

Ботанический журнал, 2019, T. 104, № 9, стр. 1431-1445
ДИНАМИКА СОСТАВА ЛУГОВ РАИФСКОГО УЧАСТКА ВОЛЖСКО-КАМСКОГО ЗАПОВЕДНИКА (ТАТАРСТАН)
Т. В. Рогова 1, *, О. В. Бакин 2, **, В. Е. Прохоров 1, ***, Г. А. Шайхутдинова 1, ****
1 Казанский (Приволжский) федеральный университет
420008 г. Казань, ул. Кремлёвская, 18, Россия
2 Волжско-Камский государственный природный биосферный заповедник
422537 Республика Татарстан, пос. Садовый, Зеленодольский р-н, ул. Вехова, 1, Россия
* E-mail: tatiana.rogova@kpfu.ru
** E-mail: vkz@mail.ru
*** E-mail: vadim.prokhorov@gmail.com
**** E-mail: gshaykhu@gmail.com
Поступила в редакцию 17.07.2019
После доработки 03.09.2019
Принята к публикации 10.09.2019
Аннотация
Сравнение результатов трех инвентаризаций флористического разнообразия Раифского участка Волжско-Камского государственного природного биосферного заповедника показывает относительную стабильность видового состава луговых сообществ на протяжении более 100 лет. С прекращением сенокошения происходит зарастание луговых участков лесом. Геоинформационный анализ спутниковых данных методами группы Change Detection и материалов государственного учета лесного фонда за двадцатилетний период показал, что скорость и характер экспансии лесных растений на луга зависит от исходной площади лугового участка, его формы и расположения в лесном массиве. Информативным показателем данного процесса может являться вероятностная оценка потенциального присутствия видов в целевом сообществе, поступающих из локального пула – набора видов, встречающихся в окружающем целевое сообщество ландшафте.
Разнообразие, состав и экологические особенности природных комплексов современных заповедных территорий и важнейших составляющих их компонентов – флоры и растительности – являются результатом процесса взаимодействия видов с абиотическими условиями среды и их реакции на антропогенное воздействие, включая и природоохранные режимы. Объяснение процессов формирования и динамики видового разнообразия сообщества является одним из центральных вопросов экологии, имеющим большое природоохранное значение. Видовой состав сообществ, определяемый условиями экотопов, экологическими особенностями видов и характером биотических связей, зависит и от таких факторов, как видообразование, расселение и вымирание видов. Попытки решения этих вопросов привели к разработке концепции пула видов (Taylor et al., 1990). Последний обычно рассматривается как набор видов, потенциально способных сосуществовать в определенном сообществе (Erikson, 1993). Вхождение видов в сообщество связано с их “фильтрацией” под воздействием комплекса абиотических и биотических факторов (Zobel et al., 1998). М. Пяртель и соавт. (Pärtel et al., 1996) предложили различать три пространственных уровня пула видов: актуальный, локальный и региональный. Актуальный пул видов – это набор видов, существующих в конкретном экотопе и формирующих определенное (целевое) сообщество. Локальный пул видов – набор видов, встречающихся в окружающем целевое сообщество экотопе и способных относительно быстро мигрировать в целевое сообщество. Региональный пул представляет собой набор видов, способных сосуществовать в целевом сообществе и встречающихся на достаточно большой территории ландшафта с относительно однородными физико-географическими условиями. Очевидно, что при масштабировании пула видов может быть использована базирующаяся на ландшафтоведении отечественная классификация биохорологического разнообразия флоры, где выделяются уровни: микроэкотопов и фитоценозов – флоротопологические комплексы; мезоэкотопов, урочищ и местностей – экотопологические парциальные флоры; макроэкотопов, ландшафтов – конкретные флоры по Толмачеву (Tolmachev, 1974); флоры вышестоящих единиц биогеографического районирования (Yurtsev, 1987). Согласно концепции пула видов, видовой состав на каждом уровне определяется видовым составом уровня более высокого ранга. Актуальный пул видов можно рассматривать как нечеткое множество видов, к которому с разной степенью вероятности могут принадлежать все виды локальной и региональной флоры, при этом вероятность участия вида в сообществе зависит от степени соответствия экологических требований вида условиям экотопа. Это позволяет оценивать пространственный масштаб и закономерности формирования видового и ценотического разнообразия отдельных территорий.
При наличии многолетних рядов наблюдений оценка динамики состава видового разнообразия может быть проведена с применением понятия исторический пул видов (Giarrizzo et al., 2015, 2017). Динамические аспекты формирования растительных сообществ успешно учитываются моделью “Carousel Model” Ван Дер Маареля (Van der Maarel et al., 1993), согласно которой растительные сообщества в ходе динамических изменений пополняются видами за счет локального пула. Ранее предпринимались попытки оценки биологического разнообразия растительного покрова Раифского участка Волжско-Камского государственного природного биосферного заповедника (Раифы) на основе концепции пула видов (Lubina, Rogova, 2008).
Относительно хорошая сохранность Раифского лесного массива, около двухсот лет принадлежащего Богородицкому Раифскому монастырю, привлекала к себе внимание ботаников с конца XIX в. В первых публикациях, наряду с описаниями лесной растительности, приводятся сведения и по луговой флоре. Так, С.И. Коржинский в работе “Северная граница черноземно-степной области восточной полосы европейской России в ботанико-географическом и почвенном отношении” (Korzhinskii, 1888) приводит список из 46 видов растений, характерных для лесных опушек и дорог. А.Я. Гордягиным (Gordyagin, 1889) дано описание лесного луга в квартале 75; автор упоминает 74 вида, в том числе виды, позднее в Раифе не отмечавшиеся (Gentiana cruciata L., Potentilla erecta (L.) Raeusch., Succisa pratensis Moench). В 1925 г. Л.Н. Васильевой и А.Д. Плетнёвой-Соколовой под руководством Гордягина была проведена первая инвентаризация флоры Раифы (Spisok…, 1968). После организации в 1960 г. заповедника И.И. Гараниной в 1964–1968 гг., проводилась вторая инвентаризация флоры (Garanina, 1968), а в 2004–2008 гг. и по настоящее время сотрудниками кафедры общей экологии Казанского университета – третья. Предпринимались и специальные исследования флоры лугов (Atsekhovskii et al., 1985).
При изучении различных аспектов динамики растительного покрова все шире используют данные дистанционного зондирования Земли. Регулярность получения спутниковых данных позволяет наблюдать за текущим состоянием сообществ, выявлять изменения и закономерности их проявления, прогнозировать дальнейшее развитие. Исследования базируются на анализе характерных признаков растительности – спектральной отражательной способности, различающейся для разных длин волн. Для работы со спектральной информацией, получаемой с помощью космосъемки, разработаны различные “индексы” (вегетационные – NDVI, EVI; листовой поверхности – LAI; фотосинтетически активной радиации, поглощаемой растительностью – FPAR и проч.). На основе расчета индексов формируются индексные изображения, что позволяет выделить исследуемый объект и оценить его состояние через характер вегетации. Для оценки динамики растительности и выявления сукцессионных смен эффективно применяется группа методов Change Detection, основанных на расчетах вероятности изменений при сравнении пар индексных изображений одной и той же территории, выполненных в условиях одной фенофазы через длительный промежуток времени (Song et al., 2001). Для территории Раифского участка по серии данных спутника Landsat ранее был выполнен анализ вероятности изменения растительного покрова за двадцатилетний период с помощью трех методов группы Change Detection: “вычитанием” (subtraction), методом главных компонент (Principal Component Analysis – PCA) и методом многомерного детектирования изменений (Multivariate Alteration Detection – MAD) (Buntova et al., 2016).
Целью настоящего исследования стало изучение динамики видового состава луговых сообществ Раифы. Экотопологические (парциальные, по Б.А. Юрцеву) флоры луговых сообществ в условиях лесной зоны обязаны своим составом регулярному сенокошению или выпасу скота в экотопах, пригодных для существования лесных экосистем. На протяжении большей части истории заповедника на его территории имелись сенокосные луговые участки, к которым было приурочено около третьей части всех видов заповедной флоры сосудистых растений. Лишь в начале XXI в. сенокошение в заповеднике было прекращено. Прекращение антропогенного воздействия на луговые сообщества на заповедных территориях ведет к проникновению в сообщества видов лесной флоры, зарастанию лугов и исчезновению луговых, в том числе редких видов растений. Изменения отражаются и в характере вегетации, что детектируется при анализе спутниковых изображений.
МАТЕРИАЛ И МЕТОДЫ
Расположенный в 25 км к западу от Казани Раифский участок Волжско-Камского государственного природного биосферного заповедника находится на южной границе подтаежной зоны, занимает надпойменные террасы Волги (четвертую, средненеплейстоценовую, и пятую, нижнеэоплейстоценовую), что характеризует его ландшафты как интразональные.
Оценка динамики флоры сосудистых растений и лугового растительного покрова Раифского участка проведена с использованием достоверной исторической информации о природопользовании на исследуемой территории за столетний период, отраженной в материалах лесоустройства, и доступных сведениях о геоботанических описаниях лугов и флористических находках, имеющихся в гербарных коллекциях Ботанического музея Казанского (Приволжского) федерального университета (KAZ) и в опубликованных материалах.
Для оценки динамики видового состава луговой флоры были выполнены современные геоботанические описания и отобраны описания из базы данных информационной системы “Флора” (Rogova et al., 2010), часть которой Vegetation Database of Tatarstan входит в европейский проект EVA (Chytrý et al., 2016). По результатам исследований воссозданы списки флоры заповедника для разных временных отрезков: до организации заповедника (1888–1925 гг.), период активного сенокошения в условиях заповедного режима на территории (1964–2005 гг.), после прекращения сенокошения (2006–2018 гг.). Большую долю при анализе составили материалы инвентаризации и мониторинга флоры выделов 124 кварталов Раифы, проводимые с 2001 г. Материалы обработаны с помощью модуля анализа видового разнообразия (МАВР) информационной системы “Флора”, позволяющего проводить анализ таксономической и хорологической структуры, анализ эколого-ценотического и биоморфологического спектров видового состава, по координатным привязкам данных осуществлять картографический анализ (Prokhorov et al., 2017). Классификация луговой растительности выполнена с использованием экспертной системы программы Juice 7, позволяющей идентифицировать синтаксоны до уровня классов системы Браун-Бланке и категории экотопов EUNIS Habitat Classification (Juice ver. 7.0, 2019).
Процесс зарастания лугов изучался на четырех модельных луговых участках, расположенных в южной полосе Раифы (кварталы 77, 79, 86, 87). Эта полоса занята широколиственным лесом на легкосуглинистых почвах, являющимся производным от елово-широколиственного леса. По данным государственного учета лесного фонда с 1925 по 2013 г. оценивалось происхождение и длительность существования луговых участков. Для анализа изменений, произошедших за последние два десятилетия в связи с прекращением сенокошения, материалы учета 1993 и 2013 годов (лесоустроительные планы и таксационные описания) были оцифрованы и представлены в виде векторных картографических слоев лесохозяйственных выделов с привязанными к ним таксационными данными. Для оценки характера зарастания луговых участков выполнено сопоставление геоданных, построены тематические карты и рассчитаны геометрические характеристики луговых выделов. По паре снимков спутника Landsat, датированных 11.06.1994 г. и 15.06.2013 г., методом многоканального обнаружения изменений (MAD) группы Change Detection выполнен анализ вероятности изменения NDVI луговых модельных участков.
Сравнение видового состава модельных луговых сообществ в период проведения сенокошения и после его прекращения было выполнено на основе 20 геоботанических описаний (по 10 для каждого периода). Для определения локального пула видов, потенциально способных существовать на модельных луговых участках, из базы данных “Флора” также были отобраны описания лесных сообществ, занимающих мезофильные экотопы в полосе широколиственного леса, к которой приурочены изучаемые луговые участки. Для расчета вероятности присутствия луговых и лесных видов на участках из выборок периода проведения сенокошения и после его прекращения использовался индекс сглаживания Билса, который оценивает вероятность произрастания вида на площадке на основе фактического видового состава (Beals, 1984; De Cáceres, Legendre, 2008):
где bij – вероятность вида j встретиться на площадке i; Si – число видов на площадке i; Mij – число совместных встреч видов j и k; Nk – число встреч вида k на всех площадках.Вероятности присутствия видов рассчитывались с помощью пакета Vegan (Oksanen et al., 2017) в среде статистического программирования R. Сравнение двух выборок позволило оценить динамику видового разнообразия и тенденции развития растительного покрова.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Луговая растительность Раифы представлена сообществами, относящимися к двум классам: Molinio-Arrhenatheretea Tx. 1937 (MOL), представляющим собой пастбища и сенокосные луга на плодородных почвах, распространенные по всей Европе, и Epilobietea angustifolii Tx. et Preising ex von Rochow 1951 (EPI), представляющим высокотравные травостои на лесных полянах, нарушенных лесных опушках и вырубках, на богатых питательными веществами почвах в умеренной и бореальной зонах Евразии (Mucina et al., 2016). Согласно классификации типов местообитаний, первые соответствуют категории E 2.2 (Low and medium altitude hay meadows, Низинные и средние сенокосные луга), вторые – Е 5.4 (Moist or wet tall-herb and fern fringes and meadows, Низинные увлажненные нарушенные луга и сообщества лесных опушек) (EUNIS…, 2019).
За весь период наблюдений на территории Раифы зафиксировано 764 вида сосудистых растений. Этот набор видов можно рассматривать как региональный пул видов. Динамика эколого-ценотического спектра флоры представлена в табл. 1.
Таблица 1.
Динамика эколого-ценотического спектра флоры Раифы в 1888–2018 гг. Table 2. The dynamics of ecological-cenotic spectrum of the Raifa flora in 1888–2018
| Эколого-ценотическая группа Ecological-cenotic group |
1888–1925 | 1964–2005 | 2006–2018 | |||
|---|---|---|---|---|---|---|
| Число видов Number of species |
% | Число видов Number of species |
% | Число видов Number of species |
% | |
| Лесная бореальная/Forest boreal | 33 | 6.1 | 28 | 5.1 | 33 | 5.0 |
| Лесная неморальная/Forest nemoral | 48 | 8.9 | 46 | 8.3 | 51 | 7.7 |
| Лесная бореально-неморальная/Forest boreal-nemoral | 24 | 4.4 | 22 | 4.0 | 24 | 3.6 |
| Лесная боровая/Pine forest | 26 | 4.8 | 26 | 4.7 | 25 | 3.8 |
| Болотно-лесная/Mire-forest | 22 | 4.1 | 27 | 4.9 | 28 | 4.2 |
| Опушечно-луговая/Forest fringe-meadow | 39 | 7.2 | 39 | 7.0 | 51 | 7.7 |
| Луговая/Meadow | 86 | 15.9 | 78 | 14.1 | 98 | 14.7 |
| Лугово-степная/Meadow-steppe | 22 | 4.1 | 23 | 4.2 | 33 | 5.0 |
| Лугово-болотная/Meadow-mire | 44 | 8.1 | 48 | 8.7 | 60 | 9.0 |
| Низинно-болотная/Lowland mire | 32 | 5.9 | 32 | 5.8 | 39 | 5.9 |
| Верхово-болотная/Bogs | 23 | 4.2 | 25 | 4.5 | 26 | 3.9 |
| Водно-болотная/Wetland | 30 | 5.5 | 32 | 5.8 | 41 | 6.2 |
| Водная/Aquiatic | 19 | 3.5 | 18 | 3.2 | 18 | 2.7 |
| Рудеральная/Ruderal | 90 | 16.6 | 96 | 17.3 | 102 | 15.3 |
| Культурная/Cultivated | 4 | 0.7 | 14 | 2.5 | 34 | 5.1 |
| Всего/Total | 542 | 100.0 | 554 | 100.0 | 665 | 100.0 |
Территории, занятые модельными луговыми участками, в конце XIX в. представляли собой пашенные наделы крестьян хутора Новопольский. Они были заброшены в 90-х годах XIX столетия и заросли лесом с участием липы, дуба, березы и единично ели, и только в квартале 79 участок пашни просуществовал до Великой Отечественной войны, когда был переведен под сенокос. Остальные три луговых участка были созданы в 1930-х годах на месте вновь расчищенного леса. В кв.кв. 77 и 87 они имели площадь 7.7 и 9.8 га соответственно, а в кварталах 79 и 86 – менее 1.5 га. Далее площади луговых участков, “сжимаемые” окружающим их лесом, постепенно сокращались, а в начале XXI в., после прекращения сенокошения, этот процесс ускорился (табл. 2).
Таблица 2.
Зарастание луговых участков Раифы в 1993–2013 гг. Table 2. Overgrowing of Raifa meadow areas in 1993–2013
| № участка/
№ квартала Site number/ Quarter number |
Площадь, га Area, ha |
Периметр, км Perimeter, km |
Зарастание лесом, % Forest overgrowing, % |
Формула древостоя/
Запас древесины, м3/га Stand composition/ Wood stock, m3/ha |
||
|---|---|---|---|---|---|---|
| 1993 | 2013 | 1993 | 2013 | |||
| 1/87 | 3.2 | 0.3 | 0.73 | 0.27 | 91 | 10Б/3 |
| 2/77 | 1.4 | 0 | 0.58 | 0 | 100 | 10Б/2 |
| 3/79 | 1.1 | 0.5 | 0.51 | 0.26 | 55 | 5Б 3Ив 2Лп/1 |
| 4/86 | 0.9 | 0.2 | 0.4 | 0.2 | 78 | 6Б 4Ив/2 |
Изменения, произошедшие за последние два десятилетия в связи с прекращением сенокошения, хорошо заметны на картах запаса древесины в лесохозяйственных выделах, построенных на основе оцифровки лесоустроительных планов. Отсутствие запаса древесины на выделе маркирует луговые участки, при сопоставлении карт оценивается характер их зарастания лесом (рис. 1). Заметно, что произошло частичное или полное зарастание лугов древесной растительностью, преимущественно березой (табл. 2); степень зарастания от 55 до 100%. Характер экспансии древесных растений на луга зависит не только от исходной площади лугового участка, но и от его формы и расположения в лесном массиве. Так, участки 1 и 2, находящиеся в глубине массива, почти полностью поглощены лесом. При этом участок 1, имеющий округлую форму и наименьшее соотношение периметра к площади, демонстрирует “охватывающий” тип внедрения древесных видов (от краев к центру). Участки 3 и 4, хотя и имеют небольшие площади и вытянутую форму, частично сохраняют луговой статус благодаря проявлению краевого эффекта вблизи опушки леса.
Рис. 1.
Динамика зарастания модельных луговых выделов Раифы по данным государственного учета лесного фонда 1993 и 2013 г. Исследуемые модельные участки выделены рамкой.
Fig. 1. Overgrowing dynamics of Raifa model meadow plots according to the 1993 and 2013 forest state registration. The studied model areas are outlined.
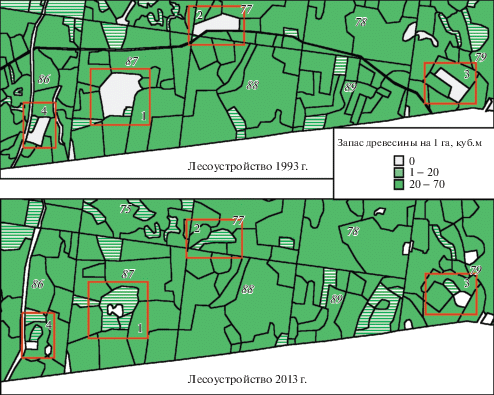
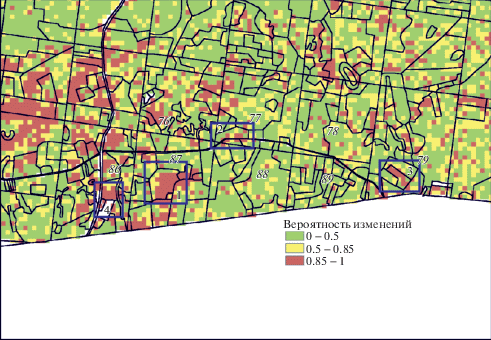
При анализе спутниковых данных выявляются выделы, где детектируются достоверные изменения NDVI в кварталах южной полосы Раифы, в том числе и на бывших луговых участках, заросших лесом (участки 2, 4). Достоверные изменения NDVI детектируются и для выделов, частично сохранивших луговой статус к 2013 г. (участки 1 и 3), что объясняется сменой состава травянистых видов и изменением продуктивности сообществ (рис. 2).
Рис. 2.
Вероятностная оценка изменения NDVI в южной полосе Раифы по данным Landsat, выполненная методом MAD группы Change Detection. Модельные участки выделены рамкой, на растровое изображение наложена сетка лесохозяйственных выделов 1993 г. Карта вероятностей отражает изменения NDVI в каждом пикселе спутникового изображения; значения вероятности в диапазоне от 0.85 до 1 подтверждают выводы об изменении луговых сообществ.
Fig. 2. Probabilistic assessment of NDVI changes in the southern part of Raifa according to Landsat data, performed by the Change Detection MAD method. Model meadow areas are outlined. Forestry allotment grid of 1993 overlaid on raster image. The probability map reflects the NDVI changes in each pixel of the satellite image; probability values in the range from 0.85 to 1 confirm the conclusions about the change in meadow communities.
Сравнение флоры сосудистых растений модельных луговых участков в период проведения сенокошения и после его прекращения выявил значительное сокращение видового состава: с 215 до 107 видов. При сохранении в целом соотношения доли видов разных эколого-ценотических групп прослеживается некоторое усиление роли лесных и опушечно-луговых видов. Сокращается как разнообразие, так и участие рудеральных видов растений в структуре видового состава лугов (табл. 3).
Таблица 3.
Эколого-ценотические спектры флоры модельных луговых участков Раифы в 1964–2018 гг. (по данным геоботанических описаний) Table 3. Ecological-cenotic flora spectrum of the model meadow sites of Raifa in 1964–2018 (according to geobotanical relevés)
| Эколого-ценотическая группа Ecological-cenotic group |
1964–2005 | 2006–2018 | ||
|---|---|---|---|---|
| Число видов Number of species |
% | Число видов Number of species |
% | |
| Лесная бореальная/Forest boreal | 1 | 0.5 | 0 | 0.0 |
| Лесная неморальная/Forest nemoral | 17 | 7.9 | 11 | 10.3 |
| Лесная бореально-неморальная/Forest boreal-nemoral | 11 | 5.1 | 8 | 7.5 |
| Лесная боровая/Pine forest | 7 | 3.3 | 1 | 0.9 |
| Болотно-лесная/Mire-forest | 1 | 0.5 | 0 | 0.0 |
| Опушечно-луговая/Forest fringe-meadow | 26 | 12.1 | 19 | 17.8 |
| Луговая/Meadow | 76 | 35.3 | 40 | 37.4 |
| Лугово-степная/Meadow-steppe | 11 | 5.1 | 3 | 2.8 |
| Лугово-болотная/Meadow-mire | 29 | 13.5 | 11 | 10.3 |
| Низинно-болотная/Lowland mire | 4 | 1.9 | 3 | 2.8 |
| Водно-болотная/Wetland | 1 | 0.5 | 2 | 1.9 |
| Рудеральная/Ruderal | 30 | 14.0 | 8 | 7.5 |
| Культурная/Cultivated | 1 | 0.5 | 1 | 0.9 |
| Всего/Total | 215 | 100.0 | 107 | 100.0 |
Локальный пул видов, характеризующий флору южной полосы Раифского лесного массива, с преобладанием сообществ класса Carpino-Fagetea sylvaticae Jakucs ex Passarge 1968 (Mucina et al., 2016), в 2005 г. насчитывал 280 видов, а в 2018 г. – 242 вида. По сравнению с региональным пулом видов в нем отсутствуют виды таких эколого-ценотических групп, как верхово-болотная и водная. Оценка возможной миграции видов локального пула в целевое сообщество (табл. 4) показывает, что после прекращения сенокошения растут вероятности присутствия на модельных участках лесных элементов флоры, как древесных форм, так и травянистых (Aegopodium podagraria L., Mercurialis perennis L., Asarum europaeum L. и др.). Более стабильным является участие в целевом сообществе лесных видов злаков и осок, а также хвощей. Также свои позиции сохраняют Urtica dioica L. и некоторые опушечно-лесные формы (Angelica sylvestris L., Anthriscus sylvestris (L.) Hoffm., Clinopodium vulgare L.). Значительно снижаются вероятности присутствия всех луговых злаков, за исключением Dactylis glomerata L., осок и видов разнотравья (Achillea millefolium L., Centaurea jacea L., Geranium pratense L., Ranunculus polyanthemos L. и др.). Характерен рост вероятности присутствия в целевом сообществе ряда гигромезофитных форм (Matteuccia struthiopteris (L.) Todaro, Agrostis stolonifera L.), что, наряду с развитием в моховом покрове лесоболотных форм (Plagiomnium cuspidatum (Hedw.) T.J. Kop., P. ellipticum (Brid.) T.J. Kop.), указывает на увеличение влажности экотопа. Последнее нужно связать с уменьшением испарения почвенной влаги после прекращения сенокошения.
Таблица 4
(фрагмент). Динамика вероятности присутствия видов в модельных луговых сообществах (Раифа, кв. 77, 79, 86, 87) Table 4 (fragment). The dynamics of the species presence probability in model meadow communities (Raifa 77, 79, 86, 87)
| Виды с уменьшающейся вероятностью Species with decreasing probability |
2005 | 2018 | Виды с уменьшающейся вероятностью Species with decreasing probability |
2005 | 2018 | Виды с увеличивающейся вероятностью Species with increasing probability |
2005 | 2018 |
|---|---|---|---|---|---|---|---|---|
| Achillea millefolium L. | 0.659 | 0.081 | Hypericum perforatum L. | 0.489 | 0.275 | Adoxa moschatellina L. | 0.163 | 0.278 |
| Agrostis capillaris L. | 0.675 | 0.039 | Knautia arvensis (L.) Coult. | 0.610 | 0.232 | Aegopodium podagraria L. | 0.747 | 0.888 |
| Agrostis gigantea Roth | 0.287 | 0.057 | Lathyrus pratensis L. | 0.637 | 0.253 | Agrostis stolonifera L. | 0.002 | 0.070 |
| Alchemilla subcrenata Buser | 0.156 | 0.085 | Leontodon hispidus L. | 0.475 | 0.006 | Asarum europaeum L. | 0.371 | 0.701 |
| Anthoxanthum odoratum L. | 0.591 | 0.068 | Leucanthemum vulgare (Vaill.) Lam. | 0.662 | 0.039 | Athyrium filix-femina (L.) Roth | 0.168 | 0.258 |
| Briza media L. | 0.441 | 0.072 | Luzula pallescens Sw. | 0.233 | 0.068 | Betula pendula Roth | 0.433 | 0.577 |
| Bupleurum longifolium L. | 0.489 | 0.179 | Lysimachia nummularia L. | 0.510 | 0.248 | Bromus inermis Leyss. | 0.062 | 0.221 |
| Campanula patula L. | 0.586 | 0.320 | Pedicularis kaufmannii Pinzger | 0.226 | 0.058 | Campanula latifolia L. | 0.067 | 0.121 |
| Carex leporina L. | 0.635 | 0.085 | Phleum pratense L. | 0.601 | 0.081 | Lactuca macrophylla subsp. uralensis (Rouy) N.Kilian & Greuter | 0.024 | 0.043 |
| Carex muricata L. | 0.324 | 0.179 | Plantago lanceolata L. | 0.592 | 0.104 | Corylus avellana L. | 0.094 | 0.169 |
| Carex pallescens L. | 0.386 | 0.179 | Plantago media L. | 0.532 | 0.029 | Dryopteris filix-mas (L.) Schott | 0.183 | 0.318 |
| Centaurea jacea L. | 0.562 | 0.198 | Polemonium caeruleum L. | 0.507 | 0.203 | Geranium sylvaticum L. | 0.089 | 0.256 |
| Silene flos-cuculi (L.) Greuter & Burdet | 0.359 | 0.041 | Potentilla thuringiaca Bernh. | 0.375 | 0.220 | Geum urbanum L. | 0.377 | 0.508 |
| Deschampsia cespitosa (L.) P.Beauv. | 0.543 | 0.289 | Prunella vulgaris L. | 0.480 | 0.060 | Glechoma hederacea L. | 0.552 | 0.722 |
| Dianthus deltoides L. | 0.381 | 0.017 | Ranunculus acris L. | 0.591 | 0.222 | Lamium maculatum (L.) L. | 0.177 | 0.286 |
| Festuca pratensis Huds. | 0.638 | 0.067 | Ranunculus polyanthemos L. | 0.048 | 0.006 | Lathyrus vernus (L.) Bernh. | 0.029 | 0.045 |
| Filipendula ulmaria (L.) Maxim. | 0.577 | 0.278 | Rumex acetosa L. | 0.489 | 0.200 | Lonicera xylosteum L. | 0.022 | 0.042 |
| Fragaria vesca L. | 0.600 | 0.169 | Stellaria graminea L. | 0.490 | 0.257 | Matteuccia struthiopteris (L.) Tod. | 0.144 | 0.392 |
| Galium mollugo L. | 0.625 | 0.351 | Tanacetum vulgare L. | 0.226 | 0.043 | Mercurialis perennis L. | 0.287 | 0.469 |
| Gentianella amarella (L.) Harry Sm. | 0.226 | 0.040 | Taraxacum officinale (L.) Weber ex F.H. Wigg. aggr. | 0.367 | 0.012 | Molinia caerulea (L.) Moench | 0.019 | 0.027 |
| Geranium pratense L. | 0.579 | 0.215 | Trifolium montanum L. | 0.332 | 0.048 | Picea × fennica (Regel) Kom. | 0.115 | 0.310 |
| Geum rivale L. | 0.499 | 0.318 | Trifolium pratense L. | 0.540 | 0.010 | Pulmonaria obscura Dumort. | 0.248 | 0.361 |
| Helictotrichon pubescens (Huds.) Schult. & Schult. f. | 0.226 | 0.048 | Trollius europaeus L. | 0.400 | 0.248 | Sorbus aucuparia L. | 0.132 | 0.239 |
| Heracleum sphondylium subsp. sibiricum (L.) Simonk. | 0.551 | 0.168 | Valeriana officinalis L. | 0.470 | 0.053 | Stellaria holostea L. | 0.349 | 0.708 |
| Pilosella caespitosa (Dumort.) P.D.Sell & C.West | 0.259 | 0.028 | Veronica chamaedrys L. | 0.637 | 0.361 | Tilia cordata Mill. | 0.290 | 0.528 |
| Hypericum maculatum Crantz | 0.456 | 0.156 | Vicia sepium L. | 0.405 | 0.292 | Vicia sylvatica L. | 0.062 | 0.324 |
ЗАКЛЮЧЕНИЕ
В условиях заповедника после прекращения сенокошения сокращаются площадь и состав луговых сообществ, что выявляется при анализе данных регулярной лесной таксации, материалов космической съемки и подтверждается данными наземных наблюдений. Картографический анализ данных показал, что характер экспансии лесных растений на луга зависит от исходной площади лугового участка, его формы и расположения в лесном массиве. Участки, расположенные в пограничных кварталах, частично сохраняют луговой статус благодаря проявлению краевого эффекта вблизи опушки леса. Вероятностная оценка изменения NDVI свидетельствует о достоверной смене состава и продуктивности луговых сообществ. Наличие флористических списков сосудистых растений Раифского участка заповедника разных лет позволяет использовать их в качестве исторических пулов видов для оценки динамики растительного покрова. Сравнение по ним таксономической структуры флоры луговой фракции убеждает в относительной стабильности видового разнообразия в первой половине ХХ века, до создания заповедника, и его увеличения в условиях заповедного режима при продолжающемся сенокошении на луговых участках. Прекращение использования лугов как сенокосов привело к зарастанию луговых участков, выпадению светолюбивых видов растений и увеличению роли лесных видов. Наиболее информативным показателем протекающих процессов являются вероятностные оценки потенциального присутствия видов в целевом сообществе, поступающих из локального пула видов.
Список литературы
[Atsekhovskii et al.] Ацеховский А.Р., Бочкарева С.Г., Рогова Т.В. 1985. Флора луговых формаций Раифы и проблемы ее охраны. – В сб.: Тезисы докладов и сообщений к республиканской научно-технической конференции, посвященной 25-летию ВКГЗ: “Охрана и исследования заповедных экосистем Волжско-Камского края”. Казань. С. 66–68.
Beals E.W. 1984. Bray-Curtis-ordination: an effective strategy for analysis of multivariate ecological data. – Adv. Ecol. Res. 14: 1–55.
[Buntova et al.] Бунтова О.Ю., Мухарамова С.С., Шайхутдинова Г.А. 2016. Оценка динамики лесного покрова по данным дистанционного зонирования Земли. – В сб.: Труды Волжско-Камского гос. природ. биосфер. заповедника. Вып. 7. Казань. С. 199–206.
Chytrý M., Hennekens S.M., Jiménez-Alfaro B., Knollová I., Dengler J., Jansen F., Landucci F., Schaminée J.H., Aćić S., Agrillo E., Ambarli D., Angelini P., Apostolova I., Attorre F., Berg C., Bergmeier E., Biurrun I., Botta-Dukát Z., Brisse H., Campos J. A., Carlón L., Čarni A., Casella L., Csiky J., Ćušterevska R., Dajić Stevanović Z., Danihelka J., De Bie E., de Ruffray P., De Sanctis M., Dickoré W.B., Dimopoulos P., Dubyna D., Dziuba T., Ejrnæs R., Ermakov N., Ewald J., Fanelli G., Fernández-González F., Fitzpatrick Ú., Font X., García-Mijangos I., Gavilán R.G., Golub V., Guarino R., Haveman R., Indreica A., Işik Gürsoy D., Jandt U., Janssen J.A., Jiroušek M., Kacki Z., Kavgaci A., Kleikamp M., Kolomiychuk V., Krstivojević Ćuk M., Krstonošić D., Kuzemko A., Lenoir J., Lysenko T., Marcenò C., Martynenko V., Michalcová D., Moeslund J.E., Onyshchenko V., Pedashenko H., Pérez-Haase A., Peterka T., Rašomavičius V., Rodríguez-Rojo M.P., Rodwell J.S., Rogova T., Ruprecht E., Rūsiņa S., Seidler G., Šibík J., Šilc U., Škvorc Z., Sopotlieva D., Stančić Z., Svenning J.-C., Swacha G., Tsiripidis I., Turtureanu P.D., Uğurlu E., Uogintas D., Valachovič M., Vashenyak Y., Vassilev K., Venanzoni R., Virtanen R., Weekes L., Willner W., Wohlgemuth T., Yamalov S. 2016. European Vegetation Archive (EVA): An integrated database of European vegetation plots. – Appl. Veget. Sci. 19 (1): 173–180. https://doi.org/10.1111/avsc.12191
De Cáceres M., Legendre P. 2008. Beals smoothing revisited. – Oecologia. 156: 657–669. https://doi.org/10.1007/s00442-008-1017-y
Erikson O. 1993. The species-pool hypothesis and plant community diversity. – Oikos. 68: 371–474.
[Garanina] Гаранина И.И. 1968. Некоторые предварительные итоги изучения флоры Раифского участка Волжско-Камского заповедника. – В сб.: Труды Волжско-Камского заповедника. Вып. 1. Казань. С. 60–68.
Giarrizzo E., Burrascano S., Chiti T., de Bello F., Leps J., Zavattero L., Blasi C. 2017. Re-visiting historical semi-natural grasslands in the Apennines to assess patterns of changes in species composition and functional traits. – Appl. Veget. Sci. 20: 247–258. https://doi.org/10.1111/avsc.12288
Giarrizzo E., Burrascano S., Zavattero L., Blasi C. 2015. New methodological insights for the assessment of temporal changes in semi-natural dry grasslands plant species composition based on field data from the northern Apennines. – Hacquetia. 14 (1): 19–32. https://doi.org/10.1515/hacq-2015-0014
[Gordyagin] Гордягин А. 1889. Ботанико-географические исследования в Казанском и Лаишевском уездах. – Тр. об-ва естествоиспытателей при Имп. Казанском ун-те. 22 (2): 1–92.
Juice version 7.0 http://www.sci.muni.cz/botany/juice
[Korzhinskii] Коржинский С. 1888. Северная граница черноземно-степной области восточной полосы Европейской России в ботанико-географическом и почвенном отношении. 1. Введение. Ботанико-географический очерк Казанской губернии. – Тр. об-ва естествоиспытателей при Имп. Казанском ун-те. 18 (5): 1–256.
[Lyubina, Rogova] Любина О.Е., Рогова Т.В. 2008. Оценка биоразнообразия растительного покрова Раифского участка ВКГПБЗ с позиций концепции пула видов. – Учен. зап. Казанского гос. ун-та. Сер. естественные науки. 150 (3): 203–218.
Mucina L., Bültmann H., Dierßen K., Theurillat J.-P., Raus T., Čarni A., Šumberová K., Willner W., Dengler J., García R.G., Chytrý M., Hájek M., Di Pietro R., Iakushenko D., Pallas J., Daniëls F.J.A., Bergmeier E., Santos-Guerra A., Ermakov N., Valachovič M., Schaminée J.H.J., Lysenko T., Didukh Ya.P., Pignatti S., Rodwell J.S., Capelo J., Weber H.E., Solomeshch A., Dimopoulos P., Aguiar C., Hennekens S.M., Tichý L. 2016. Vegetation of Europe: hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. – Appl. Veget. Sci. 19 (1): 3–783. https://doi.org/10.1111/avsc.12257
Oksanen J., Guillaume Blanchet F., Friendly M., Kindt R., Legendre R., McGlinn D., Minchin P.R., O’Hara R.B., Simpson G.L., Solymos P., Stevens M.H.H., Szoecs E., Wagner H. 2017. vegan: Community Ecology Package. R package version 2.4-5. https://CRAN.R-project.org/package=vegan.
Pärtel M., Zobel M., Zobel K., van der Maarel E. 1996. The species pool and its relation to species richness: evidence from Estonian plant communities. – Oikos 75: 111–117.
Prokhorov V.E., Rogova T.V., Kozhevnikova M.V. 2017. Vegetation Database of Tatarstan. – Phytocoenologia. 47 (3): 309–313. https://doi.org/10.1127/phyto/2017/0172
[Rogova et al.] Рогова Т.В., Прохоров В.Е., Шайхутдинова Г.А., Шагиев Б.Р. 2010. Электронные базы фитоиндикационных данных в системах оценки состояния природных экосистем и ведения кадастров биоразнообразия. – Учен. зап. Казанского ун-та. Сер. естественные науки. 152 (1): 174–181.
Song C., Woodcock C.E., Seto K.C., Lenney M.P., Macomber S.A. 2001. Classification and change detection using Landsat TM data: When and how to correct atmospheric effects? – Remote Sensing of Environment. 75: 231–232. https://doi.org/10.1016/S0034-4257(00)00169-3
[Spisok…] 1968. Список сосудистых растений Раифского леса, составленный Л.Н. Васильевой и А.Д. Плетневой-Соколовой (по материалам исследований 1925 г.), дополненный данными А.П. Ильинского (1943 г.). – В сб.: Тр. Волжско-Камского гос. заповедника. Вып. 1. Казань. С. 40–59.
Taylor D.R., Aarsen L.W., Loehle C. 1990. On the relationship between r/K selection and environmental carrying capacity: a new habitat template for plant life history strategies. – Oikos 58: 239–250.
[Tolmachev] Толмачев А. И. 1974. Введение в географию растений. Л. 244 с.
Van der Maarel E., Sykes M.T. 1993. Small-scale plant species turnover in a limestone grassland – the Carousel model and some comments on the niche concept. – J. Veget. Sci. 4: 179–188. https://doi.org/10.2307/3236103
[Yurtsev] Юрцев Б.А. 1987. Флора как базовое понятие флористики: содержание понятия, подходы к изучению. – В кн.: Теоретические и методические проблемы современной флористики. Л. С. 13–28.
Zobel M., van der Maarel E., Dupre C. 1998. Species pool: the concept, its determination and significance for community restoration. – Appl. Veget. Sci. 1: 55–66. https://doi.org/10.2307/1479085
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал