

Ботанический журнал, 2019, T. 104, № 4, стр. 626-643
ПОЛИМОРФИЗМ ХРОМОСОМ ВИДОВ CHONDRILLA (ASTERACEAE) ЕВРОПЕЙСКОЙ ЧАСТИ АРЕАЛА
А. С. Пархоменко 1, *, А. С. Кашин 1, **, Л. В. Гребенюк 1
1 Саратовский национальный исследовательский государственный университет им. Н. Г. Чернышевского
410012 Саратов, ул. Астраханская, 83, Россия
* E-mail: parkhomenko_as@mail.ru
** E-mail: kashinas2@yandex.ru
Поступила в редакцию 04.02.2019
После доработки 10.03.2019
Принята к публикации 12.03.2019
Аннотация
Приводятся результаты исследований кариотипов растений 26 популяций 8 видов Chondrilla методом рутинной окраски хромосом. По морфометрическим параметрам и визуальному сходству в кариотипах растений каждой популяции выделяли пять пар / групп хромосом. Хромосомы I и V групп в популяциях всех исследованных видов представлены метацентриками, варьирующими лишь по размерам. Хромосомы I группы имели вторичные перетяжки и спутники. По соотношению длин плеч все хромосомы растений C. ambigua (2x = 10) метацентрические. Хромосомы в популяции C. pauciflora (3x = 15) морфологически не отличались от хромосом исследованных популяций C. ambigua. C. laticoronata (3х = 15) отличалась от двух предыдущих видов по морфологии хромосом IV группы: первая в группе – длинный субметацентрик, вторая – акроцентрик средних размеров, а третья – субметацентрик и самая короткая в кариотипе. Две популяции вида C. laticoronata из Астраханской области были полиморфные еще и по морфологии хромосом III группы (мета- или субметацентрики). В популяциях C. brevirostris наблюдался не только внутривидовой, но и внутрипопуляционный полиморфизм по морфологии хромосом III и IV групп. Популяции C. latifolia (3х = 15), C. canescens (3х = 15) и C. juncea (3х = 15) подвержены максимальному варьированию морфологии хромосом III и IV групп на внутри- и межпопуляционном уровне с тем же характером изменчивости, что и у C. brevirostris. В популяциях C. graminea отмечен полиморфизм только по III группе хромосом. Обсуждаются возможные пути генезиса кариотипов исследованных таксонов: от C. ambigua через C. pauciflora к C. laticoronata с последующим радиальным расхождением к остальным слабо дифференцированным между собой таксонам.
ВВЕДЕНИЕ
В роде Chondrilla L. числятся около 30 видов (Ilyin, 1930; Leonova, 1989). В европейской части России естественно встречаются, по крайней мере, семь видов. Шесть из них принадлежат к секции Chondrilla подрода Chondrilla (C. acantholepis Boiss., C. brevirostris Fisch. et Mey, C. canescens Kat. et Kir., C. graminea Bieb., C. juncea L., C. latifolia Bieb. и C. laticoronata Leonova) и один (C. ambigua Fisch.) – к подроду Brachyrynchus (Leonova, 1964). Для Нижнего Поволжья указывался также C. pauciflora Ledeb. (Leonova, 1989), однако, нами он на этой территории не был обнаружен.
В настоящее время нет однозначного представления о таксономической структуре рода. В частности, C. juncea и C. graminea одни авторы рассматривают как самостоятельные виды (Leonova, 1964, 1989; Maevskii, 2014), другие объединяют в один вид C. juncea, включая также C. latifolia, C. canescens и C. brevirostris (Ilyin, 1930; Tutin et al., 1976; Tuzlacı, 2006; Türkçe, 2007; Elenevskii, 2008; Nasseh, Joharchi, 2008; Flann, 2009; Nasseh, 2010).
По результатам морфологического (Kashin et al., 2018) и молекулярно-генетического (Kashin et al., 2017) анализов показано, что из таксонов, указанных для европейской части ареала, лишь C. ambigua хорошо обособлен морфологически и заслуживает признания в ранге вида. Менее очевидна видовая самостоятельность C. brevirostris и C. laticoronata. C. juncea, C. latifolia, C. graminea, C. acantholepis и C. canescens по морфологическим признакам и по ISSR маркерам не разделяются и их следует считать синонимами C. juncea.
Особый интерес в отношении кариотипической изменчивости представляют виды с гаметофитным апомиксисом, у которых изменчивость числа хромосом проявляется не только на меж- и внутрипопуляционном уровнях (Gustafsson, 1946, 1947; Gadella, 1988; Schmelzer, 1997; Mrӓz et al., 2008; Kelley et al., 2009; Cosendai, Hörandl, 2010; Sato, 2011), но и в потомстве отдельных растений и у одного растения (Kashin, 1999, 2000, 2012; Kashin et al., 2000). В отношении изменчивости морфологии хромосом, а также об их числе и плоидности у апомиктичных видов Chondrilla имеются очень ограниченные сведения (Poddubnaya-Arnoldi, 1933; Bergman, 1952; Parkhomenko, Kashin, 2018), а сравнение морфологии хромосом на видовом уровне ранее не проводилось.
Цель данного исследования состояла в анализе изменчивости морфологии хромосом видов рода Chondrilla европейской части ареала.
МАТЕРИАЛЫ И МЕТОДЫ
Семянки для исследования собраны в 2015–2017 гг. в 26 популяциях 8 видов Chondrilla (C. ambigua, C. brevirostris, C. canescens, C. graminea, C. juncea, C. laticoronata, C. latifolia и C. pauciflora) из Астраханской, Волгоградской, Воронежской, Ростовской и Саратовской областей, Краснодарского края, Республик Крым и Калмыкия, а также Западного Казахстана (табл.).
Таблица 1.
Места сбора популяций видов рода Chondrilla Table 1. Collection sites of populations of the genus Chondrilla species
| Усл. №
популяции Population number |
Место произрастания/Locality |
|---|---|
| C. ambigua | |
| 30 | Астраханская обл., Красноярский р-н, окр. п. Досанг (46°54'56.1'' N, 47°55'07.6''E) Astrakhan Region, Krasnoyarsky District, environs of Dosang village (46°54'56.1'' N, 47°55'07.6''E) |
| 31 | Р. Калмыкия, Яшкульский р-н, окр. п. Хулхута (46°17'38.8''N, 46°41'55.6''E) Republic of Kalmykia, Yashkul District, environs of Khulkhuta village (46°17'38.8''N, 46°41'55.6''E) |
| C. brevirostris | |
| 29 | Астраханская обл., Ахтубинский р-н, окр. с. Болхуны (47°59'17.8''N, 46°25'52.6''E) Astrakhan Region, Akhtubinsk District, environs of Bolhkuny village (47°59'17.8''N, 46°25'52.6''E) |
| 46 | Астраханская обл., Харабалинский р-н, окр. с. Вольное (47°08'07.5''N, 47°42'12.4''E) Astrakhan Region, Kharabali District, environs of Volnoye village (47°08'07.5''N, 47°42'12.4''E) |
| 55 | Астраханская обл., Ахтубинскй р-н, окр. пос. Верблюжий (47°43'32.2''N, 46°52'37.9''E) Astrakhan Region, Akhtubinsk District, environs of Verblyuzhii village (47°43'32.2''N, 46°52'37.9''E) |
| 58 | Астраханская обл., Красноярский р-н, окр. с. Досанг (46°54'56.1''N, 47°55'07.6''E) Astrakhan Region, Krasnoyarsky District, environs of Dosang village (46°54'56.1''N, 47°55'07.6''E) |
| 59 | Р. Калмыкия, Яшкульский р-н, окр. пос. Хулхута (46°17'38.8''N, 46°41'55.6''E) Republic of Kalmykia, Yashkul District, environs of Khulkhuta village (46°17'38.8''N, 46°41'55.6''E) |
| 77 | Казахстан, Бокейординский р-н, окр. с. Жиеккум (48°50'53.2''N, 47°49'29.5''E) Kazakhstan, Bokeyorda District, environs of Zhiyekkum village (48°50'53.2''N, 47°49'29.5''E) |
| C. canescens | |
| 40 | Саратовская обл., Хвалынский р-н., гора Беленькая (52°28'44.2''N, 48°03'32.6''E) Saratov Region, Khvalynsk District, mountain Belenkaya (52°28'44.2''N, 48°03'32.6''E) |
| 54 | Астраханская обл., Ахтубинский р-н, окр. ж/д ст. Капустин Яр (48°34'03.5''N, 45°45'57.6''E) Astrakhan Region, Akhtubinsk District, environs of Kapustin Yar railway station (48°34'03.5''N, 45°45'57.6''E) |
| 74 | Воронежская обл., Богучаровский р-н, окр. с. Терешково (49°54'38.4''N, 40°40'28.8''E) Voronezh Region, Bogucharovsky District, environs of Tereshkovo village (49°54'38.4''N, 40°40'28.8''E) |
| C. graminea | |
| 42 | Саратовская обл., Хвалынский р-н., гора Беленькая (52°28'44.2''N, 48°03'32.6''E) Saratov Region, Khvalynsk District, mountain Belenkaya (52°28'44.2''N, 48°03'32.6''E) |
| 45 | Саратовская обл., Базарно-Карабулакский р-н., окр. с. Алексеевка (52°13'49.3''N, 46°19'05.9''E) Saratov Region, Bazarno-Karabulaksky District, environs of Alekseyevka village (52°13'49.3''N, 46°19'05.9''E) |
| C. juncea | |
| 19 | Краснодарский край, Ейский р-н, окр. ст. Должанская (46°39'35.7''N, 37°46'36.0''E) Krasnodar Territory, environs of Yeisk town, Dolzhanskaya station (46°39'35.7''N, 37°46'36.0''E) |
| 27 | Саратовская обл., Краснокутский р-н, окр. с. Дьяковка (50°43'07.3''N, 46°46'52.0''E) Saratov Region, Krasnyi Kut District, environs of Dyakovka village (50°43'07.3''N, 46°46'52.0''E) |
| 34 | Саратовская обл., Аткарский р-н, окр. с. Приречное (51°44'47.3''N, 44°55'44.9''E) Saratov Region, Atkarsky District, environs of Prirechnoye village (51°44'47.3''N, 44°55'44.9''E) |
| 48 | Ростовская обл., Константиновский р-н, окр. с. Базки (47°47'05.6''N, 41°02'46.7''E) Rostov Region, environs of Bazki khutor (47°47'05.6''N, 41°02'46.7''E) |
| 50 | Р. Крым, окр. г. Севастополя (44°36'51.8''N, 33°31'29.9''E) Republic of Crimea, environs of Sevastopol city (44°36'51.8''N, 33°31'29.9''E) |
| C. laticoronata | |
| 56 | Астраханская обл., Ахтубинский р-н, окр. п. Верблюжий (47°43'32.2''N, 46°52'37.9''E) Astrakhan Region, Akhtubinsk District, environs of Verblyuzhii village (47°43'32.2''N, 46°52'37.9''E) |
| 70 | Астраханская обл., Харабалинский р-н, окр. с. Сасыколи (47°33'04.7''N, 46°58'55.9''E) Astrakhan Region, Kharabali District, environs of Sasykoli village (47°33'04.7''N, 46°58'55.9''E) |
| 71 | Астраханская обл., Харабалинский р-н, окр. с. Тамбовка (47°19'02.0''N, 47°22'20.3''E) Astrakhan Region, Kharabali District, environs of Tambovka village (47°19'02.0''N, 47°22'20.3''E) |
| 76 | Казахстан, Бокейординский р-н, окр. с. Жиеккум (48°51'42.2''N, 47°48'49.2''E) Kazakhstan, Bokeyorda District, environs of Zhiyekkum village (48°51'42.2''N, 47°48'49.2''E) |
| C. latifolia | |
| 25 | Волгоградская обл., окр. г. Калач-на-Дону (48°43'04.2''N, 43°30'67.9''E) Volgograd Region, environs of Kalach-na-Donu town (48°43'04.2''N, 43°30'67.9''E) |
| 60 | Ростовская обл., Константиновский р-н, окр. х. Базки (47°47'05.6''N, 41°02'46.7''E) Rostov Region, Konstantinovsky District, environs of Bazki khutor (47°47'05.6''N, 41°02'46.7''E) |
| 61 | Волгоградская обл., окр. г. Камышин (50°08'05.9''N, 45°26'14.9''E) Volgograd Region, environs of Kamyshin city (50°08'05.9''N, 45°26'14.9''E) |
| C. pauciflora | |
| 78 | Казахстан, Абайский р-н, окр. с. Орда (48°49'10.8''N, 47°30'17.5''E) Kazakhstan, Abay District, environs of Orda village (48°49'10.8''N, 47°30'17.5''E) |
Для анализа морфологии метафазных хромосом использовали клетки апикальных меристем корешков проростков. По достижении корешками длины 10–12 мм их обрабатывали водным раствором бромнафталина в течение 3.5 часов при t° = 4°С, затем фиксировали в фиксаторе Кларка в течение суток. Окрашивали в ацетогематоксилине и готовили временные давленые препараты в 80%-ном растворе хлоралгидрата. Материал анализировали под масляной иммерсией, используя микроскоп Carl Zeiss Axio Scope A1 (×1600). Микрофотографирование производили с использованием цифровой камеры AxioCam MRc 5 (D) и адаптера 60N-C 1'' 1.0x. На каждом препарате в 10 и более различных клетках изучали морфометрические показатели (локализацию вторичной перетяжки, абсолютную и относительную длину, центромерный индекс, индекс спирализации и общую длину генома), подсчитывали число хромосом и определяли их тип. В кариотипах диплоидов, на основании морфометрических показателей и визуального сходства, выделяли 5 пар хромосом. В случае кариотипов триплоидных и тетраплоидных особей, вместо термина “пара” мы употребляли “группа” хромосом. Пары (группы) хромосом обозначались римскими цифрами от I до V (Grif, Agapova, 1986). При обнаружении изменчивости хромосом по морфологии в пределах одной группы каждая из хромосом обозначалась арабскими цифрами от 1 до 3.
РЕЗУЛЬТАТЫ
Chondrilla ambigua. Соматическое число хромосом в двух исследованных популяциях из Астраханской обл. равно 10 (рис. 1b). Метод построения поликариограмм выявил сходный для обеих популяций характер распределения точек по парам хромосом (рис. 2a). Хромосомы I–III пар образовывали одно общее облако, в то время как хромосомы IV–V пар – второе. За счет сходного центромерного индекса хромосом на поликариограмме эти облака располагались друг над другом. При этом облако, образованное I–III парами, за счет большей относительной длины составляющих их хромосом, располагалось на поликариограмме выше облака, образованного IV–V парами (рис. 2а). По соотношению длин плеч все хромосомы C. ambigua равноплечие (рис. 3а, b). При этом первая пара хромосом отличалась наличием в дистальном районе вторичной перетяжки (Sc = 76.3±2.56%). Вторая и третья пары были близки по длине к первой, но не содержали вторичных перетяжек. Четвертую пару составили хромосомы, имеющие длину хромосом в 1.5 раза короче, чем хромосомы первых трех пар, а пятая пара была самой короткой. В целом, морфология хромосом в обеих популяциях C. ambigua очень сходна, что, скорее всего, является типичным для всего вида.
Рис. 1.
Примеры метафазных пластинок изученных видов рода Chondrilla: a – C. juncea (4х); b – C. ambigua (2х); c – C. brevirostris (3х-1); d – C. pauciflora (3х); e – C. canescens (3х); f – C. laticoronata (3х).
Fig. 1. Examples of metaphase plates of the studied Chondrilla species: a – C. juncea (4х); b – C. ambigua (2х); c – C. brevirostris (3х-1); d – C. pauciflora (3х); e – C. canescens (3х); f – C. laticoronata (3х).
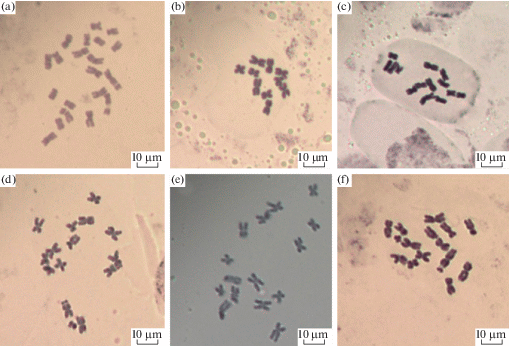
Рис. 2.
Поликариограммы популяций Chondrilla. По оси абсцисс – центромерный индекс (Ic), %; по оси ординат – относительная длина (Lr), %. I-V – номера групп хромосом: a – C. ambigua (№ 30 и 31); b – C. pauciflora (№ 78); c – C. laticoronata (№ 56, 70, 71 и 76); d – C. brevirostris (№ 29, 46 и 55), C. juncea (№ 27), C. latifolia (№ 60); e – C. brevirostris (№ 58); f – C. brevirostris (№ 59); g – C. brevirostris (№ 77); h – C. canescens (№ 40); i – C. canescens (№ 54), C. juncea (№ 48), C. latifolia (№ 25); j – C. canescens (№ 74); k – C. juncea (№ 19); l – C. juncea (№ 34 и 50), C. graminea (№ 42 и 45), C. latifolia (№ 61).
Fig. 2. Polycaryograms of Chondrilla populations. X-axis – centromeric index (Ic),%; Y-axis – relative length (Lr),%. I–V – numbers of chromosome groups: a – C. ambigua (№ 30 and 31); b – C. pauciflora (№ 78); c – C. laticoronata (№ 56, 70, 71 and 76); d – C. brevirostris (№ 29, 46 and 55), C. juncea (№ 27), C. latifolia (№ 60); e – C. brevirostris (№ 58); f – C. brevirostris (№ 59); g – C. brevirostris (№ 77); h – C. canescens (№ 40); i – C. canescens (№ 54), C. juncea (№ 48), C. latifolia (№ 25); j – C. canescens (№ 74); k – C. juncea (№ 19); l – C. juncea (№ 34 and 50), C. graminea (№ 42 and 45), C. latifolia (№ 61).
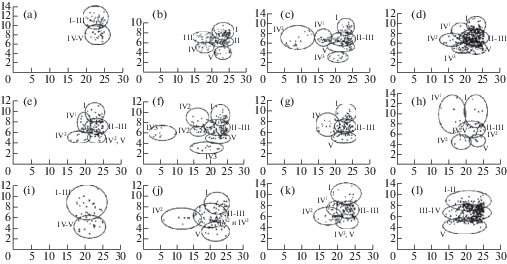
Рис. 3.
Систематизированные кариотипы популяций подрода Brachyrhynchus: a – C. ambigua (№ 30); b – C. ambigua (№ 31); c – C. pauciflora (№ 78).
Fig. 3. Systematized karyotypes of the subgenus Brachyrhynchus: a – C. ambigua (№ 30); b – C. ambigua (№ 31); c – C. pauciflora (№ 78).
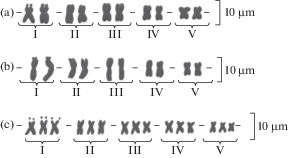
Chondrilla pauciflora. Число хромосом в соматических клетках исследованных образцов 3x = 15. Средняя суммарная абсолютная длина хромосомного набора за счет три- плоидности на 20% больше соответствующей величины по популяциям C. ambigua. В целом хромосомы растений C. pauciflora морфологически не отличались от исследованных популяций C. ambigua. На поликариограмме выделяется пять областей точек, соответствующих каждой группе хромосом (рис. 2b).
Chondrilla laticoronata. Соматическое число хромосом всех иследованных образцов равно 3х = 15. На поликариограмме области точек, соответствующие II и III группам хромосом, не разделяются (рис. 2c). Выше этой области находится область точек, соответствующая хромосомам I, а ниже – хромосомам V группы. Все эти области слабо разделяются. Каждая из хромосом IV группы образует собственные области хорошо обособленных друг от друга и от облаков точек, соответствующих хромосомам других групп. Морфология и размеры первых двух групп хромосом C. laticoronata были сходными с таковыми у других изученных видов Chondrilla (рис. 3, 4а, b).
Рис. 4.
Систематизированные кариотипы исследованных популяций подрода Chondrilla: a – C. laticoronata (№ 56, 70, 71 и 76); b – C. laticoronata (№ 56 и 70); c – C. brevirostris (№ 29, 46, 55, 58 и 77), C. canescens (№ 40, 54 и 74), C. juncea (№ 19, 27 и 50), C. graminea (№ 42 и 45), C. latifolia (№ 25, 60 и 61); d – C. canescens (№ 40, 54 и 74), C. brevirostris (№ 46 и 55), C. juncea (№ 19, 27, 34, 48 и 50), C. graminea (№ 42 и 45), C. latifolia (№ 60 и 61); e – C. canescens (№ 74), C. juncea (№ 27), C. latifolia (№ 60); f – C. canescens (№ 74), C. latifolia (№ 60); g – C. brevirostris (№ 29, 46, 55, 58 и 59), C. canescens (№ 40 и 54), C. latifolia (№ 60 и 61); h – C. juncea (№ 27); i – C. brevirostris (№ 59), C. juncea (№ 19 и 27); j – C. brevirostris (№ 59); k – C. canescens (№ 40 и 54); l – C. canescens (№ 40); m – C. brevirostris (№ 29); n – C. canescens (№ 74).
Fig. 4. Systematized karyotypes of the subgenus Chondrilla: a – C. laticoronata (№ 56, 70, 71 and 76); b – C. laticoronata (№ 56 and 70); c – C. brevirostris (№ 29, 46, 55, 58 and 77), C. canescens (№ 40, 54 and 74), C. juncea (№ 19, 27 and 50), C. graminea (№ 42 and 45), C. latifolia (№ 25, 60 and 61); d – C. canescens (№ 40, 54 and 74), C. brevirostris (№ 46 and 55), C. juncea (№ 19, 27, 34, 48 and 50), C. graminea (№ 42 and 45), C. latifolia (№ 60 and 61); e – C. canescens (№ 74), C. juncea (№ 27), C. latifolia (№ 60); f – C. canescens (№ 74), C. latifolia (№ 60); g – C. brevirostris (№ 29, 46, 55, 58 and 59), C. canescens (№ 40 and 54), C. latifolia (№ 60 and 61); h – C. juncea (№ 27); i – C. brevirostris (№ 59), C. juncea (№ 19 and 27); j – C. brevirostris (№ 59); k – C. canescens (№ 40 and 54); l – C. canescens (№ 40); m – C. brevirostris (№ 29); n – C. canescens (№ 74).
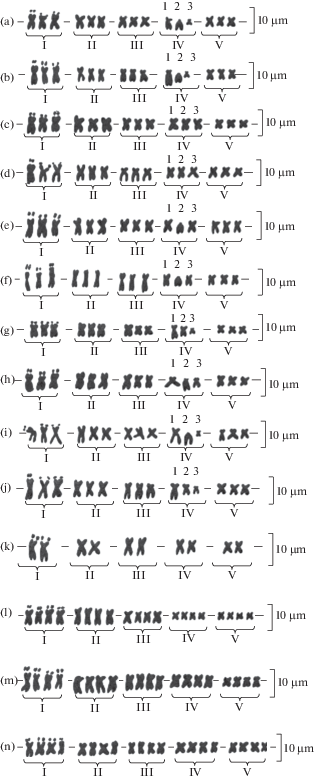
Третья группа хромосом в кариотипах популяций 71 и 76 состояла из метацентриков. В популяции 56 эта группа хромосом в кариотипах 52% исследованных растений состояла из метацентриков, у 48% особей – из субметацентриков, а в популяции 70 в кариотипах 30% растений – из метацентриков, у 70% особей – из субметацентриков (рис. 4а, b). При этом полиморфизм хромосом наблюдался даже в соседних клетках одного апекса. Однако если выявленная перестройка наблюдалась в кариотипе, то охватывала все хромосомы группы (находилась в гомозиготном состоянии).
Четвертая группа во всех четырех исследованных популяциях C. laticoronata в кариотипах всех особей состояла из трех хромосом, значительно отличающихся между собой по морфологическим параметрам. Первая хромосома в этой группе характеризовалась большими размерами и являлась субметацентриком. Вторая была акроцентриком средних размеров. Третья хромосома была субметацентриком и самой короткой в кариотипе.
В кариотипах растений популяций C. laticoronata, помимо мутаций, встречались единичные случаи выпадения хромосом: в популяции 56 отмечен случай нехватки одной хромосомы в V группе, в 71 – в III группе, а в 76 – во II группе. В популяции 70 на одной метафазной пластинке не хватало двух хромосом в I группе и двух хромосом в V группе, а на двух метафазных пластинках отсутствовала акроцентрическая хромосома в IV группе.
Chondrilla brevirostris. Соматическое число хромосом 3x = 15 (рис. 1c), но в популяции 29 была отмечена одна тетраплоидная особь с добавочной хромосомой в четвертой группе (4x + 1 = 21).
На поликариограммах все исследованные популяции C. brevirostris имеют аналогичное распределение областей точек, соответствующих разным группам хромосом, в том числе и каждой хромосоме IV группы (рис. 2d–g). Все облака точек слабо обособлены, а в ряде случаев и вовсе образуют единое рыхлое облако. Только в популяции 59 имеется область точек, хорошо обособленная от остальных и соответствующая второй хромосоме IV группы (рис. 2f). В то же время этой хромосоме соответствует еще одно, но уже плохо обособленное от остальных облако. Такое распределение точек на поликариограмме объясняется двумя вариантами морфологического строения этой хромосомы, один из которых, скорее всего, возник в результате мутации.
Морфология хромосом I, II и V групп у данного вида идентична таковым у предыдущих таксонов (рис. 3, 4).
В кариотипе популяции 77 все хромосомы были метацентрическими, как и в части кариотипов популяций 29, 46, 55 и 58 (рис. 4c).
У тетраплоидной особи в популяции 29 хромосомы I, II, IV и V групп также были метацентриками (рис. 4e). В то же время хромосомы III группы в кариотипе этой особи были субметацентриками, что может быть следствием внутрихромосомных перестроек. В кариотипах популяций 46, 55 и 59 также наблюдался полиморфизм III группы хромосом (рис. 4c, d, g, i). На большинстве метафазных пластинок (83%) эта группа была представлена метацентриками, но в 17% – субметацентриками.
В кариотипах C. brevirostris в IV группу входили хромосомы как сходные по морфологическим параметрам, так и весьма различающиеся. На метафазных пластинках 40–70% особей из популяций 29, 46, 55 и 58 эта группа состояла из метацентрических хромосом (рис. 4c, g). Встречались кариотипы, где первая и третья хромосомы в IV группе являлись субметацентриками (рис. 4b), и в 1.5–2 раза отличались друг от друга по длине. В популяции 59 такая разница в длине между этими хромосомами носила постоянный характер. Кроме того, здесь наблюдался полиморфизм по морфологии второй хромосомы в IV группе. На 17% метафазных пластинок эта хромосома была акроцентрической (рис. 4b, i), а в остальных случаях – метацентрической (рис. 4g). Таким образом, кариотипы особей C. brevirostris, которые содержали акроцентрическую хромосому, были очень похожи на кариотипы C. laticoronata, что, скорее всего, объясняется аналогичными хромосомными перестройками и свидетельствует о близкородственности этих таксонов.
В популяциях 29, 46, 55, 58 и 77 были отмечены случаи миксоплоидии, связанные с потерей хромосом. Так, в популяциях 29 и 58 на одной метафазной пластинке не хватало второй хромосомы в IV группе. В популяции 46 в одном случае отсутствовала хромосома в V группе, а в другом – не хватало сразу двух хромосом (во II и в IV группах). В популяции 55 отмечено 3 метафазных пластинки с недостающими хромосомами: на одной – не хватало второй хромосомы в IV группе, на другой – отсутствовали сразу три хромосомы – одна в I и две в V группе, на третьей – не было одной хромосомы в I группе. В популяции 77 на одной метафазной пластинке во II группе было только две хромосомы, а V группа содержала только одну хромосому (рис. 1).
Очевидно, что для C. brevirostris характерна не только межпопуляционная, но и внутрипопуляционная изменчивость морфологии хромосом. Кариотип популяции 77 морфологически аналогичен или близок кариотипу популяции 78 (C. pauciflora), а некоторые кариотипы особей в популяции 59 имеют общие черты строения с исследованными кариотипами популяций C. laticoronata. Таким образом, C. brevirostris, скорее всего, является эволюционно более молодой, чем C. pauciflora и C. laticoronata, либо является следствием отдаленной гибридизации между ними или близкими к ним таксонами. Самыми консервативными в отношении морфологии, как и у C. laticoronata, были хромосомы I, II и V групп, самой полиморфной – IV группа.
Chondrilla canescens. Соматическое число хромосом равно 15, хотя были обнаружены особи с набором хромосом 2x = 10 и 4x = 20. На поликариограмме первая и третья хромосомы IV группы в популяции 40 формируют обособленные области (рис. 2h), а в популяции 74 отдельная область точек соответствует второй хромосоме IV группы (рис. 2j), что связано со структурными перестройками, произошедшими в этих хромосомах.
Все хромосомы диплоида в популяции 40 были метацентрическими (рис. 4c). Однако у тетраплоидов в популяции 74 все хромосомы II группы, а в кариотипе одного из триплоидов, по крайней мере, одна хромосома этой группы были представлены субметацентриками (рис. 4m). Это обстоятельство может указывать на то, что кариотип этих особей является следствием межвидовой гибридизации, в том числе и с одновременным увеличением числа гаплоидных наборов хромосом. Характерно, что в популяции 74 тетраплоиды имели примерно в 1.5 раза меньшую длину хромосом внутри групп, чем триплоиды.
Популяции 40, 54 и 74 были полиморфны по III группе хромосом. На 30% метафазных пластинок хромосомы этой группы были субметацентриками (рис. 4d, f), на остальных – метацентриками (рис. 4c, e, g).
В популяциях 40 и 54 у трети особей наблюдался полиморфизм и по IV группе хромосом (рис. 4b, k), аналогичный соответствующим отклонениям в морфологии хромосом этой группы у C. laticoronata и C. brevirostris. При этом отклоняющиеся в морфологии первая и третья хромосомы IV группы в соответствующих кариотипах популяций 40 и 54 встречались совместно в одном кариотипе, в то время как акроцентрическая вторая хромосома IV группы, обнаруженная в кариотипе некоторых растений популяции 74, была в одной группе с метацентриками, а не с субметацентриками. То есть, у большей части особей популяции 74 в IV группу входили метацентрики, а у 1/4 части эта группа отличалась присутствием акроцентрической хромосомы.
Таким образом, у C. canescens отклонения в морфологии 1–3 хромосом IV группы, никогда не встречались не только в одном кариотипе, но и в генофонде одной популяции. При этом отклоняющиеся формы хромосом IV группы всегда обнаруживались в кариотипах, где хромосомы III группы – субметацентрики. Лишь в одном кариотипе III группу образуют субметацентрические хромосомы, в то время как в IV группу хромосом входят метацентрики. Все это также указывает на возможную гибридную природу указанных кариотипов таксона.
Все популяции этого вида по морфологии хромосом являются полиморфными, но в особенно сильной степени полиморфизм представлен в популяции 74 из Воронежской обл., т.е. самой северо-западной из исследованных популяций.
Chondrilla juncea. Соматическое число хромосом равно 15, но обнаружены две особи (в популяции 27) с набором хромосом 4x = 20 (рис. 1a).
Все хромосомы C. juncea в популяции 48 (рис. 4c) и хромосомы тетраплоида в популяции 27 (рис. 4n) были метацентрическими. Поликариограммы таксона в некоторых случаях оказались сходными с поликариограммами других таксонов (рис. 2d, i, k, e). Точки на поликариограмме, соответствующие хромосомам IV группы, образуют отдельно расположенные, но перекрывающиеся области. Это говорит о вариативности морфологии хромосом, входящих в данную группу (рис. 2d, j, k).
Хромосомы I, II и V групп были морфологически сходными с таковыми у других видов.
В популяциях 19, 27, 34 и 50 наблюдался внутрипопуляционный полиморфизм по III группе хромосом (рис. 4c, d). Она была представлена субметацентриками в популяции 34 у 42%, в популяции 27 – у 31%, в популяции 50 – у 29%, а в популяции 19 – у 23% растений. В остальных случаях данную группу составляли метацентрики.
Полиморфизм по IV группе хромосом отмечен в популяциях 19 и 27. Выявлены следующие варианты: 1) все три хромосомы метацентрические (в популяции 19 – 80%, в популяции 27 – 37.5%) (рис. 4c); 2) первая хромосома этой группы – длинный субметацентрик, вторая хромосома – метацентрик средних размеров, а третья – субметацентрик и самая короткая в кариотипе (в популяции 27 – 56.3%) (рис. 4g); 3) первая и третья хромосомы такие же, как и в предыдущем варианте, вторая – акроцентрик средних размеров (в популяции 19 – 20%, в популяции 27 – 37.6%) (рис. 4h).
Помимо полиморфизма в строении хромосом в популяциях 19, 34, 48 и 50 отмечены случаи миксоплоидии. В этих популяциях встречались единичные случаи (1–4 метафазные пластинки) нехватки одной, реже двух хромосом в разных группах.
Chondrilla graminea. 3x = 15. Поликариограммы популяций таксона аналогичны поликариограммам части популяций C. juncea и C. latifolia (рис. 2e). Первая, вторая, и пятая группы хромосом имели типичную для таксонов рода морфологию. Исключение составила одна метафазная пластинка в популяции 42, где во II группу входили суб-метацентрики (как у тетраплоида C. canescens в популяции 74). Четвертая группа образована метацентриками.
По III группе хромосом в изученных популяциях наблюдался внутрипопуляционный полиморфизм (рис. 4c, d). В популяции 42 эта группа состояла из субметацентриков на 60% метафазных пластинок, а в популяции 45 – на 50%. В остальных случаях III группа была образована только метацентриками.
В обеих исследованных популяциях данного таксона отмечены единичные случаи отсутствия одной хромосомы во II, III или IV группе. В популяции 45 на двух метафазных пластинках, а в популяции 42 – на одной не хватало одной хромосомы в V группе. В популяции 45 был отмечен случай потери одной хромосомы сразу в двух группах (во II и в V), а в популяции 42 на одной метафазной пластинке не хватало трех хромосом (по одной в каждой из следующих групп: III, IV и V).
Chondrilla latifolia. Соматическое число хромосом равно 15. Поликариограммы C. latifolia оказались аналогичны поликариограммам части популяций C. brevirostris (рис. 2d), C. canescens (рис. 2i), C. juncea (рис. 2d, i, e) и C. graminea (рис. 2e).
Кариотип популяции 25 состоял из метацентрических хромосом (рис. 4c). В других двух популяциях (60 и 61) такого единообразия в морфологии хромосом не было. Группы хромосом I, II и V вида C. latifolia имели типичную для всего рода морфологию.
Популяции 60 и 61 были полиморфны по III группе хромосом. На 44.7% метафазных пластинок в популяции 61 и на 12.5% – в популяции 60 эту группу составляли субметацентрики (рис. 4d). В популяции 60 у 30% особей наблюдался полиморфизм и по IV группе хромосом (рис. 4e, g). Так, в 8% случаев первая хромосома IV группы была длинным субметацентриком, вторая хромосома этой группы – метацентриком средних размеров, а третья хромосома – субметацентриком и самой короткой в кариотипе (рис. 4g). У остальных 22% особей первая и третья хромосомы IV группы были метацентриками средних размеров, в то время как вторая хромосома этой группы являлась акроцентрической (рис. 4e).
Во всех исследованных популяциях таксона отмечены случаи миксоплоидии, связанные с потерей хромосом. Встречались единичные метафазные пластинки, на которых отсутствовали одна или две хромосомы в разных группах. В популяциях 25 и 60 отмечено по одной метафазной пластинке, на которых не хватало сразу четырех хромосом: в популяции 25 – одной в I, двух в IV, и одной в V группах; в популяции 60 – одной во II, одной в IV, и двух в V группах.
ОБСУЖДЕНИЕ
Исследованные виды типового подрода оказались полиморфными по морфологии хромосом, в отличие от C. ambigua подрода Brachyrhynchus. Виды подрода Chondrilla отличаются друг от друга только уровнем плоидности и размерами хромосом. Скорее всего, C. pauciflora является триплоидным цитотипом C. ambigua. Он и морфологически близок к C. ambigua, по большинству признаков перекрываясь с ней (Leonova, 1989).
В кариотипах представителей подрода Chondrilla полиморфизм в различной степени охватывает хромосомы трех групп. Наиболее полиморфна IV группа, менее полиморфна – III, и менее всего полиморфна – II группа хромосом. По I и V группам хромосом полиморфизм не отмечен.
Во II группе хромосом полиморфизм отмечен у C. canescens (популяция 74), который выражался в том, что у тетраплоидов – все, а в кариотипе одного из триплоидов – одна из хромосом группы были субметацентрическими, в то время как у остальных особей все хромосомы этой группы были метацентрическими. У C. graminea (популяция 42) подобный характер полиморфизма наблюдался у одной триплоидной особи.
Полиморфизм по III группе хромосом (мета- или субметацентрики), отмечен в части популяций всех исследованных таксонов подрода Chondrilla, но был представлен в них с различной частотой. В двух из 4-х популяций C. laticoronata субметацентриков было 50–70%, в четырех из 6-ти популяций C. brevirostris – только около 20%, в трех из пяти популяций C. сanescens – около 30%, в четырех из пяти популяций C. juncea – от 23 до 42%, в обеих исследованных популяциях C. graminea – 50–60%, а в двух из трех популяций C. latifolia – от 12.5 до 44.7%.
IV группа хромосом во всех четырех исследованных популяциях C. laticoronata состояла из трех хромосом, значительно отличающихся между собой по морфологическим параметрам, но различий между особями и популяциями по морфологии хромосом этой группы не выявлено. В пределах кариотипа любой особи первая хромосома в этой группе являлась субметацентриком и характеризовалась большими размерами, вторая хромосома – акроцентрик средних размеров, а третья – субметацентрик и самая короткая в кариотипе.
По-видимому, такое различие в морфологии хромосом данной группы связано с внутри- и/или межхромосомными перестройками. Вероятно, изначально все хромосомы группы имели одинаковую длину и являлись метацентриками. Затем произошел перенос части плеча третьей хромосомы на плечо первой хромосомы (транслокация), в результате чего образовались две субметацентрические хромосомы: длинная – первая и короткая – третья. Во второй хромосоме этой группы, скорее всего, произошла перицентрическая инверсия (захватывающая центромерный участок), в результате чего эта хромосома стала акроцентрической.
Ранее у видов Chondrilla подобного рода хромосомные перестройки описаны не были. Безусловно, без дополнительных молекулярно-цитогенетических исследований с использованием методов дифференциального окрашивания и in situ гибридизации (FISH, GISH) нет оснований однозначно утверждать, в результате каких именно перестроек могли произойти выявленные изменения в морфологии хромосом. Показано, что сдвиг центромеры может происходить вследствие перицентрической инверсии или транслокации, которые являются распространенными сбалансированными перестройками у эукариот (Amosova et al., 2009; Bugrov, 2010). Считается, что парацентрические и перицентрические инверсии, реципрокные, нереципрокные, центрические и тандемные транслокации, являются основными типами хромосомных мутаций, лежащих в основе эволюции кариотипов. При этом именно инверсии представляют собой один из основных цитогенетических механизмов преобразования генома эукариот (Gileva, 1989). Широкое распространение перестроек в виде перицентрических инверсий и транслокаций с использованием методов С-бэндинга и FISH убедительно показано как на культурных растениях (Belay, Merker, 1998; Adonina et al., 2016; Badaeva et al., 2018), так и на диких видах, в том числе таксономически относительно близких к Chondrilla (Lim et al., 2008; Chester et al., 2015).
C. laticoronata – недавний вселенец на юго-восток европейской части России, скорее всего из Западного Казахстана. Популяция 56 – самая северная из астраханских и географически ближе всего произрастает к исследованной западно-казахстанской популяции 76. Только в этих двух популяциях выявлен полиморфизм по хромосомам III группы. В то же время во всех 4-х популяциях хромосомы IV группы морфологически отличались друг от друга. В связи с этим возможно, что в популяциях 70 и 71, более южных в пределах Астраханской обл., популяции 56 (север Астраханской обл.) и западноказахстанской популяции 76 проходили независимые миграции кариотипов этого вида из Казахстана на территорию Астраханской обл.
Исходя из гипотезы о том, что три морфологически разных хромосомы в IV группе, встречающиеся во всех кариотипах C. laticoronata, возникли как следствие предполагаемых хромосомных мутаций в виде транслокации и/или перицентрической инверсии, а в хромосомах III группы у части растений двух из четырех исследованных популяций произошла перицентрическая инверсия, можно полагать, что мутации в хромосомах IV группы гораздо древнее, чем мутации по хромосомам III группы. Эти перестройки произошли в свое время, скорее всего, в триплоидном кариотипе C. pauciflora и могли быть причиной репродуктивной изоляции и последующего возникновения таксона C. laticoronata.
У других исследованных видов подрода Chondrilla также обнаружен полиморфизм по IV группе хромосом, аналогичный C. laticoronata. Однако, распределение отклоняющихся от метацентрического типа хромосом этой группы в разных видах отличалось. Так, у C. brevirostris субметацентрики по первой и третьей хромосомам IV группы в популяции 59 обнаружены у всех особей, в популяции 58 – у 2/3, 29 и 55 – у 1/2, а в популяции 46 – лишь у 1/3 особей. Акроцентрическая хромосома в IV группе обнаружена только в кариотипах популяции 59 и всего в 17% случаев, т.е. кариотипы только этих особей были абсолютно идентичны кариотипам популяций C. laticoronata. Так как для С. laticoronata кариотип с тремя морфологически различающимися хромосомами в IV группе является типичным для всех особей всех исследованных популяций, а у C. brevirostris подобный кариотип отмечен только у части из них, можно предположить, что C. brevirostris является следствием гибридизации с С. laticoronata или близкими ему таксонами подрода. Морфологически различные хромосомы IV группы обнаружены и в трех популяциях C. canescens: в кариотипах одних популяций выявлены различия только по первой и третьей хромосомам, а в популяции 74 – только по второй хромосоме. Подобные кариотипы встречались и у C. latifolia, но в пределах одной популяции (60). У C. juncea в популяции 27 более чем для половины особей характерно присутствие двух субметацентрических хромосом разной длины и одной метацентрической хромосомы в IV группе, а у 1/3 особей все три хромосомы группы были морфологически различными (последнее характерно и для 20% особей популяции 19). В исследованных популяциях C. graminea полиморфизма по IV группе хромосом не выявлено, хотя зачастую этот вид сосуществует в симпатрических популяциях с C. juncea, образуя непрерывные ряды переходов морфотипов от одной крайней формы к другой.
Таким образом, по частоте встречаемости и характеру морфологических различий хромосом IV группы исследованные виды подрода Chondrilla распределяются по градиенту следующим образом: C. laticoronata – C. brevirostris – C. juncea – C. canescens – C. latifolia. Это может отражать степень родства между ними: чем дальше по градиенту находятся таксоны друг от друга, тем менее родственны они между собой. При этом те или иные сочетания морфологически различных хромосом III и, особенно, IV групп, вероятно, и определяют морфотипы видов подрода Chondrilla. Все таксономически значимые видовые морфологические признаки (ширина листьев, длина семянки и ее носика, степень выраженности коронки, шипиков на семянке, расположение и направление щетинок и др.) – в основном количественные. Различия C. ambigua и C. pauciflora связаны с различиями в числе полных гаплоидных наборов в кариотипе, т.е. с различным числом полных гаплокопий генома, а у остальных исследованных таксонов – с числом копий отдельных генов и их сочетаний. Сходность предполагаемых хромосомных перестроек в кариотипах исследованных видов этого подрода свидетельствует, по-видимому, о том, что они либо очень близки, либо это один “сборный” вид, возникший первоначально, возможно, в Средней Азии с кариотипом, близким к кариотипу C. laticoronata. Затем от него произошли несущественно разнящиеся между собой виды, которые расселись в западном и северо-западном направлениях до границ современных их ареалов.
С хромосомами III и IV групп может быть связана и “детерминация” гаметофитного апомиксиса у видов Chondrilla, так как морфологические различия хромосом этих групп выявлены у таксонов, которым свойственен гаметофитный апомиксис (C. laticoronata, C. brevirostris, C. canescens и C. juncea) и они не отмечены у C. ambigua, размножающейся облигатно амфимиктично (Kashin et al., 2015; Ugolnikova et al., 2017). При этом, вероятно, изначально все хромосомы пракариотипа Chondrilla были метацентрическими, как это имеет место у C. ambigua и C. pauciflora. Поэтому C. ambigua следует считать наиболее древним среди исследованных видов, а C. pauciflora – триплоидным производным от него без серьезных хромосомных перестроек. Произошедшие мутации, скорее всего, в триплоидном кариотипе C. pauciflora в хромосомах IV, а позже и III группы привела к репродуктивной изоляции. Это обеспечило последующий независимый генезис возникшей формы к C. laticoronata с факультативно апомиктичным способом воспроизводства, а также остальным исследованным таксонам, размножающимся также факультативно апомиктично. Не исключено, что последние являются следствием повторной гибридизации C. laticoronata (в качестве отцовской формы) с C. ambigua (в качестве материнской формы), как это обычно происходит в агамных комплексах (Kashin, 1999, 2000). Однако высказанные предположения носят сугубо гипотетический характер, для проверки которых необходимы молекулярно-цитогенетические исследования с использованием дифференциального окрашивания хромосом.
Список литературы
[Adonina et al.] Адонина И.Г., Леонова И.Н., Бадаева Е.Д., Салина Е.А. 2016. Генотипирование сортов мягкой пшеницы разных регионов России. – Вавиловский журнал генетики и селекции. 20 (1): 44–50. https://doi.org/10.18699/VJ16.107
Amosova A.V., Badaeva E.D., Muravenko O.V., Zelenin A.V. 2009. An improved method of genomic in situ hybridization (GISH) for distinguishing closely related genomes of tetraploid and hexaploid wheat species. – Russian J. Develop. Biol. 40 (2): 90–94. https://doi.org/10.1134/S1062360409020040
Badaeva E.D., Shishkina A.A., Goncharov N.P., Zuev E.V., Lysenko N.S., Mitrofanova O.P., Dragovich A.Yu, Kudriavtsev A.M. 2018. Evolution of Triticum aethiopicum Jakubz. from the Position of Chromosome Analysis. – Russian J. Genetics. 54 (6): 629–642. https://doi.org/10.1134/S1022795418060029
Belay G., Merker A. 1998. Cytogenetic analysis of a spontaneous 5B/6B translocation in tetraploid wheat landraces from Ethiopia, and implications for breeding. – Plant Breed. 11 (6): 537–542. https://doi.org/10.1111/j.1439-0523.1998.tb02203.x
Bergman B.T. 1952. Chromosome morphological studies in Chondrilla juncea and some remarks on the microsporogenesis. – Hereditas. 38 (2): 128–130. https://doi.org/10.1111/J.1601-5223.1952.TB02918.X
[Bugrov] Бугров А.Г. 2010. Кариотипы саранчовых России и сопредельных территорий. – Евразиатский энтомол. журн. 9 (2): 169–179.
Chester M., Riley R.K., Soltis P.S., Soltis D.E. 2015. Patterns of chromosomal variation in natural populations of the neoallotetraploid Tragopogon mirus (Asteraceae). – Heredity. 114: 309–317. https://doi.org/10.1038/hdy.2014.101
Cosendai A.-C., Hörandl E. 2010. Cytotype stability, facultative apomixis and geographical parthenogenesis in Ranunculus kuepferi (Ranunculaceae). – Ann. Bot. 105: 457–470. https://doi.org/10.1093/Aob/Mcp304
[Elenevskii et al.] Еленевский А.Г., Буланый Ю.И., Радыгина В.И. 2008. Определитель сосудистых растений Саратовской области. Саратов. 248 с.
Flann C. (ed.) 2009. Global Compositae Checklist. https://compositae.landcareresearch.co.nz/Default.aspx?Page = NameSearch&searchText = chondrilla
Gadella T.W.J. 1988. Some notes on the origin of polyploidy in Hieracium pilosella aggr. – Acta. Bot. Neerl. 37 (4): 515–522. https://doi.org/10.1111/J.1438-8677.1988.Tb02158.X
[Gileva] Гилева Э.А. 1989. Хромосомная изменчивость и эволюция. М. 141 с.
[Grif, Agapova] Гриф В.Г., Агапова Н.Д. 1986. К методике описания кариотипов растений. – Бот. журн. 71 (4): 550–553.
Gustafsson A. 1946. Apomixis in higher plants. Pt I-III. – Lunds. Univ. Arsskrift. 42: 1–68.
Gustafsson A. 1947. Apomixis in higher plants. Pt I-III. – Lunds. Univ. Arsskrift. 43: 69–370.
[Ilyin] Ильин М.М. 1930. Критический обзор рода Chondrilla L. – Бюлл. отд. каучуконосов. 3: 1–61.
Kashin A.S. 1999. Gametophytic apomixis and the problem of genomic chromosome imbalance in Angiosperms. – Russian J. Genetics. 35 (8): 890–900.
[Kashin] Кашин А.С. 2000. Геномная изменчивость, гибридогенез и возможности хромосомного видообразования при гаметофитном апомиксисе. – Успехи совр. биол. 120 (1): 502–512.
Kashin A.S. 2012. Genesis of cells of apical meristems and realization of gametophytic apomixis in flowering plants. – Russian J. Develop. Biol. 43 (2): 101–114. https://doi.org/10.1134/S1062360411040059
[Kashin et al.] Кашин А.С., Залесная С.В., Титовец В.В. 2000. Потенциал формообразования агамного комплекса Pilosella. 3. Геномная изменчивость в популяциях и потомстве отдельных растений. – Бот. журн. 85 (12): 13–28.
[Kashin et al.] Кашин А.С., Крицкая Т.А., Попова А.О., Пархоменко А.С. 2017. Генетическая дифференциация видов Chondrilla (Asteraceae) европейской части России по данным ISSR маркирования. – Бюл. Моск. об-ва испыт. природы. Отд. биол. 122 (1): 50–70.
[Kashin et al.] Кашин А.С., Петрова Н.А., Шанцер И.А., Кондратьева А.О., Шилова И.В., Пархоменко А.С. 2018. Изменчивость морфологических параметров видов Chondrilla флоры юга европейской части России в связи с их систематикой. – Бот. журн. 103 (11):1407–1436. https://doi.org/10.7868/S0006813618110030
[Kashin et al.] Кашин А.С., Попова А.О., Кочанова И.С., Угольникова Е.В., Полякова Ю.А. 2015. Некоторые параметры системы семенного размножения в популяциях видов Chondrilla L. Нижнего Поволжья. – Бот. журн. 100 (8): 828–840. https://doi.org/10.1134/S0006813615080074
Kelley A.M., Johnson P.G., Waldron B.L., Peel M.D. 2009. A survey of apomixis and ploidy levels among Poa L. (Poaceae) using flow cytometry. – Crop Sci. 49 (4): 1395–1402. https://doi.org/10.2135/cropsci2008.09.055
[Leonova] Леонова Т.Г. 1964. Род. Хондрилла – Chondrilla L. – В кн.: Флора СССР. Т. 29. М.‑Л. С. 560–586.
[Leonova] Леонова Т.Г. 1989. Хондрилла – Chondrilla L. – В кн.: Флора Европейской части СССР. Т. 8. С. 57–61.
Lim K.Y., Soltis D.E., Soltis P.S., Tate J., Matyasek R., Srubarova H., Kovarik A., Pires J.C., Xiong Z., Letch A.R. 2008. Rapid chromosome evolution in recently formed polyploids in Tragopogon (Asteraceae). – PLoS ONE. 3 (10): 33–53.
[Maevskii] Маевский П.Ф. 2014. Флора средней полосы европейской части России. М. 635 с.
Mräz P., Singliarova B., Urfus T., Krahulec F. 2008. Cytogeography of Pilosella officinarum (Compositae): Altitudinaland longitudinal differences in ploidy level distribution in the Czech republic and Slovakia and the general pattern in Europe. – Ann. Bot. 101 (1): 59–71.
Nasseh Y. 2010. A revision on the genera Chondrilla and Heteroderis (Asteraceae) in Iran. – Iran. J. Bot. 16 (1): 92–95.
Nasseh Y., Joharchi M.A. 2008. Revision the genus Chondrilla (Asteraceae) in Iran. – In: 15th National and 3rd International Conference of Biology, 19–21 August 2008. Tehran. P. 178–179.
[Parkhomenko, Kashin] Пархоменко А.С., Кашин А.С. 2018. Особенности кариотипической изменчивости у некоторых видов рода Chondrilla (Asteraceae). – Бот. журн. 103 (6): 726–739. https://doi.org/10.1134/S0006813618060030
Poddubnaya-Arnoldi W.A. 1933. Geschlechtliche und ungeschlechtliche Fortpflanzung bei einigen Chondrilla-Arten. – Planta. 19 (1): 46–86. https://doi.org/10.1007/BF01937556
Sato K., Yamazaki T., Iwatsubo Y. 2011. Cytogeography of Taraxacum albidum (Asteraceae) in Japan. – Cytologia. 76 (2): 201–212. https://doi.org/10.1508/Cytologia.76.201
Schmelzer G.H. 1997. Review of Pennisetum section Brevivalvula (Poaceae). – Euphytica. 97: 1–20. https://doi.org/10.1023/A:1003077906928
Türkçe T. 2007. Bitki Adları Sözlüğü. Ankara. 512 p.
Tutin T.G., Heywood V.H., Burges N.A., Valentine D.H. 1976. Flora Europaea: Plantaginaceae to Compositae (and Rubiaceae). Vol. 4. Cambridge. 534 p.
Tuzlacı E. 2006. Türkiye Bitkileri Sözlüğü. İstanbul. 354 p.
[Ugolnikova et al.] Угольникова Е.В., Кашин А.С., Кондратьева А.О. 2017. Частота апомиксиса в популяциях видов Chondrilla L. юга европейской части России. – Изв. Саратов. унив. Нов. сер. Сер. Химия. Биология. Экология. 17 (1): 53–61. https://doi.org/10.18500/1816-9775-2017-17-1-53-61
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал