

Ботанический журнал, 2020, T. 105, № 1, стр. 46-57
НОВЫЕ ВИДЫ ПОКРЫТОСЕМЕННЫХ ИЗ ПАЛЕОГЕНОВЫХ ОТЛОЖЕНИЙ ЗАЙСАНСКОЙ ВПАДИНЫ (ВОСТОЧНЫЙ КАЗАХСТАН)
А. Л. Аверьянова 1, *, Син Яову 2
1 Ботанический институт им. В.Л. Комарова РАН
197376 г. Санкт-Петербург, ул. Проф. Попова, 2, Россия
2 Тропический ботанический сад Сишуанбаньна (АН КНР)
666303 Менлунь, префектура Мэнгла, КНР
* E-mail: AAveryanova@binran.ru
Поступила в редакцию 15.05.2019
После доработки 02.12.2019
Принята к публикации 03.12.2019
Аннотация
Описаны новые виды покрытосеменных Alnus chaybulakensis sp. nov., Quercus borissovii sp. nov. и Hemiptelea kryshtofovichii sp. nov. из позднеэоценовой и раннеолигоценовой флор Зайсанской впадины (Восточный Казахстан), из них для двух последних видов впервые представлены валидизирующие описания. Эти таксоны входили в число теплоумеренных листопадных элементов палеофлоры, занявших на границе эоцена и олигоцена доминирующее положение, а затем, в олигоцене, полностью вытеснивших вечнозеленую растительность впадины.
ВВЕДЕНИЕ
Зайсанская впадина (Восточный Казахстан) представляет собой межгорный прогиб, ограниченный системами Южного Алтая на севере и Саур-Тарбагатая на юге, выполненный континентальными кайнозойскими отложениями (рис. 1). В палеогене и неогене на территории Джунгарской депрессии, частью которой является впадина, располагалось обширное Гобийское озеро; в результате его чередующихся ингрессий и регрессий образовалась толща осадочных отложений общей мощностью более 2000 м (Borisov, 1983), заключающих в себе разнообразный флористический и фаунистический ископаемый материал.
Рис. 1.
Расположение местонахождений ископаемых растений в Зайсанской впадине (по: Borisov, 1963, с изменениями).
1 – палеозойские отложения; 2 – палеоцен-миоценовые отложения; 3 – плиоценовые отложения; 4 – местонахождения ископаемых растений.
Fig. 1. Map of fossil floras localities in Zaissan Depression (according to: Borisov, 1963, modified).
1 – Paleozoic; 2 – Paleocene-Miocene; 3 – Pliocene; 4 – plant fossils localities.
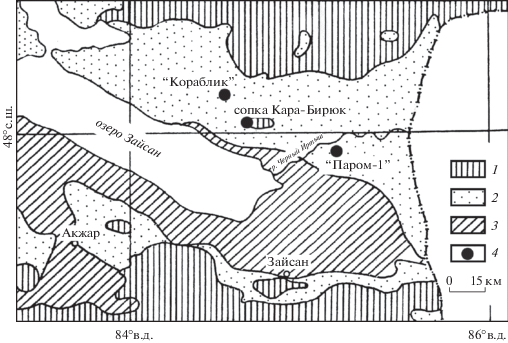
Насыщенность разрезов ископаемым материалом хорошей сохранности позволила определить систематический состав ряда локальных палеогеновых флор и характеристики климата впадины, при котором они существовали (Iljinskaja, 1986; Averyanova, 2014). На границе эоцена и олигоцена Зайсанская впадина входила в экотонную зону, проходившую по границе Тетисовой и Бореальной палеогеновых областей (Akhmetiev, 1993). Этим объясняется одновременное присутствие в пограничных локальных флорах как теплолюбивых вечнозеленых (Dryophyllum Debey, Lauraceae и др.), так и теплоумеренных таксонов (Salicaceae, Betulaceae, Fagaceae).
Ревизия систематических списков для раннего олигоцена (Akhmetiev, 1991; Iljinskaja, 1991) выявила два таксона из местонахождения на г. Киин-Кериш (кустовская свита), не имевшие описаний и не опубликованные валидно. В данной статье мы публикуем описания этих двух видов. Также приводится описание ископаемой ольхи из чайбулакской (местонахождение “Паром-1”) и среднеаксыирской подсвиты аксыирской свиты (г. Кара-Бирюк).
МАТЕРИАЛ И МЕТОДЫ
Проведенное исследование базируется на палеоботанических коллекциях из верхнего эоцена–нижнего олигоцена, собранных с 1927 по 1965 г. Б.А. Борисовым, В.К. Василенко, В.С. Ерофеевым, И.А. Ильинской, А.И. Москвитиным и М.Ф. Нейбург, и хранящихся в Лаборатории палеоботаники Ботанического института им. В.Л. Комарова РАН (Санкт-Петербург). Растительные остатки представлены преимущественно отпечатками листьев, в единичных случаях встречаются отпечатки облиственных побегов и репродуктивных органов; расположение отпечатков преимущественно в одной плоскости и хорошая сохранность свидетельствуют о захоронении вблизи места произрастания, без значительного переноса.
Сравнительно-морфологическое исследование как отпечатков, так и гербарного материала проводилось с привлечением стереоскопических микроскопов моделей МБС-2 и МС-2 при увеличении ×5–15 раз. Диагностические признаки документировались рисунками, сделанными по стандартной методике прорисовки снимка, и фотографиями (фотокамера Canon EOS 60D).
СТРАТИГРАФИЯ И ВОЗРАСТ ФЛОРОНОСНЫХ СЛОЕВ
Представление о стратиграфических границах и датировке палеогеновых свит Зайсанской впадины основаны на данных региональных стратиграфических и палеомагнитных исследований (Borisov, 1963, 1983; Suleimanova, 1999), дополненных выводами о возрасте ископаемых флоры и фауны (Gabunia et al., 1983; Iljinskaja et al., 1983; Gabunia, 1984; Iljinskaja, 1986). Согласно утвержденной стратиграфической схеме данного региона (Yakhimovich et al., 1993), отложения с ископаемыми растениями, в том числе с описанными ниже видами, входят в состав восточно-зайсанской серии по Б.А. Борисову (Borisov, 1963), охватывающей верхний эоцен (чайбулакская, кызылкаинская и нижняя подсвита аксыирской свиты) и весь олигоцен (средняя и верхняя подсвита аксыирской свиты, кустовская и буранская свиты) (табл. 1).
Таблица 1.
Стратиграфическое расчленение верхнеэоценовых-олигоценовых отложений Зайсанской впадины и положение местонахождений ископаемых растений (по: Borisov, 1963; Yachimovich et al., 1993) Table 1. Stratigraphic scheme of the Upper Eocene-Oligocene sediments of Zaissan Depression and fossil floras localities (according to: Borisov, 1963; Yachimovich et al., 1993)
| Стратиграфическая
единица Stratigraphical unit |
Серия/Series | Свита и подсвита,
суммарная мощность
отложений, м Formation and subformation, overall width, m |
Региональная
ортозона Regional orthozone |
Преобладающие
породы Dominant rocks |
Местонахождения ископаемых растений и их координаты Localities of fossil plants and their coordinates |
Номер коллекции
БИН РАН Number of the collection BIN RAS |
|
|---|---|---|---|---|---|---|---|
| Олигоцен/Oligocene Верхний эоцен/Upper Eocene |
Восточно-зайсанская/East-Zaissan | Буранская Buran (19.6) | r-Шиликты (r-Shilikty) | пески, алевриты sands, aleurites |
|||
| Кустовская Kusto (45.1) | r-Акколкан r-Akkolkan |
алевриты,
тонкозернистые пески aleurites, fine- grained sands |
г. Киин-Кериш, “Кораблик” Kiin-Kerish Mt., “Korablick” (47°59′ N, 84°47′ E) |
999, 999А | |||
| Аксыирская/Aksyir (74) | Верхняя подсвита Upper subformation |
глины, алевриты clays, aleurites |
|||||
| Средняя подсвита Middle subformation |
n-Калкан n-Kalkan |
сопка Кара-Бирюк Kara-Biruck Mt. (48°12′ N, 84°51′ E) |
987 | ||||
| пограничный
интервал border layers |
Нижняя подсвита Lower subformation |
r-Онучтениз r-Onuchteniz |
|||||
| Верхний эоцен Upper Eocene |
n-Аккум n-Akkum |
||||||
| Кызылкаинская Kysylkain (18.6) |
r-Аиршоны n-Сарыкум |
пески, алевриты sands, aleurites |
|||||
| r-Теректы r-Airshony |
|||||||
| n-Sarykum r-Terekty |
|||||||
| Чайбулакская Chaibulak (15.5) |
nr-Саржира nr-Sarzhira |
алевриты, глины, пески aleurites, clays, sands |
“Паром-1” “Parom-1” (47°59′ N, 84°47′ E) |
989 | |||
Чайбулакская свита (обнажена в местонахождении “Паром-1”) в магнитостратиграфической схеме Зайсанской впадины (Suleimanova, 1999) целиком охватывается ортозоной nr-Саржира, коррелированной с хроном С17n (Berggren et al., 1995), лежащим в основании позднего эоцена. Миоспоровый комплекс чайбулакской свиты сопоставляется Л.А. Пановой (Panova, 1965) с комплексами кумского горизонта (средний–поздний эоцен) Прикаспия; чайбулакская палинофлора характеризуется ею как разнообразная в систематическом отношении и, при этом, резко отличная от спорово-пыльцевых комплексов из нижележащих отложений. Среднеэоценовая макрофлора Зайсанской впадины также очень заметно отличается от чайбулакской.
Граница эоцена и олигоцена в Зайсанской впадине, по мнению большинства исследователей (Yakhimovich et al.., 1993; Krasheninnikov et al., 1998), основанном на магнитостратиграфических исследованиях, а также на данных по ископаемым растениям и позвоночным, находится в интервале от верхов нижне- до низов среднеаксыирской подсвит. Она пролегает в нижней трети ортозоны n-Калкан, вблизи ее границы с нижележащей ортозоной r-Онучтениз. Последняя соответствует началу хрона С13r мировой шкалы магнитных аномалий, в пределах которого проводится граница верхнего эоцена–нижнего олигоцена (стратотип Массиньяно; Premoli-Silva, Jenkins, 1993). Таким образом, флороносный горизонт сопки Кара-Бирюк (нижняя часть средней подсвиты аксыирской свиты, ортозона n-Калкан) мы относим к пограничным эоцен-олигоценовым слоям, вероятнее, к верхнему эоцену. Последнее подтверждается присутствием в ископаемой флоре этого местонахождения представителей Dryophyllum, рода, характерного для верхнего эоцена впадины и полностью отсутствующего в более поздних отложениях.
Магнитостратиграфические исследования кустовской свиты (Suleimanova, 1999) показали, что последняя, а также верхнеаксыирская подсвита аксыирской свиты составляют единый ортохрон r-Акколкан, сопоставляемый с хроном С12r мировой шкалы (ранний олигоцен). В.М. Чхиквадзе (Chkhikvadze, 1973) отмечает большое сходство представителей герпетофауны из этой свиты с олигоценовыми видами соседних с Зайсанской впадиной регионов. Палеофлора кустовской свиты (Akhmetiev, 1991; Iljinskaja, 1991; собственные наблюдения автора) резко отличается по систематическому составу как от эоценовых, так и от миоценовых зайсанских ископаемых флор. На основании этих данных для кустовской свиты нами принимается раннеолигоценовый возраст (рюпель).
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ОТДЕЛ MAGNOLIOPHYTA
КЛАСС MAGNOLIOPSIDA
Сем. FAGACEAE
Род Quercus L., 1753
Quercus borissovii Iljinskaja ex Averyanova, sp. nov.
Табл. I, 1–3
Таблица I. 1–3 – Quercus borissovii Iljinskaja ex Averyanova, sp. nov.: 1, 2 – экз. 322, 3 – край листа; кол. 999А [БИН], Восточный Казахстан, Зайсанская впадина, гора Киин-Кериш, флора останца “Кораблик”, кустовская свита, нижний олигоцен.
4 – Quercus aegilops L., современный лист. [БИН] Албания, 4 XII 1955, коллектор: А.Л. Тахтаджян.
5, 6 – Alnus japonica Sieb. et Zucc., современный лист. [БИН] Japan, 22 II 1913, № 46382, коллектор: T. Makino: 5 – общий вид листа, 6 – край листовой пластинки.
7–12 – Hemiptelea kryshtofovichii Iljinskaja ex Averyanova, sp. nov.: 7, 8 – экз. 3, 9 – он же, край листовой пластинки; 10, 11 – экз. 50; кол. 999А [БИН]; 12 – экз. 231; кол. 4337 [БИН], Восточный Казахстан, Зайсанская впадина, гора Киин-Кериш, флора останца “Кораблик”, кустовская свита, нижний олигоцен.
Масштабная линейка – 1 см.
Plate I . 1–3 – Quercus borissovii Iljinskaja ex Averyanova, sp. nov.: 1, 2 – specimen 322, 3 – leaf margin; coll. 999А [BIN], Eastern Kazakhstan, Zaissan Depression, Kiin-Kerish Mt., flora of “Korablik” remnant, Lower Oligocene, Kusto Formation.
4 – Quercus aegilops L., leaf (modern species). [BIN] Albania, 4 XII 1955, coll. А.L. Takhtajan.
5, 6 – Alnus japonica Sieb. et Zucc., leaf (modern species). [BIN] Japan, 22 II 1913, № 46382, coll. T. Makino: 5 – leaf, general view, 6 – detail of leaf margin.
7–12 – Hemiptelea kryshtofovichii Iljinskaja ex Averyanova, sp. nov.: 7, 8 – specimen 3, 9 – specimen 3, detail of leaf margin; 10, 11 – specimen 50; coll. 999А [BIN]; 12 – specimen 231; coll. 4337 [BIN], Eastern Kazakhstan, Zaissan Depression, Kiin-Kerish Mt., flora of “Korablik” remnant, Kusto Formation, Lower Oligocene.
Scale bar – 1 cm.

Quercus borissovii Iljinskaja in Ахметьев, 1991, Формирование эоценово-миоценовой флоры Казахстана и Русской равнины: 41, nom. nud.
Название вида дано в честь Бориса Александровича Борисова, геолога, в течение многих лет работавшего в Зайсанской впадине, автора стратиграфической схемы этой территории.
Holotype: leaf impression, specimen No 322, collection No 999A [BIN]; Eastern Kazakhstan, Zaissan Depression, Kiin-Kerish Mountain, flora of locality “Korablik”, Lower Oligocene, Kusto Formation. – Plate I , 1.
Description. Leaves elliptic-ovate, 5.5–8 cm long and to 2.7–4 cm wide with slightly cordate base. Margin coarsely toothed, teeth obtuse with short acute apex, apical side convex, rarely straight, basal side long, convex, occasionally with small additional teeth; sinuses rounded, rarely angular. Midvein up to 1 mm in leaf base, thinning up to the apex. Secondary veins in 5–6 pairs, straight in the lower part of the leaf, slightly arcuate in the upper part, craspedodromous. Angles of divergence of secondary veins decrease towards the leaf apex. Tertiary veins sinuous or slightly curved, diverge from secondary veins obliquely, or at a right angle, often dichotomous.
Голотип: отпечаток листа, экз. 322, кол. № 999А [БИН]; Восточный Казахстан, Зайсанская впадина, гора Киин-Кериш, флора останца “Кораблик”, нижний олигоцен, кустовская свита. – Табл. I, 1.
Описание. Листья овальные или овально-яйцевидные, 5.5–8 см дл. и 2.7–4 см шир. (табл. I, 1, 2). Основание слабосердцевидное. Край крупнозубчатый с единичными дополнительными зубцами. Зубцы низкие куполообразные, с прямой или выпуклой апикальной, и заметно более длинной выпуклой базальной стороной, с коротким толстым остроконечием на верхушке. Выемки между зубцами округлые, в единичном случае – угловатые. Главная жилка почти прямая, около 1 мм толщиной в основании, почти прямая, слегка извилистая, к верхушке истончается. Вторичные жилки в числе 5–6 пар, в нижней половине листа прямые, в верхней – слегка дуговидные, редко слегка отогнутые у края листа, нижняя пара жилок развернута почти на 180°, две следующие – на 90°, а в верхушке листа – на 60°. Третичные жилки лестничные и ветвисто-лестничные, редкие, 3–4 на 1 см, отходят от вторичных косо, реже под прямым углом, извилистые или коленчато изогнутые, нередко дихотомирующие (табл. I, 3).
Сравнение. Вид не обнаруживает сходства ни с одним из известных ископаемых дубов. Из современных видов к Quercus borissovii близок средиземноморский переднеазиатский Q. aegilops L. (табл. I, 4), от которого описываемый вид отличается слабой извилистостью главной жилки, очень короткими щетинками зубцов и дуговидно изогнутыми вторичными жилками в верхней половине листа. Подобная дуговидность наблюдается, однако, у подвида Q. aegilops ssp. ithaburensis (Decaisne) Eig. Этот засухоустойчивый листопадный дуб может встречаться одиночно в составе сосновых или платановых лесов, но чаще образует дубравы на низких высотах (до 500–1000 м над уровнем моря) (Menitsky, 1984). С упомянутым современным видом И.А. Ильинская (Iljinskaja, 1982) сближает ископаемый Q. ussuriensis Krysht.; однако, последний резко отличается краем с крупными лопастевидными зубцами, тогда как у листьев Q. borissovii зубцы низкие.
Распространение. Зайсанская впадина, Восточный Казахстан, кустовская свита, нижний олигоцен.
Материал. Голотип и кол. 999, экз. 38 с противоотпечатком 38А, кол. 999А, экз. 78, местонахождение “Кораблик”, кустовская свита.
Представители рода Quercus, достоверно известные на территории бывшего СССР с начала палеоцена (Fossil…, 1982), в Зайсанской впадине впервые зафиксированы в нижнем олигоцене, где составили, наряду с видами Castanopsis (D.Don) Spach, самую обильно представленную группу растений. Здесь они пришли на смену каштанодубам (род Dryophyllum Debey) – характерным для позднего эоцена вечнозеленым Fagaceae, полностью исчезнувшим к началу олигоцена. Последние, возможно, стояли в основании олигоценового разнообразия буковых на территории впадины, хотя нельзя исключить, что они представляли собой тупиковую ветвь эволюции, освободившую экологическую нишу для дубов и каштанов при смене климатических условий.
Сем. Betulaceae
Род Alnus Mill., 1754
Alnus chaybulakensis Averyanova, sp. nov.
Табл. II, 1–7
Таблица II. 1–7 – Alnus chaybulakensis Averyanova, sp. nov.: 1–3 – голотип экз. 19: 1, 2 – общий вид листа, 3 – верхушка листа; 4, 5 – экз. 18б; 6 – экз. 45; 7 – экз. 18а, деталь края листа; кол. 989 [БИН], Восточный Казахстан, Зайсанская впадина, местонахождение “Паром-1”; чайбулакская свита, верхний эоцен.
Масштабная линейка – 1 см.
Plate II . 1–7 – Alnus chaybulakensis Averyanova, sp. nov.: 1–3 – holotype, specimen 19: 1, 2 – leaf, general view, 3 – leaf apex; 4, 5 – specimen 18b; 6 – specimen 45; 7 – specimen 18а, detail of leaf margin; coll. 989 [BIN], Eastern Kazakhstan, Zaissan Depression, locality “Parom-1”; Сhaybulak Formation, Upper Eocene.
Scale bar – 1 cm.

Название вида – от латинской транслитерации чайбулакской свиты, к которой относится типовое местонахождение описываемого вида.
Holotype: leaf impression, specimen 19, collection 989 [BIN]; Eastern Kazakhstan, Zaissan depression, locality “Parom-1”, Upper Eocene, Сhaybulak Formation. – Plate II , 1.
Description. Leaves oblong-elliptical, 6–11 сm long and 2.7–5 сm wide. Base rounded-cuneate. Apex gradually narrowing, attenuate. Leaf margin, except apex, dentate. Teeth usually 2 per 1 сm of the margin, up to 1 mm high, acute, with 1–2 intermediate usually smaller teeth. Midvein straight, gradually thinning toward apex. Secondary veins 10–13 pairs, slightly upcurving, opposite or subopposite, 2–3 times thinner than midvein, ending craspedodromously, with conspicuous straight branching near the leaf margin, connected with tertiary vein from upper secondary vein; from these connection points the branchlets diverge into intermediate teeth. Infrabasal veins thin, connecting with secondary veins by arcuate loops. Tertiary veins 3–5 per 1 сm, distinct, diverging nearly at a right angle from the secondary veins, straight or slightly geniculately bending, sometimes dichotomous. Petiole up to 3 сm long, near 1 mm thick.
Голотип: отпечаток листа, экз. 19, кол. 989 [БИН]; Восточный Казахстан, Зайсанская впадина, местонахождение “Паром-1”, верхний эоцен, чайбулакская свита. – Табл. II, 1.
Описание. Листья продолговатоэллиптические, 6–11 см длины, 2.7–5 см ширины (табл. II, 1, 2). Верхушка постепенно сужающаяся, оттянутая (табл. II, 3). Основание ширококлиновидное (табл. II, 4, 5). Край листа, за исключением самого кончика, зубчатый; зубцы, в которых кончаются вторичные жилки, в числе около 2 на 1 см края, в верхушке более частые, направленные вверх, до 1 мм высотой, с прямой апикальной стороной и прямой или слегка вогнутой базальной стороной (табл. II, 6). Между этими зубцами имеются 1–2 более мелких, реже равных им по размеру зубчиков. Главная жилка около 0.8 мм толщиной в основании листа, почти прямая, в верхушке истончается в 3–4 раза. Вторичные жилки в числе 10–13 пар, слегка или заметно дуговидные. Нижние из них супротивные или попарно сближенные, в 2–3 раза тоньше главной, отходят от нее через интервал 0.8–1.3 см под углом 40–50° (в верхушке через 0.3–0.7 см и под 25–35°) и оканчиваются в зубцах краспедодромно, давая у края вверх прямое ответвление, обычно параллельное краю. Ответвление это соединяется под тупым углом с третичной жилкой, отходящей от вышележащей вторичной жилки; из вершины угла отходит прямая или слегка дуговидная веточка в промежуточный зубчик. На экз. 18а (кол. 989) одна из вторичных жилок ветвится в 1 см от края (табл. II, 4, 5). В основании листовой пластинки имеется пара тонких жилок, соединяющихся анастомозами с нижними вторичными жилками. Третичные жилки в количестве 3–5 на 1 см, хорошо заметные, прямые или слегка коленчато изогнутые, иногда дихотомирующие, отходят перпендикулярно, реже косо ко вторичным. Жилки четвертого порядка отходят перпендикулярно третичным, местами параллельные, а местами в середине интервала коленчато соединяются или образуют сеть из мелких 4–5-угольных ячеек, а местами – изломанную жилку, параллельную третичным, проходящую в середине интервала (табл. II, 7). Черешок до 3 см длины.
Сравнение. Новый вид по характеру зубчатости близок к листьям Alnus neuburgiae (Baik.) Iljinskaja, описанным из раннеолигоценовой флоры г. Ашутас Зайсанской впадины и известным здесь также из позднего эоцена (г. Киин-Кериш, флора “Василенко”) и раннего олигоцена (флора останца “Кораблик”) (Iljinskaja, 1986), а за пределами Зайсанской впадины обычным, по мнению Б.И. Павлюткина (Pavlytkin, 2002, 2005), в среднем (ханкайская флора) и позднем (усть-суйфунская флора) миоцене Приморья. Отличаются листья A. chaybulakensis от последнего вида более узкой и продолговатой формой всего листа и его основания, а также постепенно сужающейся оттянутой верхушкой, в то время как у A. neuburgiae верхушка сужается в оттянутый кончик, основание листа широкое, округлое до слабосердцевидного. Среди отпечатков A. neuburgiae из флоры “Кораблик” г. Киин-Кериш иногда встречаются узкие листья с клиновидным основанием, сходным с сохранившемся у одного из наших листьев (экз. 18б, кол. 989), а также, как исключение – с оттянутой верхушкой, однако у них вторичные жилки редкие и не сближенные при основании.
От характерной для олигоцена Зайсанской впадины A. schmalhausenii Grub. (Kryshtofovich et al., 1956) описываемый вид отличается разнозубчатостью и меньшим количеством промежуточных зубцов, которых у первого вида может быть до семи в одном интервале (Baikovskaja, 1974), обычно более заметными третичными жилками; кроме того, у листьев A. schmalhausenii часто присутствуют базископические ответвления от вторичных жилок, не характерные для A. chaibulakensis.
Из современных представителей рода к A. chaybulakensis наиболее близки виды из секции Haplostachys Czer.; С.К. Черепанов (Czerepanov, 1954) выделял последнюю как наиболее примитивную в пределах рода. Наибольшее сходство – по форме листа, оттянутой верхушке, характеру зубчатости и жилкования – наблюдается с A. japo-nica Sieb. et Zucc. (табл. I, 5, 6). У последнего вида, однако, вторичные жилки отходят под меньшим углом и входят в зубцы чаще апикально, а их число не превышает 7–8 пар. A. japonica распространена в теплоумеренных лесах Приморья, Кореи, Китая и Японии, предпочитает влажные местообитания на низких высотах.
Два других вида из того же ряда Japonicae секции Haplostachys, а именно, A. trabeculosa Hand.-Mazz. (Iljinskaja, 1991) и A. maritimа (Marsh.) Muhl., менее схожи с новым видом. Так, A. maritimа отличается от A. chaybulakensis притупленными зубцами, более узким основанием листа, верхушкой, оттянутой в короткий широкий кончик, и малым числом вторичных жилок, не превышающим девяти пар. Отличия описанного вида от A. trabeculosa те же, что и от A. neuburgiae.
Распространение. Чайбулакская свита и средняя подсвита аксыирской свиты, верхний эоцен, Зайсанская впадина, Восточный Казахстан.
Материал. Голотип и кол. 989, экз. 18а, 18б, 24, 45, 80, чайбулакская свита, местонахождение “Паром-1”; кол. 987, экз. 16, средняя подсвита аксыирской свиты, г. Кара-Бирюк.
Виды семейства Betulaceae (Alnus L., Betula L. и Corylus L.) заняли заметное положение в древней флоре впадины, начиная с раннего олигоцена. Первые же, еще немногочисленные, находки представителей этого семейства датируются серединой позднего эоцена. К ним относится и описанный вид Alnus. Все ископаемые ольхи из Зайсанской впадины, отличаясь деталями строения листа, имеют, тем не менее, достаточно близкие морфотипы листовой пластинки, которые характеризуются набором признаков (яйцевидная форма, мелкие, равные или почти равные по размерам зубцы, острая, часто оттянутая верхушка листа), типичных для современных представителей секции Haplostachys. Последняя включает, по мнению С.К. Черепанова (Czerepanov, 1954) наиболее древние примитивные виды рода.
Сем. ULMACEAE
Род Hemiptelea Planch., 1872
Hemiptelea kryshtofovichii Iljinskaja ex Averyanova, sp. nov.
Табл. I, 7–12
Hemiptelea kryshtofovichii Iljinskaja, 1986, Проблемы палеоботаники: 92, nom. nud.; Ахметьев, 1991, Формирование эоценово-миоценовой флоры Казахстана и Русской равнины: 41, nom. nud.
Название вида дано в честь Африкана Николаевича Криштофовича, палеоботаника, геолога, ботаника.
Holotype: leaf impression, specimen 3, collection 999A [BIN]; Eastern Kazakhstan, Zaissan Depression, Kiin-Kerish Mountain, flora of locality “Korablik”, Lower Oligocene, Kusto Formation. – Plate I , 7.
Description. Leaves 1.9–3.7 сm long, 0.9–1.3 сm wide, oblong to oblong-ovate. Base rounded, rounded-cuneate or slightly cordate. Apex gradually narrowing with obtise tip. Teeth with convex sides, somewhat overlap; basal side rarely with small tooth. The tooth at the leaf apex equal to or smaller than adjacent ones. Veins much more conspicuous on the lower leaf surface imprints. Midvein straight, thinning up to the apex. Secondary veins in 10–11 pairs, more or less straight, craspedodromous. Branchings going into sinuses are of the same thickness as tertiary veins. Intercalar veins sometimes are present. Tertiary veins very thin, diverge obliquely to the secondary veins, forming big meshes.
Голотип: отпечаток листа, экз. 3, кол. № 999А [БИН]; нижний олигоцен, кустовская свита; Восточный Казахстан, Зайсанская впадина, гора Киин-Кериш, флора останца “Kораблик”. – Табл. I, 7.
Описание. Листья продолговатые до продолговатояйцевидных, от 1.9 см длины при ширине 0.9 см до 3.7 и 2.6 см длины при ширине 1.3 см (табл. I, 7, 8, 10–12). Основание округлоклиновидное или округлое с небольшой выемкой в месте прикрепления черешка, или слабосердцевидное. Верхушка постепенно сужающаяся с притупленным кончиком. Зубец на верхушке листа обычно меньше соседних боковых, реже равен им. Зубцы с выпуклыми сторонами, единичные – с дополнительными зубчиками на базальной стороне, основания соседних зубцов слегка налегают друг на друга, верхушки зубцов расположены на уровне бухты, или же несколько ниже ее (табл. I, 9). Главная жилка тонкая, нитевидная на верхней и более заметная на нижней поверхности листа. Вторичные жилки слабо заметные на верхней поверхности листовой пластинки и относительно четкие на нижней, в числе 10–11 пар, прямые, оканчивающиеся в зубцах, развернуты в нижней части листа при округлоклиновидном основании на 100–110°, при округлом основании нижняя короткая пара развернута на 180°, а при слабосердцевидном – на 190°; в средней части листа они составляют угол 80–90°, в верхушке – 60–70°. Ответвления в бухту заметны на нижней поверхности листовой пластинки, по толщине не отличаются от третичных жилок. В широких интервалах встречаются единичные вставочные жилки, доходящие до дна бухты. Третичные жилки редкие, проходящие косо ко вторичным.
Сравнение. У рода Hemiptelea по остаткам листьев описаны два вида, из них H. iljinskiae Kutuzk. (Kutuzkina, 1973) из позднего сармата Краснодарского края не проявляет сходства с H. kryshtofovichii, а описание второго, H. paradavidii Chen из верхнего эоцена Сычуаня (Сhen et al., 1983), проиллюстрировано фотографиями, не позволяющими провести сравнение.
Листья описываемого вида очень сходны с листьями современной H. davidii (Hance) Planch. из монотипного рода; это листопадное дерево или кустарник, распространенный в Китае и Южной Корее, где произрастает на низменностях, и в предгорьях на высоте до 2000 м над уровнем моря (Schneider, 1906). Наблюдается также определенное сходство листьев H. kryshtofovichii с таковыми у Zelkova carpinifolia (Pall.) C. Koch (Losina-Losinskaja, 1951); отличает их от последней бóльшая в среднем вытянутость пластинки, постепенно сужающаяся верхушка с притупленным кончиком, зубцы, имеющие иногда дополнительный зубчик, верхушечный зубец, равный или меньший по размеру, чем соседние боковые, более многочисленные и частые вторичные жилки, отсутствие или слабая выраженность ответвлений в бухты, более тонкие третичные жилки.
Распространение. Кустовская свита, нижний олигоцен, Восточный Казахстан, Зайсанская впадина.
Материал. Кол. № 999А, экз. 3, 50, кол. № 4337, экз. 231а с противоотпечатком 231Аа, местонахождение “Кораблик”, кустовская свита.
Род Hemiptelea в ископаемом виде ранее был известен только из среднего–верхнего миоцена (Fossil…, 1982; Pavlyutkin, 2007). Таким образом, находка из Зайсанской впадины имеет самый древний возраст для территории СНГ. Интересно, что именно в пограничный эоцен-олигоценовый период во впадине уже присутствовали, наряду с родом Hemiptelea, два очень близких к нему рода подсемейства Ulmoideae – Zelkova Spach и Ulmus L. (Zavada, Kim, 1996), что свидетельствует об обособлении этих трех таксонов не позднее конца эоцена.
ЗАКЛЮЧЕНИЕ
Описанные виды входят в число теплоумеренных листопадных представителей палеофлоры Зайсанской впадины, роль которых по сравнению с более древними эпохами заметно возросла на границе эоцена и олигоцена. После закрытия меридионального Тургайского пролива и связанного с этим постепенного похолодания климата из зайсанских флор полтавского, по А.Н. Криштофовичу, экологического типа (Kryshtofovich, 1955) исчезли такие характерные эоценовые таксоны, как Broussonetia neuburgiae (Iljinskaja) Iljinskaja et Averyanova, Dryophyllum spp., Euptelea zaisanica Iljinskaja et Averyanova, Lindera vassilenkoi Iljinskaja, Ungnadia kryshtofovichii Iljinskaja и др. На смену им в начале олигоцена пришли теплоумеренные Comptonia acutiloba Brongn., Salicaceae, Betulaceae (Akhmetiev, 1991; Iljinskaja, 1991), а также особенно характерные для рюпельской флоры Зайсанской впадины многочисленные буковые. Последние подробно изучались И.А. Ильинской (Iljinskaja, 1991). Таким образом, к концу миоцена в Зайсанской впадине установилась мезофильная хвойно-широколиственная тургайская флора, в типичном виде известная из отложений г. Ашутас (Kryshtofovich et al., 1956).
Характерной особенностью флоры Зайсанской впадины, относящейся к промежутку от эоцена до раннего олигоцена, является высокая степень эндемичности, проявляющаяся порой даже на уровне отдельных местонахождений. Лишь в единичных случаях мы можем наблюдать преемственность таксонов; среди них можно упомянуть, например, Taxodium dubium (Sternb.) Heer, Periploca borissovii (Iljinskaja) Averyanova, роды Alnus, Ulmus, Zelkova, Populus. В то же время основу локальных флор составляют, как правило, виды, не встреченные за пределами конкретного местонахождения. Как отмечали еще В.С. Корнилова (Kornilova, 1966) и С.Г. Жилин (Zhilin, 1974), физико-географическая обстановка Призайсанья, очевидно, отличалась от остальной территории Казахстана. Это способствовало формированию и развитию в палеогене своеобразной растительности, не имеющей аналогов как на территории современного Казахстана, так и за ее пределами (Averyanova, 2012).
Подобная же картина постоянно меняющихся флор с неустойчивым систематическим составом наблюдалась в это время и на западе Казахстана (Akhmetiev, 2011), где связывалась с влиянием эвстатического фактора. Несомненно, это влияние сыграло роль и для Зайсанской впадины. Дальнейшее изучение зайсанских коллекций олигоценового и миоценового возраста сможет дать понимание динамики климата и эволюции растительного мира на данной территории вплоть до окончательного становления тургайского типа растительности в конце миоцена.
Список литературы
[Averyanova] Аверьянова А.Л. 2012. Позднеэоценовая флора Зайсанской впадины (Восточный Казахстан). Автореф. дис. … канд. биол. наук. СПб. 24 с.
Averyanova A.L. 2014. The climatic changes in Zaissan depression (Eastern Kazakhstan) on the Eocene-Oligocene border based on CLAMP-analysis. – In: Proceedings of the IPPC 2014. Padova. P. 9.
[Akhmetiev] Ахметьев М.А. 1991. Сравнение раннеолигоценовой флоры Киин-Кериша с другими палеогеновыми флорами. Формирование эоценово-миоценовой флоры Казахстана и Русской равнины. Л. С. 37–56.
[Akhmetiev] Ахметьев М.А. 1993. Фитостратиграфия континентальных отложений палеогена и миоцена Внетропической Азии. М. 142 с.
[Akhmetiev] Ахметьев М.А. 2011. Проблемы стратиграфии и палеогеографии палеогена средних широт Центральной Евразии. – Геология и геофизика. 52 (10): 1367–1387.
[Baikovskaja] Байковская Т.Н. 1974. Верхнемиоценовая флора Южного Приморья. Л. 196 с.
Berggren W.A., Kent D.V., Swisher C., Aubry M. 1995. A revised cenozoic Geochronology and chronostratigraphy. Special publication. 54: 129–212.
[Borisov] Борисов Б.А. 1963. Стратиграфия верхнего мела и палеоген-неогена Зайсанской впадины. Тр. ВСЕГЕИ, нов. сер. 94: 11–75.
[Borisov] Борисов Б.А. 1983. Биостратиграфия континентальных палеогеновых отложений Зайсанской впадины. Стратиграфия фанерозоя СССР. Л. С. 89–97.
Chen Ming-hong, Kong Zhao-chen, Chen Ye. 1983. On the discovery of the old Tertiary flora in western Sichuan Plateau and its significance. – Acta Bot. Sin. 25 (5): 482–491.
[Czerepanov] Черепанов С.К. 1954. Монография рода ольха – Alnus Mill. s.l. Дис. … канд. биол. наук. Л. 544 с.
[Chkhikvadze] Чхиквадзе В.М. 1973. Третичные черепахи Зайсанской котловины. Тбилиси. 100 с.
[Fossil…] Ископаемые цветковые растения СССР. Т. 2. Ulmaceae-Betulaceae. 1982. Л. 216 с.
[Gabunia] Габуния Л.К. 1984. Краткий обзор фаун палеогеновых млекопитающих Зайсанской впадины. – В кн.: Флора и фауна Зайсанской впадины. Тбилиси. С. 115–123.
[Gabunia et al.] Габуния Л.К., Гуреев А.А., Ефимов М.Б., Сычевская Е.К., Толстикова Н.В., Чхиквадзе В.М., Шевырева Н.С., Борисов Б.А. 1983. Фауна палеогена Зайсанской впадины. – В кн.: Стратиграфия фанерозоя СССР. Л. 98–115.
[Iljinskaja] Ильинская И.А. 1982. Quercus ussuriensis Krysht. – В кн.: Ископаемые цветковые. Т. 2. Л. С. 94.
[Iljinskaja] Ильинская И.А. 1986. Изменение флоры Зайсанской впадины с конца мела по миоцен. – В кн.: Проблемы палеоботаники. Л. С. 84–112.
[Iljinskaja] Ильинская И.А. 1991. Связи раннеолигоценовой флоры горы Киин-Кериш с современной флорой. – В кн.: Формирование эоценово-миоценовой флоры Казахстана и Русской равнины. Л. С. 15–36.
[Iljinskaja et al.] Ильинская И.А., Кянсеп-Ромашкина Н.П., Панова Л.А., Борисов Б.А. 1983. Палеогеновая флора Зайсанской впадины. – В кн.: Стратиграфия фанерозоя СССР. Л. С. 116–127.
[Kornilova] Корнилова В.С. 1966. Очерк истории флоры и растительности Казахстана. – В кн.: Растительный покров Казахстана. Т. 1. Алма-Ата. С. 37–190.
[Krasheninnikov et al.] Крашенинников В.А., Ахметьев М.А., Борисов Б.А., Запорожец Н.И., Серова М.Я., Яхимович В.Л. 1998. Опорные разрезы границы эоцена и олигоцена на территории бывшего СССР. Разрез Киин-Кериш (впадина озера Зайсан, Юго-Восточный Казахстан). Геологические и биотические события позднего эоцена – раннего олигоцена на территории бывшего СССР. Часть II. Геологические и биотические события. М. С. 169–172.
[Kryshtofovich] Криштофович А.Н. 1955. Развитие ботанико-географических областей северного полушария с начала третичного периода. – Вопросы геологии Азии. Т. 2. М. С. 824–844.
[Kryshtofovich et al.] Криштофович А.Н., Палибин И.В., Шапаренко К.К., Ярмоленко А.В., Байковская Т.Н., Грубов В.И., Ильинская И.А. 1956. Олигоценовая флора горы Ашутас в Казахстане. – Тр. БИН АН СССР. 8 (1): 1–180.
[Kutuzkina] Кутузкина Е.Ф. 1973. Род Hemiptelea (Ulmaceae) в позднесарматской флоре Северного Кавказа. – Бот. журн. 58 (6): 876.
[Losina-Losinskaja] Лозина-Лозинская А.С. 1951. Семейство Ulmaceae Mirb. – Ильмовые. Деревья и кустарники СССР. Т. II. М.; Л. С. 493–523.
[Menitsky] Меницкий Ю.Л. 1984. Дубы Азии. Л. 316 с.
[Panova] Панова Л.А. 1965. Новые данные о палеогеновых и неогеновых спорово-пыльцевых комплексах Зайсанской впадины. – Тр. ВСЕГЕИ. 115: 41–58.
[Pavlyutkin] Павлюткин Б.И. 2002. Позднемиоценовая флора юга Приморья. Владивосток. 192 с.
[Pavlyutkin] Павлюткин Б.И. 2005. Среднемиоценовая ханкайская флора Приморья. Владивосток. 216 с.
[Pavlyutkin] Павлюткин Б.И. 2007. Эоценовая усть-давыдовская флора юга Приморья. Владивосток. 148 с.
Premoli-Silva I., Jenkins D.G. 1993. Decision on the Eocene-Oligocene boundary stratotype. – Episodes. 16 (3): 379–382.
Schneider C.K. 1906. Illustrirtes Handbuch de Laubholzkunde. Band I. Jena. 810 p.
[Suleimanova] Сулейманова Ф.И. 1999. Шкала геомагнитных инверсий и основные черты развития геомагнитного поля в кайнозое (по опорным разрезам Зайсанской впадины и Восточно-Европейской платформы). Дис. … докт. геол.-минер. наук. Уфа. 361 с.
[Yakhimovich et al.] Яхимович В.Л., Борисов Б.А., Сулейманова Ф.И., Данукалов Н.Ф. 1993. Магнитостратиграфическая шкала кайнозоя Зайсанской котловины и ее биостратиграфическое обоснование. Уфа. 67 с.
Zavada M.S., Kim M. 1995. Phylogenetic analysis of Ulmaceae. – Pl. Syst. Evol. 200: 13–20.
[Zhilin] Жилин С.Г. 1974. Третичные флоры Устюрта. Л. 124 с.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал