

Ботанический журнал, 2019, T. 104, № 5, стр. 741-765
ШИРОКОЛИСТВЕННЫЕ ЛЕСА ЮГО-ВОСТОКА ТУЛЬСКОЙ ОБЛАСТИ
Ю. А. Семенищенков 1, *, Е. М. Волкова 2, **, О. В. Бурова 3, ***
1 Брянский государственный университет им. акад. И.Г. Петровского
241050 г. Брянск, ул. Бежицкая, 14, Россия
2 Тульский государственный университет
300012 г. Тула, пр. Ленина, 92, Россия
3 Государственный музей-заповедник “Куликово поле”
300041 г. Тула, пр. Ленина, 47, Россия
* E-mail: yuricek@yandex.ru
** E-mail: convallaria@mail.ru
*** E-mail: burova@kulpole.tula.net
Поступила в редакцию 11.04.2019
После доработки 14.05.2019
Принята к публикации 16.05.2019
Аннотация
Сохранившиеся мезофитные широколиственные леса юго-востока Тульской области, существенно трансформированные человеком, представляют собой характерную лесную растительность переходной полосы из зоны широколиственных лесов в лесостепь. С позиций подхода Ж. Браун-Бланке, они относятся к широко распространенной в Центральной России асс. Fraxino excelsioris-Quercetum roboris Bulokhov et Solomeshch 2003, объединяющей Среднерусско-Приволжские мезофитные леса широколиственно-лесной и лесостепной зон за пределами плакорного распространения ели. Сообщества ассоциации в изучаемом регионе разнообразны по составу ценофлор и объединены в 3 варианта и 3 фации, отражающие стадию сукцессии, характер нарушения лесов и имеющие выраженные экологические различия местообитаний. Анализ спектров геоэлементов и полизональных групп демонстрирует флористическую дифференциацию широколиственных лесов юго-востока Тульской области и расположенных юго-западнее регионов: северной части днепровского бассейна (зона широколиственных лесов) и бассейна Ворсклы (Центральная южная лесостепь). По составу ценофлоры и характеру ботанико-географического спектра леса изучаемого региона следует относить к типичной субассоциации (F.-Q. typicum), характерной для северной части ареала ассоциации.
ВВЕДЕНИЕ
Лесная растительность юго-восточной части Тульской области является результатом длительного антропогенного преобразования. Современная облесенность этой территории составляет менее 2% (Volkova, Burova, 2014), что требует охраны сохранившихся лесов. Однако территориальной охраной в составе ООПТ в области охвачена небольшая часть лесных массивов (Krasnaya…, 2007), а общее экологическое состояние растительности изучено недостаточно.
До периода активного преобразования человеком (XII–XIV вв.) на юго-востоке области на водоразделах преобладали остепненные луга и луговые степи в составе лесостепных ландшафтов. Лесная растительность формировалась в балках и речных долинах, реже – на водоразделах. Характерный для этого времени тип хозяйствования не приводил к существенным изменениям ландшафта (Gonyanyi et al., 2007).
В более позднее время лесостепные ландшафты подверглись сильной трансформации в результате формирования агроландшафтов, в первую очередь, вследствие распашки водораздельных пространств для расширения пахотных площадей. Это затронуло не только лугово-степные, но и лесные экосистемы. Результатом сельскохозяйственного освоения территории явилось снижение облесенности до 10% и менее (Burova, 2018). При этом в XIX–XX вв. некоторые ранее существовавшие лесные массивы были восстановлены лесными культурами, что позволило вторично сформироваться здесь лесной растительности. Среди таких массивов и мемориальная Зеленая Дубрава на ее исконном месте, известном по литературным свидетельствам (посажена в 1928 г. и имеется на картах межевания и землепользования 1941 г.), дубравы по коренному склону Дона у д. Татинки, балочные дубравы в долине р. Смолка (Зиновьев лес) и р. Нижний Дубик.
В настоящей статье дается общая характеристика наиболее крупных и значимых для сохранения флористического и фитоценотического разнообразия лесных массивов юго-востока Тульской области и приведены результаты классификации растительности мезофитных широколиственных лесов на основе подхода Ж. Браун-Бланке.
ПРИРОДНЫЕ УСЛОВИЯ РАЙОНА ИССЛЕДОВАНИЯ
Район исследования расположен на юго-востоке Тульской области, в северной части Среднерусской возвышенности. В морфоструктурном плане эта территория относится к Тульскому возвышенному карбоновому плато, в течение неотектонического этапа испытавшего устойчивое поднятие. Дочетвертичные коренные отложения, вскрытые в бортах долин Дона и Непрядвы, их притоков и в ряде балок, состоят из чередующихся известняковых и песчано-глинистых отложений нижнего карбона и песчаниковых с прослоями глин отложений мела.
Осадочный чехол местами представлен днепровскими флювиогляциальными отложениями (пески и алевриты с прослоями гравия, галечник, грубозернистый песок), а сверху повсеместно перекрыт покровными лессовидными суглинками, мощность которых колеблется от 1.5 до 5–6 м. Надпойменные террасы, речные поймы и днища балок слагают верхнечетвертичные отложения аллювиально-делювиального и озерно-болотного генезиса. Широко развиты делювиальные отложения, связанные с хозяйственной деятельностью человека, начавшейся здесь с конца XVI – начала XVII вв., перекрывающие практически всю пойму рек Дона, Непрядвы и их притоков, а также днища балок и ложбин. Их мощности на пойме – 0.3–1.0 м, а в отдельных балках – до 2 м.
Рельеф территории достаточно сложный и представлен доледниковой эрозионно-денудационной равниной, переработанной последующими геоморфологическими процессами (Glasko et al., 2005). Водоразделы выражены системой узких протяженных вершинных поверхностей с высотами 180–220 м. Водораздельные склоны с высотами от 170 до 200 м имеют сложную форму: преобладают участки длинных и протяженных склонов, как правило, прямые или слабо вогнутой формы с уклонами поверхности от 1.5 до 6°.
Речные долины глубоко врезаны – до 40–60 м, что обусловлено устойчивым поднятием территории в течение всего неотектонического этапа, и характеризуются попеременно-асимметричным поперечным профилем. В крупнейших речных системах района исследования надпойменно-террасовые комплексы Дона и Непрядвы слабо представлены. Наиболее четко прослеживаются первая и вторая надпойменные террасы верхнечетвертичного возраста. Высокие надпойменные террасы среднечетвертичного возраста – третья и четвертая, установленные только геологической съемкой. Днище речных долин занято поймой, имеющей сложный рельеф.
Водоразделы и водораздельные склоны расчленены долинно-балочной сетью. Для нижних и средних звеньев балок типичны прибалочные и балочные склоны с уклонами поверхности более 8°, осложненные оползнями, эрозионными промоинами; развиты донные взрезы. Верхние звенья балок представлены, в основном, плоскодонными лощинами и ложбинами стока (Glasko et al., 2005).
Климат территории умеренно-континентальный. Среднегодовое количество осадков – 600–750 мм. Среднегодовая температура воздуха – 8°С. Средняя температура июля составляет 18°С, января – –10°С.
Район исследования лежит в междуречье долин рек Дона и Непрядвы и их притоков: рек Курцы, Смолки, Нижнего и Среднего Дубиков, Мокрой и Сухой Таболы и др.
Почвенный покров представлен зональными типами почв северной лесостепи – черноземами (выщелоченные, оподзоленные и типичные) и серыми лесными, а также группой интразональных почв – аллювиальных пойменных, лугово-черноземных, болотных низинных.
По ботанико-географическому районированию, юго-восток Тульской области лежит в пределах Среднерусской (Верхнедонской) подпровинции Восточноевропейской лесостепной провинции Евразиатской степной области; у северо-восточной ее границы. Зональной растительностью являются широколиственные леса без участия ели и луговые степи (Isachenko, Lavrenko, 1980).
Начиная со второй половины XIX в. юго-восточная часть Тульской области была охвачена флористическими исследованиями, обзор которых выполнили И.С. Шереметьева с соавторами (Sheremetyeva et al., 2008). Специального геоботанического изучения лесной растительности этого региона никогда не проводилось. Следует отметить многотомную работу “Очерк флоры Епифанского уезда Тульской губернии” В.М. Голицына (Golitsyn, 1905, 1906, 1907), который в течение нескольких лет проводил флористические обследования лесов и прилегающих к ним участков в среднем течении р. Мокрая Табола с указанием состава и примерного возраста лесов. Сравнение описаний В.М. Голицына с современной картиной распространения лесов показывает, что большинство изучаемых лесных массивов сохранили свою локализацию. Тем не менее высоковозрастные насаждения здесь отсутствуют, так как были уничтожены в XX в.
Рис. 1.
Исследованные лесные урочища юго-востока Тульской области (показаны серой заливкой). Обозначения обследованных урочищ: 1 – Бучальский лес, 2 – Водяное поле, 3 – Исаковский лес, 4 – Грязновская балка, 5 – Лес Дубрава, 6 – Рыхотка, 7 – Стрельцы.
Fig. 1. Researched forest natural landmarks of southeastern Tula Region (shown by gray filling). Designation of teh studied landmarks: 1 – Buchal’sky Forest, 2 – Vodyanoye Polye, 3 – Isakovsky Forest, 4 – Gryasnovskaya ravine, 5 – Forest Dubrava, 6 – Rykhotka, 7 – Strel’tsy.
Обозначения с карты:
д. Казановка – village Kazanovka
д. Хворостянка – village Khvorostyanka
Климовский район – Klimovsky District
Куркинский район – Kurkinsky District
Кимовский район – Kimovsky District
с. Суханово – village Sukhanovo
с. Красное – village Krasnoe
с. Себино – village Sebino
р. Сухая Табола – Sukhaya Tabola River
р. Мокрая Табола – Mokraya Tabola River
р. Непрядва – Nepryadva River
р. Смолка – Smolka River
р. Дон –Don River
р. Рыхотка –Rykhotka River
Тульская область – Tula Region
Липецкая область – Lipetsk Region километры – kilometers
населенные пункты – settlements
леса – forests
реки – rivers
дороги – roads
административные границы – administrative boundaries
Рис. 2.
Спектры геоэлементов и полизональные группы мезофитных щироколиственных лесов юго-востока Тульской области (ЮВТ), северной части бассейна Верхнего Днепра (БВД) и бассейна Ворсклы (БВ). Обозначения геоэлементов: B — бореальный, Bs — суббореальный, N — неморальный, Sp — cубпонтический, Js — южносибирский, P — плюрирегиональный, Po — понтический; полизональных групп: Pct — субсредиземноморско-умеренная, Pcb — субсредиземноморско-бореальная, Pca — субсредиземноморско-арктическая, Pza — средиземноморско-арктическая, Pzb — средиземноморско-бореальная, Pzt — средиземноморско-умеренная (по: Булохов, Соломещ, 2003
Fig. 2. Spectra of geoelements and polyzonal groups of mesophyte broad-leaved forests of southeastern Tula Region (ЮВТ), southern part of the Upper Dnieper basin (БВД) and the Vorskla basin (БВ).
Designation of geoelements: B — boreal, Bs — subboreal, N — nemoral, Sp — subpontic, Js – South Siberian, P — pluriregional, Po — pontic; polyzonal groops: Pct — sub-Mediterranean-temperate, Pcb — sub-Mediterranean-boreal, Pca — sub-Mediterranean-arctic, Pza — Mediterranean-arctic, Pzb — Mediterranean-boreal, Pzt — Mediterranean-temperate (after Bulokhov, Solomeshch, 2003).
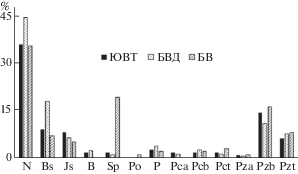
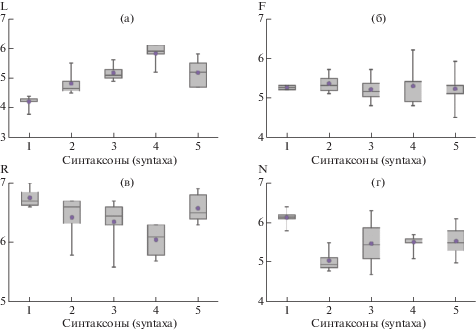
Рис. 3.
Экологические амплитуды местообитаний синтаксонов по ведущим экологическим факторам: а — освещенность (L), б — влажность почвы (F), в — кислотность почвы (R), г — обеспеченность почвы минеральным азотом (N) (определены по шкалам H. Ellenberg et al., 1992). Обозначения синтаксонов: 1 — асс. Fraxino excelsioris-Quercetum roboris typica var., 2 — асс. F. e.-Q. r. typica var. Tilia cordata fac., 3 — асс. F. e.-Q. r. typica var. Populus tremula fac., 4 — асс. F. e.-Q. r. Lysimachia nummularia var. Betula pendula var., 5 — асс. F. e.-Q. r. inops var.
Fig. 3. Ecological amplitudes of habitats of syntaxa by main ecological factors: a — illuminance (L), б — soil moisture (F), в — soil reaction (R), г — mineral nitrogen soil richness (N) (by H. Ellenberg et al., 1992). Designation of syntaxa: 1 — association Fraxino excelsioris-Quercetum roboris typica var., 2 — association F. e.-Q. r. typica var. Tilia cordata fac., 3 — association F. e.-Q. r. typica var. Populus tremula fac., 4 — association F. e.-Q. r. Lysimachia nummularia var. Betula pendula var., 5 — association F. e.-Q. r. inops var.
С начала прошлого века геоботаники и лесотипологи сосредоточили внимание на лесной растительности северо-западной “лесной” части Тульской области. При этом на юго-востоке области отмечались так называемые остепненные дубравы (Levitskii 1927; Alyokhin, 1947), которые в последнее десятилетие стали предметом специального изучения (Semenishchenkov et al., 2013; Semenishchenkov, Poluyanov, 2014). Как указывают И.С. Шереметьева и др. (Sheremetyeva et al., 2008), такие сообщества несут следы антропогенного воздействия (наличие пней, свидетельствующее о распространении в прошлом коренных пород, упрощенная структура насаждений); дубравы имеют порослевое происхождение или выращены на землях-“неудобьях”.
Следуя В.В. Алехину (Aljokhin, 1947), широколиственные леса являются зональным типом растительности юго-восточнее Оки на территории современной Тульской области. Однако уже в XVIII в. эти леса были практически полностью уничтожены, а на месте широколиственных насаждений отмечались “осиновые и березовые рощи” (Bolotov, 1766). Постоянное участие мелколиственных пород, по свидетельству С.Ф. Курнаева (Kurnaev, 1980), является прямым следствием сведения широколиственных лесов. Использование земель на их месте под пашню стало причиной исчезновения многих неморальных видов, которые хорошо сохранились только в пределах Тульских засек (Kurnaev, 1980). В настоящее время в изучаемом регионе господствуют березняки и осинники, возникшие на месте вырубленных широколиственных лесов или на залежах. Впоследствии мелколиственные породы замещаются дубом, липой, кленом и ясенем (Sheremetyeva et al., 2008).
ОБЩАЯ ХАРАКТЕРИСТИКА ИЗУЧАЕМЫХ ЛЕСНЫХ МАССИВОВ
Объектами нашего исследования стали лесные массивы в урочищах Бучальский лес, Водяное поле, Исаковский лес, Грязновская балка, Дубрава, Рыхотка, Стрельцы (рис. 1).
Исаковский лес площадью 197 га располагается на обширной части водораздельного пережима, образованного между верхними звеньями балочных комплексов Безымянного и Белого. По водораздельным склонам лесной массив спускается к долине р. Мокрая Табола. Бóльшая часть современного Бучальского леса лежит в верховье и вдоль склонов крупного долинно-балочного комплекса р. Веденец. Площадь леса в настоящее время составляет 1388 га. Грязновский лес (43 га) расположен в верховье одноименного крупного балочного комплекса в бассейне р. Дон. Леса Стрельцы (137 га) и Дубрава (106 га) сформировались в балках на водоразделе между реками Мокрая и Сухая Табола. Возможно, что в прошлом эти леса представляли собой единый массив (Себинский лес), который занимал водораздел р. Дон и его притоков рек Мокрая и Сухая Таболы. До XIV в. леса локально вырубались и использовались под подсеку. Позднее, до конца XVI в., антропогенная трансформация этих лесов была минимальна. В конце XVI – начале XVII вв. здешние земли были переданы в собственность землевладельцев, что привело к их массовой вырубке (Kulikovo…, 1999). В результате вместо единого лесного массива образовались отдельные лесные угодья, названные по близкому расположению тех или иных поселений или других топонимов: Бучальский лес (у д. Бучалки), Исаковский лес (у д. Исаковские выселки), Грязновский лес (в верховьях балки Грязновка), лес Стрельцы (балка в ур. Стрелицы и одноименная деревня), лес Дубрава (его исторические названия – Пугачевский лес у д. Пугачевка, или Краснинская роща). Дальнейшая судьба этих лесов связана с управлением землевладельцев и исполнением ими лесоохранительных законов того времени. Например, природопользование на землях крупных землевладельцев (князья Голицыны, графы Олсуфьевы) велось более прогрессивными методами, что сопровождалось оптимизацией лесов и лесовосстановлением (Bassein…, 1802, 1804). Считается, что это сыграло положительную роль в сохранности Исаковского леса. Другие хозяева земель относились к лесу потребительски, поскольку леса высокого бонитета имели более высокую рыночную стоимость. Результатом этого периода освоения явилось снижение естественной лесистости с 80% в начале XII в. до 15% – в начале-середине XIX в. и, начиная с конца XIX в., – до 5%. Тем не менее, все описываемые леса сохранили свое историческое местоположение и никогда окончательно не исчезали с исторических и современных карт, хотя с течением времени меняли свою конфигурацию – неуклонно уменьшалась их площадь.
Леса Водяное поле и Рыхотка располагаются на правобережье р. Дон, для которого была характерна более высокая “открытость” ландшафта (Glasko et al., 2005). Лес Водяное поле находится на склоне водораздела между балками, впадающими в долину р. Нижний Дубик (приток р. Непрядва). По данным палеогеографических реконструкций, на этой территории в XII–XIV вв. господствовали травяные сообщества. Леса занимали нижние уровни балочной сети – склоны и днища (Gonyanyi et al., 2007). Позднее на месте современного леса находились пашенные угодья, поскольку территория принадлежала графу Ольсуфьеву (Generalnyi…, 1790). Леса располагались только в балках. Постепенно исконные балочные леса вырубали, но на их месте формировался либо порослевой лес, либо землевладельцы, укрепляя склоны оврагов, проводили высадку саженцев (Bassein…, 1902, 1904). Впервые лесной массив Водяное поле (назван в честь одноименной небольшой деревни) появляется на картах 1928 г. (Tulskaya…, 1928). В 1990 г. массиву присвоен статус памятника природы регионального значения (Красная книга…, 2007).
Лесной массив Рыхотка расположен в нижнем течении р. Рыхотка (приток Дона) и занимает высокую позицию в прибровочной части левого коренного склона долины. На приводораздельный склон лес поднимается по небольшому разветвленному овражно-балочному комплексу. Первое картографическое изображение лесного массива, которое сходно с современными очертаниями, появляется на картах 1928 г. (Tulskaya…, 1928). До этого времени здесь была пашня крестьян из прилегающих деревень (Bassein…, 1902, 1904). В 1990 г. массиву вместе с прилегающим степным склоном присвоен статус памятника природы регионального значения (Krasnaya…, 2007). Таким образом, исследуемые лесные массивы имеют искусственное происхождение или испытывали существенное антропогенное воздействие в прошлом. В настоящее время эти леса используются в рекреационных целях, а также являются частью охотничьих хозяйств. Редкость лесных массивов в лесостепной части Тульской области делает их уникальными компонентами лесостепного ландшафта.
МАТЕРИАЛЫ И МЕТОДЫ
Флористико-геоботанические исследования на юго-востоке Тульской области были проведены в 2013 г. в пределах Кимовского и Куркинского районов. В анализ включены и несколько описаний, выполненных в лесном ур. Рыхотка (Липецкая область) у границы с Тульской областью. Геоботанические описания лесных сообществ выполнены на площадках в 400 м2. Оценка количественного участия видов в формировании сообществ произведена с использованием комбинированной шкалы обилия-покрытия Ж. Браун-Бланке (Braun-Blanquet, 1964).
Классификация растительности разработана на основе общих установок метода Ж. Браун-Бланке (Braun-Blanquet, 1964) и соответствует последним представлениям о синтаксономии широколиственных лесов Центральной России (Semenishchenkov, 2016).
В настоящей статье антропогенные и нарушенные леса рассматриваются в пределах единой синтаксономической схемы. Ранее такой подход неоднократно был использован при классификации лесной растительности в Центральной Европе (Hadač, Sofron, 1980; Zerbe, 2003; Ellenberg, 2009; Vegetace…, 2013) и Европейской России (Tikhonova et al., 2012; Semenishchenkov, 2016; Morozova et al., 2017; Shirokikh et al., 2018). Кроме того, большинство искусственных лесов изучаемого региона воссоздавались на месте ранее существовавших и уничтоженных лесных массивов, что обеспечивало преемственность лесорастительных условий и экологическую обусловленность формирования здесь именно мезофитных широколиственных лесов зонального типа. Антропогенное нарушение сообществ по характерным изменениям состава ценофлоры в данном случае отражается на уровне синтаксонов низших рангов. Следуя классическим установкам метода Ж. Браун-Бланке (Braun-Blanquet, 1964), доминирование видов, определяющих облик отдельных сообществ и имеющих в них высокое фитоценотическое значение, может стать основанием для установления синтаксона ранга фации. Фактически к фации можно отнести и производные лесные сообщества, в которых временно господствуют характерные для реализующихся в разных экологических условиях стадий сукцессии виды древесных растений. Например, Betula pendula, Populus tremula, Acer platanoides, Tilia cordata на месте коренных сообществ, формируемых Quercus robur и Fraxinus excelsior. Установленные подобным образом фации для лесной растительности ранее отмечались для разных регионов Центральной России (Bulo-khov, Solomeshch, 2003; Semenishchenkov, 2016). Наличие же выраженного блока дифференцирующих видов может быть использовано для установления варианта, отражающего особенности экологии местообитания, истории формирования сообщества и даже характер преобразования человеком: при этом среди дифференцирующих будут виды-индикаторы антропогенного воздействия. Данные представления лежат в основе разработанной в настоящей статье синтаксономии лесной растительности юго-востока Тульской области.
Оценка экологических режимов местообитаний сообществ проведена с использованием шкал Х. Элленберга (Ellenberg et al., 1992) в программе Indicator для MS Excel (Bulokhov, Semenishchenkov, 2006).
Названия сосудистых растений даны по П.Ф. Маевскому (Majevskii, 2014); мохообразных – по М.С. Игнатову и др. (Ignatov et al., 2006). Идентификация мохообразных проводилась к.б.н. В.В. Телегановой (Национальный парк “Угра”, г. Калуга).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Сообщества мезофитных широколиственных лесов юго-востока Тульской области относятся к широко распространенной в Центральной России асс. Fraxino excelsioris-Quercetum roboris Bulokhov et Solomeshch 2003. Она представляет Среднерусско-Приволжские мезофитные леса широколиственно-лесной и лесостепной зон за пределами плакорного распространения ели, соответствующие южной полосе широколиственных лесов. Основные ценообразователи таких лесов – Quercus robur, Tilia cordata, Fraxinus excelsior, Acer platanoides, виды Ulmus (Rastitelnost…, 1980). Эту ассоциацию мы рассматриваем в качестве зональной для данного природного региона в смысле, близком к понятию эпитаксона, объединяющего коренные и производные, формирующиеся на месте коренных, типы лесных сообществ (Sochava, 1968).
В настоящее время в литературе есть сведения о широком распространении лесов данного типа на территории западных областей России: Белгородской, Брянской, Воронежской, Калужской, Липецкой, Московской, Орловской, Тульской (Bulokhov, Solomeshch, 2003; Bulokhov, Semenishchenkov, 2015; Zaugolnova, Braslavskaya, 2003; Vostochnoyevropeiskie…, 2004; Starodubtseva, Khanina, 2009; Semenishchenkov, 2015, 2016; Semenishchenkov et al., 2015; Semenishchenkov et al.,., 2018). Наиболее хорошо они сохранились в мемориальных лесных массивах Калужских и Тульских засек; на большей же части своего обширного ареала эти леса представлены лишь небольшими фрагментами на фоне сельскохозяйственных земель.
Состав и структура. Древесный ярус состоит из двух подъярусов. Первый из наиболее типичных сообществ формируют Quercus robur и Fraxinus excelsior в разном соотношении. Во втором подъярусе – Tilia cordata, Betula pendula, Populus tremula, Acer platanoides, Ulmus glabra, преобладание которых характерно для разнообразных производных сообществ.
Сомкнутость древостоя колеблется от 45 до 80%.
Подлесок мозаичный. В подросте нередко обильны Acer platanoides, Tilia cordata, Fraxinus excelsior, Sorbus aucuparia. Среди кустарников наибольшую фитоценотическую значимость имеют Padus avium и Corylus avellana; высококонстантны Euonymus verrucosa, Lonicera xylosteum, в отдельных сообществах – Frangula alnus. Широко распространены сообщества с высоким обилием Padus avium, которая особенно успешно расселяется на нарушенных участках в лесах, чему способствуют кабаны. Редко отмечены типичные виды широколиственных лесостепных лесов – Acer tataricum и Rhamnus cathartica. В некоторые сообщества внедряются заносные кустарники Amelanchier spicata и Caragana arborescens, иногда встречается подрост Picea abies, проникающей в широколиственные леса из еловых культур. Потенциал распространения ели невысокий; она сильно страдает от сухости в условиях сильной ксерофитизации климата в последнее десятилетие, совпадающей со вспышками численности вредителя короеда типографа, массово уничтожающего ель в культурах (ур. Водяное поле).
Чрезвычайно редко встречается в лесах изучаемого типа характерный вид ассоциации Acer campestre, отмеченный всего один раз в ур. Бучальский лес. Причиной его низкой встречаемости в лесных массивах на юго-востоке Тульской области, по-видимому, следует считать чрезвычайно малую площадь сохранившихся лесных массивов и их высокую фрагментированность (Sheremetyeva et al., 2008), которые не позволили ему активно распространиться после уничтожения лесов. Редко встречается и другой характерный вид ассоциации – Euonymus europaea, встреченный в наиболее ее типичных сообществах в ур. Исаковский лес. Здесь бересклет европейский тяготеет к днищам балок и подножиям их склонов, по-видимому, с наибольшим увлажнением. Тем не менее, даже незначительное присутствие этих маркерных географических видов-индикаторов является отличительной особенностью лесостепных широколиственных лесов изучаемого типа и указывает на их связь с описываемой ассоциацией.
Сомкнутость кустарникового яруса колеблется в широких пределах – от 1 до 60%, что связано, в первую очередь, с характером нарушения сообществ. Сомкнутый кустарниковый полог нередко образуется в нижних частях балок, а также после выборочных рубок, способствующих осветлению после образования окон в древостое.
Травяной покров формируют преимущественно виды неморального сциофитного широкотравья. Иногда локально доминируют Aegopodium podagraria, Carex pilosa, Mercurialis perennis.
Полное преобладание в составе ценофлоры неморальных видов порядка Carpinetalia betuli P. Fukarek 1968 и класса Carpino-Fagetea Jakucs ex Passarge 1968 представляет сообщества этого типа как зональные широколиственные леса. В отличие сообществ данной ассоциации, распространенных южнее (Белгородская, Воронежская области) (Semenishchenkov, 2016; Semenishchenkov et al., 2018), в изучаемых тульских лесах слабо представлена комбинация термофильных видов союза Aceri tatarici-Quercion Zólyomi 1957 и термофильного порядка Quercetalia pubescenti-petraeae Klika 1933. Виды последнего в сообществах немногочисленны и рассеяны: Acer tataricum, Geranium sylvaticum, Primula veris, Pyrethrum corymbosum, Rhamnus cathartica, Rosa canina, Viola hirta.
Травостой чрезвычайно мозаичный, а его проективное покрытие сильно колеблется – от 1 до 90%. Мертвопокровные сообщества образуются чаще всего при высокой сомкнутости кустарникового яруса, а также после пожаров. При этом видовое богатство сообществ сильно снижается.
Сообщества с высоким проективным покрытием травяного яруса и высоким видовым богатством характерны для разреженных вторичных лесов. В целом видовое богатство колеблется от 7 (наиболее нарушенные сообщества с обедненным составом ценофлоры) до 37 (производные березняки) видов на 400 м2.
При низовых пожарах, прогоне скота и выпасе, а, в некоторых случаях, при активной роющей деятельности кабанов травостой лесов сильно повреждается, а в условиях пониженной конкуренции получают массовое развитие вегетативно подвижные неморальные виды или виды с высокой энергией семенного размножения. На территории обследованных лесных массивов отмечены на большой площади такие монодоминантные сообщества со сплошным доминированием Aegopodium podagraria, Galium odoratum, Lysimachia nummularia, Mercurialis perennis, Viola mirabilis.
Всего в составе ценофлоры лесов изучаемого типа выявлены 112 видов сосудистых растений и 12 – мохообразных. На рис. 2 показано сравнение спектров геоэлементов и полизональных групп ценофлор мезофитных широколиственных лесов юго-востока Тульской области (ЮВТ), удаленной к западу северной части бассейна Верхнего Днепра (БВД) (зона широколиственных лесов; Брянская, Орловская, север Курской областей) (Semenishchenkov, 2016) и бассейна Ворсклы (лесостепная зона; Белгородская область) (Semenishchenkov , 2012). Сравнительный анализ спектров демонстрирует похожее соотношение участия зональных геоэлементов и полизональных групп. Это подтверждает высокое флористическое сродство лесов данного типа на Русской равнине и возможность отнесения их к единой ассоциации.
В широколиственных лесах юго-востока Тульской области, представленных у северо-западной границы Среднерусской лесостепной подпровинции, так же, как и в бассейне р. Ворскла (Центральная южная лесостепь) существенно снижается участие суббореальных (подтаежных) видов, которые более широко встречаются в лесах изучаемого типа в северной части днепровского бассейна. Снижается и общая доля неморальных видов, что соответствует переходу из зоны широколиственных лесов в лесостепь. Однако небольшое участие свето- и теплолюбивых южносибирских и субпонтических (лесостепных) видов сохраняется примерно одинаковым – сообщества мезофитных широколиственных лесов не являются типичным фитоценотическим окружением для видов этих геоэлементов. Наиболее остепненной можно считать ценофлору лесов из бассейна Ворсклы, в которой представительны субпонтические (лесостепные) и даже отмечены единичные понтические (степные) виды. Минимальным во всех сравниваемых ценофлорах остается и участие бореальных (таежных) видов, которые в целом не характерны для мезофитных широколиственных лесов (в бассейне Ворсклы не выявлены). Перечисленные различия в составе ценофлор подтверждают дифференциацию мезофитных широколиственных лесов Центральной южной лесостепи, которые относят к субасс. F.-Q. crataegetosum curvisepalae Semenishchenkov 2012, от более северо-западных лесов (субасс. typicum) (Semenishchenkov, 2012, 2016). По спектру геоэлементов, а также из-за чрезвычайной редкости или отсутствия группы “южных” видов (Acer tataricum, Corydalis marschalliana, Crataegus curvisepala, Pyrus pyraster, Scilla sibirica, Viola odorata, V. suavis) леса юго-востока Тульской области в большей степени тяготеют к субасс. typicum.
Заметное долевое участие полизональных видов в составе ценофлор (в среднем в сумме – 28%) может свидетельствовать о благоприятных условиях для сохранения в лесных сообществах луговых и опушечных видов, большинство из которых имеют полизональное распространение. Это в большой мере связано с сильной антропогенной фрагментированностью сохранившихся лесов, нарушением их структуры при рубках, выпасе, пожарах. Ранее тенденция олуговения ценофлор широколиственных лесов при антропогенном нарушении отмечалась для днепровского бассейна (Semenishchenkov, 2016). Следует отметить, что участие полизональных видов возрастает в производных березовых и осиновых лесах на месте коренных сообществ.
Синтаксономическое разнообразие. Сообщества изучаемых лесов достаточно разнообразны по составу ценофлор, который отражает, в первую очередь, стадию сукцессии и характер нарушения.
Продромуc
Класс Carpino-Fagetea Jakucs ex Passarge 1968
Порядок Carpinetalia betuli P. Fukarek 1968
Cоюз Aceri campestris-Quercion roboris Bulokhov et Solomeshch in Bulokhov et Semenishchenkov 2015
Асс. Fraxino excelsioris-Quercetum roboris Bulokhov et Solomeshch 2003
Вар. typica
Tilia cordata fac.
Populus tremula fac.
Вар. Lysimachia nummularia
Betula pendula fac.
Вар. inops
Наиболее типичные сообщества ассоциации объединяет вариант typica (табл. 1, оп. 1–6). Он не имеет собственных дифференцирующих видов. В лесах этого варианта хорошо выражен смешанный древостой с преобладанием Quercus robur или Fraxinus excelsior и вторым подъярусом из Acer platanoides. В сообществах богато представлены виды неморального широкотравья, среди которых абсолютными доминантами обычно выступают Mercurialis perennis и Aegopodium podagraria.
Таблица 1.
Геоботанические описания асс. Fraxino excelsioris-Quercetum roboris. Table 1. Geobotanical relevés of association Fraxino excelsioris-Quercetum roboris
| Номера описаний/Relevés numbers | Ярус Layer |
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | К | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Синтаксоны/Syntaxa | 1 | 2 | 3 | 4 | 5 | 1 | 2 | 3 | 4 | 5 | ||||||||||||||||||||||||||
| Древесный ярус, сомкнутость крон, % Tree layer, crown density, % |
75 | 60 | 75 | 65 | 40 | 75 | 80 | 40 | 60 | 60 | 60 | 45 | 60 | 45 | 40 | 60 | 45 | 60 | 60 | 45 | 60 | 45 | 45 | 65 | 45 | 60 | 60 | 50 | 60 | 45 | ||||||
| Высота древостоя максимальная, м Forest stand height maximum, m |
25 | 24 | 24 | 25 | 24 | 25 | 18 | 24 | 24 | 16 | 25 | 24 | 16 | 18 | 24 | 18 | 18 | 14 | 14 | 18 | 20 | 18 | 18 | 18 | 20 | 22 | 16 | 18 | 25 | 26 | ||||||
| Диаметр деревьев преобладающей породы максимальный, см Diameter of trees of the prevailing species maximum, cm |
38 | 60 | 60 | 60 | 50 | 35 | 35 | 32 | 30 | 25 | 35 | 40 | 22 | 45 | 30 | 45 | – | – | – | – | 40 | 35 | 35 | – | 30 | 38 | – | – | 35 | 50 | ||||||
| Кустарниковый ярус, сомкнутость крон, % Shrub layer, crown density, % |
30 | 10 | 25 | 20 | 10 | 10 | 10 | 15 | 10 | 5 | 45 | 45 | 25 | 20 | 30 | 15 | 20 | 1 | 5 | 10 | 10 | 40 | 60 | 20 | 40 | 10 | 20 | 50 | 45 | 30 | ||||||
| Травяной ярус, проективное покрытие, % Herb layer, cover, % |
70 | 45 | 50 | 45 | 30 | 50 | 5 | 70 | 50 | 85 | 10 | 15 | 10 | 5 | 15 | 5 | 65 | 70 | 80 | 25 | 90 | 20 | 10 | 70 | 10 | 9 | 5 | 9 | 1 | 8 | ||||||
| Количество видов/Number of species | 25 | 29 | 25 | 28 | 24 | 25 | 13 | 28 | 24 | 23 | 28 | 29 | 30 | 17 | 26 | 17 | 23 | 21 | 39 | 21 | 30 | 16 | 18 | 28 | 18 | 20 | 18 | 25 | 10 | 22 | ||||||
| Индекс Шеннона–Уивера Shannon–Weaver’s index | 2.1 | 1.9 | 2.1 | 2.1 | 2.3 | 2 | 0.9 | 1.9 | 1.9 | 1.4 | 1.8 | 2.1 | 1.8 | 2 | 2.3 | 1.5 | 2 | 1.5 | 2 | 2.2 | 1.8 | 1.7 | 1.6 | 2.2 | 1.9 | 1.8 | 1.5 | 2.2 | 1.1 | 2.2 | ||||||
| Крутизна склона, о/Slope inclination, о | – | 15 | 8 | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | 12 | 10 | 8 | 8 | 5 | – | – | – | – | ||||||||
| Экспозиция склона/Slope exposition | – | ю | св | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | в | – | с | с | c | – | – | – | – | ||||||||
| Характеристики местообитаний по шкалам Элленберга / Characteristics of habitats by Ellenberg’s scales | ||||||||||||||||||||||||||||||||||||
| освещенность/illuminance | 4.4 | 4.3 | 4.2 | 4.3 | 4.3 | 3.8 | 5.5 | 4.5 | 4.7 | 4.6 | 5.2 | 4.9 | 5.0 | 5.0 | 5.3 | 5.6 | 6.0 | 6.1 | 5.9 | 6.1 | 5.8 | 5.2 | 5.3 | 6.1 | 5.9 | 5.2 | 4.7 | 5.5 | 5.8 | 4.7 | ||||||
| влажность почвы/soil moisture | 5.2 | 5.3 | 5.3 | 5.3 | 5.2 | 5.2 | 5.1 | 5.4 | 5.7 | 5.2 | 5.2 | 5.4 | 5.1 | 5.0 | 5.7 | 4.8 | 4.9 | 6.2 | 5.7 | 4.8 | 4.8 | 5.4 | 5.4 | 5.4 | 5.0 | 5.9 | 5.1 | 5.3 | 4.5 | 5.3 | ||||||
| реакция почвы/ soil reaction | 6.6 | 6.7 | 6.7 | 7.0 | 6.6 | 6.9 | 5.8 | 6.5 | 6.7 | 6.7 | 5.6 | 6.7 | 6.6 | 6.3 | 6.6 | 6.3 | 6.0 | 6.3 | 6.3 | 6.1 | 6.3 | 5.7 | 5.7 | 6.3 | 5.8 | 6.4 | 6.5 | 6.8 | 6.3 | 6.9 | ||||||
| богатство почвы минеральным азотом/mineral nitrogen soil richness | 5.8 | 6.1 | 6.2 | 6.4 | 6.1 | 6.2 | 5.0 | 4.8 | 4.9 | 5.5 | 5.1 | 6.3 | 5.8 | 5.9 | 5.1 | 4.7 | 5.5 | 5.6 | 5.4 | 5.5 | 5.5 | 5.6 | 5.7 | 5.7 | 5.1 | 5.3 | 6.1 | 5.5 | 5.0 | 5.8 | ||||||
| Характерные виды (х. в.) асс. Fraxino excelsioris-Quercetum roboris / Characteristic species (ch. sp.) of association Fraxino excelsioris-Quercetum roboris | ||||||||||||||||||||||||||||||||||||
| Quercus robur | A | 1 | 1 | r | 1 | 1 | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | 1 | 1 | 2 | 2 | 3 | 1 | 2 | 4 | 2 | V | . | . | III | V |
| Q. robur | B | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . | . | I | I | . |
| Q. robur | C | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | r | . | . | . | . | . | . | . | . | r | . | . | . | r | . | . | I | I | II |
| Fraxinus excelsior | A | 4 | . | 4 | 4 | 3 | 4 | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | V | . | I | . | . |
| F. excelsior | C | + | 1 | + | + | 1 | + | . | . | . | . | . | . | r | + | r | . | . | . | . | . | . | r | r | . | . | . | r | r | . | r | V | . | III | II | III |
| F. excelsior | B | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 3 | . | . | . | . | . | . | I |
| Euonymus europaea | C | 2 | . | 1 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | II | . | . | . | . |
| Acer campestre | C | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | I |
| Дифференцирующий вид (д. в.) фации Tilia cordata / Differential species (d. sp.) of facies Tilia cordata | ||||||||||||||||||||||||||||||||||||
| Tilia cordata | A | . | . | . | . | . | . | 5 | 5 | 4 | 4 | . | . | . | . | . | . | 3 | . | . | 3 | 1 | . | . | . | . | . | . | . | . | . | . | 4 | . | II | . |
| T. cordata | C | + | . | . | . | . | . | + | 1 | 1 | . | + | . | r | + | r | 2 | 1 | . | . | 1 | + | . | . | . | . | . | . | . | . | . | I | 3 | V | II | . |
| T. cordata | B | 1 | . | . | . | . | . | . | . | . | . | 1 | . | . | 1 | 1 | 1 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | I | . | IV | . | . |
| Д. в. фации Populus tremula / D. sp. of facies Populus tremula | ||||||||||||||||||||||||||||||||||||
| Populus tremula | A | . | . | . | . | . | . | . | . | . | . | 4 | 3 | 4 | 3 | 3 | 4 | . | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . | . | V | . | II |
| P. tremula | C | . | . | . | . | . | . | r | + | + | r | + | . | . | + | r | . | . | . | r | . | . | . | . | . | . | . | + | r | . | . | . | 4 | III | I | III |
| Д. в. вар. Lysimachia nummularia / D. sp. of var. Lysimachia nummularia | ||||||||||||||||||||||||||||||||||||
| Lysimachia nummularia | D | . | . | . | . | . | . | . | r | r | . | . | . | . | . | r | . | 4 | 4 | 5 | 2 | 5 | + | + | 1 | 1 | . | . | . | . | . | . | 2 | I | V | . |
| Galium aparine | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | 1 | + | + | r | r | r | + | r | . | . | . | . | . | . | . | . | V | . |
| Sambucus racemosa | C | . | r | . | + | r | . | . | . | . | r | . | . | . | . | . | . | r | . | . | + | r | r | r | + | . | . | . | . | . | . | III | 1 | . | IV | . |
| Hypericum perforatum | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | r | r | r | . | . | . | . | . | . | . | . | . | . | . | . | III | . |
| Carex contigua | D | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | r | + | r | r | + | + | . | . | . | . | . | . | r | . | . | . | . | II | III | II |
| Rubus idaeus | C | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | r | r | . | . | . | . | r | + | . | . | . | . | . | . | . | I | III | . |
| Д. в. фации Betula pendula / D. sp. of facies Betula pendula | ||||||||||||||||||||||||||||||||||||
| Betula pendula | A | . | . | . | . | . | . | . | . | . | . | . | 2 | . | . | 1 | . | 2 | 4 | 4 | 2 | 4 | 4 | 4 | 2 | 1 | . | . | . | . | . | . | . | II | V | . |
| Х. в. класса Carpino-Fagetea / Ch. sp. of class Carpino-Fagetea | ||||||||||||||||||||||||||||||||||||
| Acer platanoides | A | . | 4 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 3 | . | . | . | . | . | I | . | . | I | . |
| A. platanoides | B | 3 | . | 3 | 2 | 1 | 3 | . | . | . | . | . | . | . | 2 | . | . | . | . | . | . | + | 2 | 2 | 2 | . | 3 | . | 2 | . | 3 | V | . | I | III | III |
| A. platanoides | C | 1 | 1 | 2 | 1 | 1 | 1 | 1 | . | . | . | + | 1 | + | 1 | . | + | . | . | . | . | + | 3 | 4 | 2 | 3 | + | + | 2 | . | 2 | V | 1 | V | III | IV |
| Corylus avellana | C | 2 | . | 1 | r | . | . | . | + | + | + | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | 2 | 3 | 2 | r | III | 3 | I | . | IV |
| Ulmus glabra | C | . | . | 1 | + | . | 1 | . | . | . | . | . | + | + | r | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | r | III | . | III | . | II |
| U. glabra | A | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 4 | 1 | . | . | . | . | . | . | II |
| U. glabra | B | . | + | . | . | + | . | . | . | . | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | II | . | I | . | I |
| Euonymus verrucosa | C | . | + | + | + | + | + | . | r | r | r | r | r | r | . | r | r | r | . | . | . | + | r | r | r | . | 1 | + | + | r | + | V | 3 | V | III | V |
| Lonicera xylosteum | C | r | + | + | r | + | + | . | + | r | + | + | + | r | . | + | . | . | . | r | . | . | . | . | . | . | . | 1 | + | r | r | V | 3 | IV | I | IV |
| Mercurialis perennis | D | 4 | 3 | 3 | 3 | 2 | 3 | . | + | 2 | 5 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | V | 3 | . | . | . |
| Aegopodium podagraria | D | 2 | 1 | 2 | + | 2 | 2 | . | . | . | . | r | 1 | . | . | + | . | . | . | r | . | . | . | . | . | . | . | r | r | . | . | V | . | III | I | II |
| Galium odoratum | D | . | + | r | r | + | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | V | . | . | . | . |
| Lamium maculatum | D | . | r | r | + | r | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | V | . | . | . | . |
| Carex pilosa | D | r | r | + | . | r | . | . | 4 | 3 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | IV | 2 | . | . | . |
| Pulmonaria obscura | D | . | + | r | r | + | r | . | + | + | r | . | r | + | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | + | V | 3 | II | . | II |
| Polygonatum multiflorum | D | r | + | + | . | + | r | . | r | r | r | . | + | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | V | 3 | II | . | I |
| Lathyrus vernus | D | r | r | . | r | r | r | . | r | r | r | . | . | . | . | r | + | . | . | + | . | . | . | . | . | . | . | . | . | . | . | V | 3 | II | I | . |
| Dryopteris filix-mas | D | r | . | r | r | . | r | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | IV | . | I | . | . |
| Paris quadrifolia | D | r | r | . | . | r | r | . | r | r | . | . | r | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | r | IV | 2 | I | . | II |
| Campanula latifolia | D | . | r | r | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | III | . | . | . | . |
| Malus sylvestris | C | . | r | . | . | r | . | . | . | . | . | r | + | . | . | + | . | . | . | . | . | r | . | . | r | r | . | r | r | . | + | II | . | III | II | III |
| Actaea spicata | D | . | r | . | . | . | r | . | . | . | . | . | r | r | . | . | r | . | . | . | . | . | . | . | . | . | . | r | . | . | . | II | . | III | . | I |
| Geum urbanum | D | . | . | . | r | . | r | . | . | . | . | . | r | . | . | . | . | r | . | . | r | r | + | + | + | r | . | r | . | . | . | II | . | I | IV | I |
| Asarum europaeum | D | . | . | r | r | . | . | . | . | . | . | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | II | . | I | . | . |
| Anemonoides ranunculoides | D | . | r | . | . | r | . | . | . | . | r | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | + | r | . | . | r | II | 1 | I | . | III |
| Ranunculus cassubicus | D | + | . | . | r | . | . | . | . | . | r | . | . | . | . | r | . | . | . | . | . | . | . | . | . | . | . | r | r | . | . | II | 1 | I | . | II |
| Stachys sylvatica | D | . | . | 1 | r | . | . | . | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | II | 1 | . | . | . |
| Viola mirabilis | D | . | . | . | + | . | r | . | r | r | . | . | . | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | II | 2 | I | . | I |
| Lathraea squamaria | D | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | I | . | . | . | . |
| Viburnum opulus | С | r | . | . | . | . | . | r | . | . | . | r | r | r | . | r | r | r | . | r | . | . | . | . | . | . | r | . | r | r | . | I | 1 | V | II | III |
| Swida sanguinea | C | . | . | . | . | . | . | . | . | . | . | . | r | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | II | . | . |
| Ranunculus auricomus | D | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | + | . | + | + | + | . | . | . | . | r | . | . | . | . | . | . | I | III | I |
| Vicia sepium | D | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | r | r | . | r | r | . | . | . | . | . | . | . | . | . | . | . | I | III | . |
| Campanula trachelium | D | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | r | r | . | . | . | . | . | . | . | . | . | . | . | . | . | I | II | . |
| Athyrium filix-femina | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | r | . | . | . | . | . | . | . | . | . | . | II | . |
| Stellaria holostea | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | I | . |
| Scrophularia nodosa | D | . | . | . | . | . | . | . | . | . | r | . | . | . | . | r | . | . | . | . | . | r | . | . | . | . | . | . | . | . | r | . | 1 | I | I | I |
| Lapsana communis | D | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | 1 | . | I | . |
| Neottia nidus-avis | D | . | . | . | . | . | . | . | r | . | r | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . | r | . | . | . | . | 2 | . | I | I |
| Х. в. порядка Quercetalia pubescenti-petraeae и класса Trifolio-Geranietea / Ch. sp. of order Quercetalia pubescenti-petraeae and class Trifolio-Geranietea | ||||||||||||||||||||||||||||||||||||
| Acer tataricum | C | . | . | . | . | . | . | r | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | I | . | . |
| Rosa canina | C | . | . | . | . | . | . | . | . | . | . | r | . | . | r | . | . | r | . | . | + | . | . | . | . | . | . | . | . | . | . | . | . | II | II | . |
| Geranium sylvaticum | D | . | . | . | . | . | . | . | . | . | . | r | r | . | . | . | . | . | + | r | . | . | . | . | . | . | . | . | . | . | . | . | . | II | II | . |
| Pyrethrum corymbosum | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | I | . | I |
| Rhamnus cathartica | C | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . | r | . | . | . | r | . | . | . | . | I | I | I |
| Primula veris | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | r | . | . | . | . | r | . | . | . | II | I |
| Viola hirta | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | r | . | . | . | . | . | . | . | . | . | . | . | . | II | . |
| Прочие виды / Other species | ||||||||||||||||||||||||||||||||||||
| Padus avium | C | 1 | + | r | 2 | + | 1 | . | 1 | + | + | . | 3 | 2 | 2 | 2 | . | 2 | . | 1 | 1 | 1 | r | + | . | + | 1 | r | 1 | 3 | 2 | V | 3 | IV | IV | V |
| Convallaria majalis | D | r | + | . | r | + | + | . | . | . | . | r | + | r | 1 | r | r | r | . | . | r | . | . | . | . | + | . | . | . | . | . | V | . | V | II | . |
| Sorbus aucuparia | C | + | + | r | . | + | . | + | + | + | r | 1 | 1 | + | r | + | r | + | . | . | + | + | + | + | + | 1 | . | + | + | + | r | IV | 4 | V | IV | IV |
| Rubus saxatilis | D | . | r | . | . | . | . | . | r | r | . | r | r | . | . | . | . | 1 | + | r | 1 | 1 | . | . | r | . | + | . | . | . | . | I | 2 | II | IV | I |
| Urtica dioica | D | . | . | . | r | . | . | . | . | . | . | . | r | r | . | . | . | . | + | . | . | + | . | . | r | . | . | . | . | . | . | I | . | II | II | . |
| Equisetum pratense | D | . | . | . | . | . | . | . | 1 | + | + | . | . | . | . | 1 | . | . | 1 | . | . | . | 2 | 1 | . | . | . | . | . | . | . | . | 3 | I | II | . |
| Dryopteris carthusiana | D | . | . | . | . | . | . | . | r | + | . | + | . | . | . | r | . | . | . | . | . | . | + | + | . | . | r | . | . | . | . | . | 2 | II | II | I |
| Atrichum undulatum | D | . | . | . | . | . | . | . | r | r | . | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | 2 | I | . | I |
| Geum rivale | D | . | . | . | . | . | . | . | r | r | . | . | . | . | . | + | . | . | . | 2 | . | . | . | . | . | . | . | . | . | . | . | . | 2 | I | I | . |
| Rubus caesius | C | . | . | . | . | . | . | . | 1 | + | . | . | . | . | . | 2 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 2 | I | . | . |
| Fragaria vesca | D | . | . | . | . | . | . | + | . | . | . | r | r | . | r | . | r | + | r | + | 1 | . | r | r | + | . | . | . | . | . | . | . | 1 | IV | IV | . |
| Frangula alnus | C | . | . | . | . | . | . | r | . | . | . | 3 | . | . | . | . | r | . | . | r | . | . | . | . | r | . | + | . | . | . | . | . | 1 | II | II | I |
| Picea abies | C | . | . | . | . | . | . | 1 | . | . | . | . | . | . | + | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | 1 | I | . | I |
| Amelanchier spicata | C | . | . | . | . | . | . | r | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | I | . | . |
| Maianthemum bifolium | D | . | . | . | . | . | . | . | r | . | . | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | I | . | . |
| Ranunculus repens | D | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . | r | r | . | . | . | . | . | . | . | . | . | . | . | . | 1 | . | II | . |
| Filipendula ulmaria | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | . | r | . | . | . | . | . | . | . | . | . | . | . | . | . | I | I | . |
| Veronica chamaedrys | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | r | r | . | . | r | . | . | . | . | . | II | I |
| Valeriana officinalis | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | I |
| Platanthera chlorantha | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | I |
| Lysimachia vulgaris | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | + | . | . | . | . | . | . | . | . | . | . | . | . | . | . | II | . |
| Leucanthemum vulgare | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | r | . | . | . | r | . | . | . | . | . | . | . | . | II | . |
| Caragana arborescens | C | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | r | + | . | . | . | . | . | . | . | . | II | . |
| Gagea minima | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | . | . | . | . | . | . | I | . |
| Bistorta major | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | r | + | . | . | . | . | . | . | . | . | II | . |
| Veratrum lobelianum | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . | II | . |
| Carex pallescens | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . | II | . |
| Alchemilla sp. | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . | II | . |
| Taraxacum officinale | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | . | . | r | r | . | . | . | . | . | . | . | . | II | . |
| Arctium sp. | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | r | . | . | . | . | . | . | . | . | . | . | . | . | . | II | . |
| Anthriscus sylvestris | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | r | . | . | . | . | . | . | . | . | . | . | . | . | . | II | . |
| Chelidonium majus | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | . | r | . | . | . | . | . | . | . | . | . | II | . |
| Cirsium polonicum | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | r | . | . | . | . | . | . | . | . | . | II | . |
| Glechoma hederacea | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | r | . | . | . | . | . | . | . | . | . | II | . |
| Viola × neglecta | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | r | . | . | . | . | . | . | . | . | . | . | . | . | II | . |
| Fissidens taxifolius | E | r | . | r | r | . | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | . | IV | . | . | . | I |
| Brachytecium salebrosum | E | . | r | . | . | . | r | . | . | . | . | r | r | r | . | r | . | . | r | . | . | . | . | . | . | . | r | r | r | r | . | II | . | IV | I | V |
| Fissidens bryoides | E | r | r | . | . | . | . | . | r | . | r | . | . | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | I | 2 | I | . | . |
| Oxyrrhynchium hians | E | . | . | r | . | r | . | . | r | . | r | . | . | r | . | . | . | . | r | r | . | . | . | . | . | . | . | . | . | . | . | I | 2 | I | I | . |
| Plagiomnium cuspidatum | E | . | r | . | r | . | . | . | r | r | . | . | . | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | I | 2 | I | . | . |
| Eurhynchiastrum angustirete | E | . | . | . | . | . | . | . | . | . | . | r | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | . | II | . | I |
| Rhytidiadelphus triquetrus | E | . | . | . | . | . | . | . | . | . | . | r | r | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | II | . | . |
| Pleurozium schreberi | E | . | . | . | . | . | . | . | . | . | . | . | . | . | . | r | . | . | r | r | . | . | . | . | . | . | . | . | . | . | . | . | . | I | I | . |
Примечание. Отмечены в одном описании: Allium oleraceum D (20,r), Bryum sp. E (30,r), Campanula patula D (19,r), Carex cespitosa D (18,r), C. sylvatica D (10,r), Crataegus curvisepala C (30,+), Epilobium montanum D (19,r), Epipactis helleborine D (7,r), Galium boreale D (24,+), Grossularia reclinata C (14,r), Hieracium sylvularum D (24,4), Leontodon hispidus D (19,r), Lonicera tatarica C (24,r), Lychnis flos-cuculi D (18,r), Matteucia struthiopteris D (6,r), Melica nutans D (19,r), Picea abies C (13,r), Polygonatum odoratum D (7,r), Potentilla alba D (11,r), P. goldbachii D (18,r), Pylaisia polyantha E (26,r), Ribes nigrum C (9,r), Sciuro-hypnum reflexum E (13,+), Solidago virgaurea D (21,r), Sonchus arvensis D (24,r), Vicia sylvatica D (10,r), Weissia squarrosa E (2,r).
Обозначения ярусов и подъярусов: A – первый подъярус древостоя, B – второй подъярус, С – кустарниковый ярус, подлесок, D – травяной ярус, E – моховой ярус. Класс постоянства (К) дан по пятибалльной шкале: 1 – вид присутствует, менее чем в 20% описаний, 2 – 21–40%, 3 – 41–60%, 4 – 61–80%, 5 – в более 80% описаний.
Обозначения синтаксонов: 1 – асс. Fraxino excelsioris-Quercetum roboris typica var., 2 – асс. F. e.-Q. r. typica var. Tilia cordata fac., 3 – асс. F. e.-Q. r. typica var. Populus tremula fac., 4 – асс. F. e.-Q. r. Lysimachia nummularia var. Betula pendula var., 5 – асс. F. e.-Q. r. inops var.
Локализация описаний. Тульская обл., Кимовский р-н: оп. 1–6 – ур. Исаковский лес, 07.06.2013; оп. 7, 14, 16 – ур. Водяное поле, 05.06.2013; оп. 8–10, 15, 18, 19, 30 – ур. Бучальский лес, 08.06.2013; оп. 11, 12, 29 – ур. Стрельцы, 08.06.2013; оп. 13, 27, 28 – ур. Грязновская балка, 06.06.2013; оп. 17, 20, 21, 25 – ур. Рыхотка, 06.06.2013; оп. 26 – ур. Лес Дубрава, 08.06.2013. Липецкая обл., Данковский р-н: оп. 22–24 – ур. Рыхотка, 06.06.2013.
Авторы описаний: 1–6, 8–12, 15, 18, 19, 26, 29, 30 – Е. М. Волкова, Ю. А. Семенищенков; 7, 13, 14, 16, 17, 20–25, 27, 28 – Ю. А. Семенищенков.
Note. Following species are noted in one relevé each: Allium oleraceum D (20,r), Bryum sp. E (30,r), Campanula patula D (19,r), Carex cespitosa D (18,r), C. sylvatica D (10,r), Crataegus curvisepala C (30,+), Epilobium montanum D (19,r), Epipactis helleborine D (7,r), Galium boreale D (24,+), Grossularia reclinata C (14,r), Hieracium sylvularum D (24,4), Leontodon hispidus D (19,r), Lonicera tatarica C (24,r), Lychnis flos-cuculi D (18,r), Matteucia struthiopteris D (6,r), Melica nutans D (19,r), Picea abies C (13,r), Polygonatum odoratum D (7,r), Potentilla alba D (11,r), P. goldbachii D (18,r), Pylaisia polyantha E (26,r), Ribes nigrum C (9,r), Sciuro-hypnum reflexum E (13,+), Solidago virgaurea D (21,r), Sonchus arvensis D (24,r), Vicia sylvatica D (10,r), Weissia squarrosa E (2,r).
Designations of layers and sublayers: A – first sublayer of the forest stand, B – second sublayer, C – shrub layer, underbrush, D – herb layer, E – moss layer. The class of constancy (K) is given on a five-point scale: 1 – species is present in less than 20% relevés, 2 – 21–40%, 3 – 41–60%, 4 – 61–80%, 5 – over 80% relevés.
Designations of syntaxa: 1 – association Fraxino excelsioris-Quercetum roboris typica var., 2 – association F. e.-Q. r. typica var. Tilia cordata fac., 3 – association F. e.-Q. r. typica var. Populus tremula fac., 4 – association F. e.-Q. r. Lysimachia nummularia var. Betula pendula var., 5 – association F. e.-Q. r. inops var.
Localities of relevés. Tula Region, Kimovsky District: relevés 1–6 – natural landmark Isakovsky Forest, 07.06.2013; relevés 7, 14, 16 – natural landmark Vodyanoye Polye, 05.06.2013; оп. 8–10, 15, 18, 19, 30 – natural landmark Buchal’sky Forest, 08.06.2013; relevés 11, 12, 29 – natural landmark Strel’tsy, 08.06.2013; relevés 13, 27, 28 – natural landmark Gryaznovskaya ravine, 06.06.2013; relevés 17, 20, 21, 25 – natural landmark Rykhotka, 06.06.2013; relevé 26 – natural landmark Forest Dubrava, 08.06.2013. Lipetsk Region, Dankovsky District: relevés 22–24 – natural landmark Rykhotka, 06.06.2013.
Authors of relevés: 1–6, 8–12, 15, 18, 19, 26, 29, 30 – Е. М. Volkova, Yu. A. Semenishchenkov; 7, 13, 14, 16, 17, 20–25, 27, 28 – Yu. A. Semenishchenkov.
Высокие фитоценотические позиции и активное возобновление ясеня в лесах данного варианта коррелируют с наибольшими значениями богатства минеральным азотом (рис. 3 , г) и pH почвы (рис. 3 , в). Нередко ясеневые леса формируются в местообитаниях с почвами, подстилаемыми карбонатными породами. Однако, как показали исследования в XX в. (Vysotskii, 1929; Sukachev, 1938; Pogrebniak, 1949; Yurkevich, Adarikho, 1973), приуроченность ясеня к карбонатным почвам обусловлена в значительной мере не его кальцефильностью, а бóльшим плодородием таких почв. По-видимому, присутствие ясеня и в наиболее гигрофитных экотопах, по днищам балок и речных долин с обильным увлажнением почв, можно объяснить его предпочтением сопутствующего высокого богатства почв (Sukachev, 1938).
Видовое богатство сообществ варианта – 24–29 видов на 400 м2.
Сообщества варианта занимают выровненные пространства водоразделов с темно-серыми суглинистыми почвами и склоны балок небольшой крутизны разной экспозиции со смытыми суглинками. Они сохранились в ур. Исаковский лес и заслуживают особой охраны как эталонные леса изучаемого типа.
На месте коренных лесов ассоциации формируются производные сообщества, которые отнесены нами к временным фациям с преобладанием отдельных пород, характеризующим этапы восстановления коренного широколиственного леса.
Варианту соответствуют ассоциации, установленные на основе эколого-фитоценотической классификации: ясене-дубняк пролесниковый, ясене-дубняк снытевый.
Фация Tilia cordata (табл. 1, оп. 7–10) объединяет вторичные липовые леса, которые сформировались после вырубки коренных дубово-ясеневых сообществ на наиболее бедных (рис. 3 , г) суглинистых почвах. Липа надолго захватывает местообитания после рубок, активно восстанавливается порослевым способом, о чем свидетельствует наличие многоствольных порослевых деревьев. В некоторых местах сохранились остатки пней высоковозрастных деревьев дуба после его вырубки. Возобновление коренных пород в условиях высокого затенения сильно затруднено. Такие сообщества обычно характеризуются обедненным флористическим составом, снижением фитоценотических позиций неморальных мезофитов и высоким обилием Carex pilosa. Этот длиннокорневищный вид активно разрастается после нарушений в тенистых лесах и на отдельных участках в липняках формирует сплошное покрытие.
Видовое богатство сообществ варианта – 13–28 видов на 400 м2.
Сообщества фации распространены на выровненных водоразделах с темно-серыми суглинистыми почвами.
Фации соответствуют ассоциации, установленные на основе эколого-фитоценотической классификации: липняк волосистоосоковый, липняк пролесниковый.
Фация Populus tremula (табл. 1, оп. 11–16) объединяет производные осиновые леса, как правило, с развитым черемуховым подлеском. Во втором подъярусе древесного яруса Acer platanoides уступает место Betula pendula. Обычно осинники возникают после разрушения коренных широколиственных лесов в условиях бóльших увлажнения и богатства почвы. Эти условия характеризуют высокие константность кустарника Viburnum opulus и обилие Frangula alnus в некоторых сообществах. Подрост коренных пород – дуба и ясеня – практически отсутствует, в то время как более обильны в подросте клен и липа. В будущем при распаде осинового древостоя возможно формирование длительнопроизводного липо-кленового леса.
В травостое сообществ фации снижается разнообразие неморального широкотравья, четкие доминанты не выражены. Повышается константность светолюбивых лесных видов: Convallaria majalis, Fragaria vesca.
Видовое богатство сообществ варианта – 17–29 видов на 400 м2.
Распространены сообщества фации в тех же местообитаниях, что и для предыдущей фации.
Фации соответствуют ассоциации, установленные на основе эколого-фитоценотической классификации: осинник разнотравный, осинник черемуховый, липо-осинник разнотравный.
В условиях сильного нарушения – пожаров, выпаса, прогона скота – на месте коренных широколиственных лесов сформировались сообщества антропогенного вар. Lysimachia vulgaris (табл. 1, оп. 17–25). Его дифференцирующие виды: Carex contigua, Galium aparine, Hypericum perforatum, Lysimachia nummularia, Rubus idaeus, Sambucus racemosa. Большинство перечисленных видов достаточно светолюбивы и индицируют осветление сообществ в результате нарушений. С другой стороны, Galium aparine, Rubus idaeus и Sambucus racemosa – индикаторы богатых почв (Ellenberg et al., 1992). Для многих сообществ варианта характерно полное или локальное доминирование почвопокровного Lysimachia nummularia, который быстро захватывает нарушенный субстрат в условиях пониженной конкуренции, размножаясь вегетативным и семенным путем. Ценофлора варианта характеризуется выпадением неморальных сциофитов и повышением константности ряда светолюбивых лесных (Fragaria vesca, Ranunculus auricomus, Veronica chamaedrys, Vicia sepium), лесо-опушечных (Primula veris, Viola hirta) и лугово-опушечных (Leucanthemum vulgare) видов. Характерны и рудерально-лесные нитрофильные (Ellenberg et al., 1992) виды: Chelidonium majus, Glechoma hederacea, Taraxacum officinale, Urtica dioica.
Достаточно интересными с флористической точки зрения являются сообщества с высоким обилием заносного вида Hieracium sylvularum11. По-видимому, этот вид в Средней полосе России является реликтом интродукции начала XX в. в усадебные парки (Kaluzhskaya…, 2010). В настоящее время ястребинка рощевая массово распространяется в лесах разного состава и локально доминирует, формируя желтый аспект во время цветения.
Сообщества этого варианта существуют в виде березовой фации Betula pendula. В них береза повислая формирует первый подъярус древесного яруса. Высота деревьев 14–20 м, что свидетельствует о небольшом возрасте этих вторичных насаждений. Иногда в древостое сохраняется дуб; во втором подъярусе в некоторых сообществах обилен Acer platanoides. В сообществах единично присутствует подрост дуба или ясеня, поэтому в будущем на месте березняка возможно формирование кленового леса с небольшим участием всех перечисленных пород, с сопутствующим нарастанием затенения и выпадением гелиофитных видов из травостоя.
Видовое богатство сообществ варианта – 18–39 видов на 400 м2.
Сообщества варианта распространены на выровненных водоразделах на смытом черноземе, а также на склонах балок небольшой крутизны со смытыми суглинками.
Варианту и фации соответствуют ассоциации, установленные на основе эколого-фитоценотической классификации: березняк разнотравный, клено-березняк разнотравный, дубо-березняк разнотравный.
Перечисленные синтаксоны формируют экологический ряд на градиенте нарастания освещенности под пологом леса, оцененной методом экологических шкал: вар. typica → Tilia cordata fac. → Populus tremula fac. → вар. Lysimachia nummularia Betula pendula fac. (рис. 3 , а). В этом же ряду закономерно повышается кислотность почвы (рис. 3 , в). Местообитания всех синтаксонов характеризуются близкими значениями почвенной влажности (рис. 3 , б).
Сообщества дубрав с наиболее обедненным составом ценофлоры, сформировавшиеся после выборочных рубок или лесных пожаров, объединяет вар. inops (табл. 1, оп. 26–30). Фитоценозы отличаются высоким затенением, которое создается сомкнутым подлеском, где с высоким обилием представлены Acer platanoides, Corylus avellana и Padus avium.
Проективное покрытие травяного яруса очень мало: 1–9%; сообщества мертвопокровные. Видовое богатство в целом снижается – 10–25 видов на 400 м2, хотя в отдельных сообществах сохраняется высоким за счет многочисленных единично присутствующих растений разных видов.
Варианту соответствуют ассоциации, установленные на основе эколого-фитоценотической классификации: дубняк черемуховый, дубняк лещиновый, дубняк разнотравный, ясене-дубняк лещиновый.
ЗАКЛЮЧЕНИЕ
Сохранившиеся мезофитные широколиственные леса юго-востока Тульской области, несмотря на антропогенное происхождение и существенную антропогенную трансформацию, представляют собой характерную лесную растительность переходной полосы из зоны широколиственных лесов в лесостепь. С позиций подхода Ж. Браун-Бланке, они относятся к широко распространенной в Центральной России асс. Fraxino excelsioris-Quercetum roboris Bulokhov et Solomeshch 2003, объединяющей Среднерусско-Приволжские мезофитные леса широколиственно-лесной и лесостепной зон за пределами плакорного распространения ели. Сообщества лесов изучаемого региона достаточно разнообразны по составу, который отражает стадию сукцессии и характер нарушения.
Анализ спектров геоэлементов и полизональных групп продемонстрировал ботанико-географическую дифференциацию мезофитных широколиственных лесов юго-востока Тульской области, северной части днепровского бассейна и бассейна Ворсклы.
Несмотря на значительную трансформацию человеком, леса изучаемого типа на юго-востоке Тульской области представляют собой сохранившиеся убежища видов неморальной флоры, а также элементы эталонных ландшафтов лесостепной зоны у ее северо-западной границы.
Список литературы
[Aljokhin] Алехин В.В. 1947. Растительность и геоботанические районы Московской и сопредельных областей. М. 71 с.
[Bassein…] Бассейн р. Дон до слияния с Непрядвой. Исследования лесоводственного отдела 1895 года. 1902. – Тр. экспедиции для исследования источников главнейших рек Европейской России. М. 63 с.
[Bassein…] Бассейн Дона. Водосборы Непрядвы, Сухой и Мокрой Табол и правой стороны р. Дон до впадения р. Красивой Мечи. Исследования лесоводственного отдела. 1898 года. 1904. – Тр. экспедиции для исследования источников главнейших рек Европейской России. М. 183 с.
[Bolotov] Болотов А.Т. 1766. Описание свойства и доброты земель Каширского уезда и прочих от сего уезда касающихся обстоятельств с ответами на предложенные вопросы. – Тр. Вольного экономического о-ва. Ч. 2. С. 129–219.
Braun-Blanquet J. 1964. Pflanzensoziologie. Grundzuge der Vegetationskunde. 3 Aufl. Wien; New-York. 865 S.
[Bulokhov, Semenishchenkov] Булохов А.Д., Семенищенков Ю.А. 2006. Компьютерная программа Indicator и методические указания по ее использованию для экологической оценки местообитаний и анализа флористического разнообразия растительных сообществ. Брянск. 30 с.
[Bulokhov, Semenishchenkov] Булохов А.Д., Семенищенков Ю.А. 2015. Типификация и коррекция синтаксонов лесной растительности Южного Нечерноземья России и сопредельных регионов. – Бюл. Брянского отделения РБО. 1 (5): 26–32.
[Bulokhov, Solomeshch] Булохов А.Д., Соломещ А.И. 2003. Эколого-флористическая классификация лесов Южного Нечерноземья России. Брянск. 359 с.
[Burova] Бурова О.В. 2018. Этапы развития и антропогенная трансформация лесостепных ландшафтов в бассейне Верхнего Дона. – Сб. науч. ст. “Проблемы изучения и восстановления ландшафтов лесостепной зоны: историко-культурные и природные территории”. Вып. 4. Тула. С. 7–11.
Ellenberg H. 2009. Vegetation Ecology of Central Europe. Cambridge. 756 p.
Ellenberg H., Weber H.E., Düll R., Wirth W., Paulißen D. 1992. Zeigerwerte von Pflanzen in Mitteleuropa. 2 Aufl. Göttingen. 258 S.
[Generalnyi…] Генеральный план Епифанского уезда. Ч. 1, ч. 2. 1790. Масштаб: 2 версты в английском дюйме / РГАДА, № описи 593, ч. 1, ч. 2 Тульской губернии, Епифанский уезд. М.
[Glasko et al.] Гласко М.П., Гольева С.А., Сычева С.А., Бурова О.В. 2005. Ландшафты Донского побоища: возвращение утраченного. – Тр. ГИМ. 150: 227–256.
[Golitsyn] Голицын В.М. 1905. Очерк флоры Епифанского уезда Тульской губернии. – Тр. Ботанического сада Юрьевского университета. Т. 5 (1904 г.). 4: 230–243.
[Golitsyn] Голицын В.М. 1906. Очерк флоры Епифанского уезда Тульской губернии. (Окончание). – Тр. Ботанического сада Юрьевского университета. Т. 6 (1905 г.). 1: 20–31.
[Golitsyn] Голицын В.М. 1907. Первое дополнение к очерку флоры Епифанского уезда Тульской губернии. – Тр. Ботанического сада Юрьевского университета. Т. 7 (1906 г.). 4: 231–233.
[Gonyanyi et al.] Гоняный М.И., Александровский А.А., Гласко М.П. 2007. Северная лесостепь бассейна Верхнего Дона времени Куликовской битвы. М. 208 с.
Hadač E., Sofron J. 1980. Notes on syntaxonomy of cultural forest communities. – Folia Geobot. Phytotax. 15: 245–258.
Ignatov M.S. et al. 2006. Check-list of mosses of East Europe and North Asia. – Arctoa. 15: 10–131.
[Isachenko, Lavrenko] Исаченко Т.И., Лавренко Е.М. 1980. Ботанико-географическое районирование. – В кн.: Растительность европейской части СССР. Л. С. 10–20.
[Iurkevich, Aderikho] Юркевич И.Д., Адерихо В.С. 1973. Типы и ассоциации ясеневых лесов (по исследованиям в БССР). Минск. 256 с.
[Kaluzhskaya…] Калужская флора: аннотированный список сосудистых растений Калужской области. 2010. М. С. 439.
[Krasnaya…] Красная книга: особо охраняемые природные территории Тульской области. 2007. Тула. 316 с.
[Kulikovo…] Куликово поле. Документы по землевладению XVII в. 1999. Вып. 1. Тула.
[Kurnaev] Курнаев С.Ф. 1980. Тульские засеки, их природа и научно-хозяйственное значение. – Живая природа. 3: 86–99.
[Levitskij] Левицкий С.С. 1927. Геоботанические исследования в юго-восточной части Тульской губернии. – Мат. к изучению флоры и фауны Центрально-Промышленной области. М. С. 7–10.
[Majevskii] Маевский П.Ф. 2014. Флора средней полосы европейской части России. 11-е изд. М. 635 с.
[Morozova et al.] Морозова О.В., Семенищенков Ю.А., Тихонова Е.В., Беляева Н.Г., Кожевникова М.В., Черненькова Т.В. 2017. Неморальнотравные ельники Европейской России. – Растительность России. 31: 33–58.
[Pogrebnyak] Погребняк П.С. 1949. Результати вегатацiйних дослiдiв з мiнерального живлення деревних порiд. – Працi Iн-ту лiсiвнiцтва АН УРСР. I: 10–67.
[Rastitelnost…] Растительность европейской части СССР. 1980. Л. 429 с.
[Semenishchenkov, Polujanov] Семенищенков Ю.А., Полуянов А.В. 2014. Остепненные широколиственные леса союза Aceri tatarici-Quercion Zólyomi 1957 на Cреднерусской возвышенности. – Растительность России. 24:101–123.
[Semenishchenkov et al.] Семенищенков Ю.А., Телеганова В.В., Шапурко А.В., Кобозев Д.А. 2015. О новой субассоциации мезофитных широколиственных лесов на Юго-Востоке Калужской области. – Современные проблемы науки и образования. 3: 67–73.
[Semenishchenkov et al.] Семенищенков Ю.А., Волкова Е.М., Бурова О.В. 2013. О новой ассоциации союза Aceri tatarici-Quercion Zólyomi 1957 на территории заповедника “Куликово поле” (Тульская область). – Изв. СамНЦ РАН. 15 (3): 405–414.
[Semenishchenkov] Семенищенков Ю.А. 2015. Флористическая классификация как отражение ботанико-географического разнообразия мезофитных широколиственных лесов запада Среднерусской возвышенности. – Мат. Московского городского отделения Русского географического общества. Биогеография. Вып. 19. М. С. 45–60.
[Semenishchenkov] Семенищенков Ю.А. 2016. Эколого-флористическая классификация как основа ботанико-географического районирования и охраны лесной растительности бассейна Верхнего Днепра (в пределах Российской Федерации): Дис. … докт. биол. наук. Уфа. 558 с.
[Semenishchenkov] Семенищенков Ю.А. 2012. Cообщества союза Aceri campestris-Quercion roboris Bulokhov et Solomeshch 2003 в бассейне реки Ворсклы (Белгородская область). – Известия Тульского гос. ун-та. Сер. Естественные науки. 3: 221–230.
[Sheremetyeva et al.] Шереметьева И.С., Хорун Л.В., Щербаков А.В. 2008. Конспект флоры сосудистых растений Тульской области. Тула. 274 с.
[Shirokikh et al.] Динамика растительности на вырубках Южно-Уральского региона: основные итоги исследований Уфимской геоботанической школы. 2018. – Фиторазнообразие Восточной Европы. XII (3): 17–30.
[Sochava] Сочава В.Б. 1968. Растительные сообщества и динамика природных систем. – Докл. ин-та географии Сибири и Дальнего Востока. Вып. 20. С. 12–22.
[Starodubtseva, Khanina] Стародубцева Е.А., Ханина Л.Г. 2009. Классификация растительности Воронежского заповедника. – Растительность России. 14: 78–107.
[Sukachyev] Сукачев В.Н. 1938. Дендрология с основами лесной геоботаники. М.–Л. 115 с.
[Tikhonova et al.] Тихонова Е.В., Пестерова О.А., Семенищенков Ю.А. 2012. Синтаксономический анализ искусственных лесов Юго-Западного Подмосковья. – Изв. СамНЦ РАН. 1–4: 1135–1138.
[Tulskaya…] Тульская и Рязанская губернии. Богородицкий, Раненбургский и Скопинский уезды. N-37-90. Куркино. 1 : 100 000. 1928. Сост. Карт. отд. ГГК. М.
Vegetace České republiky. 4. Lesní a křovinná vegetace. 2013. Vyd. 1. Praha. 551 s.
Volkova E., Burova O. 2014. Natural conditions of the Kulikovo Field (Tula region). –Book of Abstracts and Excursion Gudes of the 11th European Dry Grassland Meeting “Steppes and Semi-Natural Dry Grasslands: Ecology, Transformation and Restoration”, 5–15th June 2014. Tula. P. 80–88.
[Vostochnoyevropeiskie…] Восточноевропейские леса: история в голоцене и современность. Кн. 2. 2004. М. 575 с.
[Vysotskii] Высоцкий Г.Н. 1929. Позиция ясеня в наших лесах (и морской полыни в степях). – Очерки по фитосоциологии и фитогеографии. М. С. 17–19.
[Zaugolnova, Braslavskaya] Заугольнова Л.Б., Браславская Т.Ю. 2003. Анализ ассоциаций мезофитных широколиственных лесов в центре Европейской России. – Растительность России. 4: 3–28.
Zerbe S. 2003. The differentiation of anthropogenous forest communities: a syn-systematical approach. – Mitt. Naturwiss. Ver. Steiermark. 133:. P. 109–117.
[Zony…] Зоны и типы поясности растительности России и сопредельных территорий. Карта масштаба 1 : 8000000. 1999. М.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал