

Ботанический журнал, 2019, T. 104, № 8, стр. 1228-1248
СОВРЕМЕННЫЕ СПОРОВО-ПЫЛЬЦЕВЫЕ СПЕКТРЫ ЕВРОПЕЙСКОЙ РОССИИ: 10 ЛЕТ НАБЛЮДЕНИЙ
М. Б. Носова 1, *, Е. Э. Северова 2, **, ***, О. В. Волкова 2
1 Главный ботанический сад им. Н.В. Цицина РАН
127276 г. Москва, ул. Ботаническая, 4, Россия
2 Московский государственный университет им. М.В. Ломоносова, Биологический факультет
119234 г. Москва, Ленинские горы, д. 1, стр. 12, Россия
* E-mail: mashanosova@mail.ru
** E-mail: elena.severova@mail.ru
*** E-mail: centaurea57@yandex.ru
Поступила в редакцию 02.06.2019
После доработки 25.06.2019
Принята к публикации 02.07.2019
Аннотация
Современные палинологические спектры были исследованы с применением ловушек Таубера в рамках Программы мониторинга пыльцы (Pollen Monitoring Programme – PMP) в шести модельных регионах средней полосы европейской части России в период 2007–2017 гг. Наблюдения проводились в Псковской, Тверской, Московской и Тульской областях, в трех растительных зонах: хвойно-широколиственных лесов, широколиственных лесов и в лесостепном экотоне. По результатам исследований предложены значимые уровни процентного содержания и скоростей аккумуляции пыльцы для основных средообразующих таксонов умеренной зоны. Определены специфические сочетания палинологических типов, определяющие зональную принадлежность, а также локальные особенности палинологических спектров и окружающей растительности.
При палеофитоценологических реконструкциях перед палинологами встает проблема соответствия количественных показателей ископаемых спектров (процентного содержания или скорости аккумуляции пыльцы) и ископаемой растительности.
При интерпретации ископаемых спектров появляется ряд закономерных вопросов, от возможности ответа на которые зависит качество реконструкции динамики растительности и, шире, ландшафта:
– Как влияет удаленность источника пыльцы на состав спорово-пыльцевого спектра?
– Каков вклад региональных и локальных источников пыльцы?
– Высокое участие пыльцевого типа в спектре соответствует высокому участию таксона в составе растительности или отражает его более высокую пыльцевую продукцию?
– Какой временной промежуток отражают поверхностные образцы мха? Почвы?
– Как влияет характер воспринимающей поверхности на состав спектра?
– Каков минимальный уровень содержания пыльцы (значимый уровень) того или иного таксона в образце, чтобы можно было однозначно подтвердить присутствие таксона in situ?
Эти вопросы были предметом внимания как за рубежом, так и в СССР, а позже в России (Федорова, 1952; Andersen, 1970; Birks, 1973; Bradshaw, 1981; Язвенко, 1992; Hicks, Birks, 1996; Филимонова, 2005). Естественным этапом развития палинологии в этом направлении был переход к выявлению количественных закономерностей формирования спорово-пыльцевых спектров (СПС). На первом этапе определяли только процентное содержание пыльцы в спектрах. Этот метод используется до сих пор, но имеет существенный недостаток – взаимозависимость величин процентного содержания друг с другом. Например, при высоком участии в составе растительности сосны, обладающей высокой пыльцевой продуктивностью, ее пыльца в процентном выражении преобладает, снижая процентные доли прочих таксонов и искажая реконструируемую картину их соотношения в составе растительности.
По мере разработки подходов к исследованию современных спектров происходило совершенствование методики наблюдений и отбора образцов. Использовались, помимо поверхностных спектров (почва, моховые подушки, которые накапливают пыльцу в течение нескольких, обычно 2–5 лет), также современные годичные спектры, получаемые с использованием ловушек Таубера. Широкое использование ловушек Таубера в исследованиях современного пыльцевого дождя стартовало в рамках Программы мониторинга пыльцы (Pollen Monitoring Programme – PMP, Hicks et al., 1996). Ловушка Таубера представляет собой емкость объемом 2 литра с отверстием определенного (5 см) диаметра. Их использование дает возможность оценить абсолютные характеристики пыльцевого дождя, такие как приток (influx) или скорость аккумуляции пыльцы (pollen accumulation rate – PAR), позволяющие, в отличие от процентного содержания, сохранить независимость компонентов в ряду данных. При определении этих показателей используется метод расчета числа пыльцевых зерен, попадающих на единицу площади улавливающей поверхности за один год (Seppä, Hicks, 2006), являющийся модификацией метода расчета концентрации пыльцы в ископаемых отложениях (Stockmarr, 1971).
В идеальном случае, для каждого географического региона и для каждого пыльцевого типа необходимо определение того значимого уровня (процентного содержания или скорости аккумуляции) пыльцы в спектре, который более или менее однозначно говорит о присутствии данного таксона в составе локальной, либо региональной растительности. Некоторые авторы в качестве маркера присутствия ископаемого таксона используют эмпирическую границу – уровень на пыльцевой диаграмме, выше которого кривая содержания пыльцы данного типа в спорово-пыльцевом спектре становится непрерывной (Vuorela, 1986). Однако для многих таксонов этот выбор системы отсчета не подходит из-за высокой пыльцевой продуктивности и значительной дальности переноса пыльцы. На территории европейской части России к ним относятся, например, Betula и Pinus. Напротив, таксоны с низкой пыльцевой продукцией могут быть не отражены в спектрах, несмотря на свое постоянное присутствие в составе растительности. Как правило, это энтомофильные таксоны с тяжелой пыльцой (например, Acer и, в меньшей степени, Tilia). Однако единичные пыльцевые зерна малопродуктивного таксона в образцах также сложно однозначно трактовать как признак его произрастания.
Основной современной работой, в которой обсуждаются значимые уровни для главных лесообразующих пород Европы, является работа О.В. Лисицыной с соавторами (Lisitsyna et al., 2011), одно из достоинств которой – использование, в том числе, данных с территории России (табл. 1). Однако полученные авторами этой работы значимые уровни требуют подтверждения на более обширном местном материале (пыльцевая продуктивность может различаться в разных частях ареала) и дополнения данными о скорости аккумуляции пыльцы.
Таблица 1.
Значимые уровни основных средообразующих таксонов центра Европейской России Table 1. Threshold values of main landcover-forming taxa from the Central European Russia
| Таксон Taxon |
Значимые уровни, % (литературные данные –
по Lisitsyna et al., 2011 и Kozharinov, 1994) Threshold values, % (published data cited after Lisitsyna et al., 2011, and Kozharinov, 1994) |
Значимые уровни, %,
предложенные О. В. Лисицыной
и соавт. (Lisitsyna et al., 2011) Threshold values, %, proposed by O. V. Lisitsyna et al. (2001) |
|
|---|---|---|---|
| для современной пыльцы modern pollen |
для ископаемой пыльцы fossil pollen |
||
| Picea | 1–5 | 2.5–5 | 1 |
| Pinus | 35–42 | 20–55 | 10 |
| Betula | 4.3–25 | 10–70 | 5 |
| Alnus | 0.5–2.5 | 0.5–8 | 2.5 |
| Corylus | 2–10 | pr*.–2 | 1 |
| Quercus | 1.5–10 | pr.–2 | 1.5 |
| Ulmus | 1 | pr.–2 | 0.5** |
| Tilia | pr. | pr.–5 | 0.5** |
| Acer | pr. | pr.–0.1 | 0.5** |
| Fraxinus | 2 | pr.–1 | 0.5** |
Примечание. * – pr. (presence) – присутствие единичных пыльцевых зерен. ** – возможен более низкий значимый уровень в том случае, когда в этом месте достигаются более высокие процентные значения для таксона.
Note. * – pr. (presence) – single pollen grains. ** – lower threshold value is possible in the case when the taxon has higher percentage in local vegetation communities.
Более ранняя работа А.В. Кожаринова (1994), посвященная динамике и миграциям основных лесообразующих пород Восточной Европы в позднеледниковье и голоцене, также рассматривает значимые уровни процентного содержания пыльцевых типов и является на сегодняшний день наиболее масштабным исследованием этой проблемы для территории европейской части России. Однако миграции таксонов в течение голоцена обсуждаются в ней без учета их составного характера (пыльцевой таксон Alnus включает пыльцу двух видов ольхи, которые, очевидно, мигрировали отдельно, аналогично для Ulmus). Кроме того, в работе отсутствуют данные об абсолютных показателях.
Последние исследования истории растительности голоцена в пределах лесной зоны европейской части России разных авторов (Novenko et al., 2009; Новенко, 2016; Nosova et al., 2019) показали, что зональной растительностью на пространстве от Балтийского моря до современного местоположения Москвы в течение последних 2000–3000 лет являлись различные варианты хвойно-широколиственных лесов с постепенным переходом на юге и западе к широколиственным лесам. В более ранний период, в течение термического оптимума голоцена и вплоть до субатлантического похолодания 2500 л.н., граница широколиственных лесов была смещена к северу и востоку. Субатлантическое похолодание усилило позиции ели и вновь раздвинуло границы хвойно-широколиственных лесов на юг и запад. Примерно к этому времени (Бронзовый век) относились первые значительные трансформации растительности человеком: происходил повсеместный переход к производящему хозяйству. Вплоть до славянской колонизации во второй половине I тыс. н.э. антропогенная нагрузка была невелика и преобладала зональная растительность – хвойно-широколиственные леса примерно в пределах современной соответствующей зоны (Nosova et al., 2019), широколиственные леса к северу и к югу от Оки и лесостепь – южнее (Novenko et al., 2012).
Таким образом, основными “индикаторными” зональными элементами палинологических диаграмм европейской части России являются ель и широколиственные породы, а также травянистые ветроопыляемые таксоны (Poaceae, Cyperaceae, Chenopodiaceae, Artemisia), формирующие зональные спектры лесостепных территорий.
Настоящее исследование посвящено определению возможных значимых уровней основных средообразующих таксонов центра Европейской России, как в относительном выражении (%), так и в абсолютном (скорость аккумуляции пыльцы, pollen accumulation rate, PAR) на основании результатов 10-летнего мониторинга современного пыльцевого дождя в рамках программы PMP.
ОБЪЕКТЫ И МЕТОДЫ
Исследования современных спектров в средней полосе европейской части России (ЕЧР) были начаты в 2007 г. Первые ловушки Таубера были установлены в Московской, Тверской и Псковской областях, затем (в 2009 г.) район исследования был расширен на юг, в окрестности Тулы и на Куликово Поле, а также добавлена еще одна точка мониторинга в Тверской области. Таким образом, в настоящее время получены палинологические данные за 8 и 10 лет для 23 ловушек Таубера, расположенных на шести ключевых территориях (рис. 1) в четырех локальных вариантах в зоне хвойно-широколиственных лесов (точки RU-PL, RU-TR, RU-CF и RU-ZV), в зоне широколиственных лесов (RU-TU) и лесостепном экотоне (RU-KP). По мере наблюдений некоторые ловушки были уничтожены естественными нарушениями (ветровал), другие – регулярно уничтожались людьми или животными. В Полистовском заповеднике пришлось сменить местоположение группы ловушек на более доступное. Краткие описания модельных территорий и локальных условий в месте расположения ловушек, а также периоды наблюдений и все вышеупомянутые особенности показаны в табл. 2.
Рис. 1.
Расположение шести ключевых участков мониторинга пыльцы в пределах европейской части России. 1 – RU-PL; 2 – RU-TR; 3 – RU-CF; 4 – RU-ZV; 5 – RU-TU; 6 – RU-KP.
Fig. 1. The locations of Russian PMP sites within Central European Russia. 1 – RU-PL; 2 – RU-TR; 3 – RU-CF; 4 – RU-ZV; 5 – RU-TU; 6 – RU-KP.
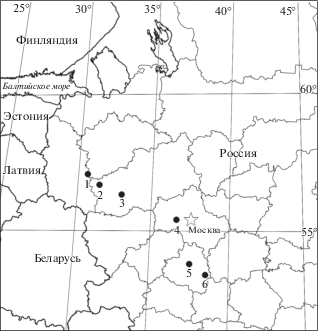
Таблица 2.
Расположение и краткая характеристика мест установки ловушек Таубера в пределах европейской части России Table 2. Location and short characteristics of the Tauber-trap sites in European Russia
| №, ID точки, местоположение Site number, ID and location |
Региональная растительность Regional vegetation |
Годы Years |
Широта
Долгота Latitude Longitude |
ID
ловушки Trap ID |
Локальная растительность Local vegetation |
|---|---|---|---|---|---|
| 1. RU-PL Псковская обл., заповедник “Полистовский” Pskov Region, Polistovsky Reserve |
Северная часть зоны хвойно-широколиственных лесов, Полистово-Ловатская болотная система, включающая различные варианты болот, в окружении вторичных смешанных и мелколиственных лесов/Nothern part of the coniferous-broadleaved forest zone, Polistovo-Lovatskaya Mire System with meso-oligotrophic mire complex. Small-leaved and mixed secondary forests with birch, aspen and alder prevail | 2008–2012 | 57°04' N 30°40' E |
RU-PL 1 | Окрайка олиготрофного болота/Oligotrophic bog edge |
| RU-PL 2 | Остров на болоте со смешанным лесом Mineral island with mixed forest | ||||
| RU-PL 3 | Мезотрофная окрайка болота/Mesotrophic bog edge | ||||
| RU-PL 4 | Открытое олиготрофное болото/Open raised bog | ||||
| 2013–2017 | 57°06' N 30°23' E |
RU-PL 5 | Мезотрофное болото/Mesotrophic bog | ||
| RU-PL 6 | Олиготрофное болото с сосной Oligotrophic bog with pine | ||||
| RU-PL 7 | Открытое олиготрофное болото/Open raised bog | ||||
| RU-PL 8 | Мезотрофное болото с черноольшаником по краю Mesotrophic bog with black alder on the edge |
||||
| 2. RU-TR Тверская обл., Биостанция “Чистый лес” Tver Region, Chisty Les Biology Station |
Зона хвойно-широколиственных лесов. Преимущественно вторичные смешанные леса с сосной, елью, березой и примесью широколиственных пород на песчаных почвах/Sand-soil type of coniferous-broadleaved forest zone. Secondary mixed forests with pine, spruce, birch and alder, and with admixture of broadleaved elements prevail | 2010–2017 | 56°44' N 31°31' E |
RU-TR 1 | Граница открытого и облесенного болота Open-forest edge on the oligotrophic bog |
| RU-TR 2 | Открытое олиготрофное болото/Open oligotrophic bog | ||||
| RU-TR 3* | Небольшое открытое олиготрофное болото Small open mesooligotropic bog |
||||
| 3. RU-CF Тверская обл., Центрально-Лесной заповедник Tver Region, Central Fores Nature Reserve |
Зона хвойно-широколиственных лесов, хвойные и смешанные леса южнотаежного облика/Southern taiga – coniferous-broadleaved forests transition. Seminatural spruce, mixed and secondary birch+aspen forests prevail | 2008–2017 | 56°28' N 33°02' E |
RU-CF 1 | Окно в смешанном лесу/Gap in mixed forest |
| RU-CF 2 | Сосняк по болоту/Bog pine forest | ||||
| RU-CF 3 | Открытое олиготрофное болото/Open raised bog | ||||
| RU-CF 4 | Открытая мезотрофная окрайка болота, граничащая с лугом Open bog edge bordered with meadow and pine forest |
||||
| 4. RU-ZV Московская обл., Звенигородская биологическая станция МГУ Moscow Region, Zvenigorod Biology Station |
Зона хвойно-широколиственных лесов. Густонаселенная пригородная территория. Вторичные еловые, сосново-еловые и елово-березовые леса 80–100-летнего возраста. Сложные ельники и широколиственные леса по речным террасам. В 2011–2012 гг. еловые насаждения нарушены типографом (Ips typographus)/Coniferous-broadleaved (mixed) forest zone. Densely populated suburban belt of Moscow with secondary spruce, pine-spruce and spruce-birch forests 80–100 years old. Terraces of the Moscow River are covered by coniferous-broadleaved forests. In 2011–2012 spruce forests were greatly damaged by bark beetle (Ips typographus) | 2008–2017 | 55°40' N 36°42' E |
RU-ZV 1 | Открытое олиготрофное болото/Open raised bog |
| RU-ZV 2 | Граница открытого и облесенного болота Open and pine-forest bog edge |
||||
| RU-ZV 3 | Смешанный лес/Mixed forest | ||||
| 2010–2017 | RU-ZV 4* | Еловый лес/Spruce forest | |||
| RU-ZV 5 | Вырубка с подростом сосны и березы Cutting with young pine-birch forest |
||||
| RU-ZV 6 | Березняк/Birch forest | ||||
| 5. RU-TU Тульская обл., Тульские засеки Tula Region, Tulskie Zaseki |
Зона широколиственных лесов. Современная растительность представлена фрагментами Засечной черты (широколиственные и мелколиственные леса) в окружении сельскохозяйственных земель и населенных пунктов./Broadleaved forest zone. Modern vegetation consists of fragmentary old broadleaved forests (remains of the forest belt which possessed defensive value in the Middle Ages), surrounded by agricultural lands and suburban areas. | 2010–2017 | 54°03' N 37°35' E |
RU-TU 1* | Карстовое мезотрофное болото в окружении сельскохозяйственных земель Karst mire surrounded with agricultural lands |
| RU-TU 2 | Мезоолиготрофное карстовое болото в окружении широколиственного леса/Mesooligotrophic bog surrounded by broadleaved forest | ||||
| RU-TU 3 | Эвтрофное карстовое болото в окружении широколиственного леса Eutrophic mire surrounded with broadleaved forest | ||||
| 6. RU-KP Тульская обл., музей-заповедник “Куликово поле” Tula Region, Kulikovo Field |
Северная лесостепь. Пахотные и залежные земли, остепненные луга, небольшие участки степной растительности и смешанных лесов./North forest-steppe. Arable and abandoned lands, steppified meadows, small areas of natural steppes and small patches of mixed forests. | 2010–2017 | 53°40' N 38°38' E |
RU-KP 1* | Открытый степной склон/Open steppe slope |
| RU-KP 2 | Пойменное эвтрофное болото в окружении залежей и полей Eutrophic mire surrounded with agricultural lands | ||||
| RU-KP 3 | Залежь/Abandoned field | ||||
| RU-KP 4 | Участок восстановления степной растительности Abandoned field under steppe restoration |
Методы работы в поле и лаборатории соответствуют стандарту Программы мониторинга пыльцы (Hicks et al., 1996; www.pollentrapping.net). Модифицированные ловушки Таубера были установлены в каждом из модельных регионов в количестве от 3 до 6 в различных местообитаниях (окно в лесу, граница леса и открытого фитоценоза, открытый фитоценоз) в октябре и заменялись новыми через год, после окончания сезона пыления. Лабораторная обработка содержимого ловушек включала в себя многократное центрифугирование для удаления избытков воды и химическую обработку ацетолизным или щелочным методом (Erdtman, 1960; Moore et al., 1991). В каждую ловушку перед началом лабораторной обработки добавляли таблетки Lycopodium (Stockmarr, 1971) для определения ежегодного притока пыльцы (п.з. см–2 год–1). Подсчет вели до 500 п.з. деревьев и кустарников в лесной зоне и до 600 п.з. в лесостепи, при этом количество подсчитанных спор Lycopodium должно быть не менее 20. Вычисления и построение диаграмм проводили с использованием программ TILIA и TGView (Grimm, 1991). Для определения скорости аккумуляции пыльцы (PAR) использовали формулу, предложенную S. Hicks с соавторами (1999).
РЕЗУЛЬТАТЫ
За время наблюдения в ловушках всего было зарегистрировано 86 пыльцевых типов, относящихся к 22 древесным, 59 недревесным и 5 криптогамным таксонам. Были отмечены все основные пыльцевые типы лесообразующих пород, характерных для Средней России. Также был отмечен дальний транспорт пыльцы, причем часто на значительные расстояния. Из нехарактерных для исследованных регионов древесных пород встречены Fagus (RU-KP), Carpinus (RU-TR, RU-CF, RU-ZV, RU-KP) и Juglans (RU-ZV), последние, возможно, из посадок в Московской области.
В связи с тем, что значения PAR и процентное содержание значительно различаются по годам, мы рассматривали медианные значения этих показателей, которые лучше, чем средние отражают характерные для данной точки величины притока пыльцы и ее процентного содержания. Максимальные и минимальные значения в случае с пыльцевыми данными часто имеют характер “выбросов” и несут в себе значительный элемент случайности, нарушая нормальное распределение.
Скорости аккумуляции и процентное содержание пыльцы (табл. 3) значительно изменяются от года к году и от ловушки к ловушке. Наиболее высокие медианные значения суммарной скорости аккумуляции пыльцы (PAR total), наблюдаются в южной части зоны хвойно-широколиственных лесов (RU-ZV) и в зоне широколиственных лесов (RU-TU), главным образом, за счет высоких PAR хвойных и мелколиственных древесных таксонов, южнее и севернее – при значительных (на порядки) колебаниях PAR, медианные значения ниже.
Таблица 3.
Медианы процентного содержания и скорости аккумуляции пыльцы для шести модельных территорий в пределах европейской части России Table 3. Median percentages and PAR values of the main landcover-forming taxa in the six model areas within European Russia
| RU-PL | RU-TR | RU-CF | RU-ZV | RU-TU | RU-KP | ||
|---|---|---|---|---|---|---|---|
| Tянв (Tjan), °С | –8.1 | –6.3 | –10.3 | –10.6 | –6.9* | –9.3 | |
| Tиюл (Tjul), °С | 17.2 | 17.8 | 16.1 | 17.2 | 19.1 | 23.1 | |
| Осадки, мм/Precipitation, mm | 685 | 760 | 740 | 650 | 550 | 504 | |
| Picea | % | 1.2 | 1.1 | 1.0 | 2.7 | 0 | 0 |
| PAR | 569 | 283 | 316 | 1406 | 0 | 0 | |
| Pinus | % | 5 | 4.6 | 5.8 | 8.8 | 0.8 | 1.9 |
| PAR | 1833 | 1379 | 1967 | 4725 | 474 | 692 | |
| Betula | % | 54.8 | 45.8 | 53 | 68.6 | 66.6 | 23.6 |
| PAR | 17 099 | 14 403 | 14 089 | 41 505 | 42 362 | 7446 | |
| Alnus | % | 15.4 | 23 | 10.6 | 2.1 | 3.9 | 2.9 |
| PAR | 6179 | 3641 | 1913 | 880 | 1942 | 1599 | |
| Quercus | % | 0 | 0.2 | 0 | 0.3 | 0.9 | 0.6 |
| PAR | 0 | 58 | 0 | 187 | 569 | 158 | |
| Tilia | % | 0 | 0 | 0 | 0.2 | 0.6 | 0.2 |
| PAR | 0 | 0 | 0 | 33 | 370 | 32 | |
| Ulmus | % | 0 | 0.2 | 0.2 | 0 | 0.3 | 0 |
| PAR | 0 | 20 | 37 | 0 | 253 | 0 | |
| Fraxinus | % | 0 | 0.2 | 0 | 0 | 1.1 | 0.2 |
| PAR | 0 | 36 | 0 | 0 | 711 | 26 | |
| Acer | % | 0 | 0 | 0 | 0 | 0.7 | 0 |
| PAR | 0 | 0 | 0 | 0 | 327 | 0 | |
| Corylus | % | 0.2 | 0 | 0.3 | 0.2 | 1 | 0 |
| PAR | 118 | 0 | 57 | 65 | 107 | 0 | |
| Cerealia | % | 0 | 0 | 0 | 0 | 0.1 | 0.9 |
| PAR | 0 | 0 | 0 | 0 | 45 | 441 | |
| Poaceae | % | 1.2 | 1.7 | 1.1 | 1.1 | 1.6 | 6.9 |
| PAR | 742 | 460 | 261 | 791 | 1174 | 1844 | |
| Cyperaceae | % | 2.4 | 0.7 | 0.7 | 0.2 | 0.3 | 0.4 |
| PAR | 837 | 115 | 106 | 88 | 196 | 181 | |
| Ericales | % | 0.6 | 0.3 | 0.3 | 0 | 0 | 0 |
| PAR | 156 | 125 | 62 | 0 | 0 | 0 | |
| Chenopodiaceae | % | 0 | 0 | 0.2 | 0.3 | 0.4 | 0.9 |
| PAR | 0 | 0 | 28 | 121 | 181 | 536 | |
| Artemisia | % | 0.6 | 0.2 | 0.7 | 0.6 | 0.9 | 3.7 |
| PAR | 209 | 79 | 130 | 302 | 632 | 1072 | |
| Пыльца трав/Grass pollen | % | 9.4 | 9.2 | 9.7 | 8 | 11.7 | 57.5 |
| PAR | 4319 | 2327 | 2328 | 4566 | 7446 | 23 401 | |
| PARtotal | 38 373 | 33 042 | 26 661 | 64 735 | 58 733 | 46 839 |
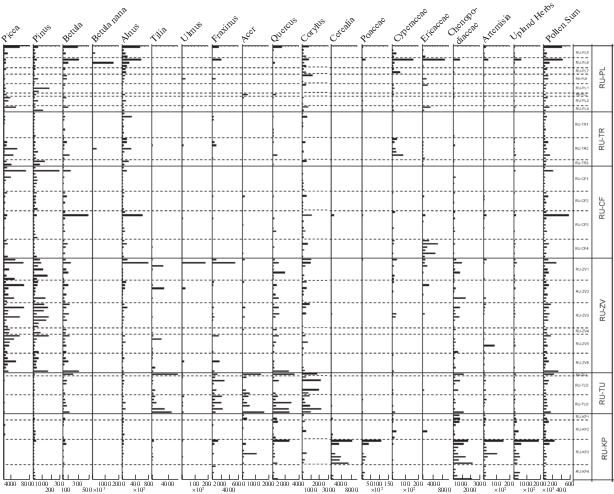
Picea. Количественная оценка содержания пыльцы ели в составе годичного пыльцевого дождя представляет особенный интерес, поскольку ель в средней полосе ЕЧР является одной из основных лесообразующих пород и эдификатором. В течение голоцена в пределах средней полосы европейской части России многие диаграммы показывают высокое (до 50–60%) содержание пыльцы Picea (Davydova et al., 2001; Носова, 2009), а скорость аккумуляции достигает 47 000 п.з. см–2 год–1 (Nosova et al., 2015). В настоящее время картина иная: процентное содержание пыльцы ели в ловушках Таубера колеблется в пределах от 0 до 14%, а PAR – от 0 до 6900 п.з. см–2 год–1 с максимальными значениями в малонарушенном участке в зоне хвойно-широколиственных лесов (RU-CF). Медианные скорости аккумуляции пыльцы сосны и ели высоки в хвойно-лесной зоне и снижаются в зоне широколиственных лесов и в лесостепи. Величины PAR, которые наблюдаются южнее Оки, обеспечиваются, вероятно, в значительной степени дальним и региональным (из более северных хвойно-лесных районов) транспортом пыльцы.
Медианное процентное содержание пыльцы ели в зоне хвойно-широколиственных лесов – от 1 до 2.7% (последнее значение – в наиболее южной точке RU-ZV).
После засухи 2010 года ельники в Подмосковье серьезно пострадали в результате массового размножения короеда-типографа (Ips typographus), при этом (рис. 3) скорости аккумуляции пыльцы ели уменьшились в первые два года после начала усыхания, а в 2015–2017 гг. – напротив, увеличились.
Рис. 2.
Процентное содержание пыльцы основных средообразующих таксонов в период 2008–2017 гг.
Fig. 2. Pollen percentages of main landcover-forming taxa for the period 2008–2017. Incomplete data rows are explained in the Materials and Methods section.
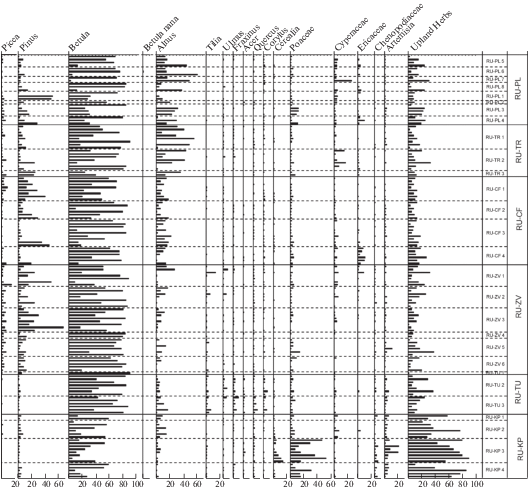
Рис. 3.
Скорости аккумуляции пыльцы основных средообразующих таксонов в период 2008–2017 гг.
Fig. 3. Annual PAR values of main landcover-forming taxa for the period 2008–2017. Incomplete data rows are explained in the Materials and Methods section.
Pinus. В четырех точках мониторинга (RU-ZV, RU-CF, RU-TR, RU-PL) отмечены сосновые древостои (болотные, либо суходольные). В Тульской области сосновые леса поблизости от точек исследования (RU-TU, RU-KP) отсутствуют, сосна единично присутствует в составе смешанных лесов. Невысокие значения PAR (med = 470 и 690 п.з. см–2 год–1 соответственно), характерные для этих территорий, маркируют подобную ситуацию: пыльца сосны здесь представляет собой региональный компонент пыльцевого дождя, а также частично может быть связана с дальним транспортом. Сосна встречается в составе растительности всех исследованных регионов, поэтому определить значимые уровни для нее не представляется возможным. В зоне широколиственных лесов и лесостепи, где она не определяет облик зональной растительности, ее медианные скорости аккумуляции снижаются с тысяч до сотен п.з. см–2 год–1. Следует отметить также, что в широколиственных лесах, где ловушки Таубера стоят в относительно закрытых фитоценозах, количественные показатели для пыльцы сосны ниже, чем в зоне лесостепи, где на состав пыльцевого спектра оказывает существенное влияние дальний транспорт.
Betula. Пыльца березы демонстрирует высокие показатели PAR в пределах всей лесной зоны, включая широколиственные леса. Максимальное значение PAR характерно для ловушки на открытом участке болота (RU-CF3, 297 000 п.з. см–2 год–1), однако медианные значения достаточно высоки во всех точках лесного пояса – десятки тысяч п.з. см–2 год–1. Значительное уменьшение PAR происходит в зоне лесостепи (медиана – 7500 п.з. см–2 год–1), хотя березовые леса здесь присутствуют по балкам и в посадках, а также возобновляются по залежам.
Наиболее высокие значения процентного содержания пыльцы березы (медиана) характерны для южной части зоны хвойно-широколиственных лесов (RU-ZV) – 68%, в широколиственных лесах (RU-TU) они немного уменьшаются, а южнее, в открытых ландшафтах лесостепи – снижаются до 23%.
Alnus. PAR ольхи достигает максимальных значений (60 000 п.з. см–2 год–1) в Центрально-Лесном заповеднике, в открытом фитоценозе. Высокие значения PAR типичны также и для других спорово-пыльцевых спектров из Полистовского и Центрально-Лесного заповедников (RU-PL и RU-CF). В остальных регионах значения PAR ольхи существенно ниже, высокие значения типичны лишь для спектров ловушек, расположенных в открытых местообитаниях, в лесу рядом с ольховой окрайкой болота (RU-ZV3, RU-TU3) и в близких к рекам местообитаниях в лесостепи (RU-KP1 и RU-KP2). Ольха – элемент азональной растительности, приуроченный к болотам и приречным древостоям, поэтому обсуждать ее значимые уровни в контексте зональной принадлежности спектра мы считаем нецелесообразным. Стоит отметить, однако, что наиболее низкие значения и процентного содержания, и PAR ольхи отмечены в точке с наиболее высокой пыльцевой продукцией для деревьев в целом (RU-ZV), где мягкие климатические условия юга зоны хвойно-широколиственных лесов способствуют высокой жизненности не только хвойных пород, но и широколиственных.
Скорости аккумуляции пыльцы широколиственных пород и орешника максимальны (от сотен до нескольких тысяч п.з. см–2 год–1) в зоне широколиственных лесов (RU-TU). Значительно меньшее, но постоянное участие этих пород наблюдается в южной части зоны хвойно-широколиственных лесов (RU-ZV) и, более спорадично, в лесостепи (RU-KP). В трех наиболее северных точках (RU-CF, RU-TR, RU-PL) лишь дуб и вяз (анемофильные таксоны) присутствуют регулярно и не единично. Несколько в меньших количествах встречается пыльца орешника, что, по-видимому, связано с его произрастанием под пологом леса. Минимальные значения PAR широколиственных пород могут служить как свидетельством произрастания таксона в пределах микрорегиона, так и отражать дальний транспорт пыльцы.
Tilia. Максимальное значение PAR отмечено в составе спектра ловушки RU-TU1– 5900 п.з. см–2 год–1, расположенной на границе зоны широколиственных лесов, в открытом ландшафте и вблизи (<300 м) от широколиственного леса. Медианное значение PAR составляет 370 п.з. см–2 год–1. Процентное содержание пыльцы липы в составе СПС не достигает 1% даже в зоне широколиственных лесов, в лесостепи (RU-KP) и на юге зоны хвойно-широколиственных лесов (RU-ZV) медианное процентное содержание составляет 0.2%. Вне зоны широколиственных лесов пыльца липы встречается единично, а медианные значения процентного содержания и PAR равны нулю.
Ulmus. В современных СПС пыльца вяза встречается регулярно и в заметных количествах во всех изученных зонах, максимальное значение PAR (18 000 п.з. см–2 год–1) было зафиксировано в точке RU-ZV1 (Московская область) на открытом участке. Однако, если исключить это выпадающее значение, то медианное значение PAR будет максимальным в зоне широколиственных лесов под Тулой (RU-TU). Во всех исследованных точках вяз является немногочисленным, но стабильным элементом растительности. Он занимает неудобные для земледелия, рубок и строительства участки, например, крутые склоны речных долин, а также низинные болота. Медианное процентное содержание и PAR колеблются около 0.2–0.3% и 20–250 п.з. см–2 год–1 соответственно (рис. 2 и рис. 3).
Fraxinus. Пыльца ясеня на всех точках регулярно встречается в небольших количествах, достигая максимального содержания в спектрах зоны широколиственных лесов и снижая свое участие в СПС лесостепи. PARmax составляет 5200 п.з. см–2 год–1 в точке RU-ZV (Московская область), самое высокое медианное значение PAR характерно для зоны широколиственных лесов. Медианное процентное содержание достигает 1.1% в зоне широколиственных лесов и не превышает 1% в других зонах.
Acer. Максимальная скорость аккумуляции пыльцы клена составляет 1600 п.з. см–2 год–1 (медиана 327 п.з. см–2 год–1) в широколиственных лесах. В других регионах пыльца клена в составе СПС встречается единично, а все медианные значения равны нулю.
Quercus. В зоне широколиственных лесов (RU-TU) значения PAR колеблются от десятков до тысяч п.з. см–2 год–1 (медиана – 569 п.з. см–2 год–1, максимум 3300 п.з. см–2 год–1). В значительных количествах, помимо широколиственных лесов, пыльца дуба отмечена в составе СПС южной части зоны хвойно-широколиственных лесов и лесостепи, достигая PARmax = 2400 п.з. см–2 год–1. В этих же зонах в ловушках RU-PL и RU-CF, расположенных на территориях обширных болотных массивов, пыльца дуба отсутствует.
Corylus. Скорость аккумуляции пыльцы орешника в хвойно-лесных районах не превышает 1000 п.з. см–2 год–1, медиана составляет 50–120 п.з. см–2 год–1. Максимальные значения медианы PAR отмечены в зоне широколиственных лесов и в Полистовском заповеднике (PAR max = 107 и 118 соответственно п.з. см–2 год–1). Медиана процентного содержания – 1% в зоне широколиственных лесов и 0.2–0.3% в прочих зонах за исключением лесостепи, где в составе СПС изредка встречаются лишь единичные зерна орешника.
Cerealia. Единственным регионом, в котором у культурных злаков значения PAR велики (PAR med = 440, PAR max = 8000 п.з. см–2 год–1), является Куликово Поле, где сельское хозяйство хорошо развито, и посевы занимают значительные площади. В зоне широколиственных лесов процентное содержание пыльцы культурных злаков в составе СПС достигает лишь 0.1%, а PAR med составляет 45 п.з. см–2 год–1. Во всех регионах зоны хвойно-широколиственных лесов медианные значения PAR для культурных злаков нулевые, а индивидуальные значения более высокие в точке RU-PL (Полистовский заповедник).
Cyperaceae. Скорость аккумуляции пыльцы и процентное содержание пыльцы осок достигает максимальных значений (PAR med = 837 п.з. см–2 год–1, 2.4%) в Полистовском заповеднике (RU-PL), где обширный болотный массив служит источником пыльцы этого таксона. Медианы обоих показателей тeм выше, чем большее число ловушек в пределах данной модельной территории находится в болотных фитоценозах. В точке RU-ZV лишь две ловушки из шести расположены на болоте, поэтому показатели для осок там самые низкие.
Ericales. Пыльца вересковых встречается в лесной зоне почти во всех образцах, увеличивая свое участие в составе СПС верховых болот. В лесных образцах (RU-PL3, RU-CF1-2, RU-ZV4-6) PAR вересковых минимальна (<200 п.з. см–2 год–1), либо пыльца их вовсе отсутствует. Медианы PAR трех южных модельных территорий (RU-ZV, RU-TU, RU-KP) нулевые, однако даже на Куликовом поле (RU-KP), где вересковые не растут, встречаются их единичные тетрады.
Chenopodiaceae. Пыльца маревых традиционно рассматривается как индикатор сельскохозяйственных угодий и нарушенных местообитаний. Высокие значения PAR маревых отмечены в Московской области (RU-ZV – медиана 121 п.з. см–2 год–1) и в широколиственных лесах, поблизости от Тулы (RU-TU – медиана 181 п.з. см–2 год–1). В лесостепи высокие значения PAR для маревых (PAR max 2200 п.з. см–2 год–1, медиана 536 п.з. см–2 год–1) указывает на высокую степень антропогенной нагрузки на территорию.
Artemisia. Максимальных значений PAR полыни (максимум – 18 000 п.з. см–2 год–1, медиана – 1072 п.з. см–2 год–1) достигает в точке RU-KP, наиболее южной, открытой и максимально нарушенной. Высокие значения PAR отмечаются также и в лесной зоне (RU-PL – 3500 п.з. см–2 год–1, RU-ZV – 8200 п.з. см–2 год–1).
Пыльца трав. PAR трав максимальна в лесостепи, отличаясь от остальных зон более чем на порядок. В лесном поясе скорость аккумуляции пыльцы травянистых растений выше на открытых участках.
Суммарная скорость аккумуляции пыльцы (PAR total). Суммарная скорость аккумуляции пыльцы максимальна в южной части зоны хвойно-широколиственных лесов (RU-ZV), где по-отдельности возрастают скорости аккумуляции пыльцы почти всех средообразующих таксонов. Минимальна PAR total в наиболее суровых условиях Центрально-Лесного заповедника (RU-CF) (табл. 3).
ОБСУЖДЕНИЕ
Растительные зоны и ландшафтные варианты в пределах зоны хвойно-широколиственных лесов можно диагностировать по сочетанию преобладающих в составе СПС пыльцевых типов. Межсезонные флуктуации как процентного содержания, так и скорости аккумуляции пыльцы значительны, однако медианные значения этих параметров хорошо характеризуют тип растительности и локальные условия. Исключение составляют случаи, когда пыльцевой тип в составе спектра встречается единично, но для определения зональной и подзональной принадлежности, как правило, такие случаи не важны.
Скорости аккумуляции пыльцы лучше, чем процентное содержание, отражают преобладающие тенденции (динамику участия таксона в составе растительности). Тем не менее, мы сочли важным показать процентную диаграмму (рис. 2) и медианы для процентных значений, поскольку традиционно палинологи интерпретируют именно процентные данные.
Болотный массив Полистовского заповедника, состоящий из разных вариантов болот, от эвтрофных черноольховых до олиготрофных грядово-мочажинных участков, и окруженный преимущественно вторичными лесами и залежами (RU-PL), формирует спектры с преобладанием пыльцы ольхи и вересковых, а также с неожиданно высоким в отсутствие сельскохозяйственной деятельности содержанием пыльцы растений-антропогенных индикаторов.
Вторая точка нашей “трансекты” – смешанные, сосновые и мелколиственные леса на легких почвах биостанции Чистый лес (RU-TR), которые характеризуются низкими значениями PAR древесных таксонов в целом и относительно высокими – Cyperaceae (на сплавине болота). Среди древесных таксонов стабильно высоко участие Pinus и Betula. Нестабильно, но чаще, чем в точках RU-PL и RU-CF, в спектрах RU-TR отмечается пыльца широколиственных таксонов, которые до сельскохозяйственного освоения территории входили в состав различных вариантов хвойно-широколиственных лесов, и в настоящее время восстанавливаются вновь в отсутствие нарушений.
В Центрально-Лесном заповеднике (RU-CF), представляющем собой малонарушенный массив хвойных лесов южнотаежного облика, значения PAR total минимальны. Особо следует отметить также чрезвычайно низкое процентное содержание и PAR Picea, а также высокие значения PAR болотных таксонов (Cyperaceae, Ericales).
Парадоксальное несоответствие процентного содержания и скоростей аккумуляции Picea в голоцене и в настоящее время заставляет задуматься о возможных причинах этого явления, которое невозможно объяснить какой-либо одной причиной. Наиболее очевидное объяснение – отсутствие современных аналогов для растительных сообществ, которые занимали эту территорию до начала трансформации ее человеком. Помимо смены условий произрастания (изменения климата и гидрологических условий, а также сведения климаксных еловых лесов человеком), этот феномен может быть объяснен также с позиций концепции генетической дифференциации ели в пределах ее ареала (Latałowa and Van der Knaap, 2006). Центр европейской части России является зоной гибридизации между Picea abies (L.) Karst. и P. obovata Ledeb. с образованием гибрида P. × fennica (Regel) Kom. Возможно, суровые климатические условия Центрально-Лесного заповедника неблагоприятны для произрастающей там популяции ели с ее генетическими особенностями.
Можно принять, что значимые уровни для ели на территории ЕЧР составляют <0.5–1% и несколько десятков п.з. см–2 год–1. Результаты подробного рассмотрения участия Picea в СПС и растительности, полученные в результате обработки данных мониторинга пыльцы, а также сведений из Российской палинологической базы данных (www.pollendata.org) представлены в ранее опубликованной нами статье (Nosova et al., 2015).
В южной части зоны хвойно-широколиственных лесов (RU-ZV) общий приток пыльцы значительно возрастает, причем он пропорционально увеличивается для всех лесообразующих пород кроме ольхи, а также для злаков и таксонов-антропогенных индикаторов. Формально принадлежа той же зоне, что и предыдущие точки, RU-ZV и визуально на диаграмме, и по медианным значениями процентного содержания и PAR существенно отличается от них. По-видимому, условия в этом районе наиболее благоприятные для произрастания и пыления бореальных таксонов. При этом роль широколиственных пород и Corylus также заметно возрастает.
В зоне широколиственных лесов ловушки расположены в пределах бывшей Засечной черты, хотя и нарушенной рубками, но сохраняющей в значительной степени зональный облик растительности. В окрестностях г. Тула (RU-TU) отмечены максимальные значения PAR широколиственных таксонов и Betula, а также Poaceae и таксонов-антропогенных индикаторов.
Спорово-пыльцевые спектры зоны лесостепи (территория музея-заповедника Куликово Поле – RU-KP) обладают наиболее яркими отличиями, что связано c особенностями открытого ландшафта и большой площадью сельскохозяйственных угодий. Для СПС лесостепи характерны максимальные значениями PAR и процентного содержания Poaceae, Cerealia, Artemisia, Chenopodiaceae и пыльцы травянистых растений в целом.
Скорости аккумуляции термофильных таксонов (Tilia, Fraxinus, Acer, Ulmus, Quercus и Corylus) зависят от теплообеспеченности территории. Помимо очевидного максимума содержания пыльцы термофильных пород в зоне широколиственных лесов и уменьшения их участия в лесостепи, в зоне хвойно-широколиственных лесов также прослеживаются закономерности, обусловленные влиянием климата (табл. 3). Направление нашей “трансекты” из точек с северо-запада на юго-восток, с одной стороны, соответствует широтному градиенту теплообеспеченности. С другой стороны, влияние Балтики на западе и более континентальные условия на востоке влияют на эту широтную закономерность: в точке RU-PL под воздействием более мягкого, переходного к морскому, климата увеличивается участие термофильных таксонов, а в точке RU-CF за счет специфических микроклиматических условий (табл. 3) – напротив, снижается.
Признаки развития сельского хозяйства на окружающих территориях, в частности, присутствие в составе СПС пыльцы хлебных злаков (Cerealia), требуют пристального внимания и изучения, поскольку здесь история растительности голоцена соприкасается с историей землепользования и антропогенного воздействия на растительность. Несмотря на то, что культурные злаки анемофильны, их PAR в лесной зоне не достигает больших значений (<1000 п.з. см–2 год–1) даже на открытых пространствах. Во всех пяти точках лесной зоны ближайшие поля отделены от мест расположения ловушек расстоянием и/или лесной растительностью, которая является естественным фильтром при распространении пыльцы (Vuorela, 1973). При этом более высокие значения в RU-PL отражают, по-видимому, с одной стороны, более развитое сельское хозяйство в этой части Псковской области (пыльца регионального происхождения), а с другой – близость к Прибалтике и Беларуси, где посевами заняты значительные территории (дальний транспорт пыльцы). Таким образом, в нашем исследовании скорость аккумуляции пыльцы хлебных злаков уверенно указывает на степень развития сельского хозяйства в регионе.
Помимо климата и антропогенного воздействия, на смены растительности влияют также инвазии вредителей. Яркий пример такого воздействия – вспышка короеда-типографа (Ips typographus). После повреждения лесов короедом-типографом в 2011–2014 гг., PAR Picea в Звенигороде (RU-ZV) сначала уменьшается, а затем постепенно увеличивается в течение 2014–2017 гг. Выпадение крупных старых экземпляров ели из 1 яруса древостоя сначала снижает пыльцевую продукцию, а затем осветление 2 яруса леса приводит к улучшению условий существования и увеличению пыльцевой продуктивности. Corylus, как правило, уже обитает под пологом ельника и дает “пыльцевой” ответ на осветление и улучшение условий уже в следующем году, поэтому PAR орешника возрастает в 2012 г. и остается, с некоторыми колебаниями, высоким и далее. Данные тенденции характерны для поврежденных короедом-типографом лесов точки RU-ZV, но не наблюдаются в почти не затронутых инвазией типографа лесах RU-CF.
ЗНАЧИМЫЕ УРОВНИ ОСНОВНЫХ СРЕДООБРАЗУЮЩИХ ТАКСОНОВ
На основании данных о динамике процентного содержания и скорости аккумуляции пыльцы за 10 лет мы предлагаем значимые уровни для основных средообразующих таксонов центра европейской части России (табл. 4). Следует отметить, что данные цифры означают не единичное присутствие таксона в составе растительности, а именно участие его в составе фитоценозов. Единичные пыльцевые зерна могут быть как продуктом единичных (например, культивируемых) или угнетенных особей, так и результатом дальнего заноса пыльцы, как в случае упомянутых в начале раздела “Результаты” Carpinus и Fаgus. При этом нулевой уровень процентного содержания и PAR может наблюдаться даже при наличии таксона в непосредственной близости от места отбора пробы. Например, посадки ели в музее-заповеднике “Куликово поле” не отражаются в составе СПС, по-видимому, из-за низкой пыльцевой продукции таксона за границей естественного ареала.
Таблица 4.
Предлагаемые нами для центра европейской части России значимые уровни пыльцы основных средообразующих таксонов. Table 4. The threshold values of main landcover-forming taxa offered for the Central European Russia
| Таксон/Taxon | % | PAR (п.з. см–2 год–1/p.g. cm–2 year–1) |
|---|---|---|
| Picea | 0.5 | 50 |
| Pinus | 5 | 500 |
| Betula | 3–5 | 2000 |
| Alnus | 0.5 | 300 |
| Corylus | pr.* | 50–100 |
| Quercus | pr. | pr. 50 |
| Ulmus | 0.2 | 30 |
| Tilia | pr. | pr. |
| Acer | pr. | pr. |
| Fraxinus | pr. | pr. |
| Cerealia | 0.1 | 50 |
| Poaceae | 0.5 | 50 |
| Cyperaceae | 0.5 | 50 |
| Ericales | pr. | 50 |
| Chenopodiaceae | 0.5 | 50 |
| Artemisia | 0.5 | 50 |
Значимые уровни скорости аккумуляции для европейской части России предложены нами впервые. Эти данные могут быть использованы при интерпретации результатов анализа скорости аккумуляции пыльцы голоценовых отложений.
По сравнению с данными О. Лисицыной с соавторами (Lisitsyna et al., 2011) значимые уровни процентного содержания Picea и Pinus, предложенные нами, ниже. Снижение значимых уровней пыльцы ели отражает данные из Центрально-Лесного заповедника RU-CF, где при высоком участии ели в насаждениях медианное содержание ее пыльцы составляет всего около 1%. Снижение значимых уровней пыльцы сосны связано с анализом ее встречаемости в СПС Полистовского заповедника (RU-PL), где сосна болотных форм занимает значительные площади, однако ее медианное содержание не превышает 5%. Очевидно, имеет значение не только площадь, занимаемая тем или иным таксоном, но и его жизненное состояние.
Обсуждать значимые уровни для Betula и Alnus сложно: для березы из-за чрезвычайно широкого распространения во всех точках наблюдения и отсутствия естественных границ обитания на нашей территории, а для ольхи – из-за приуроченности к азональным типам растительности. Тем не менее, мы предлагаем для березы за значимый уровень принять значение около 2000 п.з. см–2 год–1 и 3–5% (с учетом исследований других авторов (Lisitsyna et al., 2011) и выявленных в нашей работе медианных уровней скорости аккумуляции пыльцы и процентного содержания (7500 п.з. см–2 год–1, 23%).
Значимые уровни термофильных таксонов (Quercus, Ulmus, Tilia, Fraxinus, Alnus) и Corylus находятся в интервале от простого присутствия таксона до 0.5%. Однако надо принимать во внимание различие между произрастанием единичных особей и участием таксона в сложении растительности. Единичное присутствие пыльцевых зерен широколиственных пород в спектрах достаточно характерно для территорий, где они произрастают, но не формируют 1 ярус древостоя. В зоне широколиственных лесов, где термофильные таксоны входят в состав 1 яруса, медианное содержание достигает 0.5–1%. Таким образом, мы поддерживаем предложенное другими авторами (Lisitsyna et al., 2011) содержание 0.5% для всех пород кроме Quercus – для дуба мы изменяем значимый уровень до тех же 0.5% – в южной части хвойно-широколиственных лесов (RU-ZV) уже встречаются дубравы, однако медиана составляет всего 0.3%. Медианные скорости аккумуляции у энтомофильных широколиственных таксонов (Acer, Tilia) и у Fraxinus – нулевые или близки к ним в зоне хвойно-широколиственных лесов, а в зоне широколиственных лесов принимают значение до нескольких сотен. Таким образом, значимым уровнем для них является простое присутствие пыльцы в спектрах. Ветроопыляемые широколиственные (Quercus и Ulmus) демонстрируют ненулевые медианные значения на юге зоны хвойно-широколиственных лесов и их значимый уровень – от 30 до 50 п.з. см–2 год–1.
Значимые уровни PAR травянистых таксонов (все они, кроме Ericales – ветроопыляемые) на основании наших исследований составляют 50 п.з. см–2 год–1. Значимым уровнем процентного содержания Ericales можно считать единичное присутствие пыльцы.
Для травянистых ветроопыляемых таксонов (Chenopodiaceae, Artemisia, Cyperaceae) можно принять за значимый уровень 0.5%. Исключение составляет пыльца культурных злаков, содержание которых 0.1% уже может свидетельствовать о наличии сельскохозяйственных угодий на окружающей территории.
ЗАКЛЮЧЕНИЕ
1. Растительные зоны средней полосы европейской части России уверенно различимы на основании сочетания как относительных (процентных), так и абсолютных (скорость аккумуляции пыльцы – PAR) показателей пыльцевого дождя.
2. Основными диагностическими пыльцевыми таксонами, позволяющими определять зональную принадлежность спектра, являются Picea, широколиственные породы, Corylus, а также ветроопыляемые травянистые таксоны (Poaceae, Chenopodiaceae, Artemisia). Пыльца Ericaceae и Cyperaceae маркирует болотные ландшафты, олиготрофные и эвтрофные соответственно.
3. Пыльца Betula и Pinus большого диагностического значения на территории центра Европейской России не имеет из-за высокой пыльцевой продуктивности и летучести пыльцы, а также высокого участия этих таксонов в составе вторичной растительности. Оба эти фактора приводят к тому, что климатический градиент в пределах зоны хвойно-широколиственных лесов хотя и различим в палинологических диаграммах, но несколько “смазан” этими вторичными элементами спектров.
4. Для южной части зоны хвойно-широколиственных лесов характерны наибольшие абсолютные значения для всех бореальных элементов спектра кроме Alnus, которая может рассматриваться как азональный элемент спектров в силу специфики местообитаний.
5. Скорости аккумуляции пыльцы термофильных таксонов и ели зависят от климатических параметров. Увеличение их участия в составе растительности как следствие смягчения климата хорошо заметно даже в пределах одной растительной зоны хвойно-широколиственных лесов.
6. Для пыльцы основных средообразующих таксонов европейской части России предложены значимые уровни процентного содержания и скорости аккумуляции пыльцы, свидетельствующие об участии таксона в сложении растительности.
Список литературы
Andersen S.Th. 1970. The relative pollen productivity and pollen representation of north European trees, and correction factors for tree pollen spectra determined by surface pollen analysis from forest. – Dan. Geol. Unders. 2 (96): 1–99.
Birks H.J.B. 1973. Modern pollen studies in some arctic and alpine environments. – In: H.J.B. Birks and R.G. West (Editors) Quaternary Plant Ecology. Blackwell, Oxford. P. 143–168.
Bradshaw R.H.W. 1981. Modern pollen representation factors for woods in south-eastern England. – Journ. Ecol. 69: 45–70.
Davydova N.N., Subetto D.A., Khomutova V.I., Sapelko T.V. 2001. Late Pleistocene-Holocene paleolimnology of three north-western Russian lakes. – Journ. Paleolimn. 26 (1): 37–51.
Erdtman G. 1960. The acetolysis method. – Svensk. Bot. Tidskr. 54: 561–564.
[Fedorova] Федорова Р.В. 1952. Количественные закономерности распространения пыльцы древесных пород воздушным путем. – Труды ИГ АН СССР. 52: 91–103.
[Filimonova] Филимонова Л.В. 2005. Динамика растительности среднетаежной подзоны Карелии в позднеледниковье и голоцене (палеоэкологические аспекты). Петрозаводск. 200 с.
Grimm E.C. 1991. TILIA and Tilia graph: Springfield, U.S.A., Software available from Illinois State Museum.
Hicks S., Latałowa M., Ammann B., Pardoe H., Tinsley H. (Eds.). 1996. European Pollen Monitoring Programme – Project Description and Guidelines University of Oulu: 28 p.
Hicks S., Tinsley H., Pardoe H., Cundill P. 1999. European Pollen Monitoring Programme, supplement to the guidelines. Oulu. University Press, Oulu.
Hicks S., Birks H.J.B. 1996. Numerical analysis of modern and fossil pollen spectra as a tool for elucidating the nature of fine-scale human activities in boreal areas. – Veget. Hist. Archaeobot. 5 (4): 257ь272.
[Kozharinov] Кожаринов А.В. (1994). Динамика растительного покрова Восточной Европы в позднеледниковье-голоцене: Дисc. … докт. биол. наук. М. 255 с.
Latałowa M., van der Knaap W.O. 2006. Late Quaternary expansion of Norway spruce Picea abies (L.) Karst. in Europe according to pollen data. – Quat. Sci. Rev. 25 (21–22): 2780–2805.
Lisitsyna O.V., Giesecke T., Hicks S. 2011. Exploring pollen percentage threshold values as an indication for the regional presence of major European trees. – Rev. Palaeobot. Palynol., 166 (3–4): 311–324.
Moore P.D., Webb J.A., Collinson M.E. 1991. Pollen analysis. Oxford. 216 p.
[Nosova] Носова М.Б. 2009. Спорово-пыльцевые диаграммы голоценовых отложений как источник информации об антропогенном воздействии на растительность в доисторический период (на примере Центрально-Лесного заповедника). – Бюлл. МОИП. Сер. Биол. 114 (3): 30–36.
Nosova M.B., Novenko E.Y., Severova E.E., Volkova O.A. 2019. Vegetation and climate changes within and around the Polistovo-Lovatskaya mire system (Pskov Oblast, north-western Russia) during the past 10,500 years. – Veget. Hist. Archaeobot. 28 (2): 123–140.
Nosova M.B., Severova E.E., Volkova O.A., Kosenko J.V. 2015. Representation of Picea pollen in modern and surface samples from Central European Russia. – Veget. Hist. Archaeobot. 24 (2): 319–330.
[Novenko] Новенко Е.Ю. 2016. Изменения растительности и климата Центральной и Восточной Европы в позднем плейстоцене и голоцене в межледниковые и переходные этапы климатических макроциклов. М.: ГЕОС. 227 с.
Novenko E.Y., Volkova E.M., Glasko M.P., & Zuganova I.S. 2012. Palaeoecological evidence for the middle and late Holocene vegetation, climate and land use in the upper Don River basin (Russia). – Veget. Hist. Archaeobot. 21 (4–5): 337–352.
Novenko E.Y., Volkova E.M., Nosova N.B., Zuganova I.S. 2009. Late Glacial and Holocene landscape dynamics in the southern taiga zone of East European Plain according to pollen and macrofossil records from the Central Forest State Reserve (Valdai Hills, Russia). – Quat. Int. 207 (1–2): 93–103.
Seppä H., Hicks S. 2006. Integration of modern pollen and past pollen accumulation rate (PAR) records across the arctic tree line: a method for more precise vegetation reconstructions. – Quat. Sci. Rev. Vol. 25. P. 1501–1516.
Stockmarr J. 1971. Tablets with spores used in absolute pollen analysis. – Pollen Spores. 13: 615–621.
Vuorela I. 1973. Relative pollen rain around cultivated fields. – Acta Bot. Fenn. 102: 1–28.
Vuorela I. 1986. Palynological and historical evidence of slash-and-burn cultivation in South Finland. – In: Antropogenic indicators in pollen diagrams. (Behre K.-E. ed.). Rotterdam: Balkema. P. 53–64.
[Yasvenko] Язвенко С.Б. 1992. Современная пыльцевая продукция и голоценовая история горных лесов Закавказья. Дисс.… канд. биол. наук. М.: МГУ. 245 с.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал