

Ботанический журнал, 2019, T. 104, № 7, стр. 1059-1071
БИОМОРФОЛОГИЯ ЭНДЕМИКА ПАМИРО-АЛАЯ NEPETA CONSANGUINEA (LAMIACEAE): ОНТОГЕНЕЗ И АРХИТЕКТУРНЫЙ АНАЛИЗ
В. А. Черемушкина 1, А. Ю. Асташенков 1, *, М. Т. Бобоев 2, **
1 Центральный сибирский ботанический сад СО РАН
630090 г. Новосибирск, ул. Золотодолинская, д. 101, Россия
2 Хатлонский научный центр АН РТ
735360 Куляб, ул. Гулбог, 1, Таджикистан
* E-mail: astal@bk.ru
** E-mail: b_mario@mail.ru
Поступила в редакцию 10.06.2019
После доработки 25.06.2019
Принята к публикации 02.07.2019
Аннотация
Изучены онтогенез и особенности побегообразования особей Nepeta consanguinea. Установлено, что в зависимости от конкретных экологических условий особи представлены длиннокаудексовой или корневищно-каудексовой стержнекорневыми биоморфами. Формирование корневищно-каудексовой связано с изменением длины, структуры резида и положением почки возобновления, а также функционированием спящих почек. Архитектурный анализ показал, что иерархическая структура особи состоит из трех отличимых модулей, которые приводят к формированию двух разветвленных комплексов. Отсроченный архитектурный комплекс можно рассматривать как структурный элемент реитерации.
Архитектурный подход к описанию растений дает обоснованную интерпретацию процессов, связанных с их адаптацией к экологическим условиям обитания (Barthélémy, Caraglio, 2007; Bruy et al., 2018). В мире используют различные подходы к анализу структуры растений. Однако они все основаны на исследовании характера нарастания и ветвления основных структурных единиц побеговой системы и характера сочленения и пространственного расположения модулей разного ранга (Serebryakov, 1962; Troll, 1964; Halle, Oldeman, 1970; Gatsuk, 1974; Mazurenko, Khokhryakov, 1977; Serebryakova, 1977; Barthélémy, Caraglio, 2007; Kolegova, Cheryomushkina, 2012). Изменение функциональности отдельных модулей побеговой сферы и ее роль в трансформации жизненной формы трав отмечены во многих работах (Gatsuk, 1974; Dorokhina, 1978; Tortosa et al., 1996; Keller, 1997; Rua, 2003; Cheryomushkina, 2004; Notov, Kusnetzova, 2004; Savinykh, 2006).
Среди губоцветных Nepeta L. – один из многочисленных родов, входящих в крупную трибу Nepeteae Benth., большая часть видов которого освоила горные районы Центральной Азии. Среди всего разнообразия значительная доля Nepeta относится к эндемикам различных географических областей, а изучение устойчивости их популяций вызывает особый интерес в экологии растений. Произрастая на ограниченной территории, эндемики в процессе эволюции выработали специфические механизмы приспособления как на организменном, так и популяционном уровнях. Биоморфологический анализ таких групп растений с применением онтогенетического подхода позволит выявить основные структурные изменения и пластичность особей, которые отражают механизмы приспособления к определенным экологическим условиям. Комплексный анализ в изучении изменения морфы (sensu Nepeta) в эколого-ценотическом ряду даст возможность установить морфофилогенетические связи в развитии близкородственных видов и проследить основной тренд в соматической эволюции. Наиболее интересная в этом отношении оказалась sect. Macronepeta Benth., представители которой сохранились в настоящее время в географически отдаленных флористических областях Центральной Азии, Сибири и Дальнего Востока.
Цель работы – изучение онтогенеза и особенностей архитектуры особей эндемика Памиро-Алая Nepeta consanguinea Pojark.
МАТЕРИАЛЫ И МЕТОДЫ
N. consanguinea (Котовник родственный) во флоре СССР приводится как эндемик Памиро-Алая (Pojarkova, 1954). В филогенетическом отношении N. consanguinea относится к среднеазиатской ser. Gontsharovianeae Pojark. sect. Macronepeta (Budantzev, 1993). Указания Т.И. Цукерваник (Tsukervanik, 1987) в “Определителе Средней Азии” о распространении вида на хребте Петра I, вероятно, ошибочно, т.к. в фондах гербариев LE и TAD хранятся сборы только с Дарвазского и Вахшского хребтов. Собственно из экземпляров, собранных с Вахшского хребта, приводится первоописание таксона. Образцы из других районов Таджикистана отсутствуют.
N. consanguinea обитает на каменисто-мелкоземистых осыпях в верхней части пояса древесно-кустарниковой растительности – чернолесья (Juglans regia L.), термофильных арчовников с Juniperus seravschanica (Kom.) Nevski. Пределы распространения вида по относительным высотам находятся в узкой амплитуде от 2300 до 2600 м над уровнем моря. Наши исследования показали, что особи N. consanguinea встречаются также в составе нарушенной растительности розариев (Rosa kokanica (Regel) Juz.) и сообществах полусаванн на каменистых сухих склонах.
Материал собран с юго-западной оконечности хребта Сурх, входящего в единую цепь трансмеридиональных отрогов Вахшского хребта (N 38°38'43,0'', E 069°36'41,0'' 2412 м над уровнем моря). Исследовались особи, произрастающие в различных экологических условиях, образующие ценопопуляционные локусы (ЦЛ): 1) красномелкозёмистый осыпной склон юго-западной экспозиции крутизной 20°; 2) серомелкозёмистый с уплотненным субстратом склон южной экспозиции крутизной 20°, покрытый гранитными плитами и крупнообломочной материнской породой; 3) антропогенно нарушенный плакор межгорной долины; 4) борт технологического желоба дорожной насыпи.
При изучении онтогенеза была принята концепция дискретного его описания (Rabotnov, 1950; Uranov, 1975) и представления о поливариантности развития особей (Zhukova, 1995). Организация побега охарактеризована с позиции его структурно-функциональной дифференциации, предложенной W. Troll (1964). Тип биоморфы определен по расположению многолетних структур особи, воздействующих на среду обитания (Smirnova, 1987). Календарный возраст особей, в том случае, когда это представлялось возможным, установлен с помощью прямого подсчета колец на анатомических срезах корней и сохранившихся резидах. Архитектурный анализ выполнен с позиции концепции об архитектонике растений, предложенных F. Hallé et R.A.A Oldeman (1970), F. Hallé et al. (1978), D. Barthélémy, Y. Caraglio (2007). Архитектурное развитие N. consanguinea основано на трех иерархически соподчиненных единицах: модуль, разветвленный комплекс, особь (в понимании Barthélémy, Caraglio, 2007; Charles-Dominique, 2010).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Онтогенез
Биоморфологический анализ растений N. consanguinea показал, что типичная биоморфа особей – длиннокаудексовая стержнекорневая моноцентрическая. Однако этой биоморфе до генеративного состояния предшествует компактнокаудексовая, которая нами рассматривается как промежуточная. В зависимости от конкретных условий обитания биоморфа особей меняется с типичной на корневищно-каудексовую стержнекорневую неявнополицентрическую. Ниже приводится описание онтогенеза и механизмы формирования различных биоморф.
Длиннокаудексовая биоморфа
Прорастание семян надземное. Проросток – двусемядольное растение с одной парой настоящих зеленых листьев и главным корнем. У проростка слабо выражены гипокотиль и эпикотиль. В год прорастания семян, после отмирания семядолей, продолжая нарастать моноподиально, растения переходят в ювенильное состояние. Первичный побег – удлиненный, ортотропный, высотой от 1.5 до 4.5 см. За вегетационный сезон на побеге формируются 5–6 пар настоящих зеленых листьев. В пазухах всех листьев закладываются почки. Главный корень толщиной 0.1 см, длиной 5–10 см. В конце вегетации побег отмирает до первого удлиненного метамера. Сохранившаяся часть побега – резид, состоит из семядольного узла и оси первого метамера. За счет контрактильной деятельности корня резид погружается в почву, в результате чего почки, расположенные в узлах, оказываются под землей. Формируется первое звено каудекса. Обычно почки семядольного узла наиболее развитые, они прикрыты 4-мя крупными чешуями.
На второй год после таяния снега развертываются побеги возобновления, растения переходят в имматурное состояние. Нарастание особи сменяется на симподиальное. С этого состояния у особей N. consanguinea отмечается поливариантность развития, которая проявляется в различных способах нарастания. 1) Особи остаются однопобеговыми (рис. 1А). Побег возобновления развертывается из одной почки сохранившегося резида. Как правило, трогается в рост почка семядольного узла. Нередко в побегообразовании участвует почка первого метамера. Нереализованные почки становятся спящими. 2) Особи ветвятся (многопобеговые) (рис. 1B). Ветвление происходит в результате раскрытия двух почек семядольного узла и (или) почек первого длинного метамера – формируется первичный куст, состоящий из 2–3-х ортотропных вегетативных побегов. Побеги имматурных особей высотой 5–7 см.
Рис. 1.
Онтогенез особей Nepeta consanguinea разных биоморф.
Пути развития биоморфы: A – однопобеговой, B – ветвящейся (многопобеговой), С – длиннокаудексовой, D – корневищно-каудексовой с длинным корневищем, E – корневищно-каудексовой со смешанным корневищем. Онтогенетические состояния: p – проросток, j – ювенильное, im – имматурное, $v$ – виргинильное, g1 – молодое генеративное, g2 – зрелое генеративное, g3 – старое генеративное, ss – субсенильное, s – сенильное; черта – уровень почвы, стрелки – переход из одного онтогенетического состояние в другое и варианты развития; 1 – каудекс, 2 – главный корень, 3 – придаточный корень, 4 – корневище длинное, 5 – корневище короткое, 6 – геофильная часть растущего побега, возникшего из спящей почки, 7 – побег, возникший из почки возобновления, 8 – побег, возникший из спящей почки.
Fig. 1. Ontogenesis of Nepeta consanguinea individuals of different biomorphs.
Paths of biomorph development: A – one-shooted, B – branching (many-shooted), C – long-caudex, D – rhizomatous-caudex with long rhizome, E – rhizomatous-caudex with mixed rhizome. Ontogenetic states: p – seedling, j – juvenile, im – immature, v – virginal, g1 – young generative, g2 – mature generative, g3 – old generative, ss – subsenile, s – senile, line – ground level, arrows – transition from one ontogenetic state to another and its variants; 1 – caudex, 2 – main root, 3 – adventitious root, 4 – long rhizome, 5 – short rhizome, 6 – geophilous part of the growing shoot arisen from a dormant bud, 7 – shoot arisen from an innovation bud, 8 – shoot arisen from a dormant bud.
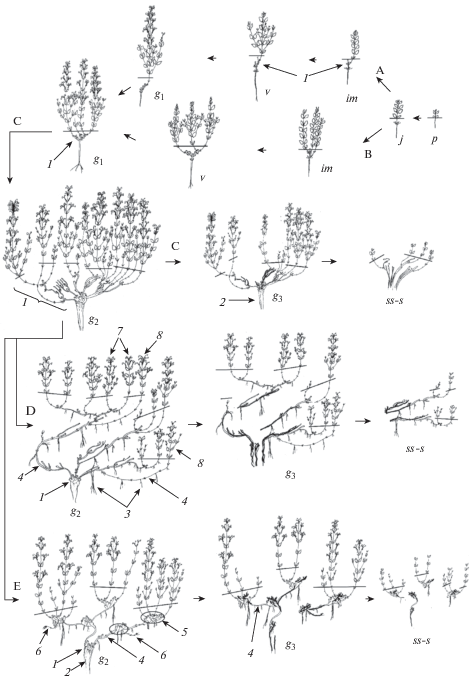
В зависимости от условий обитания подземная базальная часть побега разная. На склонах (ЦЛ 1, 2) нижняя часть побега состоит из метамеров со сближенными (3–4) и расставленными (1–2) узлами, а ее длина достигает 1.2 см. В условиях антропогенной нагрузки (ЦЛ 3, 4) базальная часть состоит из метамеров с сильно сближенными узлами (4–5) с чешуевидными и переходными листьями, длина участка не превышает 0.5 см. Почки возобновления в первом случае закладываются в пазухах листьев расставленных узлов, во втором – сближенных узлов. Нереализованные почки переходят в спящее состояние. После вегетации побег отмирает до базальной части, сохранившийся резид встраивается в общую структуру компактного каудекса. Главный корень превышает 10 см длины, он в апикальной части начинает слабо ветвиться. Ветвление не превышает двух порядков.
На третий год растения переходят в виргинильное онтогенетическое состояние. Оно характеризуется интенсивным ветвлением надземной части побегов возобновления. Боковые побеги развертываются из пазух каждого зеленого листа, их длина изменяется снизу вверх по материнскому побегу от 6.5 см до 2.5 см. После вегетации вся разветвленная надземная сфера отмирает. Виргинильные особи могут быть как однопобеговые, так и многопобеговые. Побегообразование осуществляется в первом случае за счет раскрытия только одной почки возобновления, во втором – за счет раскрытия двух и более почек. В зависимости от вариантов побегообразования компактный каудекс может быть разветвленным или одноосным. В том и другом случаях биоморфа остается моноцентрической. Главный корень слабо ветвится, утолщается до 0.4 см.
На четвертый год особи N. consanguinea зацветают. Нередко ускоренные темпы развития в прегенеративном периоде сопровождаются пропусками имматурного или виргинильного состояний. Так, нами обнаружены экземпляры, у которых первое цветение приходится на третий год жизни. У таких особей зацветают побеги, возникшие из спящих почек, расположенных в семядольном узле. Эти почки в течение двух лет подрастают, за это время происходит закладка зачаточных структур всей генеративной сферы. У особей с нормальным темпом развития генеративные побеги развертываются из почек возобновления.
У молодых генеративных растений формируется либо один генеративный побег, который надстраивает многолетнюю структуру каудекса по типу монохазия (главной оси), либо несколько (4–6) побегов (по типу дихазия/плейохазия). Однопобеговые особи после первого цветения развиваются как обычные кустящиеся растения.
Молодые генеративные растения компактнокаудексовой биоморфы представляют собой хорошо развитые кусты, в которых насчитывается от 5 до 15 цветущих побегов высотой 15–20 см. Побегообразование осуществляется за счет функционирования почек возобновления, расположенных в укороченной базальной части сохранившегося резида.
В структуре каждого генеративного побега выделяются следующие функциональные зоны: зона возобновления (ЗВ – 3–4 коротких метамера с чешуевидными листьями), средняя зона торможения (СЗТ – 1–2 длинных метамера с переходными листьями); зона обогащения (ЗО – 7–8 длинных метамеров с вегетативными побегами обогащения и паракладиями), зона главного соцветия (ГС – 5–6 длинных метамеров). Главное соцветие представляет собой рыхлую фрондозно-фрондулозную кисть, состоящую из супротивно расположенных на длинных осях дихазиев из монохазиев. После вегетации каждый побег отмирает до базальной части, близкорасположенные укороченные резиды участвуют в построении компактного каудекса. Ежегодно из почек каждого резида развертываются 2–3 побега возобновления. Главный корень хорошо выражен и достигает 1 см в диаметре. Длительность состояния составляет 4–5 лет.
На 9–10 год растения переходят в зрелое генеративное состояние. В кусте насчитывается 50–70 генеративных побегов высотой 35–50 см. Диаметр куста может превышать 50 см. Это состояние характеризуется пробуждением спящих почек, сохранившихся на многолетних частях каудекса. Возникшие из спящих почек побеги структурно отличаются от побегов, развившихся из почек возобновления. Отличия заключаются в увеличении числа коротких метамеров с чешуевидными листьями в базальной части до 7–9; отсутствии ЗО в результате ее замещения СЗТ; сокращении ГС до 3–4 метамеров, вплоть до полной редукции флоральных элементов.
В этом состоянии происходит трансформация биоморфы из компактной каудексовой в длиннокаудексовую (рис. 1С). Мы выявили два пути перестройки биоморфы: первый связан с удлинением геофильной части побега возобновления в результате его роста в плотном субстрате; второй – увеличением длины подземной части побега, возникшего из спящей почки. У некоторых экземпляров геофильная часть с длинными междоузлиями может увеличиваться до 10 см. С увеличением длины базального участка изменяется его структура. Так, почка возобновления из нижнего положения перемещается выше по оси на последний сохранившийся метамер, а нижележащие почки не реализуются и образуют протяженную нижнюю зону торможения (НЗТ). После вегетации надземная часть побега отмирает до длинного участка. Последующее побегообразование особи осуществляется за счет почек, расположенных на верхних длинных метамерах резида. Сохраняющиеся удлиненные резиды симподиально надстраивают каудекс, в результате чего формируется длиннокаудексовая биоморфа. Главный и боковые корни утолщаются (диаметр главного корня более 2.0 см), их покровная ткань частично слущивается. Длительность зрелого генеративного состояния колеблется от 10 до 15 лет.
Нами установлено, что наибольшая продолжительность пребывания особей в компактном состоянии и перестройка биоморфы в длиннокаудексовую происходит в условиях серомелкоземистого субстрата на склоне, покрытом крупнообломочной породой (ЦЛ 2). В таких условиях компактнокаудексовая биоморфа сохраняется достаточно долго (до середины зрелого генеративного состояния), а ее трансформация начинается на 10–17 году жизни особи. Наименьшая продолжительность (5–6 лет) и ранняя трансформация (в молодом генеративном состоянии) отмечены у особей, растущих на осыпном склоне с красномелкозёмистым субстратом (ЦЛ 1).
Приближаясь к 20–25-летнему возрасту, особи переходят в старое генеративное состояние. У растений происходит старческая партикуляция, которая протекает по линии паренхимных лучей, расположенных между проводящими пучками каудекса и главного корня. В результате разрушения центральной части куста частично обособляются длинные главы каудекса, но их связь с главным корнем сохраняется. Побегообразование происходит за счет функционирования почек возобновления на периферии куста. В кусте уменьшается число побегов, у некоторых экземпляров их число не превышает 20–25.
В постгенеративном периоде происходит полная дезинтеграция куста. Образуется клон, состоящий из материнской особи и дочерних партикул. Отделившиеся партикулы не омоложены. Их жизнеспособность обеспечивается участками главного корня с сохранившейся проводящей системой. Побегообразование происходит исключительно за счет спящих почек. Побеги возобновления структурно подобны побегам имматурных особей.
Произрастая на красномелкозёмистом осыпном склоне (ЦЛ 2) и в условиях антропогенной нагрузки (ЦЛ 3, 4), у особей возможна перестройка типичной биоморфы в корневищно-каудексовую. Механизм ее трансформации начинается в зрелом генеративном состоянии и связан с образованием корневища. Корневище в свою очередь может быть длинным или комбинированным (длинное в сочетании с коротким) гипогеогенного или гипоэпигеогенного происхождения.
Корневищная биоморфа
Длинное корневище формируется у особей в ЦЛ 2. Постоянное засыпание побегов, образование на них придаточных корней приводит к разрастанию особей на освободившихся участках склона (рис. 1 D). Побегообразование осуществляется за счет почек возобновления и спящих почек. Погребенные удлиненные части побегов становятся элементами длинного корневища гипоэпигеогенного происхождения, а геофильная часть побегов, возникших из спящих почек – гипогеогенного происхождения. Укореняющиеся побеги способствуют захвату территории, а за счет образования парциальных кустов – удержанию пространства. Корневая система становится смешанной. Появление парциальных кустов, связанных с семенной особью, приводит к формированию куртины, особи становятся неявнополицентрическими. Парциальные кусты малопобеговые (3–5 побега). Старение куртины сопровождается партикуляцией корневища, что приводит к обособлению неомоложенных кустов. Их онтогенез сокращенный, партикулы быстро стареют и отмирают.
Комбинированное корневище формируется у особей, растущих в нарушенных условиях (ЦЛ 3, 4). В таких местообитаниях пробуждается спящая почка, которая формирует побег с удлиненной геофильной частью (рис. 1E). Развитие такого побега можно оценить как способ продвижения особи в пространстве и удаление новых, молодых структур от компактной старой части растения. После отмирания надземной сферы побега сохраняется удлиненный резид – элемент длинного корневища. Этот резид становится основой для образования будущего парциального куста. На резиде появляются придаточные корни, один из которых в дальнейшем становится вторично стержнекорневым. Под влиянием антропогенной нагрузки (вытаптывание) основа парциального куста надстраивается короткими резидами – элементами короткого корневища. Они образуются из базальных частей поврежденных ортотропных побегов, возникших из почек возобновления. Парциальный куст многопобеговый (10–20 побегов). Пробуждение спящей почки, расположенной в кусте n-го порядка, приводит к развитию нового куста n + 1 порядка. Раскрытие спящей почки происходит нерегулярно. Так, нами установлено, что появление парциальных кустов n + 1 порядка происходит каждые 4 года. В результате появления новых укореняющихся парциальных кустов формируется куртина. Таким образом, многолетняя структура растений складывается из симподиальных систем резидов разной длины, связанных через материнскую особь с главным корнем.
В старом генеративном состоянии происходит полная дезинтеграции куртины. Парциальные кусты отделяются от материнской особи, в результате отмирания удлиненных частей корневища формируется клон. Он состоит из семенной особи и неомоложенных дочерних партикул. Партикулы находятся в том же состоянии, что и семенная особь, реже – старше на одно онтогенетическое состояние. Процесс партикуляции сопровождается задержкой в развитии и растянутыми темпам старения клона. Обособленные партикулы, за счет вторичного стержневого корня, остаются жизнеспособными и стареют одновременно с материнской особью.
Продолжительность данного состояния составляет 5–6 лет.
В постгенеративном периоде клон состоит из отдельных вегетативных партикул и живых частей семенной особи. Подсчитать длительность состояния из-за разрушения многолетних структур не представляется возможным. Однако, судя по сохранившимся элементам корневища, продолжительность постгенеративного периода не превышает 1–3 лет.
Таким образом, общая продолжительность онтогенеза семенной особи N. consanguinea колеблется от 25 до 35 лет.
Архитектурный анализ
Ранее нами были изучены биоморфология и особенности развития некоторых видов Nepeta Памиро-Алая, Тянь-Шаня и Памира. У них установлены варианты развития гомологичных структурных элементов, отражающих родство (Astashenkov, Cheryomushkina, 2016; Astashenkov et al., 2017; Astashenkov et al., 2018). Среди изученных Nepeta такой вариант развития и механизм побегообразования как у особей N. consanguinea нами описаны впервые и у изученных ранее видов нами не встречались.
Нами проведен архитектурный анализ особей N. consanguinea, ветвление которых начинается на начальных этапах онтогенеза (рис. 1B) и соответствует развитию корневищной биоморфы (1B, 1C). У особей выделены модули, отличающиеся по структуре и времени их появления в онтогенезе (рис. 2). Модуль I – ортотропный, моноподиально нарастающий (в надземной части ветвистый/не ветвистый) побег, его терминальная почка (формирует/не формирует генеративную сферу), ежегодно отмирает. Сохранившийся короткий резид симподиально надстраивает многолетнюю компактную структуру особи. Последовательное развитие модулей и сочленение их базальными участками формируют разветвленный комплекс (РК I). Модуль II гомологичен M I, он формируется в зрелом генеративном состоянии из почки возобновления; М II – анизотропный, имеющий длинный плагио-, анизотропный (геофильный/надземный) участок. Сохранившийся длинный резид – элемент удлиненного каудекса или эпигеогенного корневища, также симподиально надстраивает многолетнее тело растения и входит в состав РК I. Функционально этот модуль обеспечивает продвижение особи и удержание территории. М III – анизотропный (в надземной сфере ветвящийся/не ветвящийся) побег, возникший из спящей почки, ежегодно отмирающий. На его основе формируется отсроченный разветвленный комплекс РК II, который надстраивается модулями I и II. Сохранившаяся базальная плагиотропная геофильная часть модуля становится элементом гипогеогенного корневища. Каждый раз, с периодом кратным 4 годам, формируется повторяющийся отсроченный РК II n + 1 порядка. Процесс естественной партикуляции, связанный с некрозом многолетних структур, сопровождается обособлением РК II (в том числе РК II n-го порядка).
Рис. 2.
Архитектурный анализ особей Nepeta consanguinea (корневищной биоморфы).
М – модуль (I, II, III), 1 – отмершая надземная часть побега, 2 – сохраняющаяся подземная часть побега (резид) модуля I, 3 – резид модуля II, 4 – резид модуля III, РК – разветвленный комплекс (I, II), РК n-го порядка – разветвленный комплекс возрастающего порядка. Стрелками обозначен путь построения архитектуры особи. Пунктиром обозначены отмершие РК.
Fig. 2. Architectural analysis of Nepeta consanguinea individuals (rhizomatous biomorph).
M – modules (I, II, III), 1 – dead overground part of the shoot, 2 – remaining underground part of the shoot (residue) of module I, 3 – residue of module II, 4 – residue of module III, PK – branched complex (I, II), n-order PK – branched complex of increasing order. The arrows show the path to construct the architecture of the individual. Dotted lines indicate dead PK.
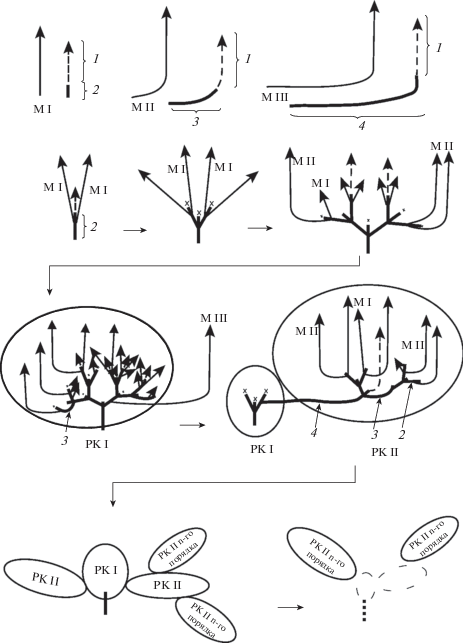
Таким образом, архитектурная структура особей N. consanguinea представляет собой совокупность трех модулей, их базитонное сочленение базальными частями приводит к формированию двух РК, один из которых – отсроченный РК II, можно рассматривать в качестве структурного элемента реитерации.
ЗАКЛЮЧЕНИЕ
Изучение особенностей развития особей эндемичного вида Памиро-Алая Nepeta consanguinea выявило высокую степень морфологической вариабельности, которая определяет устойчивое существование этого вида в сообществах Вахшского хребта. Онтогенез особей, независимо от условий обитания, сложный, полный. В конкретных местообитаниях происходит смена типа биоморфы с моноцентрической на неявнополицентрическую и образование различных морфологических структур. Типичная биоморфа N. consanguinea – удлиненнокаудексовая стержнекорневая формируется во всех изученных условиях (ЦЛ 1–4), ее перестройка на корневищно-каудексовую связана с пробуждением спящей почки и удлинением геофильных частей растущих побегов, возникших из почек регулярного возобновления. Образование парциальных кустов и смешанной корневой системы приводит к формированию куртины (ЦЛ 3, 4). В зависимости от условий обитания куртина в подземной сфере может состоять из различных элементов: удлиненных (ЦП 3) и (или) укороченных, гипо-, (и) или гипоэпигеогенного (ЦЛ 4) происхождения. Ускоренные темпы развития в прегенеративном периоде приводят к быстрому переходу растений к цветению, что положительно сказывается на пополнении и ускоренном обороте поколений в ценопопуляционных локусах. В антропогенных сообществах растянутые темпы развития в генеративном периоде, связанные с образованием куртины и клона, можно оценить как способ приспособления к существованию особей в данных условиях. Поливариантность онтогенеза и пластичность биоморфы оказываются одними из важнейших механизмов адаптации N. consanguinea к различным экологическим условиям Центральной Азии. Архитектурный анализ показал, что иерархическая организация особи состоит из трех структурно отличимых модулей, развитие которых в онтогенезе приводит к формированию двух разветвленных комплексов. Отсроченный архитектурный комплекс можно рассматривать как элемент реитерации. Детальное изучение близкородственных таксонов из sect. Macronepeta в будущем позволит сопоставить данные по биоморфологии этой группы и установить степень диверсификации биоморф в эколого-ценотическом ряду.
Список литературы
[Astashenkov, Cheryomushkina] Асташенков А.Ю., Черемушкина В.А. 2016. Архитектурная модель Nepeta mariae (Lamiaceae). – Растительный мир Азиатской России. 4 (24): 22–29.
Astashenkov A.Yu., Cheryomushkina V.A., Grebenjuk A.V., Dzumanov S.D. 2017. Transformation of life forms and ontogenetic structure of Nepeta pulchella Pojark. coenopopulations in Acsy-Zhabaglinsky nature reserve. – Contemp. Prob. Ecol. 10 (6): 758–771.
Astashenkov A.Yu., Cheryomushkina V.A., Myrzagalieva A.B., Medeubaeva B. 2018. Ontogenesis, estimation of coenopopulation state and component composition in Nepeta densiflora individuals of East – Int. J. Environ. Stud. 6: 1–4.
Barthélémy D., Caraglio Y. 2007. Plant architecture: a dynamic, multilevel and comprehensive approach to plant form, structure and ontogeny. – An. Bot. 99 (3): 375–407.
Bruy D., Hattermann T., Barrabe L., Mouly A., Barthelemy D., Isnard S. 2018. Evolution of plant architecture, functional diversification and divergent evolution in the genus Atractocarpus (Rubiaceae) for New Caledonia. – Front. Plant Sci. 9: 1–17.
[Budantzev] Буданцев А.Л. 1993. Триба Nepeteae Benth.семейства Lamiaceae Lindl. (систематика, география, возможности использования). Автореф. дис. … док. биол. наук. СПб. 33 с.
Charles-Dominique T., Edelin C., Bouchard A. 2010. Architectural strategies of Cornus sericea, a native but invasiveshrus of southern Quebec, Canada, under an open or a closed canopy. – Annals of Botany. 105: 205–220.
[Cheryomushkina] Черемушкина В.А. 2004. Биология луков Евразии. Новосибирск. 280 с.
[Dorokhina] Дорохина Л.Н. 1978. О жизненных формах полыней подрода Dracunculus (Bess.) Rydb. и переходе от трав к полукустарникам. – Бюлл. МОИП. Отд. биол. 83 (4): 97–108.
[Gatsuk] Гатцук Л.Е. 1974. Геммаксилярные растения и система соподчиненных единиц их побегового тела. – Бюлл. МОИП. Отд. биол. 1 (79): 100–113.
Halle F., Oldeman R.A.A. 1970. Essai sur l’architecture et la dynamique de croissance des arbres tropicaux. Paris. 178 p.
Hallé F., Oldeman R.A.A., Tomlinson P.B. 1978. Tropical trees and forests. Berlin, Heidelberg, New York. 441 p.
Keller R. 1997. Les divers modes de ramification de Potamogeton pectinatus L. (Potamogetonaceae) dans le Léman. – Adansonia. Ser. 3. 19 (1): 117–134.
[Kolegova, Cheryomushkina] Колегова Е.Б., Черемушкина В.А. 2012. Структура побеговых систем видов рода Thymus (Lamiaceae) в Хакасии. – Бот. журн. 97 (2): 173–183.
[Mazurenko, Khokhryakov] Мазуренко М.Т., Хохряков А.П. 1977. Структура и морфогенез кустарников. М. 158 с.
Notov A.A., Kusnetzova T.V. 2004. Architectural units, axiality and their taxonomic implications in Alchemillinae. – Wilfenia. 11: 85–130.
[Rabotnov] Работнов Т.А. 1950. Жизненный цикл многолетних травянистых растений в луговых ценозах. – Труды БИН АН СССР. Геоботаника. 3 (6): 176–196.
[Pojarkova] Пояркова А.И. 1954. Семейство Lamiaceae. – В кн.: Флора СССР. Т. 20. М., Л. С. 286–360.
Rua G.H. 2003. Growth forms, branching patterns, and inflorescence structure in Digitaria sect. Trichachne (Poaceae, Paniceae). – Flora. 198: 178–187.
[Savinykh] Савиных Н.П. 2006. Род Вероника: морфология и эволюция жизненных форм. Киров. 324 с.
[Serebryakov] Серебряков И.Г. 1962. Экологическая морфология растений. М. 378 с.
[Serebryakova] Серебрякова Т.И. 1977. Об основных “архитектурных моделях” травянистых многолетников и модусах их преобразования. – Бюлл. МОИП. Отд. биол. 5 (82): 112–128.
[Smirnova] Смирнова О.В. 1987. Структура травяного покрова широколиственных лесов. М. 205 с.
Tortosa R.D., Aagesen L., Tourn G.M. 1996. Morphological stadies in the tribe Colletieae (Rhamnaceae): analysis of architecture and inflorescences. – Bot. J. Linn. S. 122:3 53–367.
Troll W. Die Infloreszenzen. 1964. Bd. 1. Jena 615 p.
[Tsukervanik] Цукерваник Т.И. 1987. Род Nepeta. – В кн.: Определитель растений Средней Азии. Т. 9. С. 44–58.
[Uranov] Уранов А.А. 1975. Возрастной спектр фитоценопопуляций как функция времени и энергетических волновых процессов. – Биол. науки. 2: 7–34.
[Zhukova] Жукова Л.А. 1995. Популяционная жизнь луговых растений. Йошкар-Ола. 224 с.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал