

Ботанический журнал, 2019, T. 104, № 5, стр. 699-716
ЦЕНОФЛОРА ЕЛЬНИКОВ ЧЕРНИЧНЫХ ВОСТОЧНОЙ ФЕННОСКАНДИИ
Н. В. Геникова 1, *, Е. П. Гнатюк 1, А. М. Крышень 1
1 Институт леса Карельского научного центра РАН
185910 Петрозаводск, ул. Пушкинская, 11, Россия
* E-mail: genikova@krc.karelia.ru
Поступила в редакцию 15.03.2019
После доработки 06.04.2019
Принята к публикации 09.04.2019
Аннотация
Статья продолжает серию публикаций, посвященных изменению видового разнообразия лесных растительных сообществ в процессе их восстановления после катастрофических нарушений. Представлен материал по ельникам черничным – зонального и наиболее распространенного типа леса на европейском севере, являющегося фактически основой эксплуатационного фонда лесозаготовительных предприятий. Различные виды рубок и способы последующего восстановления, накладываясь на спектр природных экологических условий, определяют ценотическое разнообразие растительных сообществ в пределах каждой стадии развития. Всего ценофлора ельников черничных Восточной Фенноскандии насчитывает 240 видов сосудистых растений. В растительных сообществах на стадии вырубки обнаружено 200 видов, в молодняках – 119, в средневозрастных лесах – 129 и в лесных сообществах старше 120 лет отмечен всего 71 вид. Общими для ценофлор всех возрастных стадий являются 53 вида сосудистых растений – четверть всей ценофлоры. В ельниках черничных несколько выше число видов в средневозрастных сообществах, чем в молодняках, это отмечалось и для сосняков черничных, что не случайно. Увеличение видового богатства в средневозрастных сообществах происходит за счет заселения их аборигенными видами при одновременном снижении доли гемерофильного комплекса. Не исключаются и эколого-ценотические причины, связанные с самоизреживанием древостоя и улучшением условий освещенности.
Статья продолжает серию публикаций, которые шаг за шагом подводят к построению эколого-динамической типологии лесов на автоморфных почвах. Лесная растительность на автоморфных почвах, являющаяся основой лесоэксплуатационного фонда, в настоящее время представляет мозаику сообществ от вырубки до коренных лесов. Набор вариантов сообществ в пределах каждой сукцессионной стадии обусловлен как экотопом, так и разнообразным антропогенным влиянием и поэтому достаточно широкий. Для его систематизации необходимо понимать закономерности развития сообществ после катастрофических разрушений, определяемые разнообразием факторов отдельно для сукцессионных стадий и в различных типах лесорастительных условий. Среди причин катастрофического разрушения в настоящее время абсолютно преобладает рубка древостоя при том, что рубятся уже сообщества, едва достигшие возраста спелости, что кратно увеличивает разнообразие сообществ и видов на ранних стадиях восстановления (Kryshen, 2006). Ранее мы достаточно подробно разбирали принципы выделения типов лесорастительных условий и возрастных стадий (Kryshen, 2010), обсуждали принципы эколого-динамической модели ценотического разнообразия на автоморфных почвах (Kryshen, 2012) и представили эколого-динамическую типологию сосняков (Kryshen et al., 2018).
Ельники черничные – это зональные таежные сообщества, занимающие на северо-западе России основные площади эксплуатационных лесов. Исключением являются Карелия и Мурманская область, расположенные в пределах Фенноскандии, где природные условия обеспечивают доминирование сосновых лесов. Наши исследования ограничивались в основном этой территорией, но можно с уверенностью утверждать, что закономерности формирования состава сообществ на различных стадиях восстановления ельников черничных будут едины для таежной зоны Европейской части России. Кроме зональных ельников черничных на супесчаных и суглинистых почвах на исследуемой территории произрастают типичные для южной тайги ельники кисличные и неморально-травные. Они довольно редки и встречаются преимущественно в южной части Карелии, наиболее сильно антропогенно трансформированной. Здесь развито сельское хозяйство и практически отсутствуют коренные леса. Описаны сообщества неморально-травных и кисличных ельников, представленных только ранними стадиями, что не дает возможности построить динамические ряды, и поэтому в данном исследовании они исключены из анализа.
ОБЪЕКТЫ И МЕТОДЫ
Тип лесорастительных условий ельников черничных обладает рядом характеристик, позволяющих объективно определять растительные сообщества данного типа в природе на любой стадии развития. Наиболее важным показателем является механический состав почвы, по которому происходит разделение на ельники (супесчаные почвы) и сосняки черничные (песчаные). Это разделение объективно и определяется строением корневых систем. В то же время оно не абсолютно и существуют различные варианты произрастания сообществ с доминированием ели на песках (высокая завалуненность, 2–3 единицы сосны в первом ярусе древостоя, рельеф, закрывающий сообщества от преобладающих ветров, и т.п.). Суглинистые почвы на территории Восточной Фенноскандии встречаются очень редко, в частности, в Карелии площадь суглинистых и глинистых почв менее 6% (Fedorets et al., 2000), поэтому описаний в таких условиях немного. Здесь произрастают высокопродуктивные ельники черничные и кисличные, которые на предварительном этапе исследования были также включены в анализ для понимания роли этого фактора в формировании состава сообщества.
Важными признаками являются также состав и структура подлеска, видовое разнообразие и состав доминантов мохово-лишайникового и травяно-кустарничкового яруса. Результаты ординационного анализа (DCA-ординация) (рис. 1) наглядно демонстрируют сходство структуры напочвенного покрова ельников черничных на супесчаных и суглинистых почвах. Редкие описания ельников черничных на суглинках расположены внутри “облака” преобладающих ельников черничных на супесях, в то время как ельники кисличные и неморально-травные распределились по его периферии. Это позволило нам объединить описания ельников черничных на супесях и суглинках и рассматривать их как один тип Picea abies – Vaccinium myrtillus (P.a–V.m.).
Рис. 1.
Ординационная диаграмма геоботанических описаний напочвенного покрова растительных сообществ на супесчаных и суглинистых почвах. Условные обозначения: 1 – ельники черничные на супесчаных почвах, 2 – ельники черничные на суглинистых почвах, 3 – ельники неморально-травные, 4 – ельники кисличные.
Fig. 1. Ordination diagram of geobotanical descriptions of ground cover of plant communities on loamy sand and loamy soils. Legend: 1 – bilberry spruce forest type on loamy sand soils, 2 – bilberry spruce forest type on loamy soils, 3 – rich-herb spruce forest type, 4 – wood sorrel spruce forest type.

В анализ ценофлоры ельников черничных включены описания, выполненные авторами за почти 20-летний период исследований. Описания систематизированы в базе данных “Местообитания Восточной Фенноскандии” (Kryshen et al., 2009). Для данного исследования отобраны описания, полностью отвечающие указанным требованиям, всего – 196, в том числе вырубок 54, молодняков 43, средневозрастных 62, старовозрастных 37. В старовозрастные леса мы объединили описания со средним возрастом древостоя старше 120 лет. Дело в том, что в настоящее время единичны описания субклимаксовых сообществ и практически отсутствуют климаксовые (их описания выполнены только на территориях национальных парков и заповедников). Различий в видовом составе на возрастных стадиях старше 120 лет практически нет.
Мы понимаем ценофлору как объединение полных территориальных совокупностей видов растений в экологически однотипные сообщества (Methods …, 2005). В статье обсуждаются различия в составе видов сосудистых растений (ценофлор) на этапах восстановления ельников черничных после катастрофических нарушений. Анализ полученного материала проводился по следующей схеме: 1) объединение списков видового состава однотипных и входящих в одну возрастную стадию сообществ в ценофлоры; 2) сравнение и оценка выделенных ценофлор по таксономическим и типологическим признакам (Methods…, 2005).
ЦЕНОФЛОРА ЕЛЬНИКОВ ЧЕРНИЧНЫХ
Всего в ельниках черничных выявлено 240 видов сосудистых растений из 148 родов и 54 семейств (табл. 1). Для сравнения в сосняках лишайниковых, брусничных и черничных, площадь которых в Карелии в 1.5 раза больше, чем ельников, произрастает всего 97 видов.
Таблица 1.
Видовой состав сосудистых растений на разных стадиях восстановления ельников черничных. Table 1. Vascular plant species composition at different stages of reforestation in bilberry spruce forest type
| Виды растений Plant species |
Вырубки Loggings |
Молодняки Young forests | Средневозрастные леса/ Middle-aged forests |
Старовозрастные леса/ Old-growth forests |
Фракция флоры/ Flora fractions |
|---|---|---|---|---|---|
| Сем. Lycopodiaceae | |||||
| Diphasiastrum complanatum (L.) Holub | I+ | – | I1 | I+ | ab. |
| Lycopodium annotinum L. | I+ | II+ | II+ | II1 | ab. |
| L. clavatum L. | – | I+ | I+ | – | apoph. |
| Сем. Equisetaceae | |||||
| Equisetum arvense L. | – | – | I+ | – | ap. |
| E. palustre L. | – | – | I+ | – | ab. |
| E. pratense Ehrh. | I+ | – | – | – | apoph. |
| E. sylvaticum L. | II1 | I1 | II1 | III1 | ab. |
| Сем. Botrychiaceae | |||||
| Botrychium boreale Milde | – | – | I+ | – | ab. |
| Сем. Hypolepidaceae | |||||
| Pteridium latiusculum (Desv.) Hieron. ex Fries | I+ | I+ | I+ | – | apoph. |
| Сем. Thelypteridaceae | |||||
| Phegopteris connectilis (Michx.) Watt | I+ | – | I+ | I+ | ab. |
| Сем. Dryopteridaceae | |||||
| Dryopteris carthusiana (Vill.) H. P. Fuchs | I1 | II1 | III+ | I1 | apoph. |
| D. expansa (C. Presl) Fraser-Jenk. & Jermy | I1 | – | I1 | I+ | ab. |
| D. filix-mas (L.) Schott | – | – | I+ | – | ab. |
| Сем. Athyriaceae | |||||
| Athyrium filix-femina (L.) Roth | I1 | I+ | I2 | I+ | ab. |
| Gymnocarpium dryopteris (L.) Newm. | II1 | II1 | II1 | I1 | ab. |
| Сем. Polypodiaceae | |||||
| Polypodium vulgare L. | I+ | – | I+ | – | ab. |
| Сем. Pinaceae | |||||
| Larix sibirica Ledeb. | – | – | I | – | cult. |
| Picea abies (L.) H. Karst. | IV | V | V | V | ab. |
| Pinus sylvestris L. | III | IV | IV | IV | ab. |
| Сем. Cupressaceae | |||||
| Juniperus communis L. | I+ | I1–2 | II1–2 | II1 | ab. |
| Сем. Ranunculaceae | |||||
| Aconitum septentrionale Koelle | I+ | – | – | – | ab. |
| Actaea erythrocarpa Fisch. | – | – | I+ | – | ab. |
| Actaea spicata L. | I+ | – | – | – | ab. |
| Anemonoides ranunculiodes (L.) Holub | – | – | I+ | I+ | ab. |
| Aquilegia vulgaris L. | I+ | – | – | – | adv. |
| Caltha palustris L. | – | – | I+ | – | ab. |
| Hepatica nobilis Mill. | – | – | I1 | I2 | ab. |
| Ranunculus acris L. | I+ | I+ | I+ | – | apoph. |
| R. auricomus L. | I+ | I+ | I+ | – | apoph. |
| R. repens L. | I1 | I+ | I+ | – | apoph. |
| Trollius europaeus L. | I+ | I+ | I1 | – | apoph. |
| Сем. Fumariaceae | |||||
| Fumaria officinalis L. | I+ | – | – | – | adv. |
| Сем. Urticaceae | |||||
| Urtica dioica L. | I+ | I1 | – | – | apoph. |
| Сем. Betulaceae | |||||
| Alnus incana (L.) Moench | I | II | I | I | ab. |
| Betula nana L. | I+ | – | – | – | ab. |
| B. pendula Roth | V | V | V | V | ab. |
| B. pubescens Ehrh. | V | V | V | V | ab. |
| Acer platanoides L. | I | – | I | – | ab. |
| Сем. Caryophyllaceae | |||||
| Alsine media L. | – | – | I+ | – | adv. |
| Cerastium arvense L. | I+ | – | – | – | adv. |
| Cerastium holosteoides Fries | I+ | – | – | – | ab. |
| Hylebia nemorum (L.) Fourr. | I+ | I+ | – | – | ab. |
| Melandrium dioicum (L.) Coss. & Germ. | – | – | I+ | I+ | adv. |
| Moehringia trinerva (L.) Clairv. | I+ | I+ | I+ | – | ab. |
| Sagina procumbens L. | I+ | – | – | – | apoph. |
| Stellaria fennica (Murb.) Perf. | I+ | – | – | – | ab. |
| S. graminea L. | I+ | I1 | I+ | I+ | apoph. |
| S. holostea L. | I1 | – | I+ | – | ab. |
| S. palustris Retz. | I+ | – | – | – | ab. |
| Viscaria viscosa (Scop.) Aschers. | I+ | – | – | – | ab. |
| Сем. Chenopodiaceae | |||||
| Chenopodium album L. | I+ | – | – | – | adv. |
| Сем. Polygonaceae | |||||
| Fallopia convolvulus (L.) Á. Löve | I+ | – | – | – | adv. |
| Rumex acetosa L. | I+ | – | I+ | – | apoph. |
| R. acetosella L. | I1 | – | – | – | apoph. |
| Сем. Hypericaceae | |||||
| Hypericum maculatum Crantz | I+ | I1 | I+ | – | adv. |
| H. perforatum L. | – | I+ | – | – | ab. |
| Сем. Violaceae | |||||
| Viola arvensis Murr. | I+ | – | – | – | adv. |
| V. epipsila Ledeb. | – | – | I+ | – | ab. |
| V. mirabilis L. | I+ | I+ | I+ | – | ab. |
| Viola nemoralis Kütz. | I+ | I1 | I+ | – | ab. |
| V. riviniana Reichenb. | I+ | – | – | – | ab. |
| V. tricolor L. | I+ | – | – | – | apoph. |
| Сем. Brassicaceae | |||||
| Barbarea arcuata (Opiz ex J. et C. Presl) Reichenb. | I+ | – | – | – | adv. |
| Barbarea stricta Andrz. | I+ | – | – | – | ab. |
| Сем. Salicaceae | |||||
| Populus tremula L. | II | III | III | II | ab. |
| Salix aurita L. | I+ | – | I+ | – | ab. |
| S. caprea L. | I1 | III1–2 | I1 | I1 | ab. |
| S. phylicifolia L. | I+ | I+ | – | I+ | ab. |
| Сем. Ericaceae | |||||
| Andromeda polifolia L. | I+ | – | – | – | ab. |
| Arctostaphylos uva-ursi (L.) Spreng. | – | I+ | – | – | apoph. |
| Calluna vulgaris (L.) Hull | I1 | II1 | I+ | I1 | ab. |
| Chamaedaphne calyculata (L.) Moench | I1 | – | – | – | ab. |
| Ledum palustre L. | I2 | I+ | I+ | I+ | ab. |
| Vaccinium myrtillus L. | V1 | V2 | V2 | V3 | ab. |
| V. uliginosum L. | I1 | I1 | I1 | I2 | ab. |
| V. vitis-idaea L. | IV2 | IV2 | V1 | V2 | ab. |
| Сем. Pyrolaceae | |||||
| Moneses uniflora (L.) A. Gray | – | I+ | I+ | – | ab. |
| Orthilia secunda (L.) House | I+ | I+ | II+ | II+ | ab. |
| Pyrola chlorantha Sw. | – | – | I+ | I1 | ab. |
| P. media Sw. | – | I+ | I+ | – | ab. |
| P. minor L. | I+ | I+ | I+ | – | ab. |
| P. rotundifolia L. | I+ | – | I1 | I+ | ab. |
| Сем. Empetraceae | |||||
| Empetrum nigrum L. s. l. | I1 | I1 | I1 | II1 | ab. |
| Сем. Primulaceae | |||||
| Lysimachia vulgaris L. | – | – | I+ | I1 | ab. |
| Trientalis europaea L. | II+ | III+ | IV+ | III1 | ab. |
| Сем. Thymelaeaceae | |||||
| Daphne mezereum L. | I+ | I+ | I+ | – | ab. |
| Сем. Saxifragaceae | |||||
| Chrysosplenium alternifolium L. | I1 | – | I+ | – | ab. |
| Сем. Grossulariaceae | |||||
| Ribes nigrum L. | I+ | – | – | – | ab. |
| R. spicatum E. Robson | I+ | I+ | I1 | I+ | ab. |
| Сем. Rosaceae | |||||
| Alchemilla acutiloba Opiz | I+ | I1 | I+ | – | adv. |
| Comarum palustre L. | – | I+ | – | – | ab. |
| Filipendula ulmaria (L.) Maxim. | I1 | I+ | I1 | I+ | ab. |
| Fragaria vesca L. | I1 | II1 | I1 | I1 | apoph. |
| Geum rivale L. | I1 | I+ | – | – | ab. |
| Padus avium Mill. | – | I+ | I1 | – | ab. |
| Potentilla argentea L. | I+ | – | – | – | apoph. |
| Potentilla erecta (L.) Raeusch. | I+ | I+ | I+ | – | apoph. |
| Potentilla intermedia L. | I+ | – | – | – | adv. |
| P. norvegica L. | I+ | – | – | – | adv. |
| Rosa acicularis Lindl. | I+ | I+ | I+ | II+ | ab. |
| R. majalis Herrm. | I+ | – | – | – | ab. |
| Rubus arcticus L. | I1 | – | I+ | I+ | ab. |
| R. chamaemorus L. | I+ | – | – | I1 | ab. |
| R. idaeus L. | I2 | I1 | I2 | I+ | apoph. |
| R. saxatilis L. | II1 | III1 | III1 | II1 | ab. |
| Sorbus aucuparia L. | IV1 | IV1–2 | V1–2 | V1 | ab. |
| Сем. Fabaceae | |||||
| Chrysaspis aurea (Poll.) Greene | I+ | – | – | – | adv. |
| Lathyrus pratensis L. | I+ | I+ | – | – | apoph. |
| L. vernus (L.) Bernh. | I+ | I1 | I+ | I+ | ab. |
| Lupinus polyphyllus Lindl. | – | I2 | – | – | adv. |
| Trifolium medium L. | I+ | – | – | – | apoph. |
| T. pratense L. | I+ | I1 | – | – | adv. |
| T. repens L. | I+ | I1 | I+ | – | adv. |
| Vicia cracca L. | I+ | I1 | – | – | apoph. |
| V. sepium L. | I+ | I1 | I+ | – | apoph. |
| V. sylvatica L. | – | I1 | – | – | ab. |
| Сем. Onagraceae | |||||
| Chamaenerion angustifolium (L.) Scop. | V2 | V2 | I+ | I+ | apoph. |
| Epilobium adenocaulon Hausskn. | I+ | – | – | – | adv. |
| E. montanum L. | I+ | – | I+ | – | ab. |
| E. palustre L. | I+ | – | – | – | ab. |
| Сем. Oxalidaceae | |||||
| Oxalis acetosella L. | I+ | II1 | III2 | II2 | ab. |
| Сем. Geraniaceae | |||||
| Geranium sylvaticum L. | II1 | II1 | I1 | I+ | ab. |
| Сем. Cornaceae | |||||
| Chamaepericlymenum suecicum (L.) Asch. & Graebn. | I1 | I1 | I+ | I+ | ab. |
| Сем. Apiaceae | |||||
| Aegopodium podagraria L. | – | I1 | I+ | – | apoph. |
| Angelica sylvestris L. | I+ | II1 | I+ | I+ | ab. |
| Anthriscus sylvestris (L.) Hoffm. | I+ | I2 | I1 | – | apoph. |
| Heracleum sibiricum L. | I+ | – | – | – | apoph. |
| Pimpinella saxifraga L. | I+ | I1 | – | – | adv. |
| Сем. Rhamnaceae | |||||
| Frangula alnus Mill. | I+ | – | I1 | – | ab. |
| Сем. Rubiaceae | |||||
| Galium album Mill. | I1 | I1 | I+ | – | adv. |
| Galium boreale L. | I+ | – | – | – | apoph. |
| Galium triflorum Michaux | I+ | – | – | – | ab. |
| Сем. Viburnaceae | |||||
| Viburnum opulus L. | – | I+ | – | – | ab. |
| Сем. Caprifoliaceae | |||||
| Linnaea borealis L. | II+ | II+ | II+ | IV+ | ab. |
| Lonicera xylosteum L. | – | – | I1 | I+ | ab. |
| Сем. Dipsacaceae | |||||
| Knautia arvensis (L.) Coult. | I+ | – | I+ | – | apoph. |
| Сем. Lamiaceae | |||||
| Galeopsis tetrahit L. | I+ | I2 | I+ | – | adv. |
| Lamium album L. | I+ | – | – | – | adv. |
| Mentha arvensis L. | – | – | I+ | – | ab. |
| Prunella vulgaris L. | I+ | I2 | – | – | apoph. |
| Сем. Callitrichaceae | |||||
| Callitriche palustris L. | I+ | – | – | – | apoph. |
| Сем. Scrophulariaceae | |||||
| Melampyrum nemorosum L. | I+ | I+ | – | I1 | apoph. |
| M. pratense L. | II1 | IV2 | IV1 | III+ | ab. |
| M. sylvaticum L. | I+ | II+ | III1 | II1 | ab. |
| Pseudolysimachion longifolium (L.) Opiz | – | – | I+ | – | apoph. |
| Veronica chamaedrys L. | I+ | I+ | I+ | I+ | apoph. |
| V. officinalis L. | I+ | I+ | I+ | – | apoph. |
| Сем. Campanulaceae | |||||
| Campanula glomerata L. | – | I+ | I+ | – | apoph. |
| C. patula L. | I+ | I1 | – | – | adv. |
| C. persicifolia L. | I+ | I1 | – | – | ab. |
| C. rapunculoides L. | I+ | – | – | – | ab. |
| C. rotundifolia L. | I+ | – | I+ | – | ab. |
| Сем. Asteraceae | |||||
| Achillea millefolium L. | I+ | I+ | I+ | – | apoph. |
| Artemisia vulgaris | I+ | – | – | – | apoph. |
| Bidens radiata Thuill. | I+ | – | – | – | ab. |
| Centaurea jacea L. | I+ | – | – | – | adv. |
| Centaurea phrygia L. | – | – | I+ | – | apoph. |
| Cirsium heterophyllum (L.) Hill | I+ | I+ | I+ | – | ab. |
| C. palustre (L.) Scop. | I+ | – | – | – | apoph. |
| C. setosum (Willd.) Bess. | I+ | – | – | – | adv. |
| C. vulgare (Savi) Ten. | I+ | I1 | – | – | adv. |
| Crepis paludosa (L.) Moench | – | – | I+ | – | ab. |
| C. tectorum L. | – | I1 | – | – | apoph. |
| Hieracium umbellatum L. | I+ | I+ | I+ | – | apoph. |
| H. vulgatum Fries s. l. | I+ | I+ | I+ | I+ | ab. |
| Leontodon autumnalis L. | I+ | I+ | – | – | apoph. |
| L. hispidus L. | I+ | – | – | – | apoph. |
| Leucanthemum ircutianum Lam. | I+ | I1 | – | – | adv. |
| Omalotheca sylvatica (L.) Sch. Bip. & F. Schultz | I+ | – | – | – | ab. |
| Petasites frigidus (L.) Fries | I+ | – | – | – | ab. |
| Picris hieracioides L. | I+ | – | – | – | apoph. |
| Pilosella caespitosa (Dumort.) P. D. Sell & C. West | – | I1 | – | – | apoph. |
| P. officinarum F. Schultz & Sch. Bip. | – | I+ | – | – | apoph. |
| Solidago virgaurea L. | III1 | V1 | IV1 | III+ | ab. |
| Sonchus arvensis L. | I1 | – | – | – | adv. |
| Tanacetum vulgare L. | I+ | – | – | – | apoph. |
| Taraxacum officinale L. | I+ | I+ | – | – | apoph. |
| Trommsdorffia maculata (L.) Bernh. | – | I+ | I+ | – | ab. |
| Tussilago farfara L. | I+ | I+ | – | – | apoph. |
| Сем. Trilliaceae | |||||
| Paris quadrifolia L. | I+ | I1 | I+ | I+ | ab. |
| Сем. Convallariaceae | |||||
| Convallaria majalis L. | I1 | II2 | III1 | I1 | ab. |
| Maianthemum bifolium (L.) F.W. Schmidt | II1 | IV1 | IV1 | III1 | ab. |
| Сем. Orchidaceae | |||||
| Dactylorhiza maculata (L.) Soó | – | I+ | I+ | – | ab. |
| Epipactis palustris (L.) Crantz | I+ | – | – | – | ab. |
| Goodyera repens (L.) R. Br. | – | I+ | I+ | I+ | ab. |
| Listera cordata (L.) R. Br. | I+ | – | I+ | I+ | ab. |
| Malaxis monophyllos (L.) Sw. | – | – | I+ | – | ab. |
| Platanthera bifolia (L.) Rich. | I+ | I+ | I+ | I+ | ab. |
| Сем. Lemnaceae | |||||
| Lemna minor L. | I+ | – | – | – | apoph. |
| Сем. Alismataceae | |||||
| Alisma plantago-aquatica L. | I+ | – | – | – | ab. |
| Сем. Potamogetonaceae | |||||
| Potamogeton alpinus Balb. | I+ | – | – | – | ab. |
| Сем. Typhaceae | |||||
| Typha latifolia L. | I+ | – | – | – | apoph. |
| Сем. Juncaceae | |||||
| Juncus articulatus L. | I+ | – | – | – | apoph. |
| J. bufonius L. | I+ | – | – | – | apoph. |
| J. conglomeratus L. | I+ | – | – | – | apoph. |
| J. effusus L. | I+ | – | – | – | apoph. |
| J. filiformis L. | I+ | I+ | – | – | ab. |
| J. nodulosus Wahlenb. | I+ | – | – | – | ab. |
| Luzula multiflora (Ehrh.) Lej. | I1 | – | – | – | apoph. |
| L. pallescens Sw. | I+ | – | – | – | apoph. |
| L. pilosa (L.) Willd. | IV1 | III1 | III+ | IV+ | ab. |
| Сем. Cyperaceae | |||||
| Carex acuta L. | I1 | – | – | – | ab. |
| C. brunnescens (Pers.) Poir | I2 | – | – | I2 | ab. |
| C. canescens L. | I2 | I+ | I+ | – | ab. |
| C. cespitosa L. | I+ | – | – | – | ab. |
| C. digitata L. | I+ | – | I+ | – | ab. |
| C. echinata Murr. | I+ | – | – | – | ab. |
| C. elongata L. | I2 | – | – | – | ab. |
| C. globularis L. | I2 | I1 | I+ | II1 | ab. |
| C. juncella (Fries) Th. Fries | I+ | – | – | – | ab. |
| C. leporina L. | I+ | – | – | – | apoph. |
| C. nigra (L.) Reichard | I1 | – | – | – | ab. |
| C. pallescens L. | I+ | – | I+ | – | apoph. |
| C. paupercula Michaux | I1 | – | – | – | ab. |
| C. rhynchophysa C. A. Mey. | I+ | – | – | – | ab. |
| C. rostrata Stokes | I+ | – | – | – | ab. |
| C. vaginata Tausch | – | I+ | I1 | – | ab. |
| C. vesicaria L. | I+ | – | – | – | ab. |
| Eriophorum vaginatum L. | I+ | I+ | – | – | ab. |
| Scirpus sylvaticus L. | I2 | – | – | – | ab. |
| Сем. Poaceae | |||||
| Agrostis canina L. | I+ | – | – | – | ab. |
| A. capillaris L. | I2 | I2 | I1 | – | apoph. |
| A. stolonifera L. | I+ | – | – | – | ab. |
| Alopecurus aequalis Sobol. | I+ | – | – | – | ab. |
| Anthoxanthum odoratum L. | I+ | I+ | I+ | – | apoph. |
| Avenella flexuosa (L.) Drej. | V2 | IV2 | IV1 | V2 | ab. |
| Calamagrostis arundinacea (L.) Roth | III2 | IV2 | IV2 | II1 | apoph. |
| C. canescens (Web.) Roth | I2 | I+ | I+ | I+ | ab. |
| C. epigeios (L.) Roth | I2 | I2 | I1 | I+ | apoph. |
| C. phragmitoides C. Hartm. | I1 | I2 | I+ | I2 | ab. |
| Dactylis glomerata L. | I+ | – | – | – | adv. |
| Deschampsia cespitosa (L.) Beauv. | I+ | I1 | I1 | – | apoph. |
| Festuca ovina L. | – | I+ | – | I+ | ab. |
| Melica nutans L. | I+ | I1 | I1 | I1 | ab. |
| Milium effusum L. | I+ | – | I+ | – | ab. |
| Phleum pratense L. | I+ | – | – | – | apoph. |
| Poa nemoralis L. | – | – | I+ | – | ab. |
| P. pratensis L. | I+ | – | – | – | apoph. |
| P. trivialis L. | – | I+ | – | – | apoph. |
| Schedonorus pratensis (Huds.) P. Beauv. | I+ | – | – | – | adv. |
| Количество видов сосудистых растений: Total number of vascular plant species |
200 | 119 | 129 | 71 | |
Примечание. Классы постоянства (%): I – менее 20; II – 21–40; III – 41–60; IV – 61–80; V – 81–100. Обилие вида указано по шкале: “+” – вид редкий и имеет малое проективное покрытие до 1%; 1 – среднее проективное покрытие вида составляет 1–5%; 2 – 5–25%; 3 – 25–50%; 4 – 50–75%; 5 – выше 75%. Значения обилия не указаны для видов древесного яруса. Фракции флоры: adv. – адвентивный, ab. – аборигенный вид, apoph. – апофит, cult. – интродуцент.
Note. Constancy classes: I – under 20%; II – 21–40; III – 41–60; IV – 61–80; V – 81–100. Species abundance: “+” – rare species with weak projective cover under 1%; 1 – species cover 1–5%; 2 – 5–25%; 3 – 25–50%; 4 – 50–75%; 5 – over 75%. The abundance values are not specified for tree layer species. Flora fractions: adv. – adventive, ab. – aboriginal, apoph. – apophyte, cult. – introduced species.
Таблица 2.
Коэффициенты включения ценофлор различных этапов восстановления ельников черничных. Table 2. Inclusion coefficients of coenofloras (species compositions) at different stages of bilberry spruce forest reforestation
| a | b | |||
|---|---|---|---|---|
| Этапы развития сообщества Reforestation stages of community |
||||
| Вырубки Loggings |
Молодняки Young forests | Средневозрастные Middle-aged forests |
Старовозрастные Old-growth forests |
|
| Вырубки Cuttings |
1 | 0.50 | 0.50 | 0.32 |
| Молодняки Young forests |
0.83 | 1 | 0.75 | 0.48 |
| Средневозрастные Middle-aged forests |
0.78 | 0.69 | 1 | 0.51 |
| Старовозрастные Old-growth forests |
0.90 | 0.80 | 0.93 | 1 |
Примечание. Коэффициент расcчитывался по формуле: K = a ∩ b/a, где а – число видов в первой ценофлоре, b – число видов во второй ценофлоре, a ∩ b – число общих видов.
Note. The coefficient values were calculated according to the formula: K = a ∩ b/a, where а – number of species in the first coenoflora, b – number of species in the second coenoflora and a ∩ b is the number of shared species.
В напочвенном покрове нами отмечены 30 видов мхов и лишайников: Cetraria islandica (L.) Ach., Cladonia arbuscula (Wallr.) Flot., C. rangiferina (L.) F.H. Wigg, Peltigera aphtosa (L.) Willd., Aulacomium palustre (Hedw.) Schwägr., Brachythecium sp., Climacium dendroides (Hedw.) F. Weber & D. Mohr, Dicranum fuscescens Turn., D. majus Turn., D. polysetum Sw., D. scoparium Hedw., Hylocomium splendens (Hedw.) Bruch et al., Mnium sp., Plagiothecium laetum Bruch et al., Pleurozium schreberi (Brid.) Mitt., Pohlia nutans (Hedw.) Lindb., Polytrichum commune Hedw., P. juniperinum Hedw., Rhodobryum roseum (Hedw.) Limpr., Ptilium crista-castrensis (Hedw.) De Not., Rhytidiadelhpus triquetrus (Hedw.) Warnst., Sphagnum angustifolium (C.E.O.Jensen ex Russow) C.E.O.Jensen, S. capillifolium (Ehrh.) Hedw., S. centrale C.E.O.Jensen, S. fallax (H.Klinggr.) H.Klinggr., S. fuscum (Schimp.) H.Klinggr., S. girgensohnii Russow, S. magellanicum Brid., S. russowii Warnst., S. squarrosum Crome. Повсеместно в ельниках черничных высоко обилие зеленых мхов Pleurozium schreberi, Dicranum spp., Hylocomium splendens, в понижениях могут доминировать мхи р. Sphagnum.
Наиболее широко в ценофлоре ельников черничных (табл. 1) представлены семейства Asteraceae (27 видов), Poaceae (20), Cyperaceae (19) и Rosaceae (19). Наибольшим количеством видов отличаются рода Carex (17 видов), Juncus (6), Viola (6) и Campanula (5).
Из всего списка в Красную книгу Карелии (Red…, 2007) включены четыре вида, встреченные единично: Botrychium boreale11 (в редкостойном средневозрастном березняке злаково-разнотравном, где проводился сенокос), Epipactis palustris (на вейниковой вырубке в южной части Карелии), Hypericum perforatum (в сосновом молодняке со вторым ярусом ели на бывшем сельхозугодье в Приладожье), Malaxis monophyllos (в 45-летнем сообществе с доминированием осины и березы на заброшенных сельхозполях).
В ельниках черничных наибольшее видовое разнообразие наблюдается на ранних стадиях восстановления, наименьшее – в старовозрастных лесах. В растительных сообществах на стадии вырубки обнаружено 200 видов сосудистых растений, в молодняках – 119, в средневозрастных лесах – 129 и в лесах старше 100 лет отмечен всего 71 вид. Общими для ценофлор всех возрастных стадий являются 53 вида сосудистых растений, т.е. ¼ всей ценофлоры. Только на вырубках обнаружено 79 видов растений (33% от всей ценофлоры ельников черничных), только в молодняках встречено 10 видов, только в средневозрастных лесах – 15. Эти данные отражаются в значениях коэффициента включения, который показывает долю общих видов двух ценофлор по отношению к одной из них. Так, например, коэффициент включения общих видов ценофлор вырубок и молодняков в ценофлору вырубок составляет 0.5. Это значит, что общие виды для этих этапов развития сообществ составляют половину от всего их количества на вырубках, в то время как для молодняков эта общая часть представляет более 80% видов. Разница значительна и иллюстрирует не просто процесс обеднения видами сообщества при формировании древесного яруса, но и то, что на вырубках присутствует большое число временных видов (Kryshen, 2006).
Анализ значений коэффициентов демонстрирует направление качественного изменения состава видов. Несмотря на вторую за вырубками численность видов средневозрастных насаждений (табл. 1), старовозрастные (самые бедные по числу видов) леса по значениям коэффициента включения ближе именно к средневозрастным лесам. Это значит, что в средневозрастных лесах происходит не просто увеличение видов, а качественно меняется состав, что подтверждается данными по доли гемерофильных (адвентивных и апофитных) видов на различных этапах восстановления ельников черничных (рис. 2).
Рис. 2.
Количество адвентивных видов и эвапофитов в ценофлорах сообществ разных возрастных стадий.
Fig. 2. Number of adventive species and euapophytes in the coenofloras of communities at different age stages.
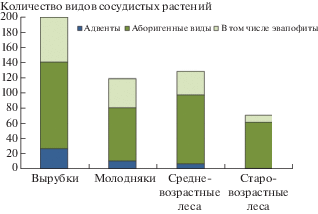
В ельниках черничных нами отмечено 30 адвентивных видов: Alchemilla acutiloba, Alsine media, Aquilegia vulgaris, Barbarea arcuata, Campanula patula, Centaurea jacea, Cerastium arvense, Chenopodium album, Chrysaspis aurea, Cirsium setosum, C. vulgare, Dactylis glomerata, Epilobium adenocaulon, Fallopia convolvulus, Fumaria officinalis, Galeopsis tetrahit, Galium album, Hypericum maculatum, Lamium album, Leucanthemum ircutianum, Lupinus polyphyllus, Melandrium dioicum, Pimpinella saxifraga, Potentilla intermedia, Potentilla norvegica, Schedonorus pratensis, Sonchus arvensis, Trifolium pratense, T. repens, Viola arvensis. Большинство присутствуют только на вырубках. С формированием древесного яруса участие заносных видов сводится практически к нулю. Их число падает с 27 видов на вырубках (13% от общего количества видов в ценофлоре данной возрастной стадии), 11 видов в молодняках (9%), 7 видов (5%) в средневозрастных древостоях и до 1 вида (Melandrium dioicum), встреченного однократно в старовозрастных лесах на границе средней и южной подзон тайги. Бóльшая часть адвентивных видов встречена единично на юге Карелии, наибольшее число их приурочено к лесам вблизи сельскохозяйственных угодий.
Лиственница сибирская считается аборигенным видом для территории Карелии, на востоке республики она обыкновенно встречается в составе древостоя. Однако в обследованных сообществах лиственница произрастает в культурах и поэтому не включена нами в состав аборигенной флоры.
Из 209 видов аборигенной фракции как апофиты (эвапофиты, т.е. аборигенные виды, осваивающие вторичные местообитания и явно положительно реагирующие на действие антропогенных факторов) в “Конспекте флоры Карелии” А.В. Кравченко (Kravchenko, 2007) отмечены 69 видов (рис. 2). Эти виды встречены преимущественно на вырубках, в молодых и средневозрастных сообществах (59, 38 и 31 вид соответственно). В старовозрастных сообществах, где участие лиственных пород в древесном ярусе незначительно, снижается и число видов-апофитов. Фактически остаются лесные травы, типичные для ельников черничных.
Географическая структура аборигенной фракции ценофлоры ельников черничных состоит из шести широтных и семи долготных групп (табл. 3). Среди широтных групп преобладают бореальные виды (141 вид – 67%). Неморально-бореальных и неморальных видов – 34, что составляет 16% от общего числа видов. Суммарная доля северных широтных элементов (гипоарктических и гипоарктобореальных) составляет только 7% и представлены они 14 видами. Среди долготных фракций преобладают циркумполярная (72 вида – 34%) и евразиатская (58 видов – 28%). Следующая по численности (49 видов – 23%) – группа евро-сибирских долготных элементов.
Таблица 3.
Широтная структура аборигенной фракции ценофлоры. Table 3. Latitudinal structure of the aboriginal fraction of coenoflora
| Географические
элементы Geographical elements |
Вырубки Loggings |
Молодняки Young forests |
Средневозрастные леса Middle-age forests |
Старовозрастные леса Old-growth forests |
||||
|---|---|---|---|---|---|---|---|---|
| кол-во number |
% part |
кол-во number |
% part |
кол-во number |
% part |
кол-во number |
% part |
|
| Северная группа: Northern group: |
12 | 6.9 | 6 | 5.6 | 7 | 5.7 | 7 | 10.0 |
| гипоарктические hypoarctic |
8 | 4.6 | 4 | 3.7 | 4 | 3.3 | 4 | 5.7 |
| гипоарктобореальные hypoarctoboreal |
4 | 2.3 | 2 | 1.9 | 3 | 2.5 | 3 | 4.3 |
| Бореальные виды Boreal species |
115 | 66.5 | 78 | 72.2 | 89 | 73.8 | 52 | 74.3 |
| Южная группа: Southern group: |
28 | 16.2 | 18 | 16.7 | 20 | 16.4 | 10 | 14.3 |
| бореально-неморальные boreo-nemoral |
17 | 9.8 | 11 | 10.2 | 11 | 9.0 | 6 | 8.6 |
| неморальные nemoral |
11 | 6.4 | 7 | 6.5 | 9 | 7.4 | 4 | 5.7 |
| Плюризональные виды Plurizonal species |
18 | 10.4 | 6 | 5.6 | 5 | 4.1 | 1 | 1.4 |
| Всего Total |
173 | 100 | 108 | 100 | 121 | 100 | 70 | 100 |
Структура широтных геоэлементов практически одинакова в ценофлорах разных возрастных стадий за исключением плюризональных видов, абсолютное и относительное количество которых уменьшается по мере развития растительного сообщества. В целом, следует отметить, что обогащение сообществ ранних стадий идет за счет всех географических групп видов, в то время как для сосняков на ранних стадиях заметно активное участие южных видов.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Продемонстрированные изменения видового состава в процессе восстановления ельников черничных сходны для таковых и для сосняков (табл. 4), они также отмечались для лиственных лесов Дании (Graae, Heskjar, 1997), для хвойных лесов Швеции (Widenfalk, Weslien, 2009), Финляндии (Tonteri, 1994). Сходные результаты были получены при изучении изменений в напочвенном покрове через 30–50 лет после рубки древостоя (Selmants, 2003) – оказалось, что видовое богатство сосудистых растений в молодняках выше, чем в спелых сообществах, в которых к тому же наблюдалось наименьшее количество адвентивных видов.
Таблица 4.
Количество видов сосудистых растений в ценофлорах сосняков и ельников разных возрастных стадий. Table 4. Vascular plant species number in coenofloras of pine and spruce forests at different age stages
| Сосняк
лишайниковый Lichen pine forest |
Сосняк
брусничный Cowberry pine forest |
Сосняк
черничный Bilberry pine forest |
Ельник
черничный Bilberry spruce forest |
|
|---|---|---|---|---|
| Вырубки Loggings |
17 | 34 | 50 | 200 |
| Молодняки Young forests |
11 | 31 | 55 | 119 |
| Средневозрастные леса Middle-aged forests |
8 | 30 | 65 | 129 |
| Старовозрастные леса Old-growth forests |
13 | 22 | 45 | 71 |
| Всего в ценофлоре Total in the coenoflora |
21 | 49 | 90 | 240 |
Примечания: в таблице приведены данные по числу видов в условиях сосняков, опубликованные ранее (Genikova et al., 2010, 2011, 2012), с дополнениями по результатам исследований последних лет.
Note. Given in the table are published data on the species number in the communities of pine forest types (Genikova et al., 2010, 2011, 2012) added with results of the studies of recent years.
В то же время видовое богатство сообщества зависит от множества факторов и определяется не только стадией сукцессии и возрастом древесного яруса. Одним из наиболее сильных факторов является состав древесного яруса, другое дело, что он также во многом зависит от стадии сукцессии. В целом можно утверждать, что в лиственных лесах разнообразие сосудистых растений выше, чем в хвойных (см. также Barbier et al., 2008; Chávez, Macdonald, 2010 и др.). Различия в видовом составе напочвенного покрова в насаждениях с доминированием разных древесных пород связаны с различиями в условиях местообитаний. Так, при сравнении хвойных и лиственных насаждений в них наблюдаются различия в освещенности, толщине лесной подстилки, водном и температурном режимах почвы (Chávez, Macdonald, 2010).
Эти общие закономерности не универсальны. Если мы переходим на уровень отдельного сообщества, то можно ожидать широкий спектр изменений состава и обилия видов напочвенного покрова. Для северотаежных, как сосняков, так и ельников, удаленных от населенных пунктов, сельскохозяйственных земель и оживленных трасс, показано, что на сплошных вырубках не происходит увеличения видового богатства, меняется только соотношение видов за счет разрастания лесных апофитов (Genikova et al., 2016). При этом практически все лесные виды находят условия для переживания неблагоприятных условий. Это, как правило, куртины и отдельные деревья подроста, пни, складированные порубочные остатки, микропонижения и т.п. (Kryshen, 2006). В средней тайге на антропогенно фрагментированных и сельскохозяйственно освоенных территориях разнообразие увеличивается в разы (Kryshen, 2006; Genikova et al., 2014) за счет моментального освоения освободившейся территории заносными видами и апофитами, а также видами, распространяющихся из соседних сообществ. Кроме самого факта наличия зачатков растений значение для формирования видового разнообразия сообщества имеют содержание элементов минерального питания и почвенное увлажнение (Hardtle et al., 2003; Hokkanen, 2006), которое с уничтожением древесного яруса может резко измениться как в сторону увеличения, так и в сторону снижения, в зависимости от рельефа (Kryshen, 2006).
В целом изменение видового богатства в процессе восстановления ельников черничных после рубки соответствует выявленным ранее в сосняках закономерностям – наибольшее число видов наблюдается на ранних стадиях восстановления за счет заселения вырубок видами, произрастающими в соседних сообществах, занесенными с сельхозполей и из других антропогенных местообитаний (табл. 4). В ельниках черничных по нашим данным несколько выше число видов в средневозрастных сообществах, чем в молодняках, что наблюдалось также и в сосняках черничных (Genikova et al., 2012). Различия в числе видов между молодняками и средневозрастными лесами не существенны, но вряд ли случайны и, скорее всего, связаны с исторически обусловленными причинами – богаты видами сообщества в сельскохозяйственно освоенной южной Карелии. Здесь часть средневозрастных сообществ образовалась на заброшенных финских хуторах, что могло в целом сказаться на разнообразии. В то же время, не исключаются и эколого-ценотические причины, связанные с самоизреживанием древостоя и улучшением условий освещенности, возможны и другие факторы. В пользу этой гипотезы свидетельствует снижение доли гемерофильного комплекса при переходе от молодняков к средневозрастным насаждениям, при возрастании общего числа видов – обогащение происходит за счет аборигенной фракции флоры. Важно влияние экологических факторов и древесного яруса на формирование напочвенного покрова. Эти вопросы будут подробно обсуждаться в специальной публикации, посвященной эколого-ценотическим факторам динамики лесных сообществ в условиях ельников черничных.
Список литературы
Barbier S., Gosselin F., Balandier P. 2008. Influence of tree species on understory vegetation diversity and mechanisms involved. A critical review for temperate and boreal forests. – Forest Ecology and Management. 254 (1): 1–15.
Chávez V., Macdonald S.E. 2010. The influence of canopy patch mosaics on understory plant community composition in boreal mixedwood forest. – Forest Ecology and Management. 259 (6): 1067–1075.
[Fedorets] Федорец Н.Г., Морозова Р.М., Синькевич С.М., Загуральская Л.М. 2000. Оценка продуктивности лесных почв в Карелии. Петрозаводск. 195 с.
[Genikova] Геникова Н.В., Гнатюк Е.П., Крышень А.М. 2012. Анализ ценофлоры лесов на автоморфных песчаных почвах в Карелии. – Бот. журн. 97 (11): 1424–1435.
[Genikova] Геникова Н.В., Гнатюк Е.П., Крышень А.М., Лейбонен Е.Э. 2010. Разнообразие сосудистых растений лесных сообществ на автоморфных песчаных почвах в Карелии. – Ученые записки ПетрГУ. 8: 11–13.
[Genikova] Геникова Н.В., Крышень А.М., Гнатюк Е.П., Кутенков С.А. 2011. Ординация геоботанических описаний лесных сообществ на песчаных почвах в Карелии. – В сб.: Матер. Всерос. научн. конф. “Отечественная геоботаника: основные вехи и перспективы”. Т. 1: Разнообразие типов растительных сообществ и вопросы их охраны. География и картография растительности. История и перспективы геоботанических исследований. Санкт-Петербург. С. 51–54.
[Genikova] Геникова Н.В., Торопова Е.В., Крышень А.М. 2016. Реакция видов напочвенного покрова ельника черничного на рубку древостоя. –Труды КарНЦ РАН. Сер. Экологические исследования. 4: 92–99. https://doi.org/10.17076/есо292
Graae B.J., Heskjar V.S. 1997. A comparison of understorey vegetation between untouched and managed deciduous forest in Denmark. – Forest Ecology and Management. 96 (1–2): 111–123.
Hardtle W., von Oheimb G., Westphal Ch. 2003. The effects of light and soil conditions on the species richness of the ground vegetation of deciduous forests in northern Germany (Schleswig-Holstein). – Forest Ecology and Management. 182 (1–3): 327–338.
Hokkanen P.J. 2006. Environmental patterns and gradients in the vascular plants and bryophytes of eastern Fennoscandian herb-rich forests. – Forest Ecology and Management. 229 (1–3): 73–87.
[Kravchenko] Кравченко А.В. 2007. Конспект флоры Карелии. Петрозаводск: КарНЦ РАН. 403 с.
[Kryshen] Крышень А. М. 2006. Растительные сообщества вырубок Карелии. М. 262 с.
[Kryshen] Крышень А.М. 2010. Типы лесорастительных условий на автоморфных почвах в Карелии. – Бот. журн. 95 (3): 281–297.
[Kryshen] Крышень А.М., Геникова Н.В., Гнатюк Е.П., Преснухин Ю.В., Ткаченко Ю.Н. 2018. Ряды восстановления сосняков Восточной Фенноскандии на песчаных автоморфных почвах. – Бот. журн. 103 (1): 5–35.
[Kryshen] Крышень А.М., Полевой А.В., Гнатюк Е.П., Кравченко А.В., Кузнецов О.Л. 2009. База данных местообитаний (биотопов) Карелии. – Труды КарНЦ РАН. 4: 3–10.
[Methods] Методы исследования ценофлор (на примере растительных сообществ вырубок Карелии). 2005. Петрозаводск. 68 с.
[Red…] Красная книга Республики Карелия. 2007. Петрозаводск. 368 с.
Selmants P.C., Knight D.H. 2003. Understory plant species composition 30–50 years after clearcutting in southeastern Wyoming coniferous forests. – Forest Ecology and Management. 185 (3): 275–289.
[Shmidt] Шмидт В.М. 1980. Статистические методы в сравнительной флористике. Л. 176 с.
Tonteri T. 1994. Species richness of boreal understorey forest vegetation in relation to site type and successional factors. – Ann. Zoo. Fennici. 31: 53–60.
Widenfalk O., Weslien J. 2009. Plant species richness in managed boreal forests. Effects of stand succession and thinning. – Forest Ecology and Management. 257 (5): 1386–1394.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал