

Ботанический журнал, 2019, T. 104, № 7, стр. 1110-1121
МОРФОЛОГИЯ ПЫЛЬЦЕВЫХ ЗЕРЕН НЕКОТОРЫХ ВИДОВ РОДА POLYGALA (POLYGALACEAE) ФЛОРЫ КАВКАЗА
И. В. Телицына 1, 2, *, В. В. Григорьева 1, **, А. Е. Поджидаев 1, ***, О. А. Гаврилова 1, ****, В. В. Шванова 1
1 Ботанический институт им. В.Л. Комарова РАН
197376 Санкт-Петербург, ул. Проф. Попова, 2, Россия
2 Пятигорский медико-фармацевтический институт, филиал ГБОН ВолгГМН Минздрава России
357532 Ставропольский край, Пятигорск, пр-т Калинина, 11, Россия
* E-mail: telitsyna.i@yandex.ru
** E-mail: mikhailov_val@mail.ru
*** E-mail: pae62@mail.ru
**** E-mail: gavrilova@binran.ru
Поступила в редакцию 15.03.2019
После доработки 05.04.2019
Принята к публикации 09.04.2019
Аннотация
С помощью светового, сканирующего электронного и конфокального лазерного сканирующего микроскопов исследована морфология пыльцевых зерен 24 образцов 11 видов рода Polygala, произрастающих, главным образом, на Северном Кавказе. Пыльцевые зерна изученных видов широкоэллипсоидальные, почти сфероидальные или слегка сплющенные, многобороздно-слитнооровые, довольно крупные, 30.0–57 мкм по максимальной оси. Характерной особенностью пыльцы истодов является эндоапертура в виде широкого внутреннего “пояска” – эндоцингулюма. По признакам скульптуры апокольпиумов и числу борозд выделено две палиногруппы, которые соответствуют 2 секциям рода Polygala. Описана индивидуальная изменчивость признаков формы пыльцевого зерна (расположение апертур). Обсуждается таксономическое значение палиноморфологических данных.
Род Polygala L. относится к семейству Polygalaceae и включает по разным оценкам от 475 до 720 видов, являясь крупнейшим в семействе (Paiva, Santos Dias, 1990; Castro et al., 2009). Представители рода распространены в разных областях Старого и Нового Света, за исключением Новой Зеландии и Арктики, 400 видов произрастают в Америке, 206 – в Африке, 70 – в Азии, 32 – в Европе и 12 – в Австралии (Paiva, Santos Dias, 1990). Polygala морфологически разнообразны, среди них встречаются травы, кустарники, лианы, деревья.
Система рода Polygala, его объем и родственные связи остаются дискуссионными и являются целью новейших интенсивных исследований с использованием и сопоставлением данных, полученных различными методами, в том числе и палинологическим. До сих пор спорными остаются вопросы внутриродового подразделения, объем и структура отдельных секций. Наиболее полной можно считать систему рода, предложенную J. Paiva (1998), согласно которой род Polygala подразделяется на 12 подродов: Acanthocladus (Klotzch ex Hassk.) Paiva, Badiera (DC.) Blake, Brachytropus (DC.) Chodat, Chodatia Paiva, Chamaebuxus (DC.) Duch., Gymnospora (Codat) Paiva, Hebecarpa (Chodat) Blake, Hebeclada (Chodat) Blake, Ligustrina (Codat) Paiva, Plebotaenia (Griseb.) Blake, Polygala (Chodat) Blake, Rhinotropis (Blake) Paiva.
С.А. Невский во “Флоре СССР” (Nevskii, 1949) описал 30 видов Polygala и выделил две секции: cекция 1. Mirgatoria Tamamsch. (3 вида) и cекция 2. Polygalon DC. c 17 видами, распределенными по 3 подсекциям. А.А. Гроссгейм во Флоре Кавказа (Grossgeim, 1962) описал 22 вида Polygala и отнес их к одноименной секции. А.С. Зернов (Zernov, 2006) во флоре Северо-Западного Кавказа признает всего 3 вида.
Несмотря на то, что первые упоминания о пыльце Polygalaceae появились еще в конце 19 века и крупнейший монограф семейства R. Chodat (1889) отмечал, что признаки пыльцевых зерен можно считать одними из важнейших характеристик семейства, работ по палиноморфологии Polygala до сих пор сравнительно немного. Имеются описания и изображения пыльцы лишь небольшого числа видов из разных секций (Larson, Skvarla, 1961; Villanueva, Ramos, 1986; Paiva, Santos Dias, 1990; Furnes, Stafford, 1995; Banks et al., 2008; Castro et al., 2009 и др.). Имеются сведения о пыльце бразильских (Aguiar et al., 2007), таиландских (Krachai et al., 2009) видов Polygala. Самой значительной остается работа H. Banks с соавторами (2008), посвященная морфологии пыльцы семейства Polygalaceae. В работе приведены описания и фотографии (СМ, СЭМ, ТЭМ) пыльцы 23 видов Polygala, кроме того приведен список еще 125 видов, сведения о пыльце которых были опубликованы ранее другими исследователями.
Палинологические данные неоднократно использовались разными авторами как для разграничения родов в семействе Polygalaceae (Chodat, 1896; Erdtman, 1944; Simpson, Skvarla, 1981; Paiva, 1998), так и для уточнения внутриродового подразделения Polygala. E. Villanueva и A. Ramos (1986), изучив пыльцу 10 видов Polygala, выделили 3 палинотипа, которые, по мнению авторов, соответствуют 3 подродам. В пределах подрода Polygala так же были установлены палинологические группы, соответствующие 2 секциям этого подрода. J. Paiva (1998) использовал палиноморфологические данные при выделении подродов Acanthocladus, Badiera, Plebotaenia из подрода Ligustrina. По морфологиии пыльцы 22 видов рода Polygala из 3 подродов (Chamamaebuxus, Chodatia, Rhinotropis) был сделан вывод, что характеристики пыльцы являются полезными при уточнении границ этих подродов и связей между ними (Castro et al., 2009). Авторы отмечают перспективность использования палинологических данных, наряду с молекулярным анализом, для уточнения вопросов систематики рода.
Значение признаков пыльцы для таксонов более низкого ранга оценивается неоднозначно из-за высокой вариабельности палиноморфологических признаков (Castro et al., 2009). Однако исследование пыльцы 15 бразильских видов истодов из подрода Hebeclada показало, что палиноморфологические признаки могут быть дополнительными характеристиками отдельных видов (Aguiar et al., 2007).
Ультраструктура оболочки пыльцы, взятой из живых пыльников, была изучена с помощью ТЭМ на примере P. fruticosa Berg. (Piava, Santos Dias, 1990). Для некоторых видов описаны срезы оболочек ацетолизированных пыльцевых зерен (Banks et. al., 2008). На ультратонких срезах показано, что в области мезокольпиумов и на полюсах экзина двуслойная, представлена слоями экт- и эндэкзины. Эктэкзина имеет толстый тектум и невысокие, тонкие, нередко слегка изогнутые, частые столбики (колумеллы), которые опираются на тонкий подстилающий слой. Эндэкзина толстая, ламеллятная, ламеллы короткие, переплетаются между собой, рыхло расположенные.
Тонкий подстилающий слой и толстая эндэкзина отчетливо видны на ультратонких срезах, исследованных с помощью ТЭМ (Paiva, Santos Dias, 1990; Banks et al., 2008). Эндэкзина ламеллятная, часть, примыкающая к подстилающему слою, образована редкими ламеллами, ниже лежащие ламеллы плотно прилегают друг к другу. Мембрана борозд образована только эндэкзиной, толщина которой заметно меньше чем на мезокольпиумах.
Пыльцу истодов флоры России до сих пор подробно не исследовали. Палиноморфологические данные известны лишь для 4 видов Polygala, произрастающих на территории Российской Федерации, два из них P. comosa и P. sibirica встечаются на Кавказе (Kupriyanova, Aleshina, 1978). Из кавказских истодов изучена пыльца P. major (Reill, 1995).
Цель данной работы – оценка таксономической значимости признаков пыльцевых зерен представителей Polygala, произрастающих на территории Кавказа и расширение представлений о пыльце рода в целом. Также была описана обнаруженная индивидуальная изменчивость признаков формы пыльцевого зерна (числа и расположения апертур, определяющих морфологический тип пыльцевого зерна и свойства его симметрии).
МАТЕРИАЛ И МЕТОДИКА
Материалом для исследования послужили зрелые пыльцевые зерна, взятые с гербарных образцов, хранящихся в Гербарии Ботанического института им. В.Л. Комарова (LE). В работе использовали методы световой (СМ), конфокальной лазерной сканирующей (КЛСМ), сканирующей электронной микроскопии (СЭМ). Для светооптического исследования применяли классический ацетолизный метод Эрдтмана (Erdtman, 1952). Методика исследования пыльцы с помощью КЛСМ описана в работе Gavrilova et al (2018). Световые микрофотографии изготовлены с увеличением 1000, на микроскопе Micmed-6 (ЛOMO).
Детали строения поверхности пыльцевых зерен уточняли на сканирующем электронном микроскопе JEOL JSM-6390 в центре коллективного пользования Ботанического института им. В.Л. Комарова. Для исследования необработанную пыльцу извлекали из пыльников и закрепляли на специальном столике с помощью двусторонней липкой ленты, затем напыляли сплавом золота и палладия в вакуумной установке.
Описания проводили по общепринятой схеме. Учитывали: тип и число апертур, форму и очертания пыльцевых зерен, размеры полярной оси и экваториального диаметра, особенности строения борозд и межапертурных участков, толщину экзины и характер скульптуры (Kuprijanova, Aleshina, 1967, 1972). Измерения проводили с помощью окулярмикрометра. В каждом образце измеряли не менее 15 пыльцевых зерен.
Исследованные образцы:
1) P. albovii Kem.:
1. Nat. Абхазская АССР, Гудаутский район, с. Лыхны, на табачных плантациях. 8.IХ.1950. Собр. Шакрон Л.И. Опр. С. Тамамшян.
2. Цебельда. Луга. Апрель. 1951. Собр. О. Деклеизо; опр. Кемулярия-Натадзе.
2) P. alpicola Rupr.:
1. Северный Кавказ, КБ АССР, Советский р-н, с.-з. окрестности сел. Безенги, левый берег р. Черек Безенгийский, подъем к Скалистому Хребту (Хуламская стена) 1700 м. н.у.м. 20 июля 1989 г. № 99 Собр. Ю. Меницкий; Опред. Т. Попова.
3) P. amoenissima Tamamsch.:
1. Дагестан, Рутульский р-н, с. Хлют, правый берег реки Самур. Северный осыпной склон, 25.VI.1961 г. Собр. и опр. А. Раджи.
2. Артвинский окр. между сс. Мериа и Даба. В лесу. 17.VII.1911 Собр. Н. Введенский; Опр. С. Тамамшян.
4) P. anatolica Boiss. et Heldr.:
1. Южно-Карабахская флористическая экспедиция 1948 г. Азербайджан, Карягинский р., между с. Туг и г. Карягино, каменистый кустарниковый склон. Собр. М.И. Кирпичников и Л.А. Смольянинова, 2.VI.1948 г.; Опред. С. Тамамшян.
2. Дагестан, Урочище Голок, субальпийские луга, южный склон 29.VII.1948 г. Собр. и опр. Косых.
5) P. caucasica Rupr.:
1. Дагестан, хр. Кукуртбаш, ущ. Бацергол, степной склон.
31.V.1956 Собр. Медведева Л.И., Надежина Т.П., Настенко Н.П., Соколов П.Д. Опред. С. Тамамшян.
2. Терраса Судахар, 23.VI.2018 г. Собр. Телицына.
3. Гуниб, 23.VI.2018 г. Собр. Коцеруба.
6) P. comosa Schkuhr.:
1. Флора Армении. Закавказская Экспедиция академии наук. 1930 г. Сев.-вост. побережье оз. Севан (Гокча) 1-е ущелье Так-Агач. Лесной склон у спуска к нижнему Асману. 29.V.1930 г. Собр. О. Полянская. Опред. С. Тамамшян.
2. Дагестан. Архасский хребет, субальпийский луг. 29.V.1964 г. Собр. Лесконог, опр. Темирсултанов.
7) P. hybrida DC. Сев-вост. побережье оз. Севан (Гокча), 1-е ущелье Так-Агач. Лесной склон у спуска к Нижнему Асману. 29.V.1930. Собр. О. Полянская. Опр. С. Тамамшян.
8) P. major Jacq. Краснодарский край, окр. г. Геленджик, 15 км к югу, Михайловский перевал, лес. 3.VIII.1990. № 22. Собр. Ю. Меницкий, Т. Попова, О. Морозенко.
9) P. sibirica L. Дагестан. VII. 1961 г. Собр. и опред. А. Раджи.
10) P. sosnowskii Kem. – Nat. 1. Северный Кавказ. Кабардино-Балкария. Чегемский район, выше водопадов по дороге к Актопраку. Лев. берег р. Чегем. 15.VII.1988. Собр. Т.Н. Попова. Опред. Н. Портениер.
11) P. transcaucasica Tamamsch.:
1. Армянская ССР Кировоканский район. Травянистый склон в грабовом лесу на склоне горы Сернех близ Кировокана (Алексондрополя) 1600 м. 14.VII.1952 г. Собр. Иг.В. Васильев, Н.Н. Васильева. Опред. С. Тамамшян.
Также для сравнения морфологии отклоняющихся форм пыльцы в статье приведены изображения пыльцы двух видов (P. austro-mexicana, P. triphylla), принадлежащих к другим секциям и не произрастающих на Кавказе.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Описания пыльцевых зерен исследованных видов представлены в табл. 1.
Таблица 1.
Описание пыльцевых зерен исследованных видов Polygala Table 1. The description of pollen grains of the studied Polygala species
| Виды Taxa |
Число
апертур Aperture number |
Полярная
ось (мкм) Polar axis length (μm) |
Экватори-
альный
диаметр (мкм) Equatorial axis length (μm) |
Ширина мезо-
кольпиума
(мкм) Mesocolpium width (μm) |
Ширина
борозд (мкм) Colpus width (μm) |
Ширина
орового
пояска (мкм) Endocingulum width (μm) |
Толщина
экзины (мкм) Exine thickness (μm) |
Скульптура
апокольпиума Sculpture on the apoco- lpium |
|---|---|---|---|---|---|---|---|---|
| P. albovii | ||||||||
| образец (sample) 1 | 14–15 | 47.0–54.6 | 50.0–57.5 | 5.6–6.0 | 5.2–5.8 | 4.7–5.0 | 3.0 | сетчатая (reticulate) |
| образец (sample) 2 | 12–14 | 47.5–52.7 | 48.8–50.0 | 6.7–8.0 | 6.0–6.5 | 5.0–6.0 | 2.5–3.0 | сетчатая (reticulate) |
| P. alpicola | 10–12 | 47.5–51.7 | 48.0–51.0 | 6.0–7.0 | 5.0–5.5 | 5.0 | 2.5–3.0 | сетчатая (reticulate) |
| P. amoenissima | 10–12 | 45.6–47.9 | 45.0–46.8 | 5.8–6.5 | 5.0–5.6 | 5.0 | 2.5–3.0 | сетчатая (reticulate) |
| P. anatolica | ||||||||
| образец (sample) 1 | 10–11 | 40.0–45.0 | 40.0–45.0 | 4.2–5.2 | 5.1–6.0 | 5.0 | 2.5–3.0 | сетчатая (reticulate) |
| образец (sample) 2 | 10–11 | 37.8–40.0 | 35.0–40.0 | 4.0–4.5 | 4.0–4.5 | 5.0 | 2.5–3.0 | сетчатая (reticulate) |
| P. caucasica | ||||||||
| образец (sample) 1 | 11–13 | 38.2–43.0 | 40.0–44.0 | 4.5–5.3 | 4.5–5.0 | 5.0 | 2.5–3.0 | сетчатая (reticulate) |
| образец (sample) 2 | 13–15 | (30)33.8–40.0 | (30)38.0–42.0 | 4.5–4.7 | 4.5–4.7 | 4.0 | 2.5–3.0 | сетчатая (reticulate) |
| образец (sample) 3 | 12–13 | 30.0–42.7 | 30.0–42.0 | 4.5–4.8 | 3.8–4.5 | 5.0 | 2.5–3.0 | сетчатая (reticulate) |
| образец (sample) 4 | 11–12 | 37.6–40.0 | 40.0–44.3 | 4.5–5.0 | 5.3–5.4 | 4.5–5.0 | 2.5–3.0 | сетчатая (reticulate) |
| P. comosa | ||||||||
| образец (sample) 1 | 11–12 | 35.0–40.0 | 35.0–40.0 | 4.5–5.0 | 5.0–5.5 | 5.0 | 2.5–3.0 | сетчатая (reticulate) |
| образец (sample) 2 | 11–12 | 40.0–45.3 | 36.5–40.0 | 4.0–7.0 | 5.3–6.4 | 4.4–5.0 | 2.5–2.7 | сетчатая (reticulate) |
| P. hybrida | 11–13 | 35.8–45.0 | 35.5–40.0 | 4.3–4.5 | 4.0–5.0 | 3.7–4.0 | 2.5–3.0 | сетчатая (reticulate) |
| P. major | 12–15 | 35.7–50.0 | 35.5–47.5 | 4.5–4.7 | 4.0–4.8 | 4.5–4.7 | 2.5–3.0 | сетчатая (reticulate) |
| P. sibirica | 18–19 | 40.0–50.0 | 38.8–51.4 | 3.0–4.5 | 2.8–3.0 | 6.0 | 2.5–3.0 | морщинистая (rugulate) |
| P. sosnowskii | 16–18 | 34.0–40.0 | 36.2–38.8 | 3.0–4.5 | 2.8–3.0 6 | 6.0 | 2.5–3.0 | морщинистая (rugulate) |
| P. transcaucasia | 11–13 | 50.0–55.0 | 46.2–53.0 | 4.0–5.0 | 5.0–6.5 | 9.0 | 2.5–3.0 | сетчатая (reticulate) |
Типичные пыльцевые зерна исследованных видов Polygala (табл. I) изополярные, радиально-симметричные, многобороздно-слитнооровые. Форма широкоэллипсоидальная или почти сфероидальная, в пределах образца меняется незначительно. Исследованные виды по этому признаку пыльцы не различаются. В очертании с полюса пыльцевые зерна округлые, многолопастные, с экватора широкоэллиптические, округлые, реже округло угловатые.
Таблица I. Морфология пыльцы представителей рода Polygala (СМ, СЭМ).
1–5 – P. albovii; 6–10 – P. caucasica; 11–15 – P. hybrida; 16–20 – P. anatolica; 21–25 – P. transcaucasica; 26–30 – P. sosnowskii; 31–35 – P. sibirica.
1, 3, 6, 11, 13, 18, 21, 23, 26, 28, 31, 33 – общий вид пыльцевого зерна с полюса. 2, 4, 7, 9, 12, 14, 17, 19, 22, 24, 27, 29, 32, 34 – общий вид пыльцевого зерна с экватора. 5, 15, 20, 25, 30, 35 – скульптура поверхности. 8, 10 – сколы пыльцевой оболочки. 1, 2, 6, 7, 11, 12, 16, 17, 21, 22, 26, 27, 31, 32 – СМ; 3–5, 8–10, 13–15, 18–20, 23–25, 28–30, 33–35 – СЭМ
Масштабные линейки, мкм: 1–4, 6–14, 16–19, 21– 24, 26– 29, 31–34 – 5; 5, 15, 20, 25, 30, 35 – 1.
Plate I . Pollen morphology of Polygala (LM, SEM).
1–5 – P. albovii; 6–10 – P. caucasica; 11–15 – P. hybrida; 16–20 – P. anatolica; 21–25 – P. transcaucasica; 26–30 – P. sosnowskii; 31–35 – P. sibirica.
1, 3, 6, 11, 13, 18, 21, 23, 26, 28, 31, 33 – polar view of pollen grain; 2, 4, 7, 9, 12, 14, 17, 19, 22, 24, 27, 29, 32, 34 – equatorial view of pollen grain; 5, 15, 20, 25, 30, 35 – exine ornamentation; 8, 10 – break of pollen wall (exine).
1, 2, 6, 7, 11, 12, 16, 17, 21, 22, 26, 27, 31, 32 – LM; 3–5, 8–10, 13–15, 18–20, 23–25, 28–30, 33–35 – SEM.
Scale bars, μm: 1–4, 6–14, 16–19, 21–24, 26–29, 31, 33 – 5; 5, 15, 20, 25, 30, 33 – 1.
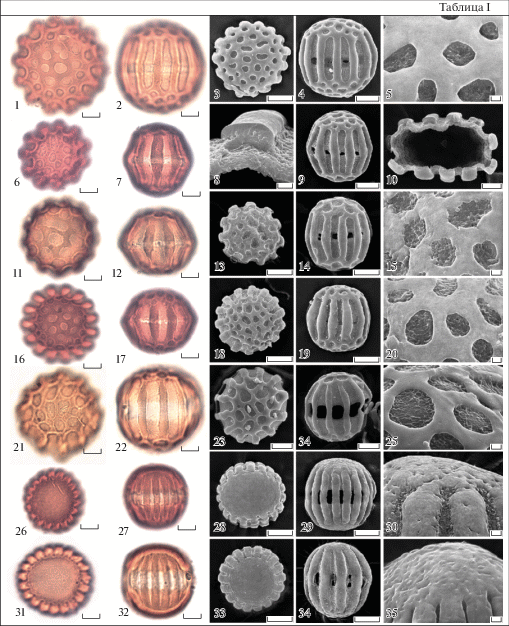
Число апертур у разных видов различно и не стабильно в пределах одного вида. Встречаются виды с 10–11, 13–15 и 18–19 бороздной пыльцой. M. Labouriau и L. Morphy (1969) показали отсутствие связи между размером пыльцевого зерна и числом борозд. К таким же выводам пришли и H. Banks с соавторами (2008). В нашем исследовании так же не наблюдалось такой связи. У одного из образцов P. albovii (табл. I, 1–5) были обнаружены самые крупные пыльцевые зерна с полярной осью 57.5 мкм, при этом максимальное число борозд у пыльцы этого вида – 15; тогда как у P. sosnowskii и P. sibirica (табл. I, 26–35) пыльцевые зерна с 18–19 бороздами, а размеры пыльцы по длинной оси не превышают 40 мкм у P. sosnowskii и 52 мкм у P. sibirica. В пределах одного образца у P. sosnowskii были обнаружены более крупные зерна с 16 бороздами и более мелкие с 18, и наоборот. В разных образцах P. caucasica (табл. I, 11–15) встречаются пыльцевые зерна одинаковых размеров с разным числом борозд.
По общепринятой классификации размеров (Erdtman, 1952) пыльца изученных видов занимает промежуточное положение между средними (25–50 мкм) и крупными (50–100 мкм). Длина максимальной оси (полярной оси или экваториального диаметра) у пыльцы изученных видов Polygala обычно варьирует в пределах 40–50 мкм, реже превышает 55 мкм. Самая крупная пыльца (полярная ось 50.0–54.6 мкм, экваториальный диаметр 50.0–57.5) у P. albovii. Самые мелкие пыльцевые зерна с максимальной осью не более 40 мкм у P. sosnowskii (полярная ось 34.0–40.0 мкм, экваториальный диаметр 36.2–38.8 мкм). Разница в размерах пыльцевых зерен из одного образца редко превышает 5 мкм. Однако в препаратах двух образцов P. caucasica, разница между самыми мелкими и самыми крупными зернами более 12.5 мкм.
На сколах пыльцевых зерен, исследованных с помощью СЭМ (табл. I, 8,10), видна структура экзины. На сколе мезокольпиума (табл. I, 8) отчетливо виден очень толстый тектум, тонкие, короткие, близко расположенные столбики и толстый слой, в котором не видна граница между подстилающим слоем и эндэкзиной. Толщина экзины 2.5–3.0 мкм. Пыльца всех исследованных видов имеет экзину близкой толщины, по этому признаку виды не различаются.
Апертуры сложные. Экзоапертуры представлены недлинными, широкими, меридиональными бороздами (кольпами). Края борозд слегка неровные, более или менее параллельные, концы большей частью широко закругленные. Эндоапертура (ора) – экваториальный поясок или endocingulum (Punt et al., 2007) с более или менее четкими, ровными, реже волнистыми краями. Исследование с помощью КЛСМ выявляет сложную конфигурацию внутренней поверхности пыльцевой оболочки в районе края эндоапертуры (табл. II, 18) у видов P. sosnowskii и P. sibirica. Пыльцевые зерна разных видов различаются по ширине пояска. У большинства видов поясок широкий, до 5.0–5.5(6.0) мкм. Самый широкий оровый пояс (около 9 мкм) отличает пыльцу P. transcaucasica (табл. I, 21–25). У пыльцы P. alpicola, P. hybrida (табл. I, 16–20) и 2 образцов P. caucasica поясок неширокий, с нечеткими границами.
Таблица II. Атипичные и типичные пыльцевые зерна представителей рода Polygala с изогнутой эндоапертурной зоной (СМ, СЭМ, КЛСМ).
1–5 – P. caucasica; 6 – P. anatolica; 7 – P. major; 8 – P. sosnowskii; 9, 10 – P. austro-mexicana; 11, 12 – P. amoenissima; 13–16 – P. triphylla; 17 – P. albovii; 18 – P. sosnowskii; 19 – P. anatolica.
1–8 – отклоняющиеся формы с изогнутой эндоапертурной зоной. 9, 10, 15, 16 – типичные формы с изогнутой эндоапертурной зоной. 1–9, 13, 15, 17 – общий вид пыльцевого зерна с экватора; 10, 14, 16 – общий вид пыльцевого зерна с полюса; 11, 12 – 3-полюсное пыльцевое зерно в двух проекциях; для каждой проекции показан верхний и нижний фокус; 18 – вид внутренней поверхности края эндоапертуры. 19 – атипичное пыльцевое зерно с разомкнутым эндоапертурным пояском.
1–14 – СМ; 15, 16 – СЭМ; 17–19 – КЛСМ.
Масштабные линейки, мкм: 1–17, 19 – 5; 18 – 1.
Plate II . Atypical and typical pollen grains of Polygala with curved endocingulum (LM, SEM, CLSM).
1–5 – P. caucasica; 6 – P. anatolica; 7 – P. major; 8 – P. sosnowskii ; 9, 10 – P. austro-mexicana; 11, 12 – P. amoenissima; 13–16 – P. triphylla; 17 – P. albovii; 18 – P. sosnowskii; 19 – P. anatolica.
1–8 – atypical forms with curved endocingulum; 9, 10, 15, 16 – typical forms with curved endocingulum. 1–9, 13, 15, 17 – equatorial view of pollen grain; 10, 14, 16 – polar view of pollen grain. 11, 12 – atypical 3-polar pollen grain in two views; the upper and lower foci are given for each view; 18 – inner surface of endoaperture edge. 19 – atypical pollen grain with open endocingulum.
1–14 – LM; 15, 16 – SEM; 17–19 – CLSM.
Scale bars, μm: 1–17, 19 – 5; 18 – 1.
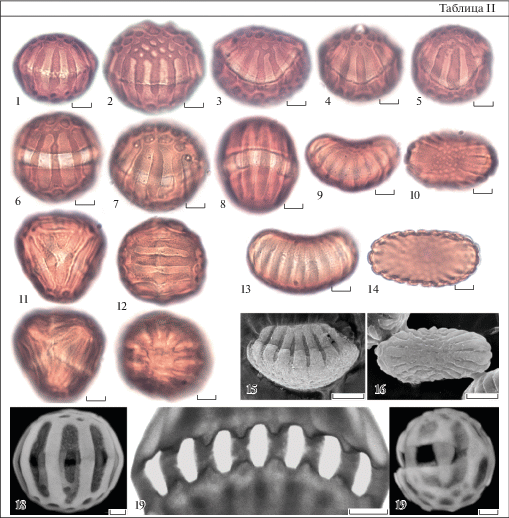
Мембрана борозд мелко шероховатая или неясно гранулярная. Пыльцевые зерна P. amoenissima отличаются от пыльцы других изученных видов отчетливо выраженным гранулярным рельефом поверхности мембраны борозд; гранулы довольно крупные, плотно прилегают друг к другу. Мембрана апертур тонкая, при обработке пыльцы часто разрывается в области ор. Лучше всего мембрана в области ор сохранилась у пыльцы разных образцов P. caucasica.
Мезокольпиумы узкие, их ширина в большинстве случаев незначительно отличается от ширины борозд. Однако у пыльцы P. transcaucasica (табл. I, 22, 24) борозды заметно шире мезокольпиумов, а у пыльцевых зерен с 16–19 бороздами, P. sosnowskii и P. sibirica (табл. I, 26–35) борозды узкие и ширина их меньше ширины мезокольпиумов. Поверхность мезокольпиумов в большинстве случаев почти гладкая.
Апокольпиумы широкие, плоские. По внешнему виду апокольпиумов и характеру рельефа на полюсах пыльцевых зерен исследованные виды можно разделить на 2 группы.
Первая группа включает P. albovii, P. alpicola, P. amoenissima, P. anatolica, P. caucasica, P. comosa, P. hibrida, P. major, P. transcaucasica (табл. I, 1–25). На полюсах пыльцевых зерен скульптура сетчатая. Просветы ячей крупные, 1.8–4.0 мкм по максимальной оси, с четко очерченными ровными или слегка волнистыми краями, в очертании округлые или округло-угловатые. Самые крупные ячеи чаще располагаются в середине апокольпиума, а у периферических размеры заметно меньше. Иногда соседние ячеи сливаются. Ячеи неглубокие, дно имеет нерегулярно морщинистую или ямчатую поверхность. Стенки гладкие, широкие, 1.8–4.0 мкм. Число ячей у разных видов неодинаково и незначительно варьирует в пределах одного образца. Изученные виды из этой группы палиноморфологически сходны. Только пыльца P. transcaucasica отличается от пыльцы других видов узкими мезокольпиумами, широкими бороздами и широким оровым пояском.
Вторая группа включает только 2 изученных вида: P. sosnowskii и P. sibirica (табл. I, 26–35). У пыльцы этих видов поверхность апокольпиумов неясно морщинистая, детали рельефа доступны для изучения только с помощью СЭМ.
С палиноморфологической точки зрения была проанализирована система рода Polygala, предложенная для Флоры СССР (Nevskii, 1949). Согласно этой системе, изученные нами виды распределяются по 2 секциям. Палиноморфологически эти 2 секции хорошо различаются и соответствуют 2 выделенным палиногруппам.
Секция Mirgatoria включает 2 изученных вида: P. sibirica и P. sosnowskii. Пыльцевые зерна на полюсах шероховатые или почти гладкие. Число борозд 18–19. Пыльцевые зерна этих двух видов незначительно различаются степенью выраженности рельефа на полюсах.
Секция Polygalon. Все изученные виды относятся к одной подсекции Eupolygalon Tamamsch. и характеризуются пыльцой с крупными ячеями на полюсах и числом борозд меньше 18. В пределах секции виды по пыльце отличаются незначительно. Среди исследованных видов выделяется пыльца P. transcaucasica с очень широким экваториальным пояском (табл. I, 22, 24).
Индивидуальная изменчивость формы пыльцы.
Кроме типичных пыльцевых зерен с описанным меридиональным расположением апертур (табл. I), в образцах 5 из 11 изученных видов были обнаружены пыльцевые зерна, отклоняющиеся от типичной формы по расположению апертур (табл. II). У этих отклоняющихся форм эндоапертурный поясок лежит не в плоскости экватора, а изогнут, напоминая по форме шов на теннисном мяче (табл. II, 1–8; Пожидаев, 2015). Степень изгиба пояска может меняться в одном образце и в разных образцах, от незначительной до хорошо заметной. Отклоняющиеся пыльцевые зерна с изогнутой эндоапертурной зоной имеют симметрию, характерную для типичной пыльцы некоторых других видов Polygala (табл. II, 9, 10, 13–16), относящихся к секциям, отсутствующим во флоре Кавказа.
Очень редко у некоторых пыльцевых зерен эндоапертурный поясок имеет форму не замкнутого кольца, а размыкается в дугу (табл. II, 19). В двух образцах P. amoenissima в качестве редкого отклонения обнаружены единичные трехполюсные пыльцевые зерна (табл. II, 11, 12). Такая конфигурация апертур никогда не бывает типичной, но, несмотря на необычность формы, расположение ее апертур не хаотично и форма выглядит геометрически упорядоченной.
Список литературы
Aguiar A.C.A., Gonçalves V.M., Gonçalves-Esteves V., Yamamoto K. 2008. Palynotaxonomy of Brazilian species of Polygala subgenus Hebeclad. – Bot. J. Linn. Soc. 157: 609–619.
Banks H., Klitgaard B.B., Claxton F., Forest F., Crane P.R. 2008. Pollen morphology of the family Polygalaceae (Fabales). – Bot. J. Linn. Soc. 156: 253–289.
Castro S., Silveira P., Navarro L., Paiva J. 2009. Pollen morphology of Chamaebuxus (DC.) Schb., Chodatia Paiva and Rhinotropis (Blake) Paiva (Polygala L., Polygalaceae). – Grana. 48 (3): 179–192. https://doi.org/10.1080/00173130902938428
Chodat R. 1889. Le pollen des Polygalacées. Arch. Sc. Phys. Nat. Guive. Sér. 3. 21 (3): 269–270.
Chodat R. 1896. Polygalaceae. – In: Die natürlichen Planzenfamilien. Vol. 3 (4). Leipzig. P. 323–345.
Erdtman G. 1944. The systematic position of genus Diclidanthera Mart. – Bot. Not. 38: 80–84.
Erdtman G. 1952. Pollen morphology and taxonomy. Angiosperms. Stockholm. 539 p.
Furnes C.A., Stafford P.J. 1995. Polygalaceae. The Northwest European Pollen Flora 55. – Rev. Palaeobot. Palynol. 88 (1–4): 61–82. https://doi.org/10.1016/0034-6667(95)00019-T
Gavrilova O., Zavialova N., Tekleva M., Karasev E. 2018. Potential of CLSM in studying some modern and fossil palynological objects. – J. Microscopy. 269 (3): 291–209. https://doi.org/10.1111/jmi.12639
[Grossgeim] Гроссгейм А.А. 1962. Флора Кавказа. Т. VI. М. Л. С. 59–67.
Krachai P., Chantaranothai P., Piwpuan N. 2009. Pollen characteristics of Polygala, Salomonia and Xanthophyllum (Polygalaceae) in Thailand. – Nat. Hist. J. Chulalongkorn Univ. 9(1): 27–34.
[Kupriyanova, Aleshina] Куприянова Л.А., Алешина Л.А. 1967. Палинологическая терминология покрытосеменных растений. Л. 84 с.
[Kupriyanova, Aleshina] Куприянова Л.А., Алешина Л.А. 1972. Пыльца и споры растений флоры Европейской части СССР. Т. 1. Л. 172 с.
[Kupriyanova, Aleshina] Куприянова Л.А., Алешина Л.А. 1978. Пыльца двудольных растений флоры Европейской части СССР (Lamiaceae-Zygophyllaceae). Л. С. 71–73.
Labouriau M.L.S., Morphy L. 1969. Pollen grains of plants of the ‘Cerrado’ – XVI. Myrsinaceae, Ochnaceae and Polygalaceae. – Anais da Academia Brasileira de Ciências. 41 (2): 249–258.
Larson D.A., Skvarla J. 1961. The morphology and fine structure of pollen of Polygala alba Nutt. & P. incarnatta L. – Pollen et Spores. 3 (1): 21–32.
[Nevskii] Невский С.А. 1949. Сем. LXXXV. Истодовые – Polygalaceae Lindl. – В кн.: Флора СССР. Т. 14. М. Л. С. 246–266.
Paiva J.A.R. 1998. Polygalarum Africanarum et Madagascar-iensium prodomus atque gerontogaei generic Hetrosamara Kuntze, a genere Polygala L. segregate et a nobis denuo recepti, synopsis monographica. – Fontqueria. 50: 1–346.
Paiva J., Santos Dias J.D. 1990. The pollen grain of Polygala fruticosa Berg. (Polygalaceae). – Anales Jard. Bot. Madrid. 47 (2): 377–385.
[Pozhidaev] Пожидаев А.Е. 2015. Рефренная структура биологического многообразия и теория филогенеза. Палеоботанический временник. Приложение к журналу “Lethaea rossica” Российский палеоботанический журнал”. 2: 115–127.
Villnueva E., Ramos A. 1986. Contribución al estudio polínico de Polygala L. (Polygalaceae) en la Península Ibérica. – Anales Jard. Bot. Madrid. 42 (2): 377–388.
[Zernov] Зернов А.С. Флора Северо-Западного Кавказа. 2006. М. С. 368.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал