

Ботанический журнал, 2019, T. 104, № 5, стр. 766-780
FOUR NEW SPECIES OF CYMBELLA (BACILLARIOPHYTA) FROM THE ANCIENT MALILI LAKES (SULAWESI ISLAND, INDONESIA)
D. A. Kapustin 1, *, J. P. Kociolek 2, 3, A. M. Glushchenko 4, M. S. Kulikovskiy 4, **
1 Papanin Institute for Biology of Inland Waters RAS Borok
152742 Nekouz Distr., Yaroslavl Region, Russia
2 Museum of Natural History, University of Colorado
80309 Boulder, Colorado, USA
3 Department of Ecology and Evolutionary Biology University of Colorado
80309 Boulder, Colorado, USA
4 K.A. Timiryazev Institute of Plant Physiology RAS, IPP RAS
127276 Moscow, Botanicheskaya Str., 35, Russia
* E-mail: dima_kapustin@outlook.com
** E-mail: max-kulikovsky@yandex.ru
Поступила в редакцию 23.11.2018
После доработки 06.04.2019
Принята к публикации 09.04.2019
Аннотация
Four new diatom species from the genus Cymbella Agardh are described on the basis of light and scanning microscopy. These are Cymbella eunotioformis sp. nov., C. densigranulata sp. nov., C. khursevichiae sp. nov. and C. margaritae sp. nov. All species are described from ancient Malili lake system in Indonesia. Morphology and comparison with similar species described previously by Hustedt from the same lakes are discussed as possible sister species. Our data show that species diversity of diatoms in general, and cymbelloid diatoms specifically, are not yet completely known in this ancient lake system. It is possible that the freshwater cymbelloid diatoms with apical pore fields, and the genus Cymbella specifically, may have originated in Asia.
The Malili lakes are located in the mountains of Central Sulawesi and are composed of five tectonic lakes namely Matano, Mahalona, Towuti, Lontoa (also known as Wawontoa) and Masapi (Brooks, 1950; Bramburger et al. 2004; Vaillant et al. 2011; von Rintelen et al. 2012). The former three lakes are directly connected to one another, while lakes Lontoa and Masapi are only indirectly joined with the other three lakes.
New species of freshwater diatoms were published first by Hustedt (in Schmidt 1934) later as part of a larger treatise on planktonic diatoms (Hustedt, 1942a). The first comprehensive study of the diatoms of the Malili lakes was made by Hustedt (1942b). He described among others 19 new taxa of Cymbella C. Agardh (1830: 1) sensu lato. Krammer reclassified them between the genera Cymbella sensu stricto, Encyonema Kützing (1833: 589), Encyonopsis Krammer (1997a: 156) and Cymbopleura (Krammer) Krammer (1999: 284) and additionally described several new taxa based on Hustedt’s types (Krammer 1997a,b; 2002, 2003). Bramburger et al. (2004) compiled the list of diatoms from the Malili lakes according to the changes in taxonomy. Recently, Kapustin et al. (2017) established new genus Celebesia D. Kapustin, Kulikovskiy et Kociolek (2017: 153) to accommodate Cymbella distinguenda Hustedt (1942: 111).
In recent years new diatom investigation were done and a number of new endemic diatom taxa have been described from Southeast Asia (Chudaev et al. 2018; Glushchenko et al., 2016; 2017a-c; 2018a,b; Glushchenko, Kulikovskiy, 2015, 2017a-d; Gusev, Kulikovskiy, 2014; Kapustin, Kulikovskiy, 2018; Kociolek et al., 2015, 2018; Kulikovskiy et al., 2015, 2018a, 2019). New diatom genera namely Tetralunata Hamsher, Graeff, Stepanek et Kociolek (2014: 361), Celebesia D. Kapustin, Kulikovskiy et Kociolek (2017: 153), Sinoperonia Kociolek, Liu, Glushchenko et Kulikovskiy (Liu et al., 2018: 149), Okhapkinia Glushchenko, Kulikovskiy et Kociolek (Kulikovskiy et al., 2018b: 122), Dorofeyukea Kulikovskiy, Maltsev, Andreeva et Kociolek (Kulikovskiy et al., 2019: 4) and Karthickia Kociolek, Glushchenko, Kulikovskiy in Glushchenko et al. (submitted) were described. We have shown that Asia is the only continent that supports endemic genera of freshwater cymbelloid diatoms (Kociolek, 2018).
In this paper, the second a series which aims to revise the cymbelloid diatoms from the Malili lakes base on the recent materials and using light and scanning electron microscopy (see Kapustin et al., 2017). Herein we formally describe four new species of Cymbella sensu stricto from lakes Matano and Towuti.
MATERIALS AND METHODS
Species described in this article were investigated from the epilithic samples which were collected from Lake Matano on November, 14th 2014 and from Lakes Matano and Towuti on September, 23th and 24th 2015 respectively. Environmental variables of the sample sites are summarized in Table 1.
Table 1.
Coordinates, collection date and environmental variables of each site
| Lake | Sampling date | Slide number | Geographic coordinates | pH | Tempera- ture (°C) | Specific conduc- tivity (μS cm–1) |
|---|---|---|---|---|---|---|
| Matano | 14 Nov 2014 | 01105 | S02°28.433', E121°15.710' | 8.53 | 28.5 | 177 |
| Matano | 23 Sept 2015 | 03981 | S02°25.703', E121°15.377' | 8.36 | 28.2 | 187 |
| Towuti | 24 Sept 2015 | 03923 | S02°39.864', E121°31.767' | 8.36 | 30.7 | 151 |
The samples were boiled in concentrated hydrogen peroxide (≈37%) to dissolve the organic matter. Then they were washed with deionized water four times at 12 h intervals. After decanting and filling with deionized water up to 100 ml, the suspension was spread onto coverslips and left to dry at room temperature. Permanent diatom slides were mounted in Naphrax®. Light microscopic (LM) observations were performed with a Zeiss Scope A1 microscope equipped with an oil immersion objective (100×, n.a. 1.4, differential interference contrast [DIC]) and Zeiss AxioCam ERc 5s camera. Valve ultrastructure was examined by means of a JSM-6510LV scanning electron microscope (Institute for Biology of Inland Waters RAS, Borok, Russia). For scanning electron microscopy (SEM), parts of the suspensions were fixed on aluminum stubs after air-drying. The stubs were sputter coated with 50 nm of gold. Types are deposited at the I.D. Papanin Institute for Biology of Inland Waters, Russian Academy of Sciences (IBIW), Borok, Russia.
RESULTS
Cymbella eunotioformis D. Kapustin, Kociolek et Kulikovskiy sp. nov. (Table I, 1–6; II, 1–6)
Table I. LM, DIC. Size diminution series, valve views.
1–6 – Cymbella eunotioformis D. Kapustin, Kociolek et Kulikovskiy sp. nov.; 2 is of the holotype. 7–12 – Cymbella margaritae D. Kapustin, Kociolek et Kulikovskiy sp. nov.; 6 is of the holotype. Scale bars – 10 µm.

Description
LM. Valves are dorsiventral with a convex dorsal and almost straight or slightly convex ventral margin. The poles are subcapitate to capitate. Length 36–51 µm, width 7.0–9.5 µm, length/width ratio 5.06–5.37. The axial area is narrow linear, the central area is not differentiated. A stigma is present, but indistinct, placed at the end of the central stria on the ventral margin. The raphe is lateral, becoming reverse-lateral towards the center and near the poles. The striae are radiate throughout, being only slightly radiate at the valve center on the ventral side of the valve; they are strongly radiate and more dense at the apices. At the valve center, the ventral striae have a density of 10–11 in 10 µm and the dorsal striae have a density of 11–13 in 10 µm (Table I, 1–6).
SEM. Externally the proximal raphe ends are slightly curved towards the ventral margin. The distal raphe endings are deflected to the dorsal margin. A pore field with a hyaline area near the raphe slit on the valve face is located at the apices. There is a round or transapically elongated stigma in the central area, positioned within a small depression at the end of the central stria. The striae are composed of apically elongated, slit-like areolae. Striae are interrupted by marginal ribs, each of which has a small spine opposite the striae or costae. Internally, the costae are broad and the alveoli are narrow without struts. The stigma opening is positioned as part of the alveolus and occluded by fine ingrowths from their perimeter. The distal raphe endings terminate in helictoglossae, the proximal raphe endings appear to be hidden. The central nodule is weakly developed. Porelli of the apical pore fields are occluded (Table II, 1–6).
Table II. Cymbella eunotioformis D. Kapustin, Kociolek et Kulikovskiy sp. nov. SEM.
1 – external view of the valve; 2 – external view of the central area; 3 – external view of the valve pole; 4 – internal view of the valve; 5 – internal view of the central area; 6 – internal view of the valve pole. Scale bars: 1, 4 – 5 µm; 2, 3, 5, 6 – 1 µm.

Type: – INDONESIA, Sulawesi Island, Lake Matano, S02°25.703' E121°15.377', collection of Maxim Kulikovskiy, I.D. Papanin Institute for Biology of Inland Waters, Russian Academy of Sciences (IBIW), holotype here designated, slide no 03981 = Table I, 2.
Etymology: – Species epithet refers to the similarity of the valve outline of this species with Eunotia.
Cymbella densigranulata D. Kapustin, Kociolek et Kulikovskiy sp. nov. (Table III, 1–9; Table IV, 1–5)
Table III. Cymbella densigranulata D. Kapustin, Kociolek et Kulikovskiy sp. nov.
LM, DIC, size diminution series, valve views (1–5), SEM (6–9); 6–9 – external view of the valve; 9 – valve in a girdle view; 2 is of the holotype. Scale bars: 14–20 – 10 µm; 21 – 5 µm.

Table IV. Cymbella densigranulata D. Kapustin, Kociolek et Kulikovskiy sp. nov. SEM.
1 – external view of the central area; 2 – external view of the valve pole; 3 – internal view of the valve; 4 – internal view of the central area; 5 – internal view of the valve pole. Scale bars: 24 – 5 µm; 22, 23, 25, 26 –1 µm.
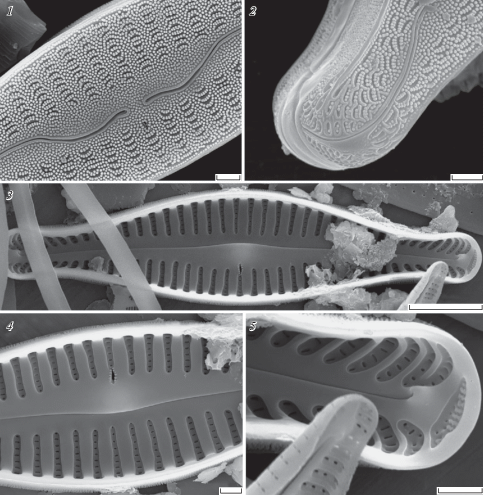
Description
LM. Valves are slightly dorsiventral, fusiform (subelliptic) with protracted subcapitate poles. Length 30–35 µm, width 6.5–7.5 µm, length/width ratio 4.0–4.77. The axial area is narrow, linear. The central area is not distinguishable from the axial area. A stigma is present, positioned at the end of the central stria on the ventral margin of the valve. The raphe is lateral, undulate, becoming reverse-lateral towards the center and filiform near the poles. The striae are parallel to slightly radiate, becoming more strongly radiate near the poles. At the valve center, the ventral striae have a density of 10–12 in 10 µm and the dorsal striae have a density of 10–11 in 10 µm (Table III, 1–9).
SEM. Externally the raphe is shown to be highly undulate along its length, ventrally curved towards the valve center, with simple, very slightly-widened proximal endings, which are slightly deflected ventrally. The distal raphe endings are deflected to the dorsal side, terminating to the dorsal margin relative to the apical pore fields. There is a round or transapically elongated stigma at the end of the central stria on the ventral margin. The striae are composed of apically elongated, slit-like areolae. The striae between the valve face and mantle are interrupted by the marginal rib. The whole valve surface and the mantle are densely covered with granules. The poles are additionally ornamented with short curved ridges. Internally the costae are broad and the alveoli narrow without struts. The stigma opening is occluded by fine ingrowths from their perimeter. The distal raphe endings terminate in helictoglossae, while the proximal raphe endings are hidden. Porelli of the apical pore fields are occluded (Table III, 6–9; IV, 1–5).
Type: – INDONESIA, Sulawesi Island, Lake Matano, S02°28.433', E121°15.710', collection of Maxim Kulikovskiy, I.D. Papanin Institute for Biology of Inland Waters, Russian Academy of Sciences (IBIW), holotype here designated, slide no 01105 = Table III, 2.
Etymology: – Species epithet refers to the dense granulate valve surface of this species.
Cymbella khursevichiae D. Kapustin, Kociolek et Kulikovskiy sp. nov. (Table V, 1–10; VI, 1–6).
Table V. Cymbella khursevichiae D. Kapustin, Kociolek et Kulikovskiy sp. nov. LM, DIC. Size diminution series, valve views (1–10). 1 is of the holotype. Scale bar – 10 µm.

Description
LM. Valves are dorsiventral with rounded apices. Dorsal margin moderately arched and the ventral margin is tumid only at the valve center. Length 43–70 µm, width 15.5–20 µm, length/width ratio 2.69–3.68. The axial area is narrow lanceolate, gradually widening into a broad semicircle central area. The raphe is lateral, becoming reverse-lateral towards valve the center and filiform near the poles. The striae are slightly radiate, becoming strongly radiate near the poles. At the valve center, the ventral striae have a density of 8 in 10 µm and the dorsal striae have a density of 7 in 10 µm; near the poles the striae have a density of 9 in 10 µm. 2–4 stigmata are present on the ventral side of the valve (Table V, 1–10).
SEM. Externally the raphe is ventrally curved towards the valve center. The proximal raphe endings are slightly dorsally deflected and the central pores are dilated and bent ventrally. The distal raphe endings are deflected to the dorsal side. A relatively small apical pore field is located at the pole apices. It composed of round and elongated porelli and separated from the raphe endings by wide hyaline area. There are apically elongated stigmata which are hardly distinguishable from areolae in the central area. The striae are composed of apically elongated, slit-like areolae (Table VI, 1–6).
Table VI. Cymbella khursevichiae D. Kapustin, Kociolek et Kulikovskiy sp. nov. SEM.
1 – external view of the valve; 2 – external view of the central area; 3 – external view of the valve pole; 4 – internal view of the valve; 5 – internal view of the central area; 6 – internal view of the valve pole. Scale bars: 1, 4 –10 µm; 2, 5, 6 – 2 µm; 3 –1 µm.

Type: – INDONESIA, Sulawesi Island, Lake Towuti, S02°39.864', E121°31.767', collection of Maxim Kulikovskiy, I.D. Papanin Institute for Biology of Inland Waters, Russian Academy of Sciences (IBIW), holotype here designated, slide no 03923 = Table V, 1.
Etymology: – The species is dedicated to well-known diatomologist Prof. Galina Khursevich.
Cymbella margaritae D. Kapustin, Kociolek et Kulikovskiy sp. nov. (Table I, 7–12; VII, 1–6).
Description
LM. Valves are dorsiventral with narrow protracted apices. Dorsal margin moderately arched and the ventral margin is convex. Length 46–59 µm, width 14–18 µm, length/width ratio 2.88–4.2. The axial area is narrow and the central area is barely differentiated. The striae are slightly radiate, becoming strongly radiate near the poles. At the valve center, the ventral striae have a density of 9–12 in 10 µm and the dorsal striae have a density of 9–10 in 10 µm; near the poles the striae have a density of 11–13 in 10 µm. A single stigma is present on the ventral side of the valve (Table I, 7–12).
SEM. Externally raphe branches are slightly undulate and ventrally curved. The proximal raphe endings are slightly widened and ventrally deflected. The distal raphe endings are deflected to the dorsal side. A relatively small apical pore field is separated from the raphe slit by a hyaline area. A transapically elongated stigma is located in a small depression in the central area. The areola openings are apically elongated and slit-like. Internally, the stigmal opening has edges with a cracked appearance. The internal distal proximal raphe ends terminate in small helictoglossae (Table VII, 1–6).
Table VII. Cymbella margaritae D. Kapustin, Kociolek et Kulikovskiy sp. nov. SEM.
1 – external view of the valve; 2 – external view of the central area; 3 – external view of the valve pole; 4 – internal view of the valve; 5 – internal view of the central area; 6 – internal view of the valve pole. Scale bars: 1 – 5 µm; 2, 3, 5 – 1 µm; 4 – 10 µm; 6 – 0.5 µm.
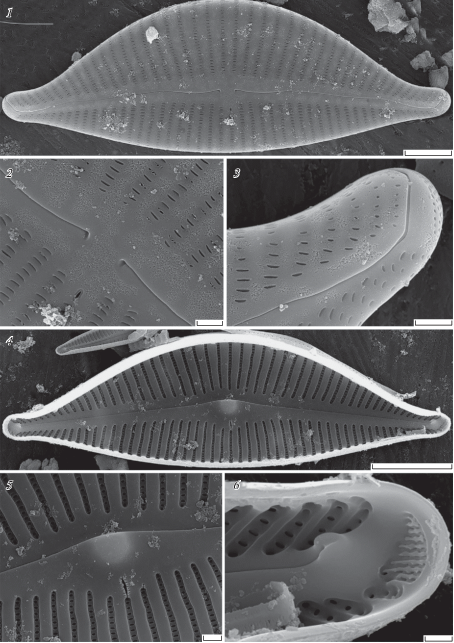
Type: – INDONESIA, Sulawesi Island, Lake Matano, S02°25.703' E121°15.377', collection of Maxim Kulikovskiy, I.D. Papanin Institute for Biology of Inland Waters, Russian Academy of Sciences (IBIW), holotype here designated, slide no 03981= Table I, 6.
Etymology: – The species is named after Margarita Faradzheva for her help in managing and sampling during expedition to Indonesia.
DISCUSSION
In general, cymbelloid diatoms of the Malili lakes are not well known, and our description of four new species is evidence that there are still species new to science to be found in these lakes. A summary of the cymbelloid diatoms described from the Malili lakes is presented in Table 2. The description of new species herein indicates that the Malili lakes still contain not only endemic species, but also species complexes that need to be verified, revised and described. All four newly-described species are similar to Hustedt’s taxa previously described from the Malili lakes (Hustedt, 1942b) but can be easily distinguished from them. In many cases the comparisons are complicated because many of the descriptions of freshwater Indonesian diatoms by Hustedt were based on a single valve only.
Table 2.
Summary of cymbelloid taxa described originally from the Malili lakes
| Current name | Original name | References |
|---|---|---|
| Encyonopsis mendosa (Van Lan-dingham) Da Silva et Menezes | Cymbella subalpina Hustedt | Da Silva, Menezes (2016) |
| Encyonopsis brevistriata (Hustedt) Krammer | Cymbella subalpina f. brevistriata Hustedt | Krammer (1997b) |
| Encyonopsis apiculata (Hustedt) Da Silva et Menezes | Cymbella subalpina f. apiculata Hustedt | Da Silva, Menezes (2016) |
| Encyonopsis dubitata (Cholnoky) Krammer | Cymbella subalpina var. dubia Hustedt | Krammer (1997b) |
| Encyonopsis cryptocephala (Hustedt) Krammer | Cymbella cryptocephala Hustedt | Krammer (1997b) |
| Encyonema tenuissimum (Hustedt) D.G. Mann | Cymbella tenuissima Hustedt | Krammer (1997a) |
| Encyonopsis cesatii var. undulata (Hustedt) Krammer | Cymbellacesatii f. undulata Hustedt | Krammer (1997b) |
| Cymbella pseudodelicatula Hustedt | Cymbella pseudodelicatula Hustedt | Krammer (2003) |
| Cymbella leptoceroides Hustedt | Cymbella leptoceroides Hustedt | Krammer (2002) |
| Cymbella leptoceroides var. sublanceolata Krammer | Cymbella leptoceroides Hustedt pro parte | Krammer (2002) |
| Cymbella towoetensis Krammer | Cymbella leptoceroides Hustedt pro parte | Krammer (2002) |
| Cymbopleura malayensis (Hustedt) Krammer | Cymbella malayensis Hustedt | Krammer (2003) |
| Cymbella malayensis f. rostrata Hustedt | Cymbella malayensis f. rostrata Hustedt | Krammer (2003) |
| Cymbella bicapitata Hustedt | Cymbella bicapitata Hustedt | Krammer (2003) |
| Cymbopleura ampullacea (Hustedt) Krammer | Cymbella ampullacea Hustedt | Krammer (2003) |
| Cymbopleura ampullacea var. lata Krammer | Cymbella ampullacea Hustedt pro parte | Krammer (2003) |
| Encyonema wallaceanum (Hustedt) Krammer | Cymbella subturgida var. wallaceana Hustedt | Krammer (1997b) |
| Cymbella gracillima Hustedt | Cymbella gracillima Hustedt | Krammer (2003) |
| Cymbella wolterecki Hustedt | Cymbella wolterecki Hustedt | Krammer (2002) |
| Cymbella wolterecki var. capitata Hustedt | Cymbella wolterecki var. capitata Hustedt | Krammer (2002) |
| Celebesia distinguenda (Hustedt) D. Kapustin et al. | Cymbella distinguenda Hustedt | Kapustin et al. (2017) |
| Cymbella mirabilis Hustedt | Cymbella mirabilis Hustedt | Krammer (2003) |
| Encyonopsis spiculiformis Krammer | Cymbella spicula Hustedt pro parte | Krammer (1997b) |
In the cases of the species we describe in the present report, three of four of them are very similar to taxa described by Hustedt in his 1942(b) flora. Differences between taxa may be by a single feature, or size metric (e.g. breadth), which might suggest Hustedt’s taxon and the new taxon proposed here are sister species, derived from a recent common ancestor.
For example, Cymbella eunotioformis sp. nov. resembles C. bicapitata Hustedt (1942b: 103–104) in general appearance and in having capitate apices. However, in C. bicapitata the dorsal margin is strongly arched and the ventral margin is convex, whereas in C. eunotioformis sp. nov. the dorsal margin is convex and the ventral margin is almost straight. Both taxa have an indistinct stigma. Simonsen (1987: 284) pointed out that “the “stigma” observed by Hustedt appears to be only a more or less isolated areola of the central stria, its optical reaction does not differ at all from the stria”. We do not know how C. bicapitata varies during the life cycle but we could not observe specimens with arched dorsal margin in C. eunotioformis sp. nov. and we believe that it represents a separate species.
A second example of possible sister species is between C. densigranulata sp. nov. and C. ampullacea Hustedt, which resemble one another with respect to valve outline. However, C. ampullacea is wider and has more strongly arched margins and more protracted ends than C. densigranulata sp. nov. (Hustedt, 1942b; Krammer, 2003). Significant differences between this species are visible under SEM. Cymbella ampulacea lacks granules and has spines on the marginal ridge as seen in C. eunotioformis sp. nov. (unpublished observations).
In the third example, Cymbella margaritae sp. nov. is similar to C. malayensis f. rostrata Hustedt (1942b: 103) but has more progressively tapering narrowly rounded ends, whereas Hustedt’s taxon has broadly rounded ends which tapering abruptly. Additionally, C. malayensis f. rostrata lacks a stigma which is present in C. margaritae sp. nov. (Hustedt, 1942b; Simonsen, 1987). Interestingly, Krammer transferred the type forma, C. malayensis f. malayensis Hustedt (1942b: 102–103), to the genus Cymbopleura under the name Cymbopleura malayensis (Hustedt) Krammer (2003: 45) but he decided to keep Cymbella malayensis f. rostrata in the genus Cymbella.
The last species we described here, C. khursevichiae sp. nov., can be easily distinguished from all known Cymbella taxa by its general appearance and semicircular central area. C. gemeinhardtii Metzeltin et Krammer (Krammer, 2002: 129) has a very similar valve shape but it differs from C. khursevichiae sp. nov. in having very large central nodule, wide axial and central area and another raphe morphology. Cymbella khursevichiae sp. nov. was probably formerly lumped as Cymbella leptoceroides Hustedt (Hustedt, 1942b; Simonsen, 1987; Krammer, 2002) but no taxon from this species complex has such remarkable central and expanded axial areas.
Under SEM both C. eunotioformis sp. nov. and C. densigranulata sp. nov. have an interesting feature, a marginal rib which interrupts the striae as the extends between the valve face and mantle. Formerly, the marginal rib was known in Oricymba Jüttner, Krammer, Cox, Van de Vijver et Tuji in Jüttner et al. (2010: 408) and Celebesia (Kapustin et al., 2017) only; and these two genera are endemics to Asia. With the occurrence of a marginal rib in these two Cymbella species from Indonesia, two conclusions might be drawn. First, the feature of the marginal rib is a primitive one in the apical pore field-bearing cymbelloid diatoms. Second, in this group, C. eunotioformis sp. nov. and C. densigranulata sp. nov. may be very primitive members of Cymbella, and the genus may have originated in Asia. These data support the idea that Asia may be important to understanding the origin and diversification of the freshwater Cymbellaceae (Kociolek, 2018).
Список литературы
Agardh C.A. 1830. Conspectus Criticus Diatomacearum. P. I. Litteris Berlingianis, Lundae. P. 1–16.
Bramburger A.J., Haffner G.D., Hamilton P.B. 2004. Examining the distributional patterns of the diatom flora of the Malili lakes, Sulawesi, Indonesia. In: Poulin, M. (ed.) Proceedings of the 17th International Diatom Symposium. Biopress Ltd., Bristol. P. 11–25.
Brooks J.L. 1950. Speciation in ancient lakes (concluded). – The Quarterly Review of Biology 25: 131–176.
Chudaev D.A., Glushchenko A.M., Blagoveshchenskaya E.Yu., Karthick B., Kulikovskiy M.S. 2018. Morphology and taxonomy of Navicula escambia and N. simulata (Bacillariophyceae). – Nova Hedwigia, Beiheft 147: 43–57. https://doi.org/10.1127/nova-suppl/2018/006
Glushchenko A.M., Genkal S.I., Kulikovskiy M.S. 2016. Aulacoseira konstantinovii sp. nov. and Aulacoseira krylovii sp. nov. – two new centric diatoms from South east Asia. – Diatom Research 31: 367–378. https://doi.org/10.1080/0269249X.2016.1253617
Glushchenko A.M., Kulikovskiy M.S. 2015. The genus Luticola (Bacillariophyceae) in water ecosystems of South-Eastern Asia. – Bot. Zhurn. 100 (8): 799–804 (In Russ.). https://doi.org/10.1134/S0006813615080037
Glushchenko A.M., Kulikovskiy M.S. 2017a. Amphipleura vavilovii: a new diatom species of the family Amphipleuraceae from Laos. – Inland Water Biology 10: 17–21. https://doi.org/10.1134/S1995082916040064
Glushchenko A.M., Kulikovskiy M.S. 2017b. Taxonomy and distribution of Eunotia camelus Ehrenberg (Bacillariophyta) infraspecific taxa from South-East Asia. – Bot. Zhurn. 102 (3): 329–339. (In Russ.). https://doi.org/10.1134/S0006813617030036
Glushchenko A.M., Kulikovskiy M.S. 2017c. Taxonomy and distribution of the genus Eunotia Ehrenberg in water ecosystems of Vietnam. – Inland Water Biology 10 (2): 130–139. https://doi.org/10.1134/S1995082917020055
Glushchenko A.M., Kulikovskiy M.S., Kociolek J.P. 2017a. New diatom species from Gomphonema subtile-group in Southeast Asia. – Phytotaxa 329 (3): 223–323. https://doi.org/10.11646/phytotaxa.329.3.3
Glushchenko A.M., Kulikovskiy M.S., Kociolek J.P. 2017b. New and interesting species from the genus Luticola (Bacillariophyceae) in waterbodies of Southeastern Asia. – Nova Hedwigia, Beiheft 146: 157–173. https://doi.org/10.1127/1438-9134/2017/157
Glushchenko A.M., Kulikovskiy M.S., Kuznetsova I.V. 2018a. The genus Eunotia Ehrenberg in waterbodies of South-East Asia. Filigran, Yaroslavl. 211 p. (In Russ.).
Glushchenko A.M., Kulikovskiy M.S., Kuznetsova I.V., Dorofeyuk N.I., Kociolek J.P. 2018b. New species and combinations in the genus Eunotia Ehrenberg 1837 (Bacillariophyceae: Eunotiaceae) from waterbodies of Southeastern Asia. – Nova Hedwigia, Beiheft 147: 69–103. https://doi.org/10.1127/nova-suppl/2018/009
Glushchenko A.M., Kulikovskiy M.S., Kuznetsova I.V., Kociolek J.P. (Submitted) Karthickia verestigmata gen. et sp. nov. – an interesting diatom with frustular morphology similar to several different cymbelloid diatom genera. – Phycologia.
Glushchenko A.M., Kulikovskiy M.S., Okhapkin A.G., Kociolek J.P. 2017d. Aneumastus laosica sp. nov. and A. genkalii sp. nov. – two new diatom species from Laos (Indochina) with comments on the biogeography of the genus. – Cryptogamie, Algologie 38 (3): 183–199. https://doi.org/10.7872/crya/v38.iss3.2017.183
Gusev E.S., Kulikovskiy M.S. 2014. Centric diatoms from Vietnam reservoirs with description of one new Urosolenia species. – Nova Hedwigia, Beiheft 143: 111–126. https://doi.org/10.1127/1438-9134/2014/006
Hamsher S.E., Graeff C.L., Stepanek J.G., Kociolek J.P. 2014. Variation in valve and girdle band morphology in freshwater Denticula (Bacillariophyceae) species: implications for the systematic position of the genus including the description of Tetralunata gen. nov. (Epithemiaceae, Rhopalodiales). – Plant Ecology and Evolution 147 (3): 346–365. https://doi.org/10.5091/plecevo.2014.990
Hustedt F. 1942a. Das Phytoplankton des Süsswässers. – In: Thienemann A., Die Binnengewässer Einzeldarstellungen aus der Limnologie und ihren Nachbargebieten, E. Schweizerbart’sche Verlagsbuchandlung, Stuttgart 16 (2): 367–549.
Hustedt F. 1942b. Süßwasser-Diatomeen des indomalayischen Archipels und der Hawaii-Inseln. – Internationale Revue der gesamten Hydrobiologie und Hydrographie 42 (1/3): 1–252. https://doi.org/10.1002/iroh.19420420102
Jüttner I., Krammer K., van de Vijver B., Tuji A., Simkhada A., Gurung S., Sharma S., Sharma C., Cox E.J. 2010. Oricymba (Cymbellales, Bacillariophyceae), a new cymbelloid genus and three new species from the Nepalese Himalaya. – Phycologia 49: 407–423. https://doi.org/10.2216/09-77.1
Kapustin D.A., Kulikovskiy M.S. 2018. Transfer of Stenopterobia and Surirella taxa (Bacillariophyceae) described from the insular Southeast Asia to the genus Iconella. – Nova Hedwigia, Beiheft 147: 237–245. https://doi.org/10.1127/nova-suppl/2018/019
Kapustin D.A., Kulikovskiy M.S., Kociolek J.P. (2017) Celebesia gen. nov., a new cymbelloid diatom genus from the ancient Lake Matano (Sulawesi Island, Indonesia). – Nova Hedwigia, Beiheft 146: 147–155. https://doi.org/10.1127/1438-9134/2017/147
Krammer K. 1997a. Die cymbelloiden Diatomeen. Eine Monographie der weltweit bekannten Taxa. Teil 1. Allgemeines und Encyonema Part. – Bibliotheca Diatomologica 36: 1–382.
Krammer K. 1997b. Die cymbelloiden Diatomeen. Eine Monographie der weltweit bekannten Taxa. Teil 2. Encyonema part., Encyonopsis und Cymbellopsis. – Bibliotheca Diatomologica 37: 1–470.
Krammer K. 1999. Validierung von Cymbopleura nov. gen. – Iconographia Diatomologica 6: 1–284.
Krammer K. 2002. Cymbella. – Diatoms of Europe 3: 1–584.
Krammer K. 2003. Cymbopleura, Delicata, Navicymbula, Gomphocymbellopsis, Afrocymbella. – Diatoms of Europe 4: 1–530.
Kociolek J.P. 2018. A worldwide listing and biogeography of freshwater diatom genera: A phylogenetic perspective. – Diatom Research.33(4): 509–534. https://doi.org/10.1080/0269249X.2019.1574243
Kociolek J.P., Glushchenko A.M., Kulikovskiy M.S. 2015. Typification, valve ultrastructure, and systematic position of Gomphonema gomphopleuroides Amossé ex Kociolek, Glushchenko & Kulikovskiy, an endemic diatom from Southeast Asia. – Diatom Research 30: 247–255. https://doi.org/10.1080/0269249X.2015.1072583
Kociolek J.P., Kapustin D.A., Kulikovskiy M.S. 2018. A new, large species of Gomphonema Ehrenberg from ancient Lake Matano, Indonesia. – Diatom Research 33: 241–250. https://doi.org/10.1080/0269249X.2018.1513868
Kulikovskiy M.S., Glushchenko A.M., Dorofeyuk N.I., Kociolek J.P. 2018a. Morphology and distribution of Ninastrelnikovia laosica sp. nov. – a new species in a previously monospecific genus. – Nova Hedwigia, Beiheft 147: 119–126. https://doi.org/10.1127/nova-suppl/2018/011
Kulikovskiy M.S., Glushchenko A.M., Kociolek J.P. 2015. The diatom genus Oricymba in Vietnam and Laos with description of one new species, and a consideration of its systematic placement. – Phytotaxa 227: 120–134. https://doi.org/10.11646/phytotaxa.227.2.2
Kulikovskiy M.S., Glushchenko A.M., Kuznetsova I.V., Kociolek J.P. 2018b. Description of the new freshwater diatom genus Okhapkinia gen. nov. from Laos (Southeast Asia), with notes on family Sellaphoraceae Mereschkowsky 1902. – Fottea, Olomouc 18 (1): 120–129. https://doi.org/10.5507/fot.2017.021
Kulikovskiy M.S., Maltzev Ye.I., Andreeva S.A., Glushchenko A.M., Gusev E.S., Podunay Y., Ludwig T.V., Tusset E., Kociolek J.P. 2019. Description of a new diatom genus Dorofeyukea gen. nov. with remarks on phylogeny of the family Stauroneidaceae. – Journal of Phycology 55 (1): 173–185. https://doi.org/10.1111/jpy.12810
Kützing F.T. 1833. Synopsis Diatomearum oder Versuch einer systematischen Zusammenstellung der Diatomeen. – Linnaea 8 (5): 529–620. https://doi.org/10.5962/bhl.title.65634
Maltsev Y.I., Andreeva S.A., Kulikovskiy M.S., Kociolek J.P. 2017. Molecular phylogeny of the diatom genus Envekadea (Bacillariophyceae, Naviculales). – Nova Hedwigia, Beiheft 146: 241–252. https://doi.org/10.1127/1438-9134/2017/241
Rintelen T. von, Rintelen K. von, Glaubrecht M., Schubart C.D., Herder F. 2012. Aquatic biodiversity hotspots in Wallacea: the species flocks in the ancient lakes of Sulawesi, Indonesia. – In: Gower D.J. (eds). Biotic Evolution and Environmental Change in Southeast Asia. Cambridge University Press, Cambridge. P. 290–315.
Schmidt A. 1934. Atlas der Diatomaceen-kunde. Series VIII, Pls 393–400. O.R. Reisland, Leipzig.
Simonsen R. (1987) Atlas and catalogue of the diatom types of Friedrich Hustedt. Volume 3. – J. Cramer, Berlin-Stuttgart. 620 p.
Vaillant J.J., Haffner G.D., Cristescu M.E. 2011. The Ancient Lakes of Indonesia: Towards Integrated Research on Speciation. – Integrative and Comparative Biology 51: 634–643. https://doi.org/10.1093/icb/icr101
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал