

Ботанический журнал, 2019, T. 104, № 8, стр. 1167-1188
ДИНАМИКА ГЕОГРАФИЧЕСКОЙ СТРУКТУРЫ ЛИХЕНОБИОТЫ БЕЛАРУСИ КАК ИНДИКАТОР СОВРЕМЕННЫХ БИОКЛИМАТИЧЕСКИХ УСЛОВИЙ
1 Гомельский государственный университет им. Франциска Скорины
246019 г. Гомель, ул. Советская, 104, Беларусь
2 Самарский национальный исследовательский университет им. академика С.П. Королева
443086 г. Самара, Московское шоссе, 34, Россия
* E-mail: tsurykau@gmail.com
Поступила в редакцию 07.07.2019
После доработки 26.07.2019
Принята к публикации 30.07.2019
Аннотация
В статье представлены результаты изучения географической структуры лихенобиоты Республики Беларусь. Выделено 7 географических элементов: аридный, бореальный, гипоарктомонтанный, монтанный, мультизональный, неморальный и субокеанический. Установлена тенденция неморализации и аридизации лихенобиоты Беларуси, проявляющаяся в увеличении доли неморальных и аридных лишайников, а также в увеличении частоты встречаемости аридных видов. Данный тренд характерен для 2000–2019 гг. и, по-видимому, связан с изменением климатических характеристик изучаемой территории в данный период. Выявлено сокращение числа видов гипоарктомонтанного и монтанного географических элементов, являющихся уязвимыми лишайниками в спектре климатических условий последних десятилетий.
Анализ географической структуры лихенобиоты является неотъемлемой частью изучения разнообразия лишайников определенной территории (Biazrov, 2013). С момента заложения основ мико- и лихеногеографии в середине XX века результаты географического анализа использовали для реконструкции истории формирования биоты лишайников региона, установления механизмов и закономерностей ее становления (Oksner, 1940–42, 1944, 1948; Urbanavichus, 1999).
Результаты последних лихенофлористических и лихеногеографических исследований показали, что реакция лишайников на макроклиматические изменения, в частности на глобальное потепление, является более быстрой по сравнению с сосудистыми растениями (Lenoir et al., 2008; Chen et al., 2011), и данные лихеногеографии могут быть использованы для проверки и прогнозирования последствий глобального потепления (Braidwood, Ellis, 2012; Nascimbene et al., 2018).
Географический анализ лихенобиоты Беларуси до настоящего времени не проводился. Имеются лишь разрозненные данные о географической структуре лишайников отдельных природных (Kobzar, 1985; Golubkov, 1992) или административных регионов страны (Kobzar, 2004; Tsurykau, 2013), а также некоторых растительных формаций ее территории (Yatsyna, 2014; Bely, 2016).
В связи с вышеизложенным представляется актуальным выявление современной географической структуры лишайников Республики Беларусь, а также выявление тенденций ее изменения в спектре климатических условий последних десятилетий.
Настоящая работа основана на анализе опубликованных списков видов лишайников и лихенофильных грибов Беларуси (Tsurykau, 2017, 2018), составленных по результатам собственных полевых исследований 2003–2017 гг., проведенных ревизий отдельных систематических групп лишайников (роды Cetrelia, Hypotrachyna, Lepraria, Parmotrema, Punctelia, Xanthoparmelia, группа Cladonia chlorophaea-pyxidata) (Bely et al. 2014; Tsurykau, Golubkov, 2015; Tsurykau et al., 2015a, 2016, 2018a), а также анализе содержания 386 статей и материалов конференций, опубликованных за почти 240-летнюю историю изучения лишайников Беларуси (1781–2017).
Согласно Е.Э. Мучник (Muchnik, 2003), в настоящей работе термин “географический элемент” используется в классическом понимании как группа видов, имеющих более или менее общие черты распространения и совпадающие центры (или области) массовости в пределах крупных растительно-климатических зон и их высотных аналогов (поясов) в горах (Trass, 1970; Oksner, 1974 и др.). Названия видов приведены согласно списку лишайников Беларуси (Tsurykau, 2018).
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
В лихенобиоте Республики Беларусь выделено 7 географических элементов, из которых 3 (бореальный, неморальный и аридный) связаны с определенными природными зонами и 4 (гипоарктомонтанный, монтанный, субокеанический и мультизональный) имеют азональный, часто дизъюнктивный характер распространения.
Неморальный элемент включает виды лишайников, центры массовости которых связаны с зоной широколиственных лесов Голарктики, а также аналогичными местообитаниями других флористических царств (Trass, 1970; Golubkova, 1983). В лихенобиоте Беларуси этот элемент является ведущим, объединяя 205 (30.9%) видов лишайников (рис. 1).
Рис. 1.
Географическая структура лихенобиоты Беларуси.
Fig. 1. Geographical structure of the lichen biota of Belarus.
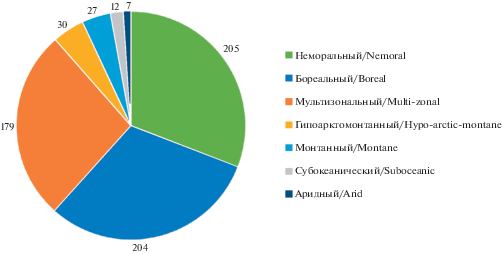
Согласно результатам исследований предыдущих лет, для географической структуры лихенобиоты различных регионов (Kobzar, 1985, 2004; Golubkov, 1992; Tsurykau, Khramchankova, 2007; Tsurykau, 2013) или растительных формаций (Bely, 2012a, 2016; Yatsyna, 2013a, 2014) Беларуси отмечалось преобладание бореального географического элемента, что является типичным для умеренной зоны Голарктики (Muchnik, 2003).
Изучение динамики представленности бореального и неморального географических элементов в лихенобиоте Беларуси за последние 50 лет (рис. 2) выявило, что увеличение доли видов лишайников неморального географического элемента произошло в течение второй декады XXI века (2011–2020 гг.).
Рис. 2.
Динамика представленности бореального и неморального географических элементов в лихенобиоте Беларуси за последние 50 лет.
Fig. 2. Dynamics of the shares of boreal and nemoral geographic elements in the lichen biota of Belarus over the past 50 years.
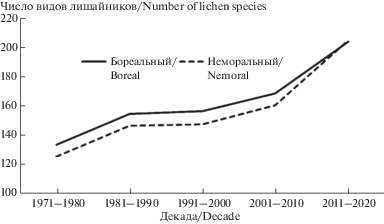
С конца 1980-х гг. на территории Республики Беларусь начался период потепления, не имеющий себе равных по продолжительности и интенсивности за весь период метеонаблюдений. В результате средняя годовая температура воздуха повысилась на 1.2°С (рис. 3), продолжительность периода со снежным покровом сократилась в среднем на 10–11 дней, увеличились продолжительность (10–12 дней) и теплообеспеченность (150–200°С) вегетационного периода (Padhornaya et al., 2015).
Рис. 3.
Карты распределения сумм активных температур на территории Беларуси до потепления климата (I) и в период его потепления (II) с учетом только сельских метеостанций (Khomich et al., 2018).
Fig. 3. Maps of distribution of growing degree days in Belarus before (I) and during (II) climate warming by only rural weather stations (Khomich et al., 2018).
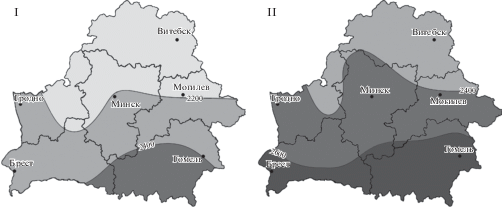
Известно, что для лишайников, как пойкилогидрических организмов, характерна тесная связь между их экофизиологией и температурой окружающей среды (Green et al., 2008). Например, исследования распределения лишайников в поясах гор показали, что некоторые бореальные виды (Bryoria fuscescens (Gyelnik) Brodo & D. Hawksw., Calicium viride Pers. и др.) достаточно быстро реагируют на повышение средних годовых температур, заселяя более высокие пояса гор и снижая свою численность в более низких (Bässler et al., 2015; Nascimbene, Spitale, 2017; Nascimbene et al., 2018). Однако, если в горах подобные сдвиги эпифитных лишайников могут быть затруднены из-за временного отсутствия необходимых видов деревьев-форофитов в более высоких поясах (Greenwood et al., 2016), на территории Беларуси изменение структуры лесов в пределах геоботанических подзон носит характер постепенного замещения (Yurkevich et al., 1979) и не препятствует распространению неморальных видов лишайников в северном направлении.
Полученные в последние годы данные о достаточно быстрой миграции многих видов лишайников вдоль зонального градиента позволяют несколько иначе взглянуть на некоторые положения, служившие основой при разработке теорий распространения лишайников в первой половине XX века. В частности, отмечалось, что лишайники обладают большей, чем у цветковых растений, ригидностью генотипической структуры и значительно лучше сохраняют свою первоначальную связь с растительно-климатическими зонами вследствие менее энергичного видообразования и, соответственно, не столь быстрой их реакции на условия окружающей среды (Oksner, 1948; Golubkova, 1983). Результаты современных молекулярных исследований указывают, что данное утверждение справедливо лишь для некоторых таксономических групп лишайников (Printzen et al., 2003; Palice, Printzen, 2004; Werth et al., 2007; Otálora et al., 2010). Скорость миграции других, в том числе многих широко распространенных видов, оказалась намного выше ожидаемой (Hewitt, 1999; Myllys et al., 2003; Muñoz et al., 2004; Buschbom, 2007). Высокая миграционная способность лишайников в сочетании с узкой экологической пластичностью многих видов (Mies, Lösch, 1995) открывает новые перспективы для биогеографической оценки региональных лихенобиот. По-видимому, современное сочетание видов лишайников в пределах определенной территории является следствием не только исторических связей, но и отражает спектр климатических условий последних десятилетий, т. е. может быть весьма динамичным.
Согласно прогнозным оценкам, к концу XXI века на территории Беларуси ожидается дальнейший рост среднегодовой температуры на 1.0–2.9°С (Bertosh et al., 2014), что в целом соответствует основным трендам климатических изменений в Европе (Gobiet et al., 2014). Вероятно, потепление климата окажет существенное влияние на лихенобиоту Беларуси, способствуя дальнейшей ее неморализации.
Бореальный элемент объединяет лишайники, центры массовости которых связаны с зоной хвойных лесов Голарктики, а также аналогичными местообитаниями других флористических царств. Мы не выделяем ното-бореальные виды в качестве отдельного географического элемента (Oksner, 1944; Muchnik, 2003) или субэлемента (Trass, 1970; Golubkova, 1983), поскольку данная классификационная единица, в отличие от остальных географических элементов флоры, выделяется не на зональной, а на региональной основе, и является скорее предметом ареалогического анализа лихенобиоты (включает мультирегиональные виды с голарктико-субантарктико-австралийским (Feuerer, Hawksworth, 2007), или биполярным распространением).
В лихенобиоте Беларуси бореальный элемент играет важную роль, лишь незначительно уступая неморальному элементу, и объединяет 204 (30.7%) вида лишайников. До начала 2010-х гг. бореальные виды занимали ведущее положение в географической структуре лихенобиоты (рис. 2), однако доля неморальных видов, найденных в Беларуси за последнее десятилетие, существенно превысила таковую бореальных лишайников. Важность данного факта возрастает в связи с тем, что основные лихенологические исследования конца 2000-х – первой половины 2010-х гг. проводились в сосновых (интразональных) (Yatsyna 2013a, 2014; Tsurykau, 2016) и еловых (бореальных) (Bely, 2011a, 2012a, b, 2016; Yatsyna, 2017) лесных формациях.
Потепление климата может негативно сказаться на частоте встречаемости многих бореальных видов лишайников. Несмотря на то, что некоторые из них (например, Hypogymnia tubulosa (Schaer.) Hav.) имеют отсроченную реакцию на потепление климата из-за их способности переносить достаточно широкий диапазон температур, другие являются крайне чувствительными к изменению климатических показателей и потому потенциально уязвимы в сценарии глобального потепления (Nascimbene et al., 2018). Например, целенаправленное изучение распространения бореального лишайника Cetraria pinastri (Scop.) Gray в Великобритании выявило существенное снижение его численности по сравнению с историческими данными, и, согласно прогнозным моделям, к 2050 году подходящие условия для его произрастания сократятся более чем на 75% (Binder, Ellis, 2008). С другой стороны, численность C. pinastri возросла в северной Европе. В частности, данные о встречаемости этого вида в Скандинавии продемонстрировали сдвиг его ареала на 151 км в северо-восточном направлении за период 1986–2003 гг. (Lättman et al., 2009).
Изменение погодно-климатических условий оказывает не только прямое влияние на бореальные лишайники, отрицательно сказываясь на их углеводном и водном обменах (Schroeter et al., 2000; Insarov, Schroeter, 2002; Wagner et al., 2012), но и способно косвенно снижать их численность и разнообразие, отражаясь на состоянии лесной растительности через изменение уровня грунтовых вод, пожары, размножение вредителей леса и усугубление развития болезней (Bertosh et al., 2014). Поскольку территория Беларуси расположена на стыке двух растительных зон – бореальной с господством хвойных и неморальной с преобладанием летне-зеленых лиственных лесов – изменение состава и структуры древесных насаждений может существенным образом отразиться на общем биоразнообразии. В частности, в современных погодно-климатических условиях еловые леса Беларуси испытывают негативное воздействие, сопровождающееся массовыми усыханиями и санитарными рубками (But’kovec, 2018). В результате, к настоящему времени граница сплошного распространения ели уже сместилась на 20–30 км севернее по сравнению с 1960-ми годами, и ожидается дальнейшая тенденция постепенного смещения границы сплошного ареала ели в северо-восточном направлении (Yurkevich, Geltman, 1962; Yermokhin, Puhacheusky, 2009; Bertosh et al., 2014). Это соответствует прогнозным оценкам изменения растительного покрова в результате климатических изменений, согласно которым Беларусь находится в зоне смены породного состава лесов (McCarthy et al., 2001).
Таким образом, многие бореальные лишайники Беларуси на протяжении последних нескольких десятилетий испытывают как непосредственное негативное воздействие повышенных среднегодовых температур атмосферного воздуха, так и находятся под угрозой изменения привычной среды обитания.
Гипоарктомонтанный элемент объединяет виды лишайников, центры массовости которых связаны с гипоарктической областью (Yurtsev, 1966), а также в результате значительной дизъюнкции широко представлены в хвойно-лесных поясах гор (Shustov, 2006). По данным Х.Х. Трасса (Трасс, 1970), многие виды этого элемента сравнительно часто произрастают в бореальных условиях, иногда внедряясь в зону смешанных и лиственных лесов и встречаясь на равнине не реже, чем в горных условиях.
В исследуемой лихенобиоте гипоарктомонтанный элемент представлен 30 видами (4.5%). Несмотря на то, что по числу видов этот элемент занимает четвертое место в географической структуре лихенобиоты Беларуси, уступая только неморальному, бореальному и мультизональному элементам, гипоарктомонтанные лишайники не принимают существенного участия в структуре лихенопокрова и в современных условиях, по-видимому, являются уязвимыми видами. В частности, 7 видов (23% числа видов данного элемента) известны лишь по историческим данным (до 1980 г., согласно (Lendemer et al., 2017)). К таким видам относятся Arctoparmelia centrifuga (L.) Hale, Megaspora verrucosa (Ach.) Hafellner & V. Wirth, Nephroma arcticum (L.) Torss., Peltigera leucophlebia (Nyl.) Gyeln., Rinodina conradii Körb., Schaereria fuscocinerea (Nyl.) Clauzade & Roux и Scytinium tenuissimum (Dickson) Otálora et al. Еще 8 видов (Buellia geophila (Flörke ex Sommerf.) Lynge, Cladonia cyanipes (Sommerf.) Nyl., C. macrophylla (Schaer.) Stenh., C. sulphurina (Michx.) Fr., Ochrolechia alboflavescens (Wulfen) Zahlbr., Rhizocarpon hochstetteri (Körb.) Vain., Tetramelas insignis (Nägeli ex Hepp) Kalb и Varicellaria lactea (L.) I. Schmitt & Lumbsch) были отмечены в 1980-х гг. Таким образом, 50% гипоарктомонтанных лишайников не подтверждены находками позднее 1990 г.
Следует отметить, что большинство видов данного элемента являются эпилитными лишайниками, произрастающими на ледниковых валунах кислых, реже основных горных пород. Количество старых неповрежденных человеком (в результате механического перемещения или дробления) эрратических валунов на территории Беларуси, подобно площади малоизмененных человеческой деятельностью первичных (коренных) лесов, значительно уменьшилось, что привело к сокращению доступных мест произрастания облигатных эпилитов (Golubkov, 1997). Негативное влияние антропогенной нагрузки на гипоарктомонтанные лишайники также отмечалось и для окрестностей Санкт-Петербурга (Malysheva, 2001).
Монтанный элемент объединяет виды лишайников, центры массовости которых связаны с лесными поясами гор Голарктики, а также других флористических царств (Makarevich, 1963; Shustov, 2006). Эти виды также могут встречаться в предгорьях и на равнинах. В Беларуси монтанный элемент по числу примерно равен гипоарктомонтанному географическому элементу и представлен 27 видами, что составляет 4.1% общего числа лишайников.
Следует отметить, что монтанные виды не достигают значительного развития на исследуемой территории. Большинство видов известны из единичных локалитетов, другие демонстрируют снижение численности в современных условиях. Так, четыре вида лишайников – Brianaria tuberculata (Sommerf.) S. Ekman & M. Svensson, Byssoloma subdiscordans (Nyl.) P. James, Rhizocarpon subpostumum (Nyl.) Arnold и Verrucaria floerkeana Dalla Torre & Sarnth. – вероятно, к настоящему времени уже исчезли на территории Беларуси, поскольку известны лишь по литературным данным начала XX века (Savicz, 1911; Savicz, Savicz, 1924; Bachmann, Bachmann, 1920). Последние находки других семи видов – Arthonia fuliginosa (Schaer.) Flot., Lecanora phaeostigma (Körb.) Almb., Lepraria neglecta (Nyl.) Erichsen, Micarea cinerea (Schaer.) Hedl., Pertusaria alpina Hepp ex Ahles, P. constricta Erichsen и P. coronata (Ach.) Th. Fr. – были сделаны в 1970–1980-х годах, и их современный статус требует уточнения (Tsurykau, 2018). Тем не менее, произрастание указанных монтанных лишайников на территории Беларуси вполне вероятно. Примером тому является Leptogium saturninum (Dicks.) Nyl., известный лишь по данным конца XIX – начала XX веков (Błoński, 1889; Kreyer, 1913; Bachmann, Bachmann, 1920; Savicz, Savicz, 1924; Savicz 1925), и найденный снова в середине 2010-х годов (Yatsyna, 2017).
Большинство монтанных лишайников, подтвержденных современными находками, являются крайне редкими и известны из одного (Rhizocarpon viridiatrum (Wulfen) Körb., Staurothele caesia (Arnold) Arnold и Thelidium minutulum Körb.), двух (Biatora ocelliformis (Nyl.) Arnold, Brianaria sylvicola (Flot. ex Körb.) S. Ekman & M. Svensson, Dimelaena oreina (Ach.) Norman, Lepraria rigidula (B. de Lesd.) Tønsberg и Staurothele drummondii (Tuck.) Tuck.) или трех (Biatora beckhausii (Körb.) Tuck.) локалитетов (Tsurykau, 2018). Несколько шире распространены Biatora vernalis (L.) Fr., Chaenotheca gracilenta (Ach.) J.-E. Mattsson & Middelb., C. laevigata Nádv., Menegazzia terebrata (Hoffm.) A. Massal., Peltigera horizontalis (Huds.) Baumg. и P. lepidophora (Nyl. ex Vain.) Bitter, однако характерной чертой их распространения в условиях Беларуси, как и многих других монтанных видов, является приуроченность к переходным (экотонным) биотопам, где в значительной мере ослаблены факторы фитоценотической конкуренции (Golubkov, 1986). Необходимость охраны таких видов была обоснована ранее (Golubkov, 1988, 1993b, 2002), и четыре из упомянутых видов (Chaenotheca gracilenta, Menegazzia terebrata, Peltigera horizontalis и P. lepidophora) включены в Красную книгу Республики Беларусь (Krasnaya… 2015).
Таким образом, участие монтанных лишайников в сложении лихенопокрова Беларуси невелико и, вероятно, ниже, чем предполагалось ранее (Gorbach, 1981). Монтанные виды не играют существенной роли в современных ценозах, являясь крайне редкими на всей территории республики. Являясь представителями азональной биоты, монтанные лишайники, тем не менее, крайне уязвимы в современных климатических условиях и нуждаются в специальных мерах охраны.
Субокеанический элемент объединяет лишайники, центры массовости которых связаны с океаническими и субокеаническими регионами материков (Golubkova, 1983; Shustov, 2006). В лихенобиоте Беларуси представлен 12 видами, что составляет 1.8% от общего числа видов лишайников. Низкая представленность данного элемента соответствует природным условиям региона. Согласно данным (Jäger, 1968; Horvat et al., 1974; Leuschner, Ellenberg, 2017), Беларусь располагается восточнее границы оптимума распространения субокеанических видов. Большая часть республики характеризуется условиями, благоприятствующими произрастанию субконтинентальных, в меньшей степени континентальных видов, но все еще обеспечивающими присутствие небольшого числа субокеанических представителей флоры (рис. 4).
Рис. 4.
Океанический градиент в Европе (на основе сосудистых растений): I – зона проникновения некоторых субокеанических видов за пределами восточной границы их распространения; составлено на основе данных Leuschner, Ellenberg (2017).
Fig. 4. Oceanic gradient in Europe (based on vascular plants): I – zone of presence of some suboceanic species east of their range limit; based on data by Leuschner, Ellenberg (2017).
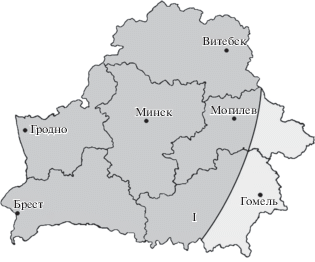
Сложно оценить уязвимость и современное состояние на территории Беларуси субокеанических видов лишайников в целом. Все лишайники субокеанического элемента являются редкими видами, некоторые из которых известны из одного (Lecania hutchinsiae (Nyl.) A.L. Sm., Ramalina subfarinacea (Nyl. ex Cromb.) Nyl., Usnea fragilescens Hav. и U. rubicunda Stirt.) (Insarov, Pchelkin, 1982; Yatsyna, 2013b) или трех–четырех локалитетов (Cladonia polydactyla (Flörke) Spreng., C. scabriuscula (Delise) Leight. и Pyrrhospora quernea (Dicks.) Körb.) (Tsurykau, 2018). Последнее упоминание Lobaria scrobiculata (Scop.) DC. для Беларуси датируется 1957 годом (Gorbach, 1957) и, вероятно, этот вид уже исчез на территории республики. Другие виды, такие как Cladonia incrassata Flörke, C. portentosa (Dufour) Coem. и C. tenuis (Flörke) Harm., спорадически встречаются в различных областях республики и подтверждены находками в течение последнего десятилетия (Tsurykau, 2018), а вид Cladonia norvegica Tønsberg & Holien был впервые указан в 2011 году (Bely, 2011b; Yatsyna, 2011) и с тех пор неоднократно приводился для различных регионов Беларуси.
Согласно Ellis (2015), чувствительность океанических и субокеанических видов лишайников к изменению климата до настоящего времени остается неясной. Результаты биоклиматического моделирования не позволили выявить каких-либо четких тенденций изменения встречаемости и распространения океанических видов в Великобритании (Ellis, Coppins, 2007; Ellis et al., 2007, 2009). Считается, что частота осадков является наиболее важным фактором, влияющим на распространение океанических и субокеанических видов лишайников. Поэтому благоприятное воздействие более высоких среднегодовых температур на океанические виды лишайников компенсируется негативным влиянием увеличения продолжительности засушливых периодов (Ellis, 2015).
В Беларуси за период потепления количество выпадающих осадков изменилось незначительно. За последние десятилетия на большей части территории республики отмечен небольшой рост годовых сумм осадков, более заметный в юго-восточных регионах (108% климатической нормы), в то время как на юго-западе страны количество выпадающих атмосферных осадков соответствует климатической норме (Padhornaya et al., 2015). По-видимому, современные тенденции в изменении основных климатических характеристик (температуры воздуха и осадков) пока не отразились существенным образом на распространении редких субокеанических видов лишайников в условиях Беларуси. Тем не менее, сложно предсказать, сохранятся ли в будущем сложившиеся гидрометеорологические условия в регионе, и как дальнейший тренд глобального потепления отразится на произрастающих за пределами своего климатического оптимума представителей субокеанической биоты.
Аридный элемент объединяет лишайники, центры массовости которых связаны с засушливыми (аридными) областями Голарктики, а также аналогичными местообитаниями других флористических царств (Lazarenko, 1956; Golubkova, 1983; Shustov, 2006).
В лихенобиоте Беларуси к аридному элементу относятся 7 видов лишайников (1.1% от общего числа видов) – Circinaria sphaerothallina (J. Steiner) Sohrabi, C. foliacea (Huds.) Willd., C. pocillum (Ach.) O.J. Rich., C. symphycarpa (Flörke) Fr., Variospora aurantia (Pers.) Arup, Søchting & Frödén, Verrucaria fusca Pers. и Xanthoparmelia pulla (Ach.) Crespo et al. Все эти виды являются редкими на территории Беларуси. В качестве примера можно привести Xanthoparmelia pulla, до недавнего времени считавшийся обычным видом, известным для всех областей республики. В результате проведенной ревизии большинство образцов, ранее приводимых под этим названием, оказались принадлежащими другому, морфологически схожему виду – X. delisei (Duby) O. Blanco et al. В Беларуси X. pulla имеет ограниченное распространение и является относительно редким, известным только на северо-западе республики видом (Tsurykau et al., 2018a, b). Такая приуроченность аридного лишайника к северной части Беларуси связана исключительно с доступностью валунного материала ледникового происхождения, отсутствующего в южных регионах страны (Golubkov, 1992, 1996; Tsurykau et al., 2018b).
Другим крайне редким видом лишайников, ранее приводимым из многих локалитетов республики (Golubkov, 1993a, 2013), является Cladonia pocillum. Большинство образцов нами были переопределены и отнесены к другим видам лишайников (преимущественно C. monomorpha Aptroot, Sipman & van Herk) в ходе проведенной ревизии лишайников группы Cladonia chlorophaea-pyxidata, и на данный момент C. pocillum известен только из одного локалитета в западной части страны (Tsurykau, Golubkov, 2015; Tsurykau et al., 2015b).
Несмотря на незначительное участие аридных видов в структуре лихеносообществ республики, эти лишайники, обладая выраженными зональными признаками распространения, могут служить индикаторными (сигнальными) видами в условиях изменяющегося климата. Следует отметить, что более половины аридных видов лишайников были приведены для территории Беларуси после 2000 г. (Golubkov, 2002; Yurchenko, 2011; Yatsyna, 2015; Tsurykau, Golubkov, 2015). На тот же период приходится 47% находок аридных лишайников за весь период изучения лихенобиоты Беларуси (рис. 5) (Tsurykau, Golubkov, 2015; Tsurykau et al., 2018b; Tsurykau, 2018).
Рис. 5.
Частота находок аридных видов лишайников на территории Беларуси (накопительный график).
Fig. 5. Frequency of the records of arid lichen species in Belarus (cumulative graph).
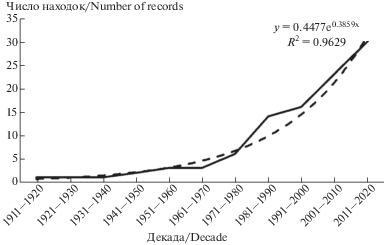
Увеличение разнообразия и встречаемости аридных видов лишайников соответствует общей динамике ксерофитизации флоры и фауны Беларуси. С конца ХХ века в республике были обнаружены растения и животные более южного происхождения (характерные для лесостепных и степных сообществ соседней Украины), которые ранее в стране не отмечались (Stsepanovich, 1997; Puhacheusky et al., 2011; Khomich et al., 2018). Расширение ареала степных видов растений и животных и их экспансия на территорию Беларуси являются результатом увеличения среднегодовых температур и появления гидротермических условий степной зоны (Khomich et al., 2018).
В условиях Беларуси аридные лишайники приурочены к хорошо освещаемым и прогреваемым локалитетам, заселяя каменистые субстраты силикатной или карбонатной природы, а также участки с легкими, преимущественно песчаными почвами.
Таким образом, выявленные особенности встречаемости ксеротермных лишайников соответствуют тенденциям аридизации климата Беларуси.
Мультизональный элемент объединяет лишайники, широко распространенные во многих растительно-климатических зонах Голарктики, а также других флористических царств (Golubkova, 1983). В лихенобиоте Беларуси мультизональный элемент играет важную роль, занимая третье место, и насчитывает 179 (27.0%) видов.
Выделение мультизонального элемента неоднократно подвергалось критике (Kondratyuk, 1988; Urbanavichus, 2001; Muchnik, 2003). Однако, ввиду различных причин (недостаточные сведения об экологии, таксономические проблемы, широкая экологическая пластичность), для многих видов лишайников затруднительно определить конкретные фитогеографические признаки, которые позволили бы однозначно выявить их центр массовости. Поэтому выделение мультизональных видов было предложено рассматривать как вынужденное, временное явление (Muchnik, 2003), пока новые данные лихеногеографии, экологии и молекулярной филогенетики не позволят с большой долей вероятности определить конкретный географический элемент для мультизональных видов лишайников.
Важную роль при определении центра массовости вида представляют данные о его ареале. 141 вид (79% от числа видов мультизонального элемента) произрастает в различных флористических царствах, т.е. характеризуется мультирегиональным типом ареала. Результаты молекулярных исследований показали генетическую неоднородность многих мультирегиональных видов. Ярким примером является Lepraria incana (L.) Ach., до недавнего времени считавшийся типичным космополитным лишайником, распространенным на всех континентах, исключая Арктические широты и Антарктиду (Saag et al., 2009). Однако таксономическая ревизия североамериканских образцов L. incana показала, что они генетически отличны от европейских и представляют 2 отдельных, ранее не описанных вида лишайников – L. pacifica Lendemer, распространенный на тихоокеанском побережье Северной Америки, и L. hodkinsoniana Lendemer, произрастающий на востоке Североамериканского континента (Lendemer, 2011). Позже L. incana s. str. была исключена из списка видов лишайников Южной Америки (образцы были переопределены как L. aff. hodkinsoniana) (Guzow-Krzemińska et al., 2019). Произрастание данного вида в Юго-Восточной Азии и Австралазии было также поставлено под сомнение после того, как видовая принадлежность образцов из Непала и Новой Зеландии, ранее опубликованных под названием Lepraria incana, была также переопределена (Kukwa, 2006). Таким образом, к настоящему времени ареал L. incana s. str. охватывает Европу и северную часть азиатского субконтинента. Такое значительное сужение географического распространения этого вида позволяет пересмотреть его зональную приуроченность. Однако, по всей видимости, даже европейский материал Lepraria incana является генетически неоднородным (Knudsen, персональное сообщение), что подтверждается его высокой морфологической вариабельностью (Saag et al., 2009), не характерной представителям данного рода.
Следует также отметить, что в Беларуси 45% мультизональных лишайников связаны с каменистым субстратом, и более половины (55%) узкоспециализированных эпилитов являются мультизональными видами (Tsurykau, 2018). Очевидно, имеющиеся данные об экологии эпилитных лишайников, а также неравномерное распространение каменистого субстрата в пределах умеренной зоны Голарктики не позволяют к настоящему времени отнести эти виды к определенному географическому элементу.
ЗАКЛЮЧЕНИЕ
В лихенобиоте Беларуси выделено 7 географических элементов. Наиболее представлены неморальный (205 видов лишайников), бореальный (204 вида) и мультизональный (178 видов). Вместе эти три элемента объединяют 88.3% лихенобиоты. Представители гипоарктомонтанного (30), монтанного (27), субокеанического (12) и аридного (7 видов) географических элементов незначительно представлены в структуре лихенобиоты и не играют существенной роли в современных ценозах, являясь преимущественно редкими видами лишайников на всей территории республики.
Установлена тенденция неморализации и аридизации лихенобиоты Беларуси, проявляющаяся в увеличении доли неморальных и аридных лишайников в ее структуре, а также в увеличении частоты встречаемости многих аридных видов. Данный тренд соответствует динамике изменения растительного и животного мира республики и связан, по-видимому, с изменением климатических характеристик изучаемой территории.
Выявлено сокращение числа видов гипоарктомонтанного и монтанного географических элементов, являющихся уязвимыми лишайниками в спектре климатических условий последних десятилетий. Сохранение этих лишайников может потребовать применения природоохранных мер.
Список литературы
Bachmann E., Bachmann F. 1920. Litauische Flechten. – Hedwigia. 61 (6): 308–342.
Bässler C., Cadotte M.W., Beudert B., Heibl C., Blaschke M., Bradtka J.H., Langbehn T., Werth S., Müller J. 2015. Contrasting patterns of lichen functional diversity and species richness across an elevation gradient. – Ecography. 39: 689–698. https://doi.org/10.1111/ecog.01789
[Bely] Белый П.Н. 2011a. Состав и особенности систематической структуры лишайников еловых лесов Беларуси. – Веснік МДПУ імя І.П. Шамякіна. 4: 9–17.
[Bely] Белый П.Н. 2011b. Аннотированный список лишайников и лихенофильных грибов еловых экосистем Беларуси. – С сб.: Особо охраняемые природные территории Беларуси. Исследования. Вып. 6. Минск. С. 146–178.
[Bely] Белый П.Н. 2012a. Географическая структура лихенофлоры еловых лесов Беларуси. – Веснік МДПУ імя І.П. Шамякіна. 1: 3–10.
[Bely] Белый П.Н. 2012b. Хорологические особенности еловых лесов Беларуси: широтное распределение лишайников. – Веснік МДПУ імя І.П. Шамякіна. 3: 3–10.
[Bely] Белый П.Н. 2016. Лишайники еловых лесов Беларуси. Минск. 230 с.
Bely P., Golubkov V., Tsurykau A., Sidorovich E. 2014. The lichen genus Cetrelia in Belarus: distribution, ecology and conservation – Bot. Lith. 20 (2): 69–76. https://doi.org/10.2478/botlit-2014-0010
[Bertosh et al.] Бертош Е., Русаков Д., Лукашевич Т. 2014. Национальный доклад: Уязвимость и адаптация к изменению климата в Беларуси. Минск. 45 с.
[Biazrov] Бязров Л.Г. 2013. Формулы ареалов лихенизированных грибов семейства Umbilicariaceae из Монголии на основе общемировой карты экорегионов суши. – Новости сист. низш. раст. 47: 179–199.
Binder M.D., Ellis C.J. 2008. Conservation of the rare British lichen Vulpicida pinastri: Changing climate, habitat loss and strategies for mitigation. – Lichenologist. 40: 63–79. https://doi.org/10.1017/S0024282908007275
Błoński F. 1888. Spis roślin skrytokwiatowych zebranych w r. 1887 w puszczy Białowieskiéj. – Pam. fizyjogr. 8: 75–119.
Braidwood D., Ellis C.J. 2012. Bioclimatic equilibrium for lichen distributions on disjunct continental landmasses. – Botany. 90: 1316–1325. https://doi.org/10.1139/b2012-103
Buschbom J. 2007. Migration between continents: geographical structure and long-distance gene flow in Porpidia flavicunda (lichen-forming Ascomycota). – Mol. Ecol. 16: 1835–1846. https://doi.org/10.1111/j.1365-294X.2007.03258.x
[But’kovec] Бутьковец В.В. 2018. Распределение Picea abies в лесных насаждениях южной части Беларуси. – В сб.: Проблемы лесоведения и лесоводства. 78. Гомель. С. 133–140.
Chen I.C., Hill J.K., Ohlemuller R., Roy D.B., Thomas C.D. 2011. Rapid range shifts of 469 species associated with high levels of climate warming. – Science. 333: 1024–1026. https://doi.org/10.1126/science.1206432
Ellis C. 2015. Biodiversity Climate change impacts report card technical paper. 8. Implications of climate change for UK bryophytes and lichens. – Biodiversity Report Card Paper. 8: 1–18.
Ellis C.J., Coppins B.J. 2007. Changing climate and historic-woodland structure interact to control species diversity of the ‘Lobarion’ epiphyte community in Scotland. – J. Veg. Sci. 18: 725–734. https://doi.org/10.1111/j.1654-1103.2007.tb02587.x
Ellis C.J., Coppins B.J., Dawson T.P., Seaward M.R.D. 2007. Response of British lichens to climate change scenarios: trends and uncertainties in the projected impact for contrasting biogeographic groups. – Biol. Conserv. 140: 217–235. https://doi.org/10.1016/j.biocon.2007.08.016
Ellis C.J., Yahr R., Coppins B.J. 2009. Local extent of old-growth woodland modifies epiphyte response to climate change. – J. Biogeogr. 36: 302–313. https://doi.org/10.1111/j.1365-2699.2008.01989.x
Feuerer T., Hawksworth D.L. 2007. Biodiversity of lichens, including a world-wide analysis of checklist data based on Takhtajan’s floristic regions. – Biodivers. Conserv. 16 (1): 85–98. https://doi.org/10.1007/s10531-006-9142-6
Gobiet A., Kotlarski S., Beniston M., Heinrich G., Rajczak J., Stoffel M. 2014. 21st century climate change in the European Alps – a review. – Sci. Total Environ. 493: 1138–1151. https://doi.org/10.1016/j.scitotenv.2013.07.050
[Golubkov] Голубков В.В. 1986. Эколого-географическая характеристика некоторых редких и реликтовых видов лишайников, произрастающих на охраняемых природных территориях Белорусской ССР. – Ботаника (Исследования). 27: 139–141.
[Golubkov] Голубков В.В. 1988. Охрана редких и исчезающих видов лишайников в Белоруссии. – Микологические исследования. 16: 125–128.
[Golubkov] Голубков В.В. 1992. Лишайники охраняемых природных территорий Беларуси (флористическая и эколого-географическая характеристика): Дис. … канд. биол. наук. Минск. 503 с.
Golubkov V.V. 1993a. About some new and rare lichen species for the territory of Belarus. – In: Abstracts of the 12th International Conference “Fungi and lichens in the Baltic region”. Vilnius. P. 137–138.
[Golubkov] Голубков В.В. 1993b. Вопросы охраны лишайников Белоруссии. – В сб.: Материалы VI съезда БРБО “Проблемы ботаники”. Ч. 1. Минск. С. 97–98.
[Golubkov] Голубков В.В. 1996. Влияние антропогенной трансформации ландшафтов на особенности распространения и разнообразия лишайников в Белорусском Поозерье. – В сб.: Тез. докл. регион. науч.-практ. конф. “Сохранение биологического разнообразия Белорусского Поозерья”. Витебск. С. 81–82.
[Golubkov] Голубков В.В. 1997. Лишайники как компоненты ландшафтов и геологических структур некоторых проектируемых и существующих памятников природы Беларуси. – В сб.: Матэрыялы канф. “Актуальныя праблемы прыродазнаўства”. Мiнск. С. 63–65.
[Golubkov] Голубков В.В. 2002. Созологический анализ лихенобиоты бассейна р. Днепр (Беларусь). – В сб.: Материалы респ. науч. конф. “Красная книга Республики Беларусь: состояние, проблемы, перспективы”. Витебск. С. 69–71.
[Golubkov] Голубков В.В. 2013. Новые, редкие и малоизвестные таксоны лишайников, обнаруженные на территории Беларуси (краткое сообщение). – В сб.: Материалы IX междунар. конф. “Актуальные проблемы экологии”. Гродно. С. 22–24.
[Golubkova] Голубкова Н.С. 1983. Анализ флоры лишайников Монголии. Ленинград. 281 с.
[Gorbach] Горбач Н.В. 1957. Материалы к флоре лишайников Белоруссии (Лишайники Беловежской пущи). – Бюл. Ин-та биологии за 1956 год. 2: 43–46.
[Gorbach] Горбач Н.В. 1981. Лишайники монтанного географического элемента в лихенофлоре Белоруссии. – В сб.: Брио-лихенологические исследования высокогорных районов и севера СССР. Апатиты. С. 86–87.
Green T.G.A., Nash III T.H., Lange O.L. 2008. Physiological ecology of carbon dioxide exchange. – In: Lichen Biology. Cambridge. P. 152–181. https://doi.org/10.1017/CBO9780511790478.010
Greenwood S., Chen J.C., Chen C.T., Jump A.S. 2016. Community change and species richness reductions in rapidly advancing tree lines. – J. Biogeogr. 43: 2274–2284. https://doi.org/10.1111/jbi.12776
Guzow-Krzemińska B., Jabłońska A., Flakus A., Rodriguez-Flakus P., Kosecka M., Kukwa M. 2019. Phylogenetic placement of Lepraria cryptovouauxii sp. nov. (Lecanorales, Lecanoromycetes, Ascomycota) with notes on other Lepraria species from South America. – MycoKeys. 53: 1–22. https://doi.org/10.3897/mycokeys.53.33508
Hewitt G.M. 1999. Post-glacial re-colonization of European biota. – Biol. J. Linn. Soc. 68: 87–112. https://doi.org/10.1111/j.1095-8312.1999.tb01160.x
Horvat I., Glavac V., Ellenberg H. 1974. Vegetation Südosteuropas. Stuttgart. 752 p.
[Insarov, Pchelkin] Инсаров Г.Э., Пчелкин А.В. 1982. Количественные характеристики состояния эпифитной лихенофлоры биосферных заповедников. Березинский заповедник. Москва. 58 с.
Insarov G., Schroeter B. 2002. Lichen monitoring and climate change. – In: Monitoring with lichens – Monitoring lichens. Dordrecht. P. 183–201. https://doi.org/10.1007/978-94-010-0423-7_13
Jäger E. 1968. Die pflanzengeographische Ozeanitätsgliederung der Holarktis und die Ozeanitätsbindung der Pflanzenareale. – Feddes Repert. 79: 157–335. https://doi.org/10.1002/fedr.19680790302
[Khomich et al.] Хомич В.С., Логинов В.Ф., Мельник В.И., Табальчук Т.Г., Семенченко В.П., Кулак А.В., Степанович И.М. 2018. Признаки остепнения южной части Беларуси. – В сб.: Материалы VIII междунар. симпозиума “Степи Северной Евразии”. Оренбург. С. 1051–1054.
[Kobzar] Кобзарь Н.Н. 1985. Географический анализ лихенофлоры Березинского государственного биосферного заповедника. – В сб.: Тезисы докл. “Грибы и лишайники в экосистеме”. Т. 2. Рига. С. 72–74.
[Kobzar] Кобзарь Н.Н. 2004. Биоразнообразие и географический анализ лихенофлоры центральной части Беларуси. – В сб.: Материалы Междунар. конф. “Биология, систематика и экология грибов в природных экосистемах и агрофитоценозах”. Минск. С. 125–128.
[Kondratyuk] Кондратюк С.Я. 1988. Некоторые проблемы географического анализа лихенофлоры. – В сб.: Тез. докл. XI симпозиума микологов и лихенологов Прибалтийских республик и Белоруссии. Таллин. С. 133–136.
Kukwa M. 2006. The lichen genus Lepraria in Poland. – Lichenologist. 38 (4): 293–305. https://doi.org/10.1017/S0024282906005962
[Krasnaya…] Красная книга Республики Беларусь: Редкие и находящиеся под угрозой исчезновения виды дикорастущих растений. 2015. Минск. 448 с.
[Kreyer] Крейер Г.К. 1913. К флоре лишайников Могилевской губернiи. Сборы 1908–1910 годов. – Труды Импер. ботан. сада. 31 (2): 263–440.
Lättman H., Milberg P., Palmer M.W., Mattson J.-E. 2009. Changes in the distribution of epiphytic lichens in southern Sweden using a new statistical method. Nord. J. Bot. 27: 418–418. https://doi.org/10.1111/j.1756-1051.2009.00425.x
[Lazarenko] Лазаренко А.С. 1956. Основнi засади классифiкацii ареалiв листяних мохiв Радянського Далекого Сходу. – Укр. бот. журн. 13 (1): 31–40.
Lendemer J.C. 2011. A taxonomic revision of the North American species of Lepraria s.l. that produce divaricatic acid, with notes on the type species of the genus L. incana. – Mycologia. 103 (6): 1216–1229. https://doi.org/10.3852/11-032
Lendemer J.C., Anderson Stewart C.R., Besal B., Goldsmith J., Griffith H., Hoffman J.R., Kraus B., LaPoint P., Li L., Muscavitch Z., Schultz J., Schultz R., Allen J.L. 2017. The lichens and allied fungi of Mount Mitchell State Park, North Carolina: a first checklist with comprehensive keys and comparison to historical data. – Castanea. 82 (2): 69–97. https://doi.org/10.2179/17-126
Lenoir J., Gégout J.C., Marquet P.A., De Ruffray P., Brisse H. 2008. A significant upward shift in plant species optimum elevation during the 20th century. – Science. 320: 1768–1771. https://doi.org/10.1126/science.1156831
Leuschner C., Ellenberg H. 2017. Ecology of central European forests, vegetation ecology of Central Europe. Vol. 1. Cham. 971 p. https://doi.org/10.1007/978-3-319-43042-3
[Makarevich] Макаревич М.Ф. 1963. Аналiз лiхенофлори Украiнських Карпат. Киiв. 262 с.
[Malysheva] Малышева Н.В. 2001. Изменение окрестностей Санкт-Петербурга. 5. Изменение видового состава лишайников парка “Осиновая роща” за 200 лет. – Новости сист. низш. раст. 34: 162–166.
McCarthy J.J., Canziani O.F., Leary N.A., Dokken D.J., White K.S. 2001. TAR Climate Change 2001: Impacts, Adaptation, and Vulnerability. Contribution of Working Group II to the Third Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge. 1042 p.
Mies B., Lösch R. 1995. Relative habitat constancy of lichens on the Atlantic islands. – Crypto-gamic Botany 5: 192–198.
[Muchnik] Мучник Е.Э. 2003. Лихенофлора Центрального Черноземья: таксономический и эколого-географический анализы, вопросы охраны: Дис. … докт. биол. наук. Воронеж. 485 с.
Muñoz J., Felicisimo A.M., Cabezas F., Burgaz A.R., Martinez I. 2004. Wind as a long-distance dispersal vehicle in the southern hemisphere. – Science. 304: 1144–1147. https://doi.org/10.1126/science.1095210
Myllys L., Stenroos S., Thell A., Ahti T. 2003. Phylogeny of bipolar Cladonia arbuscula and Cladonia mitis (Lecanorales, Euascomycetes). – Mol. Phylogenet. Evol. 27: 58–69. https://doi.org/10.1016/S1055-7903(02)00398-6
Nascimbene J., Spitale D. 2017. Patterns of beta-diversity along elevational gradients inform epiphyte conservation in alpine forests under a climate change scenario. – Biol. Conserv. 216: 26–32. https://doi.org/10.1016/j.biocon.2017.09.021
Nascimbene J., Nimis P.L., Mair P., Spitale D. 2018. Climate warming effects on epiphytes in spruce forests of the Alps. – Herzogia. 31: 374–384. https://doi.org/10.13158/heia.31.1.2018.374
[Oksner] Окснер А.Н. 1940–1942. Анализ и история происхождения лихенофлоры Советской Арктики: Дис. …. докт. биол. наук. Киев–Киров. 318 с.
[Oksner] Окснер А.Н. 1944. О происхождении ареала биполярных лишайников. – Бот. журн. 29 (6): 243–256.
[Oksner] Окснер А.М. 1948. Арктичний елемент в лiхенофлорi радянського сектора Полярноï областi. – Укр. бот. журн. 5 (1): 65–82.
[Oksner] Окснер А.Н. 1974. Определитель лишайников СССР. Вып. 2. Морфология, систематика и географическое распространение. Ленинград. 281 с.
Otálora M.A.G., Martínez I., Aragón G., Molina M.C. 2010. Phylogeography and divergence date estimates of a lichen species complex with a disjunct distribution pattern. – Am. J. Bot. 97: 216–223. https://doi.org/10.3732/ajb.0900064
[Padhornaya et al.] Подгорная Е.В., Мельник В.И., Комаровская Е.В. 2015. Особенности изменения климата на территории Республики Беларусь за последние десятилетия. – Труды Гидрометцентра России. 358: 112–120.
Palice Z., Printzen C. 2004. Genetic variability in tropical and temperate populations of Trapeliopsis glaucolepidea: Evidence against long range dispersal in a lichen with disjunct distribution. – Myco-taxon. 90: 43–54.
Printzen C., Ekman S., Tønsberg T. 2003. Phylogeography of Cavernularia hultenii: evidence of slow genetic drift in a widely disjunct lichen. – Mol. Ecol. 12: 1473–1486. https://doi.org/10.1046/j.1365-294X.2003.01812.x
[Puhacheusky et al.] Пугачевский А., Степанович И., Ермохин М. 2011. Растительность в новых природных условиях. – Наука и инновации. 4: 21–24.
Saag L., Saag A., Randlane T. 2009. World survey of the genus Lepraria (Stereocaulaceae, lichenized Ascomycota). – Lichenologist. 41 (1): 25–60. https://doi.org/10.1017/S0024282909007993
[Savicz] Савич В.П. 1911. Материалы к флоре Полесья. Список лишайников, собранных в Минской губернии в 1910 г. Л.И. Любицкой. – Труд. студ. науч. кружк. Физ.-Мат. фак. СПБ унив. 3: 57–66.
[Savicz] Савич В.П. 1925. Результаты лихенологических исследований 1923 года в Белоруссии. – Зап. Инст. С. Х. 4: 1–33.
[Savicz, Savicz] Савич В.П., Савич Л.И. 1924. Краткий предварительный отчет об исследовании флоры мхов и лишайников Белоруссии летом 1923 г. – Зап. Инст. С. Х. 3: 57–72.
Schroeter B., Kappen L., Schulz F., Sancho L.G. 2000. Seasonal variation in the carbon balance of lichens in the maritime Antarctic: long-term measurements of photosynthetic activity in Usnea aurantiaco-atra. – In: Antarctic ecosystems: models for wider ecological understanding. Christchurch. P. 258–262.
[Shustov] Шустов М.В. 2006. Лишайники приволжской возвышенности. Москва. 237 с.
[Stsepanovich] Сцепановіч І.М. 1997. Ксератэрмныя (астэпаваныя) лугавыя супольніцтвы Беларусі. – Весці АН Беларусі. Сер. біял. навук. 2: 12–20.
[Trass] Трасс Х.Х. 1970. Элементы и развитие лихенофлоры Эстонии. – Учен. зап. ун-та Тарту. Труды по ботанике. 9: 5–259.
[Tsurykau] Цуриков А.Г. 2013. Лишайники юго-востока Беларуси. Гомель. 276 с.
Tsurykau A. 2016. Epiphytic lichens in differently aged and climatic Pinus sylvestris plantations in Belarus. – In: Abstracts of the 8th IAL Symposium “Lichens in Deep Time”. Helsinki. P. 127.
Tsurykau A. 2017. New or otherwise interesting records of lichens and lichenicolous fungi from Belarus. III. With an updated checklist of lichenicolous fungi. – Herzogia. 30: 152–165. https://doi.org/10.13158/heia.30.1.2017.152
Tsurykau A. 2018. A provisional checklist of the lichens of Belarus. – Opuscula Philolichenum. 17: 374–479.
Tsurykau A., Golubkov V. 2015. The lichens of the Cladonia pyxidata-chlorophaea complex in Belarus. – Folia Cryptog. Estonica. 52: 63–71. https://doi.org/10.12697/fce.2015.52.08
Tsurykau A., Golubkov V., Bely P. 2015a. The genera Hypotrachyna, Parmotrema and Punctelia (Parmeliaceae, lichenized Ascomycota) in Belarus. – Herzogia. 28 (2): 736–745. https://doi.org/10.13158/heia.28.2.2015.736
[Tsurykau et al.] Цуриков А.Г., Голубков В.В., Цурикова Н.В. 2015b. Ревизия лишайников группы видов Cladonia pyxidata в Беларуси. – Веснiк ВДУ. 2–3: 47–52.
Tsurykau A., Golubkov V., Bely P. 2016. The genus Lepraria (Stereocaulaceae, lichenized Ascomycota) in Belarus. – Folia Cryptog. Estonica. 53: 43–50. https://doi.org/10.12697/fce.2016.53.06
Tsurykau A., Golubkov V., Bely P. 2018a. The lichen genus Xanthoparmelia (Parmeliaceae) in Belarus. – Folia Cryptog. Estonica. 55: 125–132. https://doi.org/10.12697/fce.2018.55.13
[Tsurykau et al.] Цуриков А.Г., Голубков В.В., Белый П.Н. 2018b. Ревизия лишайников рода Xanthoparmelia Беларуси: X. delisei и X. pulla. – Журнал Белорусского государственного университета. Биология. 3: 21–27.
[Tsurykau, Khramchankova] Цуриков А.Г., Храмченкова О.М. 2007. Географический анализ лихенофлоры Гомельского Полесья. – Веснік МДПУ імя І.П. Шамякіна. 1: 48–52.
[Urbanavichus] Урбанавичюс Г.П. 1999. Биогеографические закономерности формирования флоры лишайников Южного Прибайкалья: Автореф. дисс. … канд. геогр. наук. Москва. 26 с.
[Urbanavichus] Урбанавичюс Г.П. 2001. Как определить географический элемент лишайников. – В сб.: Труды Первой Российской лихенологической школы. Петрозаводск. С. 223–237.
Wagner S., Zotz G., Allen N.S., Bader M.Y. 2012. Altitudinal changes in temperature responses of net photosynthesis and dark respiration in tropical bryophytes. – Annals Bot. 111: 455–465. https://doi.org/10.1093/aob/mcs267
Werth S., Gugerli F., Holderegger R., Wagner H.H., Csencsics D., Scheidegger C. 2007. Landscape-level gene flow in Lobaria pulmonaria, an epiphytic lichen. – Mol. Ecol. 16: 2807–2815. https://doi.org/10.1111/j.1365-294X.2007.03344.x
[Yatsyna] Яцына А.П. 2011. Новые находки лишайников и нелихенизированных грибов на территории Беларуси. – В сб.: Материалы VII Междунар. конф. “Актуальные проблемы экологии”. Гродно. С. 31–32.
[Yatsyna] Яцына А.П. 2013a. Влияние региональных факторов на широтное распределение лишайников сосновой формации Беларуси: географические и таксономические особенности. – Веснiк ВДУ. 6: 29–35.
[Yatsyna] Яцына А.П. 2013b. Новые виды лишайников и нелихенизированных грибов НП “Браславские озера”. – Веснік ВДУ. 2: 53–59.
[Yatsyna] Яцына А.П. 2014. Таксономический состав, эколого-фитоценотическая, биоморфологическая и географическая структура лихенобиоты сосновых лесов Беларуси: Автореф. дис. … канд. биол. наук. Минск. 26 с.
[Yatsyna] Яцына А.П. 2015. Лихенобиота фортов Гродненской крепости (Беларусь). – В сб.: Тезисы докладов III (XI) Междунар. ботанической конф. молодых ученых в Санкт-Петербурге. СПб. С. 46.
[Yatsyna] Яцына А.П. 2017. Лихенобиота спелых еловых насаждений двух особо охраняемых природных территорий Витебской области. – Веснік ВДУ. 3: 74–79.
[Yermokhin, Puhacheusky] Ермохин М.В., Пугачевский А.В. 2009. Современная динамика южной границы сплошного распространения ели (Picea abies Karst.) в Беларуси. – Известия НАН Беларуси. 1: 51–55.
Yurchenko E.O. 2011. Lichens of Belarus: an illustrated electronic handbook. Minsk. 1 CD.
[Yurkevich, Geltman] Юркевич И.Д., Гельтман В.С. 1962. Распространение ели в БССР. – В сб.: Лесоводственная наука и практика. Минск. С. 53–78.
[Yurkevich et al.] Юркевич И.Д., Голод Д.С., Адерихо В.С. 1979. Растительность Белоруссии, ее картографирование, охрана и использование. Минск. 248 с.
[Yurtsev] Юрцев Б.А. 1966. Гипоарктический ботанико-географический пояс и происхождение его флоры. – Комаровские чтения. 19: 1–94.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал