

Ботанический журнал, 2019, T. 104, № 9, стр. 1333-1354
МХИ В РАСТИТЕЛЬНЫХ СООБЩЕСТВАХ НА СЕВЕРО-ЗАПАДЕ БОЛЬШЕЗЕМЕЛЬСКОЙ ТУНДРЫ (НЕНЕЦКИЙ АВТОНОМНЫЙ ОКРУГ)
О. М. Афонина 1, *, О. В. Лавриненко 1, 2, **, И. А. Лавриненко 1, 3, ***
1 Ботанический институт им. В.Л. Комарова РАН
197376, С.-Петербург, ул. Проф. Попова, 2, Россия
2 Государственный природный заповедник “Ненецкий”
166002 Нарьян-Мар, ул. Заводская, 2, Россия
3 Нарьян-Марский филиал Федерального исследовательского центра комплексного изучения
Арктики им. академика Н.П. Лаверова РАН –
“Нарьян-Марская сельскохозяйственная опытная станция”
166000 Нарьян-Мар, ул. Рыбников, 1А, Россия
* E-mail: stereodon@yndex.ru
** E-mail: lavrino@mail.ru
*** E-mail: lavrinenkoi@mail.ru
Поступила в редакцию 02.07.2019
После доработки 09.09.2019
Принята к публикации 10.09.2019
Аннотация
Аннотированный список мхов северо-запада Большеземельской тундры (Ненецкий автономный округ) включает 169 видов и составлен на основе определения образцов, собранных при выполнении геоботанических описаний в 10 пунктах исследования в период с 2009 по 2017 г. Распространение мхов в 22 типах растительных сообществ приведено с указанием константности и среднего значения обилия. Бóльшая часть (66%) видов встречена в 1–3 и лишь 7% – в 10 и более типах сообществ. Приведены сведения о фитоценотическом участии мхов. Впервые для Большеземельской тундры отмечены 47 видов, из них один – Oncophorus integerrimus впервые указан для округа.
Флора мхов Ненецкого автономного округа до сих пор изучена слабо и неравномерно. В обобщающей сводке по мхам для Канино-Печорского региона, который согласно флористическому делению Российской Арктики (Yurtsev et al., 1978) в значительной степени совпадает с границами округа, О.М. Афониной и И.В. Чернядьевой приведено 255 видов (Afonina, Czernyadjeva, 1995, 1996). Более поздние бриофлористические и геоботанические исследования в регионе, опубликованные в ряде статей, дополнили эти данные (Kuzmina, 2001; Afonina, 2006; Afonina et al., 2007, 2008, 2015; Zheleznova et al., 2007; Zeleznova, 2010; Zheleznova, Shubina, 2015; Afonina, Lavrinenko, 2018). Провизорно для этого сектора Арктики в настоящее время известно более 350 видов мхов.
Начало изучения бриофлоры Большеземельской тундры связано со сборами А.Г. Шренка, который в 1837 г. пересек современную территорию Ненецкого автономного округа с запада на восток и обратно. Для арктической части он привел 26 видов мхов (Schrenk, 1854). Позднее (с 1899 г. по 1911 г.) сборы мохообразных в различных северных регионах Европейской Арктики проводил Р.Р. Поле. Его сборы были обработаны В.Ф. Бротерусом, и по результатам обработки была опубликована сводка, в которой для Большеземельской тундры приводится 68 видов (Pole, 1905). Сведения о мохообразных юго-восточной части Большеземельской тундры содержатся в ряде статей, и обобщающий список для этого района включает 184 вида (Katenin, Boch, 1970; Kildyushevsky, 1973; Zheleznova, 1978, 1982). Данные о мхах (54 вида) в нарушенных и естественных местообитаниях на территории Варандейского и Торавейского нефтяных месторождений (Большеземельская тундра) содержатся в работе Е.Ю. Кузьминой (Kuzmina, 2001). В ходе геоботанических исследований, проводимых О.В. Лавриненко и И.А. Лавриненко в последние годы, получены дополнительные данные для бриофлоры Большеземельской тундры.
В настоящей статье приведены сведения о распространении и фитоценотическом участии мхов, полученные, в основном, при выполнении геоботанических описаний, а не при целенаправленных бриологических исследованиях на северо-западе Большеземельской тундры. Такие данные важны для уточнения ареалов видов и их экотопической принадлежности.
Район исследования
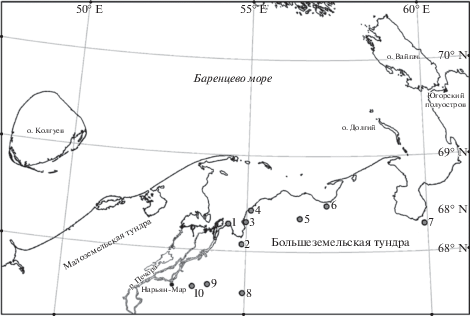
Детальные сборы мхов проводили в период с 2009 по 2017 г. при изучении растительности в 10 пунктах на северо-западе Большеземельской тундры (рис. 1).
Рис. 1.
Картосхема района работ.
1–10 – номера пунктов исследования: 1 – п-ов Болванский, северная часть; 2 – дельта рек Нерута–Ячей, Болванская губа; 3 – бассейн р. Хыльчую, Печорская губа; 4 – бассейн р. Большая Двойничная, Печорская губа; 5 – бассейн р. Большая Хэхэганьяха, возвышенность Вангуреймусюр; 6 – дельта рек Фатей–Луцаяха, Паханческая губа; 7 – дельта р. Море-Ю, Хайпудырская губа; 8 – бассейн р. Шапкина; 9 – бассейн р. Северная; 10 – бассейн р. Куя.
Fig. 1. Map of the study area.
1–10 – numbers of research sites: 1 – Bolvanskiy Peninsula, northern part; 2 – Neruta–Yachey Rivers delta, Bolvanskaya Bay; 3 – Khylchuyu River basin, Pechorskaya Bay; 4 – Bol’shaya Dvoynichnaya River basin, Pechorskaya Bay; 5 – Bol’shaya Khekhegan’yakha River basin, Vangureimusyur Upland; 6 – Fatey–Lutsayakha Rivers delta, Pahancheskaya Bay; 7 – More-Yu River delta, Haipudyrskaya Bay; 8 – Shapkina River basin; 9 – Severnaya River basin; 10 – Kuya River basin.
Большеземельская тундра занимает Печорскую низменность от правобережья р. Печоры на западе до хр. Пай-Хой на востоке и представляет собой аккумулятивную равнину с максимальной высотной отметкой – 242 м над ур. м. От берега Баренцева моря в глубь материка она поднимается уступами нескольких древних террас, сложенных глинами и морскими песками, перекрытыми прерывистым чехлом торфа 0.5–5.0 м толщиной. Плоский рельеф прибрежной зоны сменяется холмистым и холмисто-увалистым вдали от морского побережья. Холмы и вытянутые на десятки километров увалы – это конечные морены, сложенные, в основном, суглинками с включением валунно-галечного материала. Обширные понижения между ними заняты болотами и озерными котловинами остаточно-ледникового и термокарстового происхождения. Глубина эрозионного расчленения меняется от 10 до 70 м (Nenets …, 2000).
Пункты исследования: 1–4, 6 и 7 – приморские. Пункт 1 расположен в северной части п-ова Болванский, другие – находятся в бассейнах рек, впадающих в губы Баренцева моря: Нерута и Ячей (2), формирующие общую дельту, – в Болванскую губу, Хыльчую (3) и Большая Двойничная (4) – в Печорскую, Фатей и Луцаяха (6), формирующие общую дельту, – в Паханческую, Море-Ю (7) – в Хайпудырскую. Пункт 5 находится в бассейне р. Большая Хэхэганьяха на возвышенности Вангуреймусюр, остальные – реки Шапкина (8), Северная (9) и Куя (10) – принадлежат бассейну р. Печоры.
Низкие (1–3 м над ур. м.) современные аккумулятивные террасы в дельтах и приустьевых частях рек, впадающих в губы (пункты 2–4, 6 и 7), сильно заозеренные и изрезанные многочисленными связанными с морем протоками, заняты солеными и солоноватыми маршами. При продвижении от моря в глубь материка они сменяются более высокими флювиогляциально-морскими террасами, отграниченными абразионным уступом от одного до нескольких метров высотой. Берега п-ова Болванский в районе мыса Болванский нос (1) обрывистые, суглинистые, местами с нависающими сверху толщами торфа.
В пунктах 1, 5 и 8 рельеф мелкосопочный и холмисто-увалистый (высоты от 20 до 170 м над ур. м.) – суглинистые сопки и увалы с плоскими вершинами и пологими склонами разделены заболоченными понижениями.
Реки Северная (9) и Куя (10) пересекают толщу песков, отложенных бореальной морской трансгрессией. Большую площадь имеют песчаные обнажения с развитым эоловым рельефом и представленные котловинами выдувания.
Растительные сообщества
Растительные сообщества, в которых проводили сборы мхов, объединены в следующие типы:
1. Редкоивовые кустарничково-осоково-моховые сообщества, как правило, с пятнами-медальонами на плакорах, в пунктах 1, 3, 5 и 6 относящиеся к асс. Dryado octopetalae–Hylocomietum splendentis Andreyev ex Lavrinenko et Lavrinenko 201811, в пункте 7 – к асс. Calamagrostio lapponicae–Hylocomietum splendentis Lavrinenko et Lavrinenko 2018 (Lavrinenko O.V., Lavrinenko I.A., 2018а).
2. Кустарничковые, кустарничково-лишайниковые и ерниковые (Betula nana)22 кустарничково-лишайниковые сообщества на песчаных почвах водоразделов (союз Loiseleurio-Arctostaphylion Kalliola ex Nordhagen 1943, класс Loiseleurio procumbentis–Vaccinietea Eggler ex Schubert 1960).
3. Кочкарники (Eriophorum vaginatum) кустарничково-сфагновые на торфянисто-глеевых почвах в типичных тундрах.
4. Осоково (Carex globularis)-кустарничково-моховые сообщества на торфянисто-глеевых почвах в южных тундрах и лесотундре – асс. Carici globularis–Pleurozietum schreberi Lavrinenko et Lavrinenko 2015 (Lavrinenko O.V., Lavrinenko I.A., 2015).
5. Разнотравные (Astragalus subpolaris, Ranunculus propinquus, Trollius europaeus, Viola biflora) луга на склонах.
6. Разнотравно (Astragalus frigidus, Hedysarum arcticum, Oxytropis sordida)-кустарничково (Dryas octopetala, Salix hastata, S. reticulata)-моховые сообщества на склонах и бровках террас.
7. Сообщества черники (Vaccinium myrtillus) травяно-лишайниково-моховые на склонах обычно южной экспозиции на почвах легкого механического состава (союз Phyllodoco–Vaccinion myrtilli Nordhagen 1943, класс Loiseleurio procumbentis–Vaccinietea Eggler ex Schubert 1960).
8. Сообщества дерена (Chamaepericlymenum suecicum) лишайниково-моховые на склонах обычно южной экспозиции на почвах легкого механического состава (союз Phyllodoco–Vaccinion myrtilli Nordhagen 1943, класс Loiseleurio procumbentis–Vaccinietea Eggler ex Schubert 1960).
9. Бугристо-западинные комплексы, распространенные в нижних частях и на подошвах склонов, – травяно-кустарничково-моховые сообщества на буграх (0.5–0.8 (до 1.0) м выс., 1.0–2.0 м в диаметре) и мохово-печеночниковые – в межбугровых понижениях с суглинистыми почвами (в виде пятен или периодически обводненных западин).
10. Разнотравно (Draba sibirica, Saxifraga cernua, Tofieldia pusilla, Veratrum lobelianum)-ивково (Salix polaris, S. reticulata)-моховые сообщества среди курумников (россыпей камней) на подошвах склонов.
11. Нивальные травяно (Omalotheca sylvatica, Sibbaldia procumbens, Veronica alpina)-ивково (Salix herbacea, S. polaris)-моховые сообщества в местах долгого лежания снега (союз Salicion herbaceae Br.-Bl. ap. Br.-Bl. et Jenny 1926, класс Salicetea herbaceae Br.-Bl. 1948).
12. Ивняки (Salix glauca, S. lanata и S. phylicifolia) разнотравные, разнотравно-хвощовые и травяно-моховые в поймах рек и ручьев, в ложбинах стока и на низких морских террасах.
13. Ивняки (Salix myrsinites) разнотравно-моховые на водоразделах.
14. Ерники (Betula nana) зеленомошные.
15. Разнотравно-злаковые (Alopecurus alpestris, Elymus fibrosus, Geranium albiflorum, Lathyrus pratensis, Solidago lapponica, Veronica longifolia) пойменные луга.
16. Вейниково-пушицево-осоковые сообщества низинных болот с участием гипновых мхов, относящиеся к союзу Drepanocladion exannulati Krajina 1933 в классе Scheuchzerio palustris–Caricetea fuscae Tx. 1937 (Lavrinenko O.V. et al., 2016).
17. Пушицево-осоково-сфагновые сообщества в топях плоскобугристых комплексных болот и сфагновые сообщества по трещинам полигональных торфяников, относящиеся к союзу Scheuchzerion palustris Nordhagen ex Tx. 1937 (синоним Sphagnion baltici Kustova 1987 ex Lapshina 2010) в классе Scheuchzerio palustris–Caricetea fuscae (Lavrinenko O.V. et al., 2016).
18. Багульниково-морошково-мохово-лишайниковые сообщества на торфяных буграх плоскобугристых комплексных болот – асс. Rubo chamaemori–Dicranetum elongati Lavrinenko et Lavrinenko 2015 в союзе Rubo chamaemori–Dicranion elongati Lavrinenko et Lavrinenko 2015 и классе Oxycocco-Sphagnetea Br.-Bl. et Tx. ex Westhoff et al. 1946 (Lavrinenko O.V., Lavrinenko I.A., 2015).
19. Сообщества соленых и солоноватых маршей разных уровней с участием галофитов – Arctanthemum hultenii, Calamagrostis deschampsioides, Carex glareosa, C. salina, C. subspathacea, Potentilla egedii, Plantago schrenkii, Stellaria humifusa (класс Juncetea maritimi Br.-Bl. in Br.-Bl. et al. 1952) (Lavrinenko O.V., Lavrinenko I.A., 2018б).
20. Группировки бриофитов на зарастающих аллювиальных песках.
21. Погруженные заросли в небольших водоемах.
22. Группировки бриофитов на антропогеннонарушенных местообитаниях (карьеры, зимники).
Видовой состав мхов в сообществах
Проанализировано 429 геоботанических описаний (от 5 до 65 в разных типах сообществ). Список мхов, включающий 169 видов, приведен в таблице в алфавитном порядке. Номенклатура мхов дана в основном в соответствии со Списком мхов Восточной Европы и Северной Азии (Ignatov et al., 2006) c учетом последних монографических обработок ряда систематических групп, вошедших в опубликованные два тома Флоры мхов России (Moss flora …, 2017; 2018). Нами были приняты во внимание изменения в роде Oncophorus, предложенные L. Hedenӓs (2017). Им был описан новый близкий к O. virens вид – O. integerrimus Hedenӓs, который обнаружен в сборах из Большеземельской тундры. Согласно этому автору, Oncophorus wahlenbergii – это комплекс, представленный 3 видами – O. wahlenbergii Brid., O. elongates (I. Hagen) Hedenäs и O. demetrii (Renauld et Cardot) Hedenäs. Все они есть в Большеземельской тундре, однако сейчас невозможно ревизовать весь собранный нами материал и представить распределение этих видов по растительным сообществам, поэтому в данной статье O. wahlenbergii приведен в прежнем объеме. Согласно последним молекулярным исследованиям рода Hypnum (Schlesak et al., 2018), Stereodon (Hypnum) bambergeri (Schimp.) Lindb. переведен в род Campylium (C. bambergeri (Schimp.) Hedenäs, Schlesak, D. Quandt); Stereodon (Hypnum) plicatulus Lindb. – в род Aquilonium (A. plicatulum (Lindb.) Hedenäs, Schlesak, D. Quandt.), но в статье оставлены их прежние названия.
В пунктах работ богатство бриофлоры определяется не только степенью изученности (числом выполненных геоботанических описаний), но и разнообразием растительности. Наименьшее число видов (33 и 39) отмечено в низменных пунктах 2 и 10 (реки Нерута–Ячей и Куя соответственно), наибольшее (81 и 112) – в пунктах 1 и 5 с мелкосопочным рельефом (п-ов Болванский и р. Большая Хэхэганьяха на возвышенности Вангуреймусюр соответственно).
Численность мхов в разных типах сообществ варьирует от 6–7 видов (пойменные луга, песчаный аллювий и водные местообитания, сообщества 15, 20, 21 в таблице) до 90 (сообщества на плакорах, 1). Ранее уже было показано (Lavrinenko O.V., Lavrinenko I.A., 2018а), что именно зональные синтаксоны в восточноевропейских тундрах, по сравнению с любыми интразональными, самые богатые флористически (при пропорциональном числе видов сосудистых растений, мохообразных и лишайников), что обусловлено выровненными условиями среды (средние условия увлажнения, укрытости снегом, относительно богатые почвы) на плакорах, позволяющими сосуществовать наибольшему числу видов. Высокой насыщенности зональных сообществ видами способствует также хорошо выраженный микро- и нанорельеф, обусловливающий многообразие экологических ниш (пятна грунта, повышенные валики и бугорки, ложбинки и западинки) для мхов с разной экологией и конкурентной способностью. Больше половины видов (59) встречены менее чем в 20% зональных сообществ с низким (r, +) обилием. Постоянно или часто встречаются 15 видов, из которых содоминантами являются лишь 5 – Aulacomnium turgidum, Hylocomium splendens, Rhytidium rugosum (на повышенных элементах рельефа), Stereodon bambergeri и Tomentypnum nitens (на пятнах). Только в редкоивовых кустарничково-осоково-моховых сообществах отмечено 17 мхов – Brachythecium cirrosum, B. coruscum, Brachytheciastrum velutinum, Bryum wrightii, Conostomum tetragonum, Dicranum leioneuron, Didymodon rigidulus, Distichium inclinatum, Encalypta rhaptocarpa, Flexitrichum gracile, Hypnum cupressiforme, Myurella julacea, Orthothecium strictum, Splachnum vasculosum, Stereodon holmenii, S. plicatulus, S. subimponens.
В кустарничково-лишайниковых сообществах в ветрообдуваемых местообитаниях с хорошо дренированными бедными и кислыми автоморфными почвами (2) постоянны и часты Dicranum elongatum, Hylocomium splendens (с невысоким обилием), Pogonatum dentatum, Polytrichum hyperboreum, P. piliferum, Racomitrium lanuginosum. Только в этих сообществах найдены Niphotrichum canescens (бывает обильным), Ptilium crista-castrensis, Tetraplodon paradoxus, T. urceolatus.
Богаты мхами разнотравно-кустарничково-моховые сообщества на склонах (6) и сообщества, сформированные в основании склонов холмов и перегибов террас, где есть разные элементы рельефа – бугорки, межбугорковые понижения, пятна суглинка, скопления каменистого материала (9 и 10). В них обнаружено по 38–40 видов, среди доминантов – Aulacomnium turgidum, Hylocomium splendens, Sanionia uncinata и Tomentypnum nitens. На склонах и бровках террас часты и бывают обильны – Abietinella abietina, Climacium dendroides, Dicranum acutifolium, D. bonjeanii, Rhytidiadelphus triquetrus и Rhytidium rugosum, в основании склонов среди курумников – Brachythecium erythrorrhizon, Campylium stellatum, Dicranum spadiceum, Hylocomiastrum pyrenaicum, Oncophorus integerrimus, O. wahlenbergii, Orthothecium chryseum, Pohlia cruda, в бугристо-западинных комплексах – Pleurozium schreberi и Polytrichum commune на повышениях, Catoscopium nigritum, Kiaeria glacialis, Pseudocalliergon turgescens, Scorpidium revolvens – в межбугорковых понижениях. Только в сообществах на склонах собран Mnium thomsonii, среди курумников – Bryum uliginosum, Dicranum polysetum, Didymodon icmadophilus, Hymenoloma crispulum, Polytrichastrum septentrionale, Schistidium sordidum, S. submuticum.
В сообществах черники и дерена (7 и 8), занимающих хорошо дренированные и укрытые зимой снегом местообитания, мхов немного (17 и 12 видов соответственно), среди доминантов – Hylocomium splendens и Pleurozium schreberi, в черничниках также – Dicranum majus, обычны Polytrichum commune и Sanionia uncinata.
Эти же мхи (Hylocomium splendens, Pleurozium schreberi и Sanionia uncinata) обычны и на склонах с разнотравными сообществами (5), но обилие их невысоко. Всего здесь отмечено 19 видов, среди которых бывают заметны также Bryum elegans, Mnium blyttii, Polytrichastrum fragile, Rhytidiadelphus triquetrus, Sciuro-hypnum reflexum.
В травяно-ивково-моховых сообществах нивальных местообитаний (11) всего 33 вида, из которых постоянны и часты Bartramia ithyphylla, Kiaeria glacialis, Polytrichastrum alpinum, Polytrichum commune, Sanionia uncinata; только здесь обнаружен Brachythecium rutabulum.
В большой группе ивняков (12), сложенных основными тундровыми ивами – Salix glauca, S. lanata и S. phylicifolia, обнаружен 61 вид, из которых постоянны лишь 2 – Rhizomnium pseudopunctatum и Sanionia uncinata. Они отмечены и в тех пойменных ивняках, где напочвенный моховой покров почти отсутствует, а мхи (в том числе Brachythecium spp.) поселяются в основании стволов и на комлях ив. В ивняках с высоким покрытием мхов, занимающих понижения на водоразделах, обильны Aulacomnium palustre, Brachythecium mildeanum, Bryum pseudotriquetrum, Hylocomium splendens и Sphagnum fimbriatum. Только в этой группе сообществ отмечены Aongstroemia longipes, Brachythecium salebrosum, Didymodon asperifolius, Mnium stellare, Plagiothecium berggrenianum, P. denticulatum, Pseudobryum cinclidioides, Rhizomnium punctatum, Warnstorfia pseudostraminea. В ивняках из Salix myrsinites (13) моховой покров многовидовой, кроме Aulacomnium palustre и Hylocomium splendens, в нем содоминируют Sphagnum warnstorfii и Tomentypnum nitens, часто встречаются Aulacomnium turgidum, Paludella squarrosa, Pleurozium schreberi.
В зеленомошных ерниках (14) из 24 выявленных видов мхов доминируют Hylocomium splendens и Pleurozium schreberi, бывает много Aulacomnium palustre, занимающего пониженные элементы нанорельефа. В примеси обычны политриховые (Polytrichum commune, P. hyperboreum, P. strictum) и сфагновые (Sphagnum angustifolium, S. fimbriatum, S. teres) мхи.
На соленых и солоноватых маршах (19), где условия специфичные, выявлено 25 видов. Большинство мхов не способны выносить сильное засоление и частое затопление водами приливов и участвуют в сложении сообществ только на маршах высокого уровня, подверженных воздействию морских брызг и затоплению лишь в самые сильные приливы и нагоны воды. В таких экотопах чаще других покрытия (до 70%) образуют обычные тундровые и болотные виды – Aulacomnium palustre, Bryum pseudotriquetrum, Drepanocladus aduncus, Sanionia uncinata. На маршах низкого уровня растут только бриумы, отнесенные к группе Bryum salinum s. l. Только в сообществах маршей отмечены Amblystegium serpens, Drepanocladus arcticus, Hennediella heimii, Platydictya jungermannioides.
В кочкарниках из Eriophortum vaginatum и в сообществах с Carex globularis на торфянисто-глеевых почвах (3 и 4) число мхов примерно равное (23 и 28) и состав их близкий. Обычны Aulacomnium palustre и A. turgidum, Dicranum elongatum и D. laevidens, Pleurozium schreberi, Polytrichum jensenii, Sphagnum russowii, в подзоне типичных тундр также – S. balticum и S. lenense, а южных – S. fuscum.
На торфяниках в сообществах асс. Rubo chamaemori–Dicranetum elongati (18) несомненным доминантом является Dicranum elongatum, постоянны D. laevidens, Pleurozium schreberi, Polytrichum strictum, всего выявлено 29 видов.
В осоково-сфагновых сообществах топей плоскобугристых болот господствуют Sphagnum lindbergii – в сырых центральных частях и S. balticum – в менее обводненных и примыкающих к торфяным буграм. Бывает много и других сфагновых мхов – S. compactum, S. jensenii, S. riparium. В сплошном их покрове обычны побеги Polytrichum jensenii. Всего обнаружен 21 вид, из них 4 сфагновых мха отмечены только в этих местообитаниях.
В вейниково-пушицево-осоковых сообществах низинных болот (17) всего 21 вид, постоянны и наиболее часты Bryum pseudotriquetrum, Calliergon giganteum, Drepanocladus polygamus, Plagiomnium ellipticum, Warnstorfia exannulata. Только в этих экотопах найдены Hygroamblystegium varium и Meesia longiseta.
Мхов, которые доминируют в сообществах Большеземельской тундры и образуют сомкнутые покровы, немного. Среди них есть виды с широкой экологической амплитудой, растущие в различных местообитаниях. Это, прежде всего, Hylocomium splendens, имеющий высокие показатели константности и обилия в тундровых ценозах, сообществах на склонах и зарослях кустарников и в то же время являющийся характерным преферентным видом сообществ на плакорах, где среднее обилие этого мха максимально (табл. 1). У Pleurozium schreberi, встречающегося в аналогичных местообитаниях, а также на торфяниках, наиболее высокое обилие отмечено в зеленомошных ерниках. В 10 и более типах сообществ найдены Bryum pseudotriquetrum (высококонстантен в низинных болотах), Dicranum majus (в черничниках) и политрихумы – Polytrichum commune (в сообществах в нижних частях и на подошвах склонов), P. hyperboreum (в кустарничково-лишайниковых тундрах) и P. strictum (в сообществах на торфяниках). Широкая амплитуда также у Sanionia uncinata (высоко константного в сообществах на склонах, маршах и в ивняках) и у обоих видов Aulacomnium, из которых A. palustre является характерным преферентным для ивняков из Salix myrsinites, а A. turgidum – для зональных тундр. Tomentypnum nitens имеет наибольшее постоянство в зональных тундрах, бугристо-западинных комплексах и в ивняках из Salix myrsinites. В кочкарниках и сообществах с Carex globularis на торфянисто-глеевых почвах и в асс. Rubo chamaemori–Dicranetum elongati на торфяниках моховые покровы образуют Dicranum elongatum и D. laevidens. Первый вид бывает обильным также и в других тундровых сообществах. К эвритопным видам можно отнести Pohlia nutans, отмеченную в большинстве изученных типов сообществ, но с низкими показателями константности и обилия. Сфагнумы образуют сомкнутые ковры, но большинство из них стенотопны и растут только в болотах и близких к ним заболоченных сообществах (кочкарниках и сообществах с Carex globularis) (см. табл. 1).
Таблица 1.
Участие мхов в сообществах Большеземельской тундры Table 1. Participation of mosses in communities of the Bolshezemelskaya tundra
| Местообитания и номера типов сообществ/Habitats and numbers of communities types | Пункты иссле- дования/ Research sites | ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Тундры/Tundras | Склоны/Slopes | Кустарниковые заросли/ Shrubs | Луга/ Meadows | Болота/ Fens and palsa bogs | Марши/ Marshes | Другое/ Other | |||||||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | ||
| Abietinella abietina (Hedw.) Fleisch. | I+ | . | . | . | . | III+ | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1, 5, 6 |
| Amblystegium serpens (Hedw.) Bruch, Schimp. et W. Gümbel | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | II2a | . | . | . | 6, 7 |
| *Aongstroemia longipes (Sommerf.) Bruch, Schimp. et W. Gümbel | . | . | . | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | 7 |
| Aulacomnium palustre (Hedw.) Schwägr. | IV1 | . | IV2a | IV+ | . | II1 | . | . | II+ | . | . | III2a | V2b | III2b | . | III2a | . | II+ | III2b | . | . | . | 1−9 |
| A. turgidum (Wahlenb.) Schwägr. | V2b | II+ | IV1 | I+ | . | IV2a | . | . | I+ | . | . | . | III+ | III1 | . | . | . | III+ | . | . | . | . | 1−10 |
| Barbula unguiculata Hedw. | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | 8 |
| Bartramia ithyphylla Brid. | . | . | . | . | . | . | . | . | . | . | III+ | I+ | . | . | . | . | . | . | . | . | . | . | 1, 4, 6 |
| Brachytheciastrum velutinum (Hedw.) Ignatov et Huttunen | I+ | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 6 |
| Brachythecium cirrosum (Schwägr.) Schimp. | II+ | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1, 5, 6 |
| *B. coruscum I. Hagen | I+ | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 5 |
| B. erythrorrhizon Bruch, Schimp. et W. Gümbel | . | . | . | . | . | I+ | . | . | . | IV+ | . | . | . | . | . | . | . | . | . | . | . | . | 5 |
| B. mildeanum (Schimp.) Schimp. | . | . | . | . | . | . | . | . | . | . | . | III2a | . | . | . | II+ | . | . | . | . | . | . | 2, 4, 6, 7 |
| *B. rutabulum (Hedw.) Bruch, Schimp. et W. Gümbel | . | . | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | . | 4 |
| B. salebrosum (F. Weber et D. Mohr) Bruch, Schimp. et W. Gümbel | . | . | . | . | . | . | . | . | . | . | . | II+ | . | . | . | . | . | . | . | . | . | . | 3, 5 |
| B. turgidum (Hartm.) Kindb. | I+ | . | . | . | . | I+ | . | . | . | . | I+ | II+ | . | . | . | . | . | . | . | . | . | . | 1, 5, 8 |
| Bryoerythrophyllum recurvirostrum (Hedw.) P. C. Chen | II+ | . | . | . | . | . | . | . | I+ | . | I+ | I+ | . | . | . | . | . | . | . | . | . | . | 3–5 |
| *Bryum argenteum Hedw. | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | 8 |
| *B. elegans Nees in Brid. | . | . | . | . | II+ | . | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | . | 4, 6 |
| *B. neodamense Itzigs. | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | I+ | . | . | I+ | . | . | . | 4 |
| B. pseudotriquetrum (Hedw.) P. Gaertn., B. Mey et Scherb. | I+ | . | . | . | . | I+ | . | . | I+ | . | I+ | III1 | I+ | I+ | . | V+ | . | . | III2a | III+ | + | . | 1-9 |
| *B. rutilans Brid. | I+ | . | . | . | . | . | . | . | . | I+ | I+ | . | . | . | . | . | . | . | I+ | . | . | . | 1, 3–5, 8 |
| *B. salinum I. Hagen ex Limpr. s. l. | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | III1 | . | . | . | 3, 4, 6, 7 |
| *B. teres Lindb. | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | 8 |
| *B. uliginosum (Brid.) Bruch, Schimp. et W. Gümbel | . | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | . | . | 5 |
| *B. wrightii Sull. et Lesq. | I+ | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 |
| Calliergon cordifolium (Hedw.) Kindb. | . | . | . | . | . | . | . | . | . | . | . | I+ | . | . | . | I+ | . | . | III2a | I+ | . | . | 2–7, 9, 10 |
| C. giganteum (Schimp.) Kindb. | I+ | . | . | . | . | . | . | . | . | . | . | I+ | . | . | . | V2a | . | . | I+ | . | + | . | 1–7, 9 |
| Calliergonella lindbergii (Mitt.) Hedenäs | . | . | . | . | . | . | . | . | . | I+ | I+ | I+ | I+ | . | . | I+ | . | . | . | . | + | . | 3–5, 8 |
| Campylium stellatum (Hedw.) C.E.O. Jensen | I+ | . | . | . | . | . | . | . | I+ | V+ | . | II+ | I+ | . | . | . | . | . | I+ | . | . | . | 2, 3, 5-7 |
| *Catoscopium nigritum (Hedw.) Brid. | II+ | . | . | . | . | . | . | . | V1 | . | . | . | I+ | . | . | . | . | . | . | . | . | . | 1, 3, 5, 6 |
| Ceratodon purpureus (Hedw.) Brid. | I+ | . | . | . | . | I+ | . | . | . | . | . | I+ | . | . | . | . | . | . | I+ | . | . | + | 2, 5, 6, 8 |
| Cinclidium arcticum (Bruch, Schimp. et W. Gümbel) Schimp. | I+ | . | . | . | . | I+ | . | . | II+ | . | . | . | I+ | . | . | . | . | . | . | . | . | . | 3, 5 |
| *C. stygium Sw. | . | . | . | . | . | . | . | . | . | . | . | . | I+ | . | . | I+ | . | . | . | . | . | . | 4 |
| *C. subrotundum Lindb. | I+ | . | . | . | . | . | . | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | 1, 4, 5 |
| Climacium dendroides (Hedw.) F. Weber et D. Mohr | Ir | . | . | . | I+ | III1 | . | . | Ir | . | I+ | I+ | I+ | . | . | . | . | . | . | . | . | . | 1, 4, 5, 7, 9 |
| *Conostomum tetragonum (Hedw.) Lindb. | Ir | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 5 |
| Cyrtomnium hymenophyllum (Bruch, Schimp. et W. Gümbel) Holmen | I+ | . | . | . | . | . | . | . | II+ | I+ | . | . | I+ | . | . | . | . | . | . | . | . | . | 1, 3, 5 |
| *Dichodontium pellucidum (Hedw.) Schimp. | I+ | . | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | . | 1, 4 |
| Dicranella cerviculata (Hedw.) Schimp. | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | . | . | + | 4, 8 |
| D. grevilleana (Brid.) Schimp. | . | I+ | . | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | 4, 7 |
| D. schreberiana (Hedw.) Hilf. ex H.A. Crum et L.E. Anderson | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | 8 |
| D. subulata (Hedw.) Schimp. | I+ | I+ | . | I+ | . | . | . | . | . | . | . | . | . | . | . | . | . | I+ | . | . | . | + | 1, 8 |
| D. varia (Hedw.) Schimp. | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | 8 |
| Dicranum acutifolium (Lindb. et Arnell) C.E.O. Jensen | II+ | I+ | . | . | . | III1 | II+ | I+ | I+ | . | . | . | I+ | I+ | . | . | . | I+ | . | . | . | . | 1, 3–10 |
| D. bonjeanii De Not. | II+ | I+ | . | . | . | III1 | II2a | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 5–7, 10 |
| D. elongatum Schleich. ex Schwägr. | V+ | IV2a | IV1 | III1 | . | I+ | I+ | I+ | II+ | . | I+ | I+ | I+ | I+ | . | . | I+ | V3 | . | . | . | . | 1–10 |
| D. flexicaule Brid. | I+ | III+ | . | I+ | . | . | II+ | . | II+ | . | I+ | . | . | I+ | . | . | . | I+ | . | . | . | . | 1, 4–10 |
| D. fuscescens Turner | . | . | I+ | I+ | . | . | . | . | . | . | . | . | . | I+ | . | . | . | I+ | . | . | . | . | 5, 8, 9 |
| D. groenlandicum Brid. | I+ | I+ | I+ | I+ | . | . | . | . | . | . | . | . | I+ | . | . | . | I+ | I+ | . | . | . | . | 1, 3–5, 7–10 |
| *D. laevidens R.S. Williams | III1 | I+ | IV2a | V1 | . | . | . | . | . | . | . | I+ | I+ | I+ | . | . | III+ | V+ | . | . | . | . | 1–10 |
| D. leioneuron Kindb. | I+ | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 5, 8 |
| D. majus Sm. | I+ | I+ | I+ | I+ | II1 | . | IV2b | II1 | I+ | I+ | I+ | II1 | . | I+ | . | . | . | I+ | . | . | . | . | 1–10 |
| *D. polysetum Sw. | . | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | . | . | 8, 9 |
| D. scoparium Hedw. | I+ | I+ | . | . | . | . | I+ | I+ | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1, 3, 10 |
| D. spadiceum Zett. | II1 | . | . | . | . | I+ | II2a | . | I+ | IV1 | I+ | . | . | . | . | . | . | . | . | . | . | . | 1, 5, 6, 8–10 |
| D. undulatum Schrad. ex Brid. | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | I+ | . | . | . | . | 8 |
| Didymodon asperifolius (Mitt.) H.A. Crum, Steere et L.E. Anderson | . | . | . | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | 3 |
| D. rigidulus Hedw. | I+ | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 |
| *D. icmadophilus (Schimp. ex Müll. Hal.) K. Saito | . | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | . | . | 5 |
| Distichium capillaceum (Hedw.) Bruch, Schimp. et W. Gümbel | III+ | . | . | . | . | I+ | . | . | I+ | III+ | I+ | I+ | I+ | . | . | . | . | . | . | . | . | . | 1, 3–5, 6, 8 |
| *D. inclinatum (Hedw.) Bruch, Schimp. et W. Gümbel | I+ | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1, 5 |
| Ditrichum heteromallum (Hedw.) E. Britton | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | 8 |
| *D. pusillum (Hedw.) Hampe | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | 8 |
| Drepanocladus aduncus (Hedw.) Warnst. | . | . | . | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | II2a | II+ | . | . | 2–4, 6, 7 |
| D. arcticus (R.S. Williams) Hedenӓs | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | II+ | . | . | . | 2, 6, 7 |
| D. polygamus (Bruch, Schimp. et W. Gümbel) Hedenäs | I+ | . | . | . | . | . | . | . | . | . | . | I+ | I+ | . | . | III+ | . | . | II+ | . | . | . | 1–7 |
| *D. sendtneri (Schimp. ex H. Müll.) Warnst. | . | . | I+ | . | . | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | . | . | . | 5, 8 |
| *Encalypta alpina Sm. | I+ | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | . | . | 5, 6 |
| E. rhaptocarpa Schwägr. | I+ | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 5 |
| Eurhynchiastrum pulchellum (Hedw.) Ignatov et Huttunen | I+ | . | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 5, 6 |
| *Fissidens osmundoides Hedw. | I+ | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | . | . | 5 |
| Flexitrichum flexicaule (Schwägr.) Ignatov et Fedosov | V+ | I+ | . | . | . | I+ | . | . | I+ | IV+ | I+ | . | I+ | . | . | . | . | . | . | . | . | . | 1, 3, 5, 6 |
| F. gracile (Mitt.) Ignatov et Fedosov | I+ | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 5 |
| *Hennediella heimii (Hedw.) R.H. Zander | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | I+ | . | . | . | 7 |
| Hygroamblystegium varium (Hedw.) Mönk | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | 4 |
| *Hylocomiastrum pyrenaicum (Spruce) M. Fleisch. | . | . | . | . | I+ | . | . | . | . | III+ | . | . | . | . | . | . | . | . | . | . | . | . | 5, 6 |
| Hylocomium splendens (Hedw.) Bruch, Schimp. et W. Gümbel | V3 | IV2a | III1 | III+ | IV1 | V2b | V2a | IV2b | V2b | IV2a | . | III2a | V2b | V2b | . | . | . | I+ | . | . | . | . | 1–10 |
| Hymenoloma crispulum (Hedw.) Ochyra | . | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | . | . | 5 |
| *Hypnum cupressiforme Hedw. | II+ | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 6 |
| *Kiaeria glacialis (Berggr.) I. Hagen | I+ | . | . | . | I+ | . | I1 | I+ | III2a | I+ | V2b | I+ | . | . | . | . | . | . | . | . | . | . | 1, 3–6 |
| Leptobryum pyriforme (Hedw.) Wilson | . | . | . | . | . | . | . | . | . | I+ | . | I1 | . | . | . | . | . | . | I+ | . | . | . | 3, 5–7 |
| *Loeskypnum badium (Hartm.) H.K.G. Paul | I+ | . | . | . | . | . | . | . | I+ | . | . | . | I+ | . | . | I+ | . | . | . | . | . | . | 3–5 |
| *Meesia longiseta Hedw. | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | 4 |
| *M. triquetra (Jolycl.) Ångstr. | . | . | . | . | . | . | . | . | . | . | . | I+ | I+ | . | . | I+ | . | . | . | . | . | . | 1, 2, 4, 5 |
| M. uliginosa Hedw. | II+ | . | . | . | . | . | . | . | I+ | I+ | . | . | I+ | . | . | . | . | . | . | . | . | . | 1, 3–6 |
| Mnium blyttii Bruch, Schimp. et W. Gümbel | . | . | . | . | II1 | . | . | . | . | I+ | I+ | I+ | . | . | . | . | . | . | . | . | . | . | 1, 4, 5, 8 |
| M. spinosum (Voit) Schwägr. | . | . | . | . | I+ | I+ | . | . | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | 1, 3, 5 |
| M. stellare Hedw. | . | . | . | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | 3, 5 |
| *M. thomsonii Schimp. | . | . | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 5 |
| Myurella julacea (Schwägr.) Bruch, Schimp. et W. Gümbel | II+ | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1, 3, 5, 6 |
| Niphotrichum canescens (Hedw.) Bednarek-Ochyra et Ochyra | . | II2b | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 8, 10 |
| *Oncophorus integerrimus Hedenäs | . | . | . | . | . | . | . | . | . | III2a | . | I+ | . | . | . | . | . | . | . | . | . | . | 5 |
| O. virens (Hedw.) Brid. | I+ | . | . | . | . | . | . | . | . | . | I+ | I+ | . | . | . | . | . | . | . | . | . | . | 1, 4, 5, 8 |
| O. wahlenbergii Brid. | II1 | I+ | . | I+ | . | . | . | . | I+ | III2a | . | . | . | I+ | . | . | . | Ir | . | . | . | . | 1, 3–6, 8, 9 |
| Orthothecium chryseum (Schwägr.) Bruch, Schimp. et W. Gümbel | II+ | . | . | . | . | . | . | . | I+ | IV1 | I+ | . | I+ | . | . | . | . | . | . | . | . | . | 1, 5 |
| *O. strictum Lorentz | Ir | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1, 5, 6 |
| Paludella squarrosa (Hedw.) Brid. | . | . | . | . | . | . | . | . | . | . | . | I+ | III1 | . | . | II2a | . | . | . | . | . | . | 3–6 |
| Philonotis fontana (Hedw.) Brid. | I+ | . | . | . | . | I+ | . | . | I+ | II1 | I+ | I+ | I+ | . | . | . | . | . | . | I+ | + | . | 1, 4, 5, 7 |
| P. tomentella Molendo | . | . | . | . | . | . | . | . | . | . | I+ | I+ | . | . | . | . | . | . | . | . | . | . | 4 |
| Plagiomnium ellipticum (Brid.) T. J. Kop. | . | . | . | . | . | I+ | . | . | . | . | . | I+ | I+ | . | I+ | IV2a | . | . | I+ | . | . | . | 2–7, 9, 10 |
| *Plagiothecium berggrenianum Frisvoll | . | . | . | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | 6 |
| P. denticulatum (Hedw.) Bruch, Schimp. et W. Gümbel | . | . | . | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | 4, 6 |
| *Platydictya jungermannioides (Brid.) H.A. Crum | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | I+ | . | . | . | 4 |
| Pleurozium schreberi (Brid.) Mitt. | IV1 | II1 | V1 | V2a | IV+ | I+ | V2a | IV1 | V2a | I+ | . | Ir | III1 | IV2b | . | . | . | IV+ | . | . | . | . | 1–10 |
| Pogonatum dentatum (Brid.) Brid. | IIr | IV+ | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | . | 2, 3, 5–10 |
| P. urnigerum (Hedw.) P. Beauv. | II+ | Ir | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | + | 1, 4, 6, 8 |
| Pohlia cruda (Hedw.) Lindb. | IV+ | + | . | . | Ir | I+ | . | . | Ir | III1 | I+ | . | . | . | . | . | . | . | . | . | . | . | 1, 5, 8 |
| P. nutans (Hedw.) Lindb. | I+ | I+ | I+ | I+ | . | . | I+ | I+ | Ir | . | I+ | I+ | I+ | I+ | I+ | I+ | I+ | Ir | I+ | . | . | . | 1, 3–10 |
| P. wahlenbergii (F. Weber et D. Mohr) A. L. Andrews | . | . | . | . | . | . | . | . | . | I+ | . | I+ | . | . | I+ | I+ | . | . | I+ | V2a | + | . | 2, 4, 5, 7 |
| Polytrichastrum alpinum (Hedw.) G.L. Smith | I+ | I+ | . | . | . | I+ | II+ | . | . | I+ | III2a | II1 | . | . | . | . | . | . | . | . | . | . | 1, 2, 5–8, 10 |
| P. fragile (Bryhn) Schljakov | . | . | . | . | II+ | . | . | . | . | I+ | . | I+ | . | . | . | . | . | I+ | II+ | . | . | . | 5, 6 |
| P. septentrionale (Brid) E.I. Ivanova | . | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | . | . | 5 |
| Polytrichum commune Hedw. | I+ | II+ | . | II+ | I+ | . | III1 | II1 | V1 | . | IV1 | I+ | . | III1 | I+ | . | . | I+ | . | . | . | . | 1, 3–10 |
| P. hyperboreum R. Brown | III1 | V2a | I+ | III+ | I+ | I+ | II1 | I+ | . | . | . | . | . | III+ | . | . | . | I+ | . | . | . | . | 1–10 |
| P. jensenii I. Hagen | Ir | . | III1 | III+ | . | . | . | . | . | . | I+ | I+ | . | . | . | 1+ | V1 | II+ | . | . | . | . | 1–10 |
| P. juniperinum Hedw. | I+ | I+ | . | . | II+ | I+ | . | . | . | . | . | I+ | . | . | . | . | I+ | . | . | . | . | . | 1, 3, 5, 6, 8 |
| P. piliferum Hedw. | I+ | III+ | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | . | I+ | . | . | . | . | 1, 3–5, 8–10 |
| P. strictum Brid. | IV+ | I+ | II+ | . | . | I+ | I+ | I+ | III+ | . | II1 | . | I+ | III+ | . | . | . | V1 | . | . | . | . | 1–10 |
| Pseudobryum cinclidioides (Hüeb.) T.J. Kop. | . | . | . | . | . | . | . | . | . | . | . | I+ | . | . | . | I1 | . | . | I+ | . | . | . | 1, 4–6, 8, 10 |
| *Pseudocalliergon turgescens (T. Jensen) Loeske | I+ | . | . | . | . | . | . | . | III+ | . | . | . | I+ | . | . | . | . | . | . | . | . | . | 5 |
| Ptilium crista-castrensis (Hedw.) De Not. | . | I+ | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 8 |
| Racomitrium lanuginosum (Hedw.) Brid. | IV+ | V+ | . | . | . | I+ | . | . | I+ | . | . | . | . | . | . | . | . | . | . | . | . | . | 1–10 |
| Rhizomnium pseudopunctatum (Bruch et Schimp.) T.J. Kop. | I+ | . | . | . | . | . | . | . | . | . | I+ | V2a | I+ | . | . | . | . | . | . | . | . | . | 1–6, 9, 10 |
| *R. punctatum (Hedw.) T.J. Kop. | . | . | . | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | 5 |
| Rhytidiadelphus triquetrus (Hedw.) Warnst. | I+ | . | . | . | II1 | III1 | . | . | I+ | . | . | I+ | . | . | . | . | . | . | . | . | . | . | 3, 5, 7, 9 |
| Rhytidium rugosum (Hedw.) Kindb. | V2a | I+ | I+ | . | . | III+ | . | . | I+ | . | . | . | . | . | . | . | . | . | . | . | . | . | 1, 3, 5, 6, 8 |
| Sanionia uncinata (Hedw.) Loeske | I+ | I+ | . | I+ | IV1 | III2a | II1 | II2b | V1 | V2a | V2b | V2a | . | I+ | II2a | III+ | I+ | . | IV2a | I+ | . | . | 1–10 |
| Schistidium sordidum I. Hagen | . | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | . | . | 5 |
| *S. submuticum Broth. ex H.H. Blom | . | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | . | . | 5 |
| Sciuro-hypnum reflexum (Starke) Ignatov et Huttunen | . | . | . | . | II+ | . | I+ | . | . | . | I+ | I+ | . | . | I+ | . | . | . | . | . | . | . | 1, 3–6 |
| Scorpidium revolvens (Sw. ex anon.) Rubers | II1 | . | . | . | . | I+ | . | . | III1 | I+ | . | . | I+ | . | . | I+ | . | . | . | . | . | . | 1, 3–6 |
| Sphagnum angustifolium (C.E.O. Jensen ex Russow) C.E.O. Jensen | I+ | . | I+ | II1 | . | . | . | . | . | . | . | . | . | II2a | . | . | II+ | I+ | . | . | . | . | 1, 2, 7–9 |
| S. aongstroemii Hartm. | I+ | . | I1 | . | . | . | . | . | . | . | . | . | . | . | . | I+ | I+ | . | . | . | . | . | 1, 4, 9 |
| S. balticum (Russow) C.E.O. Jensen | . | . | V2b | I1 | . | . | . | . | . | . | . | . | . | . | . | . | V3 | III+ | . | . | . | . | 1, 3, 5–10 |
| S. capillifolium (Ehrh.) Hedw. | . | . | I+ | I1 | . | . | . | . | . | . | . | . | I+ | . | . | . | . | I+ | . | . | . | . | 1, 3, 4, 8–10 |
| S. compactum Lam. et DC. | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | . | I2a | . | . | . | . | . | 3, 7, 9, 10 |
| S. fimbriatum Wilson | I+ | . | I+ | . | . | . | . | . | . | . | . | III2a | . | II2a | . | I+ | I+ | . | I+ | . | . | . | 1, 2, 4, 6, 7, 9 |
| S. fuscum (Schimp.) H. Klinggr. | . | . | I+ | V2b | . | . | . | . | . | . | . | . | . | . | . | . | . | II+ | . | . | . | . | 8–10 |
| S. girgensohnii Russow | I+ | . | V1 | I+ | . | . | . | . | . | . | . | . | . | I+ | . | . | II2a | I+ | . | . | . | . | 1, 3, 5, 8, 9 |
| S. jensenii H. Lindb. | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | II2a | . | . | . | . | . | 10 |
| S. lenense H. Lindb. ex L.I. Savicz | . | . | V2b | . | . | . | . | . | . | . | . | . | . | . | . | . | . | I+ | . | . | . | . | 1, 3, 5, 7 |
| S. lindbergii Schimp. | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | V4 | . | . | . | . | . | 1, 5, 7–10 |
| S. obtusum Warnst. | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | 10 |
| S. riparium Ångstr. | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | I2a | . | . | . | . | . | 1, 8 |
| S. russowii Warnst. | I+ | . | IV2a | V2a | . | . | . | . | . | . | . | . | . | I+ | . | . | III2a | I+ | . | . | . | . | 1, 5, 8–10 |
| S. squarrosum Crome | I+ | . | . | . | . | . | . | . | . | . | . | II1 | . | III1 | . | I+ | . | . | I+ | . | . | . | 1-6, 9, 10 |
| S. teres (Schimp.) Ångstr. | . | . | I+ | . | . | . | . | . | . | . | . | . | I+ | III2a | . | . | . | . | . | . | . | . | 2, 4 |
| S. warnstorfii Russow | I+ | . | . | . | . | . | . | . | . | I+ | . | II+ | V2a | . | . | I+ | . | . | . | . | . | . | 3–5, 8–10 |
| *Splachnum sphaericum Hedw. | . | . | Ir | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 5 |
| *S. vasculosum Hedw. | I+ | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 |
| *Stereodon bambergeri (Schimp.) Lindb. | IV2a | I+ | . | . | . | I+ | . | . | I+ | . | . | . | . | . | . | . | . | . | . | . | . | . | 1, 5, 6 |
| *S. holmenii (Ando) Ignatov et Ignatova | II+ | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 |
| *S. plicatulus Lindb. | I+ | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 3, 6 |
| *S. subimponens (Lesq.) Broth. | I+ | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 |
| Straminergon stramineum (Dicks. ex Brid.) Hedenӓs | I+ | . | I+ | . | . | . | . | . | . | . | . | I+ | I+ | I+ | . | . | I+ | . | . | . | . | + | 1, 2, 4–9 |
| Syntrichia norvegica F. Weber | . | . | . | . | I+ | I+ | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 5 |
| S. ruralis (Hedw.) F. Weber et D. Mohr | II+ | . | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 5, 6 |
| *Tayloria lingulata (Dicks.) Lindb. | . | . | Ir | . | . | . | . | . | . | I+ | . | I+ | I+ | . | . | . | . | . | . | . | . | . | 4, 5 |
| Tetraplodon mnioides (Hedw.) Bruch, Schimp. et W. Gümbel | . | Ir | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | Ir | . | . | . | . | 1, 3, 5, 8 |
| T. paradoxus (R. Br.) I. Hagen | . | Ir | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 4 |
| *T. urceolatus (Hedw.) Bruch, Schimp. et W. Gümbel | . | Ir | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 10 |
| Timmia austriaca Hedw. | I+ | . | . | . | I+ | I+ | . | . | . | I+ | II1 | . | . | . | . | . | . | . | . | . | . | . | 1, 3, 5 |
| Tomentypnum nitens (Hedw.) Loeske | V2b | . | . | . | . | II2a | . | . | V2a | III2a | . | I+ | IV2b | . | . | . | . | . | . | . | . | . | 1, 3, 5–8 |
| Tortella fragilis (Hook. et Wilson) Limpr. | I+ | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | . | . | 3, 5 |
| Warnstorfia exannulata (Bruch, Schimp. et W. Gümbel) Loeske | . | . | . | . | . | . | . | . | . | . | . | I+ | . | . | . | III2b | III+ | . | III2b | . | . | . | 1, 2, 4–6, 8 |
| W. fluitans (Hedw.) Loeske | . | . | . | . | . | . | . | . | . | . | . | I+ | . | . | . | . | I+ | . | I+ | . | . | + | 1, 2, 6–8, 10 |
| W. pseudostraminea (Müll. Hal.) Tuom. et T.J. Kop. | . | . | . | . | . | . | . | . | . | . | . | I+ | . | . | . | . | . | . | . | . | . | . | 6 |
| W. sarmentosa (Wahlenb.) Hedenӓs | I+ | . | . | . | . | . | . | . | I+ | . | . | I+ | . | . | . | I+ | . | . | . | . | . | . | 1, 3–5 |
| W. trichophylla (Warnst.) Tuom. et T.J. Kop. | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | + | . | 5, 6 |
| Всего видов/Total species | 90 | 34 | 28 | 23 | 19 | 38 | 17 | 12 | 40 | 40 | 33 | 61 | 37 | 24 | 7 | 28 | 21 | 29 | 26 | 6 | 6 | 13 | |
| Число описаний/Number of relevés | 52 | 64 | 14 | 13 | 5 | 12 | 13 | 8 | 7 | 5 | 14 | 37 | 12 | 20 | 8 | 16 | 21 | 36 | 65 | 7 | |||
Примечание. Звездочкой отмечены виды, новые для Большеземельской тундры. Для видов в каждом типе сообщества указана константность по шкале: I – >0–20%, II – 21–40%, III – 41–60%, IV – 61–80%, V – 81–100% и среднее значение обилия по шкале Браун-Бланке: r – единично; + – менее 1%; 1 – 1–5%; 2а – 6–12%; 2b – 13–25%; 3 – 26–50%; 4 – 51–75% (Barkman et al., 1964); в водных и антропогенно нарушенных местообитаниях присутствие видов обозначено знаком “+”. Цифрами 1–22 (в шапке таблицы) обозначены типы растительных сообществ (см. текст статьи), цифрами 1–10 – пункты исследований (как на рис. 1).
Note. Asterisks denote the species, new to the Bolshezemelskaya tundra. In each type of community, the constancy is given for the species on a scale: I – >0–20%, II – 21–40%, III – 41–60%, IV – 61–80%, V – 81–100%, and the average value of abundance is given on the Braun-Blanquet scale: r – single; + – under 1%; 1 – 1–5%; 2а – 6–12%; 2b – 13–25%; 3 – 26–50%; 4 – 51–75% (Barkman et al., 1964); in aquatic and anthropogenically disturbed habitats, the presence of species is indicated by the “+” sign. The numbers 1–22 (in the header of the table) indicate the types of plant communities (see the text), the numbers 1–10 – the research sites (see Fig. 1).
Из 169 видов 109, или 66%, встречены лишь в 1–3 типах сообществ. Среди них есть виды-кальцефилы, обитающие на карбонатном обнаженном грунте в районе полуострова Болванский (1) и возвышенности Вангуреймусюр (5), – это Bryum wrightii, Catoscopium nigritum, Didymodon asperifolius, Distichium inclinatum, Orthothecium chryseum, O. strictum, Stereodon bambergeri. Стенотопные мхи, а также те, которые обилием и константностью показывают предпочтительные им местообитания, используются, наряду с сосудистыми растениями, в качестве характерных (диагностических) видов при флористической классификации растительности.
Впервые для Большеземельской тундры приведены 47 видов мхов (отмечены звездочкой в табл. 1), из них один вид – Oncophorus integerrimus впервые указывается для Ненецкого автономного округа.
ЗАКЛЮЧЕНИЕ
В ходе исследований в 10 пунктах на северо-западе Большеземельской тундры была собрана коллекция мхов с привязкой к геоботаническим описаниям. В результате обработки коллекции составлен аннотированный список мхов, включающий 169 видов, из них 46 приводятся для бриофлоры Большеземельской тундры впервые, один вид – Oncophorus integerrimus впервые указывается для Ненецкого автономного округа, 7 видов (Bryum uliginosum, Dicranum leioneuron, Didymodon asperifolius, Flexitrichum gracile, Oncophorus integerrimus, Polytrichastrum septentrionale, Warnstorfia trichophylla) недавно опубликованы нами (Afonina, Lavrinenko, 2018) как новые для этого региона. Один вид – Plagiothecium berggrenianum, найденный в районе Паханческой губы, – включен во второе издание Красной книги Ненецкого автономного округа (в печати). На настоящий момент разнообразие мхов Большеземельской тундры, с учетом литературных сведений, составляет 245 видов.
Данные, полученные при изучении мхов в 22 типах растительных сообществ, показали, что из 169 выявленных видов 109 (66%) встречены в 1–3 и лишь 12 (7%) – в 10 и более типах. Самые насыщенные мхами сообщества – это редкоивовые кустарничково-осоково-моховые, как правило, с пятнами-медальонами на плакорах, где отмечено 90 видов. Это подтверждает высказанное ранее мнение, что зональные синтаксоны в восточноевропейских тундрах, по сравнению с любыми интразональными, самые богатые флористически (при пропорциональном числе видов сосудистых растений, мохообразных и лишайников).
Список литературы
Afonina O.M. 2006. New moss records from Nenetskij Autonomous District. 1. – Arctoa. 15: 251. https://doi.org/10.15298/arctoa.15.14
Afonina O.M., Czernyadjeva I.V. 1995. Mosses of the Russian Arctic: check-list and bibliography. – Arctoa. 5: 99–142.
[Afonina, Czernyadjeva] Афонина О.М., Чернядьева И.В. 1996. Итоги изучения флоры листостебельных мхов Русской Арктики. – Новости систематики низших растений. 31: 151–167.
[Afonina, Lavrinenko] Афонина О.М., Лавриненко О.В. 2018. Новые находки мхов в Ненецком автономном округе 5. – Arctoa. 27(1): 60–61. https://doi.org/10.15298/arctoa.27.1
[Afonina et al.] Афонина О.М., Лавриненко О.В., Матвеева Н.В. 2007. К флоре мхов арктической части Ненецкого автономного округа. – Новости систематики низших растений. 41: 281–302.
Afonina O.M., Lavrinenko O.V., Kholod S.S., Lavrinenko I.A. 2008. New moss records from Nenets Autonomous District. 2. – Arctoa. 17: 192–194. https://doi.org/10.15298/arctoa.17.16
[Afonina et al.] Афонина О.М., Золотов В.И., Лавриненко О.В. 2015. Новые находки мхов в Ненецком автономном округе. 3. – Arctoa. 24: 224–225. https://doi.org/10.15298/arctoa.24.23
Barkman J.J., Doing H., Segal S. 1964. Kritische Bemerkungen und Vorschlӓge zur quantitativen Vegetationsanalyse – Acta Bot. Neerl. 13(3): 394–419.https://doi.org/10.1111/j.1438-8677.1964.tb00164.x
[Cherepanov] Черепанов С.К. 1995. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). СПб. 992 с.
Hedenäs L. 2017. Scandinavian Oncophorus (Bryopsida, Oncophoraceae): species, cryptic species, and intraspecific variation. – European Journal of Taxonomy. 315: 1–34. https://doi.org/10.5852/ejt.2017.315
Ignatov M.S., Afonina O.M., Ignatova E.A. et al. 2006. Check-list of mosses of East Europe and North Asia. – Arctoa. 15: 1–130. https://doi.org/10.15298/arctoa.15.01
Катенин А.Е., Боч М.С. 1970. Флора лесотундрового стационара. Печеночники, мхи и лишайники. – В кн.: Экология и биология растений восточноевропейской лесотундры. Л. С. 47–54.
[Kildyushevsky] Кильдюшевский И.Д. 1973. В защиту флористических обследований по квадратам. – Бот. журн. 58(4): 519–522.
[Kuzmina] Кузьмина E.Ю. 2001. Листостебельные мхи нарушенных и естественных местообитаний Варандейского и Таравейского нефтяных месторождений (Ненецкий автономный округ, Архангельская область). – Новости систематики низших растений. 35: 229–238.
[Lavrinenko O.V., Lavrinenko I.A.] Лавриненко О.В., Лавриненко И. А. 2015. Сообщества класса Oxycocco-Sphagnetea Br.-Bl. et R. Tx. 1943 в восточноевропейских тундрах. – Растительность России. 26: 55–84.
[Lavrinenko O.V., Lavrinenko I.A.] Лавриненко О.В., Лавриненко И. А. 2018а. Зональная растительность равнинных восточноевропейских тундр. – Растительность России. 32: 35–108. https://doi.org/10.31111/vegrus/2018.32.35
[Lavrinenko O.V., Lavrinenko I.A.] Лавриненко О.В., Лавриненко И.А. 2018б. Классификация растительности соленых и солоноватых маршей Большеземельской тундры (побережье Баренцева моря). – Фиторазнообразие Восточной Европы. 12 (3): 82–143. https://doi.org/10.24411/2072-8816-2018-10028
[Lavrinenko et al.] Лавриненко О.В., Матвеева Н.В., Лавриненко И.А. 2016. Сообщества класса Scheuchzerio–Caricetea nigrae (Nordh. 1936) Tx. 1937 в восточноевропейских тундрах. – Растительность России. 28: 55–88.
[Moss flora …] Флора мхов России. Том 2. Oedipodiales – Grimmiales. 2017. / М.С. Игнатов (отв. ред.). М. 560 с. (Арктоа том 26, приложение 1).
[Moss flora …] Флора мхов России. 2018. Bartramiales – Aulacomniales. Том 4. / М.С. Игнатов (отв. ред.). М. 543 с.
Mucina L., Bültmann H., Dierßen K., Theurillat J.-P., Raus T., Čarni A., Šumberová K., Willner W., Dengler J., Gavilán García R., Chytrý M., Hájek M., Di Pietro R., Iakushenko D., Pallas J., Daniëls F.J.A., Bergmeie E., Santos Guerra A., Ermakov N., Valachovič M., Schaminée J.H.J., Lysenko T., Didukh Ya.P., Pignatti S., Rodwell J.S., Capelo J., Weber H.E., Solomeshch A., Dimopoulos P., Aguiar C., Freitag H., Hennekens S.M., Tichý L. 2016. Vegetation of Europe: Hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities – Applied Vegetation Science. 19(1): 1–264. https://doi.org/10.1111/avsc.12257
[Nenets …] Ненецкий автономный округ. Современное состояние и перспективы развития. 2005. В.М. Макеев (ред.). СПб. 512 с.
[Pole] Поле Р.Р. 1915. Материалы для познания растительности северной России. 1. К флоре мхов России. – Тр. Имп. Бот. Сада. 33(1): 1–148.
Schlessak S., Hedenäs L., Nebel M., Quandt D. 2018. Cleaning a taxonomic dustbin: placing the European Hypnum species in a phylogenetic context! – Bryophyte Diversity et Evolution. 40(2): 37–54. https://doi.org/10.11646/bde.40.2.3
Schrenk A.G. 1854. Reise nach dem Nordosten des europaeschen Russlands, durch die Tundren der Samojeden, zum Arktischen Uralgebirge, auf Allerhocchsten Befehl fuer den kais. botanischen Garten zu St.-Petersburg im Jahre 1837 ausgefuehrt. – Zweiter Theil. Wissenschaftliche Beilagen. Dorpat. P. 541–546.
[Yurtsev et al.] Юрцев Б.А., Толмачев А.И., Ребристая О.В. 1978. Флористическое ограничение и разделение Арктики. – В кн.: Арктическая флористическая область. Л. С. 9–104.
[Zheleznova] Железнова Г.В. 1978. Мохообразные. Флора и фауна водоемов Крайнего Севера (на примере Большеземельской тундры). Л. С. 26–31, 151–161.
[Zheleznova] Железнова Г.В. 1982. Бриофлора юго-восточной части Большеземельской тундры. – Труды Коми научного центра УрО РАН, № 49 “Споровые растения тундровых биогеоценозов”. Сыктывкар. С. 95–108.
[Zheleznova] Железнова Г.В. 2010. Новые находки мхов в Ненецком Автономном округе. 3. – Arctoa. 19: 262. https://doi.org/10.15298/arctoa.19.24
Zheleznova G.V., Shubina T.P. 2015. Mosses of the Belaya River basin (Northern Timan, Nenets Autonomous District). – Arctoa. 24: 204–209. https://doi.org/10.15298/arctoa.24.20
[Zheleznova et al.] Железнова Г.В., Шубина Т.П., Дулин М.В., Бакалин В.А. 2007. Бриофиты юго-западной части острова Вайгач. – В сб.: Материалы всероссийской конференции “Биоразнообразие растительного покрова Крайнего Севера: инвентаризация, мониторинг, охрана”. Сыктывкар. С. 30–39.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал