn. Vanamo. 22 (1): 1–206.
Hiitonen I. 1962. Über die natürliche Südostgrenze des östlichen Fennoskandien unter besonderer Berücksichtigung der Karelischen Landenge. – Memoranda Soc. Fauna Fl. Fenn. 37: 13–69.
[Ignatov et al.] Игнатов М.С., Афонина О.М., Игнатова Е.А., Aболиня A.A., Акатова T.В., Баишева Э.З., Бардунов Л.В., Барякина E.A., Белкина O.A., Безгодов A.Г., Бойчук М.А., Черданцева В.Я., Чернядьева И.В., Дорошина Г.Я., Дьяченко A.П., Федосов В.Э., Гольдберг И.Л., Иванова E.И., Юкониeне И., Каннукене Л., Казановский С.Г., Харзинов З.Х., Курбатова Л.Е., Максимов А.И., Маматкулов У.K., Манакян В.A., Масловский O.M., Напреенко M.Г., Отнюкова T.Н., Партыка Л.Я., Писаренко O.Ю., Попова Н.Н., Рыковский Г.Ф., Тубанова Д.Я., Железнова Г.В., Золотов В.И. 2006. Список мхов Восточной Европы и Северной Азии. – Arctoa. 15: 1–130.
[Il’insky] Ильинский А.П. 1921. Материалы к познанию раменей окрестностей Петрограда. – Тр. Петрогр. общ. естествоисп. I–II. 8: 35–65.
[Ipatov et al.] Ипатов В.С., Герасименко Г.Г., Трофимец В.И. 1998. Динамическая классификация сосново-еловых лесов на скалах. – Бот. журн. 82 (8): 13–24.
[Isachenko et al.] Исаченко Г.А., Пенин Р.Л. 1995. Динамика ландшафта за 40 лет. Эколого-флористический анализ. – В кн.: Длительные изменения и современное состояние ландшафтов Приладожья. СПб. С. 36–48.
[Isachenko, Reznikov] Исаченко Г.А., Резников А.И. 2003. Ландшафтная карта Карельского перешейка. М 1:500 000. – В кн.: Природная среда побережья и акватории Финского залива (район порта “Приморск”). СПб. Карта-вкл.
[Kamelin] Камелин Р.В. 2017. Флора Севера Европейской России (в сравнении с близлежащими территориями): Учебное пособие. СПб. 241 с.
[Karpov] Карпов В.Г. 1973. Структура и смены еловых лесов бореального и неморального типов в экспериментальном освещении. – В кн.: Проблемы биогеоценологии, геоботаники и ботанической географии. Л. С. 88–102.
[Kozharinov et al.] Кожаринов А.В., Борисов П.В., Горшкова И.И. 2010. Палеоареал ели европейской (Picea abies (Karst.) L.) на территории Восточной Европы за последние 13500 лет. – Изв. РАН. Сер. геогр. 1: 71–82.
[Korchagin, Senyaninova-Korchagina] Корчагин А.А., Сенянинова-Корчагина М.В. 1957. Леса Молого-Шекснинского междуречья − Тр. Дарвин. зап. 4: 291–402.
[Kutenkov] Кутенков С.А. 2006. Растительность болотных лесов среднетаежной подзоны Карелии. – Болотные экосистемы севера Европы: разнообразие, динамика, углеродный баланс, ресурсы и охрана. – Мат-лы межд. симпоз. Петрозаводск. С. 145–171.
[Kutenkov] Кутенков С.А. 2011. Ельники чернично-сфагновые северотаежной подзоны Карелии. − Развитие геоботаники: история и современность. – Сб. мат-лов конф. СПб. С. 65
[Kucherov et al.] Кучеров И.Б, Филимонова Л.В., Кутенков С.А., Максимов А.И., Максимова Т.А. 2006. Географическая структура лесных ценофлор заповедника “Кивач”. – Тр. Кар. НЦ РАН. Петрозаводск. 10: 71–84.
[Kucherov et al.] Кучеров И.Б., Разумовская А.В., Чуракова Е.Ю. 2010. Еловые леса национального парка “Кенозерский” (Архангельская область). – Бот. журн. 95(9): 1268–1298.
Makarova M.A. 2008. Peculiarities of forest vegetation on the northern border of southern taiga (NW Ladoga region, Russia). – Proceedings of the Intern. scien. conf. “Climate Change and Forest Ecosystems”. October 22–23. Vilnius. Lithuania. P. 91–94.
Makarova M.A. 2009. The natural and regenerated forest communities of lake terraces (NW Ladoga region). – Materials of IX Intern. Conf. of Young Scientists “Eurasian forests – Polish forests”. Moskow – Kórnik. P. 67–70.
Makarova M. 2010. Large-scale mapping of actual vegetation in heterogeneous landscape conditions (NW Ladoga region, Russia). – Conf. proc. “Mapping and monitoring of Nordic vegetation and landscapes”. 115, NO-1431 As, Norway. Norsk institutt for skog og landskap. Viten fra Skog og landskap [Norwegian forest and landscape institute. Forest and landscape research]. 01 (10): 75–79.
Makarova M. 2011. Spruce and pine forests of North-West Ladoga region and large-scale vegetation mapping. – Abstr. of 20 years anniversary symposium of the Finnish-Russian Nature Reserve “Friendship”. 4–7 October 2010. Kuhmo. Finland. P. 38.
[Makarova, Galanina] Макарова М.А., Галанина О.В. 2012. Изучение растительного покрова малых болот Северо-Западного Приладожья для целей крупномасштабного картографирования. – Бот. журн. 97 (1): 44–58.
[Mirin] Мирин Д.М. 2003. Комплексы фитоценозов в долинах ручьев. – Бот. журн. 8(5): 93–111.
[Morozova, Korotkov] Морозова О.В., Коротков В.Н. 1999. Классификация лесной растительности Костомукшского заповедника. – Лесное дело. 5: 56–78.
[Morozova et al.] Морозова О.В., Заугольнова Л.Б., Исаева Л.Г., Костина В.А. 2008. Классификация бореальных лесов севера Европейской России. I. Олиготрофные хвойные леса. – Растительность России. 13: 61–81.
[Morozova et al.] Морозова О.В., Семенищенков Ю.А., Тихонова Е.В., Беляева Н.Г., Кожевникова М.В., Черненькова Т.В. 2017. Неморальнотравные ельники Европейской России. – Растительность России. 31: 33–58.
[Nitsenko] Ниценко А.А. 1958. К вопросу о границе среднетаежной и южнотаежной подзон в пределах Ленинградской области. – Бот. журн. 43 (5): 684–694.
[Nitsenko] Ниценко А.А. 1959. Геоботанический очерк территории Ладожской станции Ленинградского университета. – В кн.: Очерки растительности Ленинградской области. Л. С. 111–135.
[Nitsenko] Ниценко А.А. 1960. Еловые леса Ленинградской области. – Вестник ЛГУ. Сер. биол. 2 (9): 5–16.
[Nitsenko] Ниценко А.А. 1964. Хозяйственно-геоботаническое районирование Ленинградской области. Л. 128 с.
[Obzor …] Обзор санитарного и лесопатологического состояния лесов Ленинградской области в 2009 году и прогноз лесопатологической ситуации на 2010 год. 2010. Фил. ФГУ “Рослесозащита”. Центр защиты леса Ленинградской области.
Pählsson L. 1994. Vegetationstyper i Norden. Stockholm. 627 p.
Rekdal Y., Larsson J.Y. 2005. Veiledning i vegetasjonskartlegging M 1: 20 000–50 000. NIJOS-instruks 1/05. Norsk institutt for jord- og skogkartlegging, Ås. 108 p.
[Rysin, Saveleva] Рысин Л.П., Савельева Л.И. 2002. Еловые леса России. М. 335 с.
[Semenova-Tian-Shanskaya] Семенова-Тян-Шанская А.М. 1960. Картосхема растительности Ленинградской области. – Тр. БИН АН СССР. Сер. III. Геоботаника. 12: 60–75.
[Smirnova] Смирнова З.Н. 1928. Лесные ассоциации северо-западной части Ленинградской области. – Тр. Петергоф. науч.-иссл. ин-та. 5: 119–259.
[Sokolov] Соколов С.Я. 1929. К вопросу о классификации типов еловых лесов. – В кн.: Очерки по фитосоцилогии и фитогеографии. М. С. 205–255.
[Sukachev] Сукачев В.Н. 1931. Руководство к исследованию типов лесов. М.–Л. 328 с.
[Vasilevich] Василевич В.И. 1983. О растительных ассоциациях ельников Северо-Запада. – Бот. журн. 68 (12): 1604–1613.
[Vasilevich] Василевич В.И. 2004а. Травяные ельники европейской России. – Бот. журн. 89 (1): 13–27.
[Vasilevich] Василевич В.И. 2004b. Ельники черничные европейской России. – Бот. журн. 89 (11): 1728–1739.
[Vasilevich] Василевич В. И. 2015. Видовое разнообразие в еловых лесах европейской России. – Бот. журн. 100 (12): 226–236.
[Vasilevich, Bibikova] Василевич В.И., Бибикова Т.В. 2004а. Сфагновые ельники европейской России. – Бот. журн. 89 (5): 734–748.
[Vasilevich, Bibikova] Василевич В.И., Бибикова Т.В. 2004б. Ельники кисличные европейской России. – Бот. журн. 89 (10): 1573–1587.
[Vilikaynen] Виликайнен М.И. 1957. О флоре еловых лесов Карелии. – Тр. Карел. фил. АН СССР. VII: 5–14.
[Vilikaynen] Виликайнен М.И. 1971. Типологическая характеристика объектов исследования. – В сб.: Лесные растительные ресурсы Карелии. С. 7–21.
[Volkova et al.] Волкова Е.А., Галанина О.В., Макарова М.А., Храмцов В. Н. 1999. Очерк растительности района Лужской губы Ленинградской области. – Бот. журн. 84 (12): 21–38.
[Volkova, Khramtsov] Волкова Е.А., Храмцов В.Н. 2003. Характеристика растительности. – В кн.: Природная среда побережья и акватории Финского залива (район порта “Приморск”). СПб. С. 37–44.
[Volkova et al.] Волкова Е.А., Макарова М.А., Храмцов В.Н. 2007. Лесная растительность. – В кн.: Природная среда и биологическое разнообразие архипелага Березовые острова (Финский залив). СПб. С. 70–94.
[Volkova] Волкова Е.А. 2017. Растительность. – В кн.: Природа заказника “Озеро Щучье”. СПб. С. 41–59.
[Uskov] Усков С. 1930. Типы лесов Карелии. Петрозаводск. 87 с.
[Yurkovskaya, Payanskaya-Gvozdeva] Юрковская Т.К., Паянская-Гвоздева И.И. 1993. Широтная дифференциация растительности вдоль Российско-Финляндской границы. – Бот. журн. 78(12): 72–98.
[Yurkovskaya, Elina] Юрковская Т.К., Елина Г.А. 2009. Восстановленная растительность Карелии на геоботанической и палеокартах. Петрозаводск. 136 с.
[Zaugolnova et al.] Заугольнова Л.Б., Смирнова О.В., Браславская Т.Ю., Дегтева С.В., Проказина Т.С., Луговая Д.Л. 2009. Высокотравные таежные леса на востоке европейской части России. – Растительность России. 15: 3–26.
[Zinserling] Цинзерлинг Ю.Д. 1932. География растительного покрова Северо-Запада европейской части СССР. – Тр. Геоморф. ин-та. Сер. физ.-геогр. Вып. 4. 377 с.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал