

Ботанический журнал, 2020, T. 105, № 12, стр. 1207-1220
ОСОБЕННОСТИ МЕЙОЗА ПРИ МИКРОСПОРОГЕНЕЗЕ У PICEA OBOVATA (PINACEAE) ПРИ ИНТРОДУКЦИИ
Е. В. Бажина 1, *, М. И. Седаева 1, **, Е. Н. Муратова 1, ***, Е. А. Бажина 2, ****
1 Институт леса им. В.Н. Сукачева – обособленное подразделение Федерального исследовательского центра “Красноярский научный центр СО РАН”
660036 Красноярск, Академгородок, 50, стр. 28, Россия
2 Московский городской педагогический университет
129226 Москва, 2-й Сельскохозяйственный проезд, 4, Россия
* E-mail: genetics@ksc.krasn.ru
** E-mail: msedaeva@ksc.krasn.ru
*** E-mail: elena-muratova@ksc.krasn.ru
**** E-mail: bazhina09@list.ru
Поступила в редакцию 09.06.2020
После доработки 05.08.2020
Принята к публикации 25.08.2020
Аннотация
Представлены результаты исследований мейоза при микроспорогенезе у ели сибирской (Picea obovata Ledeb.) в условиях интродукции. Выявлены особенности мейоза – четко идентифицируются все стадии профазы первого деления, значительная асинхронность делений не только у разных деревьев, но и в пределах одного микроспорангия. Отмечены особенности ориентации веретен деления и нарушения развития на разных стадиях. Спектр нарушений был меньше, чем у ели сибирской в естественных популяциях Сибири. Особенности развития мужских репродуктивных структур свидетельствуют о наличии высокого адаптивного потенциала ели сибирской в условиях изменяющейся среды.
Ель сибирская (Picea obovata Ledeb.) распространена от границы лесотундры (59–72° с.ш.) на севере до степной зоны (53–56° с.ш.) на юге (Pravdin, 1975). В пределах ареала, адаптируясь к условиям среды, особи одного вида в разных ценопопуляциях формируют экотипы, которые могут различаться по жизнеспособности репродуктивных органов (Mamayev, Popov, 1989; Beuker, 1994; Beuker et al., 1998; Nikkanen et al., 2000). В случае отклонения условий среды от оптимальных, что часто происходит при интродукции, гомеостаз растений нарушается, наблюдаются аномалии роста и развития, изменяются сроки прохождения фенофаз (Nekrasov, 1971; Skroppa, 1994). Особенно чувствительны растения на ювенильной стадии онтогенеза и на начальных этапах репродуктивной фазы, когда процессы споро- и гаметогенеза адаптируются к новому температурному режиму и фотопериоду (Chung, 1981; Shkutko, 1991). Одним из наиболее восприимчивых к воздействию факторов внешней среды периодов в онтогенезе растений является мейоз при микроспорогенезе. Изучение различных видов хвойных при интродукции показало, что реакции репродуктивных органов растений видоспецифичны – мейоз может идти как в направлении стабилизации, так и значительной его нарушенности (Muraya et al., 1988; Bazhina et al., 2011).
Мейоз и формирование пыльцы представителей рода Picea A. Dietr., как в естественных популяциях, так и в условиях культуры, наиболее полно изучены для ели обыкновенной P. abies (L.) H. Karst. (Andersson et al., 1969; Eriksson et al., 1970). У ели сибирской эти процессы изучены в естественных популяциях (Rozhdestvensky, 1981; Bazhina et al., 2019), в том числе и в условиях техногенного стресса (Kalashnik, Yasovieva, 2012). Кроме того, есть данные для некоторых североамериканских и восточноазиатских видов – P. mariana (Mill.) Britton, Sterns et Poggenb., P. pungens Engelm., P. meyeri Rehder et E.H. Wilson, P. smithiana (Wall.) Boiss., P. wilsonii Mast. (Sax, Sax, 1933; Deng, Zhang, 2005; Bazhina et al., 2017). Описание особенностей мейоза при микроспорогенезе у видов Picea с добавочными хромосомами приведено в ряде работ (Moir, Fox, 1975; Fox, 1987). Мейоз при интродукции у видов Picea ранее не изучался.
Долговременные климатические наблюдения (таяние вечной мерзлоты, повышение уровня моря, увеличение продолжительности вегетационного периода) показывают тренды глобального повышения температуры за последние 200 лет (Hulme et al., 1999; Gordon et al. 2000; Groisman et al., 2017). Как полагают некоторые исследователи (Tchebakova et al., 2016), изменения климатических условий приведут к смещению ареалов лесообразующих видов хвойных. В связи с этим остро встает проблема оценки адаптивного потенциала видов и его реализации в условиях изменяющейся среды. Крайне важное значение получает анализ репродуктивного потенциала основных видов хвойных, который определяется особенностями процессов их половой репродукции (Batygina, 2014). Сохранение хвойных растений в условиях культуры является одним из способов поддержания их генофонда. Мониторинг онтогенеза растений вне нативных условий среды, в том числе и при интродукции, выявляет их ответные реакции на смену климатических режимов.
Цель настоящих исследований – установление особенностей мейоза у ели сибирской при интродукции.
ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЙ
Объектами исследований являлись деревья декоративных форм ели сибирской Picea obovata из коллекции Института леса им. В.Н. Сукачева СО РАН (северо-западная окраина г. Красноярска, Академгородок). Коллекция расположена на левобережной террасе р. Енисей (275 м над ур. м.). Возраст деревьев – 53 года. Они посажены в 1972 г. привитыми семилетними саженцами, привезенными из НИИ садоводства Сибири им. М.А. Лисавенко, г. Барнаул (Lucznik, 1976; Loskutov, 1991). Саженцы выращивались из семян, собранных в естественных популяциях Алтая, произрастающих в умеренно континентальном климате (среднегодовая температура +4.0°С). Климат района исследований – резко континентальный (среднегодовая температура +0.5°С, среднегодовое количество осадков – 485 мм/год) (Spravochnik …, 1967, 1969). Почва – дерново-карбонатная, характеризующаяся слабощелочной (pH = 7.01 ± ± 0.08) реакцией среды, невысоким содержанием гумуса (2.55 ± 0.13%), органическое вещество минерализовано, отмечается низкая степень подвижности азота. Несмотря на то, что коллекция находится на окраине крупного промышленного центра (г. Красноярск), территория практически не подвержена промышленному загрязнению благодаря удачному расположению относительно города и розе ветров (Varpholomeev, Maltsev, 2006).
Сбор микростробилов и пыльцы проводился в 2018 г. в период развития пыльцы (вторая декада апреля – первая декада июня). Погодно-климатические условия в период исследований оказались благоприятными для развития – положительные среднесуточные температуры воздуха. Количество осадков в период развития пыльцы составляло 39.1 мм. Сумма эффективных температур рассчитывалась по данным ближайшей метеостанции – “Красноярск. Опытное поле”.
Микростробилы фиксировали ежедневно с 23 по 30 апреля в спиртово-уксусной смеси (3:1). Материал выдерживали в фиксаторе 1–2 суток, затем переводили в 70%-й спирт на хранение. Микростробилы для исследований окрашивали ацетогематоксилином (Pausheva, 1980). В каждом образце просматривали около 300 клеток, определяли частоту встречаемости и спектр аномалий. Статистическая обработка проводилась по общепринятым методикам с использованием пакета MICROSOFT EXCEL 2013.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
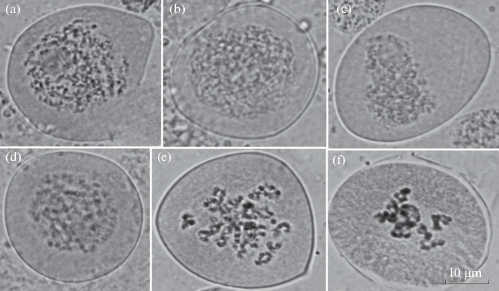
Мужские генеративные почки у ели сибирской, как и у других видов рода Picea, закладываются и начинают дифференцироваться осенью (Owens, Molder, 1985): на побегах первого порядка – 2–4 шт. над мутовкой, на побегах второго порядка – непосредственно на верхушке побега (рис. 1). На первых этапах развития (до окончания мейоза) генеративные почки морфологически практически не отличаются от вегетативных. Мейоз у деревьев ели сибирской, растущих в коллекции, как и в естественных популяциях, начинается и заканчивается весной при сумме эффективных температур 21.7 град/дн. (1.4% от годовой суммы эффективных температур). В третьей декаде апреля в микроспорангиях видны микроспороциты (материнские клетки микроспор) округлой формы на стадии профазы первого деления (рис. 2). У ели сибирской хорошо идентифицируются все стадии профазы (лептотена, зиготена, пахитена, диплотена и диакинез), имеется, как правило, одно ядрышко, которое исчезает в диакенезе. В профазе I образуется 12 бивалентов; их морфология определяется расположением и числом хиазм, которых насчитывается от 1 до 3 на бивалент.
Рис. 1.
Побег ели сибирской с генеративными (Г) и вегетативными (В) почками.
Fig. 1. Picea obovata shoot with generative (Г) and vegetative (В) buds.

Рис. 2.
Профаза первого деления: а – материнская клетка микроспор, b – лептотена, c – пахитена, d – зиготена, e – диплотена, f – диакинез. Масштабная линейка – 10 мкм.
Fig. 2. Prophase I: а – mother cell of microspores, b – leptotene, c – pachytene, d – zygotene, e – diplotene, f – diakinesis. Scale bar – 10 μm.
Мейоз проходит по классическому типу. В мейозе выделяются основные фазы (рис. 3), которые в условиях дендрария проходят достаточно быстро – от 1 до 5 суток у разных деревьев (рис. 4а–4b). Одной из особенностей мейоза данного вида является значительная асинхронность делений не только у разных деревьев, но и в пределах одного дерева (рис. 4 c, 5а–5b). Даже в разных микроспорангиях одного микростробила одновременно отмечаются как материнские клетки микроспор (2.32 ± 0.040% клеток), так и формирующиеся пыльцевые зерна (0.97 ± 0.318). Асинхронные деления наблюдались и в пределах одного микроспорангия (0.36 ± 0.085% от проанализированных клеток). При этом встречались следующие сочетания: материнские клетки микроспор – профаза второго деления, метафазы первого – профазы второго делений, диада – телофазы второго деления. Встречались также следующие сочетания стадий развития в одном микростробиле: лептотена – ранние микроспоры, профаза второго деления – ранние пыльцевые зерна. Необходимо отметить, что у деревьев, растущих в естественных популяциях, развитие микроспороцитов было более синхронным (Bazhina et al., 2019).
Рис. 3.
Мейоз у Picea obovatа: а – метафаза I, b – анафаза I, c – телофаза I, d – диады, e – профаза II, f – метафаза II, g – анафаза II, h – телофаза II, i – тетрады микроспор. Масштабная линейка – 10 мкм.
Fig. 3. Meiosis in Picea obovata: а – metaphase I, b – anaphase I, c – telophase I, d – dyads, e – prophase II, f – metaphase II, g – anaphase II, h – telophase II, i – tetrads. Scale bar – 10 μm.
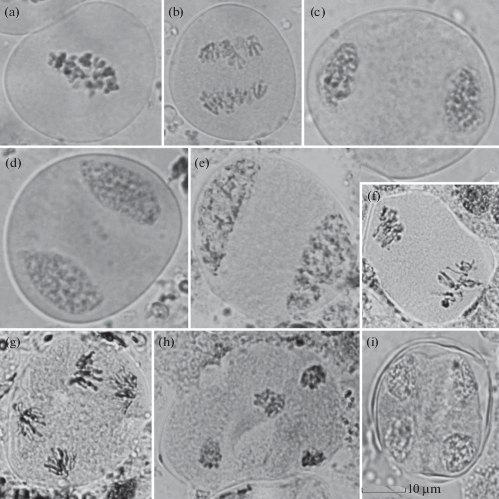
Рис. 4.
Динамика развития клеток.
а – распределение мейоцитов по стадиям профазы первого деления, дерево № 1, ось Х – даты. b – профаза I – пыльцевые зерна, дерево № 1, ось Х – даты. с – профаза I – пыльцевые зерна у разных деревьев 26.04.2018, ось Х: 1–2 – микростробилы дерева № 1, 3–4 – микростробилы дерева № 2, 5–7 – микростробилы дерева № 3. Ось Y – частота встречаемости клеток, %.
Fig. 4. Dynamics of cell development.
а – distribution of meiocytes on prophase I, tree No 1, X-axis – dates. b – prophase I – pollen grains, tree No 1, X-axis – dates. с – prophase I – pollen grains in different trees, April, 26, 2018, X-axis: 1–2 – microstrobiles from tree No 1; 3–4 – microstrobiles from tree No 2; 5–7 – microstrobiles from tree No 3. Y-axis – frequency of cells, percentage.
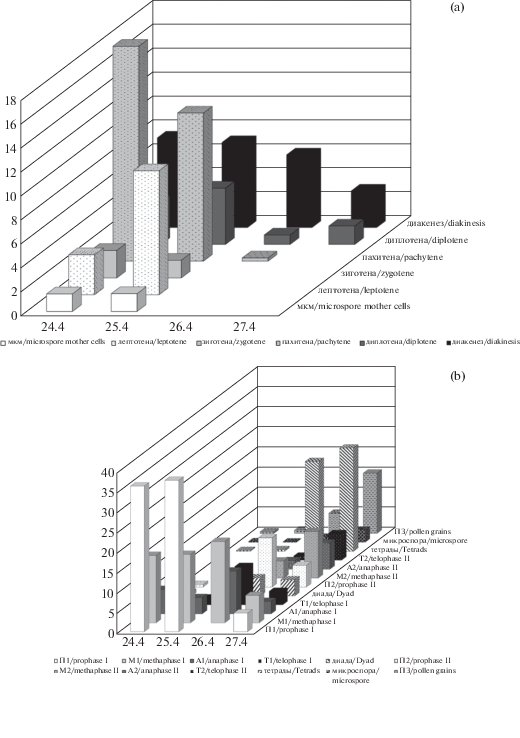
Рис. 4.
Окончание.
Рис. 5.
Особенности мейоза при микроспорогенезе ели сибирской в условиях дендрария: a – асинхронное развитие в пределах одного микроспорангия: метафаза II – анафаза II, b – асинхронное развитие в пределах одного спорангия: метафаза II – формирующаяся микроспора, c – перпендикулярная ориентация веретен деления на стадии анафазы II; d – линейная ориентация веретена деления на стадии метафазы II, e – параллельная ориентация веретена деления на стадии ана-телофазы II. Масштабная линейка – 10 мкм.
Fig. 5. Peculiarities of meiosis in Picea obovata at Arboretum: a – asynchronous development of cells within the same microsporangium: metaphase II – anaphase II, b – asynchronous development of cells within the same sporangium: metaphase II – microspore forming, c – perpendicular orientation of spindles at anaphase II, d – linear orientation of spindles at metaphase II, e – parallel and linear orientation of spindles at ana-telophase II. Scale bar – 10 μm.
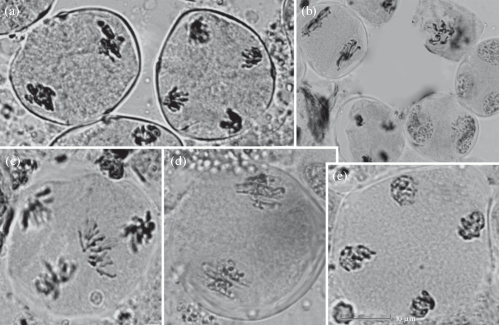
Еще одной особенностью является линейная ориентация веретена деления на стадии метафазы второго деления, а также перпендикулярная и параллельная ориентация на стадии анафазы II. Такие особенности отмечены у одного из исследованных деревьев (№ 1) в 26.03% проанализированных клеток (рис. 5 c–e). По мнению ряда авторов, во втором делении мейоза веретена должны быть ориентированы по отношению друг к другу под углом 60° (Mok, Peloquin, 1975; Butorina et al., 1985; Andreuzza, Siddiqi, 2008). Параллельное расположение может быть обусловлено мутацией гена ps и привести к слиянию одной или двух групп хромосом на противоположных полюсах и формированию пыльцевых зерен с диплоидным числом хромосом. Число мейоцитов с такой особенностью, возможно, определяет некоторое увеличение частоты трехполюсных ана- и телофаз во втором делении (табл. 1). Частота триад на стадии тетрад у этого дерева достигала 21.1%.
Таблица 1.
Частота встречаемости нарушений мейоза, % Table 1. Frequency of meiotic irregularities, percentage
| Стадии мейоза/Meiotic stages | Типы нарушений/Types of irregularities | % |
|---|---|---|
| P1 | Агглютинация хромосом в кольцо в диакенезе Agglutination of chromosomes into a ring at diakenesis |
0.76 |
| M1 | Выброс хромосом вне веретена деления Chaotic chromosome arrangement |
2.05 |
| Агглютинация хромосом в кольцо Agglutination of chromosomes into a ring |
0.41 | |
| Три группы хромосом Three groups of chromosomes |
0.41 | |
| A1 | Отстающие хромосомы Lagging chromosomes |
1.03 |
| Хромосомные мосты Chromosome bridges |
1.03 | |
| Хаотическое расхождение хромосом Chaotic chromosome separation |
1.03 | |
| A2 | Выброс хромосом вне веретена деления Chromosomes outside division spindle |
1.37 |
| Хаотическое расположение хромосом Chaotic chromosome separation |
1.37 | |
| 3-полюсное расхождение хромосом 3-polar chromosome separation |
2.74 | |
| Мосты хромосом Bridges of chromosomes |
1.37 | |
| T2 | 3-полюсное расхождение хромосом 3-polar chromosome separation |
2.55 |
| Тетрады/Tetrads | Триады микроспор Triads of microspores |
4.22 |
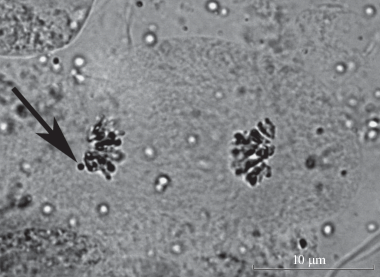
Как показали цитологические исследования, проведенные ранее, в кариотипе одного из изучаемых деревьев встречается добавочная или В-хромосома (Muratova, Vladimirova, 2001). При анализе мейоза у этого дерева В-хромосома была отмечена в анафазе первого деления (рис. 6).
Рис. 6.
В-хромосома в анафазе I (стрелка). Масштабная линейка – 10 мкм.
Fig. 6. В-chromosome at anaphase I (arrow). Scale bar –10 μm.
После распада тетрад микроспор до первых проталлиальных делений у хвойных, как правило, наблюдается период покоя, который может продолжаться до 6 дней (Kozubov, 1974; Bazhina et al., 2011). Однако, в коллекции у отдельных деревьев ели сибирской уже 26 апреля (48 часов после начала мейотических делений) в отдельных клетках наблюдались митотические деления (анафазы).
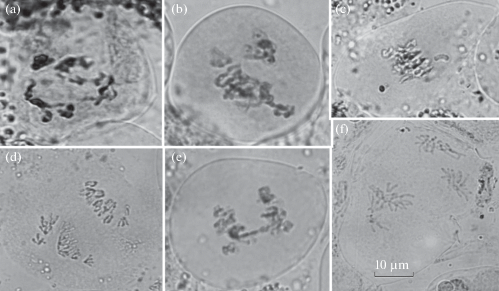
В большинстве изученных клеток мейоз проходил регулярно, хромосомы в основном правильно расходились к противоположным полюсам (табл. 1). Максимальная частота нарушений отмечалась на стадии тетрад (4.2 ± 1.67% составили триады). Некоторые типы нарушений представлены на рис. 7. Спектр наблюдающихся нарушений был несколько меньше, чем у ели сибирской в естественных популяциях Сибири (Bazhina et al., 2019). В то же время отдельные типы нарушений – такие как, агглютинация бивалентов в кольцо в диакинезе, образование трех групп хромосом на стадии метафазы первого деления, хаотическое расположение и отставание хромосом на стадии анафазы первого деления, в естественных популяциях ели сибирской в окрестностях г. Красноярска не встречались. Большая часть нарушений мейоза, как правило, элиминируется при микроспорогенезе. Доля аномальных пыльцевых зерен составляла в среднем 1.3 ± 0.86% (от 0 до 2.9% у отдельных деревьев); они были представлены в основном мелкими пыльцевыми зернами, а также с 1 или 3 воздушными мешками, либо с воротничковым мешком.
Рис. 7.
Аномалии мейоза ели сибирской: a – агглютинация хромосом в кольцо на стадии диакенеза, b – три группы хромосом на стадии метафазы I, c – выброс хромосом в метафазе I, d – выбросы группы хромосом в анафазе I, e – мост в анафазе I, f – множественные нарушения на стадии анафазы II. Масштабная линейка – 10 мкм.
Fig. 7. Irregularities of meiosis in Picea obovata: a – chromosome agglutination into a ring at diakinesis, b – three groups of chromosomes at metaphase I, c – chaotic chromosome arrangement at metaphase I: chromosome outside spindle divisions, d – chaotic arrangement of chromosome groups at metaphase I, e – bridge at anaphase I, f – multiple irregularities at anaphase II. Scale bar – 10 μm.
ОБСУЖДЕНИЕ
Сроки и продолжительность фенологических фаз развития вида могут значительно варьировать в зависимости от условий произрастания, в частности, от высоты и широты местности (Owens, Molder, 1985). Эффекты влияния климатических факторов на наступление фенофаз (начало роста почек, цветение, опыление, и пр.) описаны у различных видов рода Picea (Sirois, 2000; Messaud et al., 2007). Начало, продолжительность и особенности развития репродуктивных структур контролируются температурой воздуха и могут различаться не только у разных видов, но и в разных популяциях одного вида. Так, например, у P. sitchensis (Bong.) Carrière в условиях Шотландии мейоз при микроспорогенезе наблюдается с середины февраля по конец апреля (Moir, Fox, 1975), тогда как в Британской Колумбии (о. Ванкувер) – завершается в уже марте (Fox, 1987). Настоящие исследования показали достаточно динамичное прохождение мейоза у ели сибирской в коллекции (в течение 1–5 суток), что может свидетельствовать о высоком адаптивном потенциале вида. У другого сибирского представителя темнохвойных формаций – Abies sibirica Ledeb. продолжительность мейоза при интродукции в этих условиях значительно увеличивалась – более чем на две недели (Bazhina et al., 2011). Необходимо отметить, что мейотические деления у ели сибирской в коллекции начинались при значительно меньшей сумме эффективных температур и наблюдались несколько дольше, чем в естественных популяциях в окрестностях Красноярска (Bazhina, Sedaeva, 2019). Можно предположить, что это обусловлено локальными различиями в экологических условиях: радиационный и тепловой режим леса и города значительно различается, территория города нагревается сильнее (Khromov, 1983). Кроме того, деревья в коллекции Института произрастают на открытом пространстве, что способствует еще более интенсивному прогреванию и, как следствие, раннему началу развития.
Способность адаптироваться к иным условиям среды у ели проявляется также в значительной степени асинхронности развития мейоцитов, выявленной в настоящем исследовании. Асинхронное развитие микроспороцитов наблюдается у многих видов хвойных (Moir, Fox, 1975; Muraya et al., 1988; Muratova, 1995). Предполагается, что это может быть обусловлено неодинаковыми температурными и иными условиями развития в разных микроспорангиях, а также различиями в питании микростробилов. Однако, такой степени асинхронности, как выявлено в настоящем исследовании, у ели сибирской ранее не наблюдалось ни в естественных популяциях Сибири, ни в экстремальных условиях Крайнего Севера (Rozhdestvensky, 1981; Bazhina et al., 2019).
Большинство стадий мейоза очень чувствительно к изменениям температуры (Andersson et al., 1969; Eriksson et al., 1970; Andersson, 1980; Luomajoki, 1993). Температурные экстремумы, характерные весной для резко континентального климата Сибири, оказывают негативное влияние на развитие мужских генеративных структур, вызывая нарушения мейоза и стерильность пыльцы. Так, например, увеличение продолжительности мейоза у деревьев пихты сибирской, культивируемых в дендрарии Института леса до двух недель приводит к значительной его нарушенности (Bazhina et al., 2011). Однако, отклик растений на стрессовые условия видоспецифичен и поведение хромосом в мейозе обусловлено генетически (Shkutina, 1975; Golubovskaya, 1979; Simanovsky, Bogdanov, 2018). Показано, что критическая температура, способная вызвать хромосомные нарушения в мейозе при микроспорогенезе у видов Picea, составляет –2 – –4°С (Andersson et al., 1969). Таких экстремальных температур в период мейоза и развития пыльцы ели не наблюдалось, что, очевидно, и обусловило невысокий уровень нарушений.
Некоторые аномалии мейоза у ели сибирской, отмеченные в настоящем исследовании, могут определяться генетически. В частности, агглютинация хромосом на стадиях профазы и метафазы первого деления может быть связана с мутациями, вызывающими блокаду профазы I мейоза (Golubovskaya, 1979; Golubovskaya, Sitnikova, 1980), либо моно- или дигенными рецессивными мутациями, также приводящими к агглютинации и отставанию хромосом (Sosnikhina et al., 1994; Mehra, Rai, 1970). После цитокинеза при этом образуются полиады с микро- и макроядрами различного размера и хромосомного состава, что приводит к формированию неоднородных по размеру и форме пыльцевых зерен и снижению фертильности пыльцы (Beadle, 1937; Mehra, Rai, 1970; Muratova, 1995). Уровень таких нарушений у ели сибирской не превышает 1%.
Спектр нарушений у деревьев ели сибирской, произрастающих в условиях дендрария, несколько меньше, чем в естественных популяциях, что также может быть обусловлено их генетическими особенностями. Генами, контролирующими формирование веретена деления и расхождения хромосом, могут быть обусловлены аномалии расхождения хромосом у ели сибирской, наблюдаемые в ана-телофазах второго деления. Так, у кукурузы и лапчатки выявлены гены dv и mei-4, действие которых приводит к повреждению ахроматического веретена (Golubovskaya, 1979; Golubovskaya, Sitnikova, 1980). При полном повреждении веретена хромосомы распределяются по клетке хаотично или концентрируются в центре; при частичном может наблюдаться неравномерное распределение отдельных хромосом или их групп между двумя или тремя полюсами, а также отстающие хромосомы (Butorina et al., 1988). Аномалии мейоза, связанные с потерей генетического материала (выбросы, отставания хромосом), как правило, ведут к формированию гамет с анеуплоидным числом хромосом, образованию мелкой фракции пыльцы или дефектных пыльцевых зерен. Мосты, обнаруженные в отдельных мейоцитах ели сибирской в анафазе первого и второго делений, могут быть вызваны запоздалой терминализацией хиазм, разрывами хромосом на разных стадиях мейоза и кроссинговером между гомологичными хромосомами (Khvostova, Yachevskaya, 1975).
ЗАКЛЮЧЕНИЕ
Изучение онтогенеза растений в новых условиях среды, в частности при интродукции, выявляет ответные реакции видов на быструю смену климатических режимов. В жизненном цикле растений наиболее чувствительными к условиям стресса являются процессы репродукции, и особенно, развитие мужских генеративных структур. Анализ мейоза при микроспорогенезе у деревьев ели сибирской, интродуцированных в коллекции Института леса (Красноярск), показал быстрое прохождение мейотических делений, значительную асинхронность развития мейоцитов и достаточно узкий спектр аномалий, что, вероятно, обусловливает низкий уровень нарушений и достаточно высокую жизнеспособность пыльцы (Bazhina, Sedaeva, 2019). Изучение морфологии и развития пыльцы у различных видов рода Picea в условиях дендрария Института леса показало высокий адаптивный потенциал мужской генеративной сферы у всех представителей данного рода (Bazhina et al., 2017). Результаты исследований подтверждают высокие адаптивные способности развития мужских репродуктивных структур ели сибирской, что, возможно, характерно для рода Picea в целом.
Список литературы
Andersson E. 1980. Temperature-conditioned irregularities in pollen mother cells of Picea abies (L.) Karst. – Hereditas. 92 (1): 27–35.
Andersson E., Ekberg I., Eriksson G. 1969. A summary of meiotic investigation in conifers. – Stud. For. Suecica. 70: 1–20.
Andreuzza S., Siddiqi I. 2008. Spindle Positioning, Meiotic Nonreduction, and Polyploidy in Plants. – PLoS Genetics. 4 (11): e1000272.
[Batygina] Батыгина Т.Б. 2014. Биология развития растений. Симфония жизни. СПб. 764 с.
Bazhina E.V., Cedaeva M.I., Muratova E.N. 2019. Meiosis during Microsporogenesis in Siberian Spruce (Picea obovata Ledeb.) in the South of Central Siberia. – Russ. J. Dev. Biol. 50 (3): 113–123. https://doi.org/10.1134/S1062360419030020
Bazhina E.V., Kvitko O.V., Muratova E.N. 2011. Specific features of meiosis in the Siberian fir (Abies sibirica) in the forest Arboretum of the V.N. Sukachev Institute, Russia. – Biodivers. Conserv. 20 (2): 415–428.
[Bazhina, Sedaeva] Бажина Е.В., Седаева М.И. 2019. Особенности развития и жизнеспособность пыльцы ели сибирской в коллекции Института леса как модельный объект реакции вида на изменения климата. – В сб.: Матер. XXII Межд. науч. конференции. “Плодоводство, семеноводство, интродукция древесных растений”. Красноярск. С. 11–14.
Bazhina E.V., Sedaeva M.I., Goryachkina O.V., Muratova E.N. 2017. Pollen development and morphology in different Picea A. Dietr. species at the V.N. Sukachev Institute of Forest Arboretum. – The International Journal of Plant Reproductive Biology. 9 (1): 5–12. https://doi.org/10.14787/IJPRB.20179.1.5-12
Beadle G.W. 1937. Chromosome aberration and the gene mutation in sticky chromosome plants of Zea mays. – Cytologia. 1: 43–56.
Beuker E. 1994. Adaptation to climatic changes of the timing of bud burst in populations of Pinus sylvestris L. and Picea abies (L.) Karst. – Tree Physiol. 14 (7–9): 961–970. https://doi.org/10.1093/treephys/14.7-8-9.961
Beuker E., Valtonen E., Repo T. 1998. Seasonal variation in the frost hardiness of Scots pine and Norway spruce in old provenance experiments in Finland. – For. Ecol. Manag. 107 (1–3): 87–98. https://doi.org/10.1016/S0378-1127(97)00344-7
[Butorina at al.] Буторина А.К., Мурая Л.С., Машкина О.С., Высоцкий А.А. 1985. Новый тип мейотической мутации у сосны. – Генетика. 21 (1): 103–111.
[Butorina at al.] Буторина А.К., Мурая Л.С., Рязанцева Л.А., Высоцкий А.А. 1988. Цитогенетическая и биохимическая характеристика деревьев сосны обыкновенной (Pinus sylvestris L.), контрастных по смолопродуктивной способности. – Генетика. 24 (5): 889–899.
Chung M.-S. 1981. Flowering characteristics of Pinus sylvestris L. with special emphasis on the reproductive adaptation to local temperature factor. – Acta For. Fenn. 169: 1–69.
Deng H.-Sh., Zhang D.-M. 2005. Microsporogenesis and meiotic behavior in three species of the genus Picea (Pinaceae). – J. Hechi University. 25 (5): 23–27.
Eriksson G., Ekberg I., Jonsson A. 1970. Meiotic investigations in pollen mother cells of Norway spruce cultivated in a plastic greenhouse. – Hereditas. 66 (1): 1–20.
Fox D.P. 1987. The chromosomes of Picea sitchensis (Bong.) Carr. and its relatives. – Proceedings of the Royal Society of Edinburgh. 93 (1–2): 51–59. https://doi.org/10.1017/S0269727000006278
Golubovskaya I.N. 1979. Genetic control of meiosis. – International Review Cytology. 58: 247–290.
[Golubovskaya, Sitnikova] Голубовская И.Н., Ситникова Д.В. 1980. Три мейотические мутации кукурузы, вызывающие расхождения хромосом в первом делении мейоза. – Генетика. 16 (4): 656–666.
Gordon C., Cooper C., Senior C.A., Banks H., Gregory J.M., Johns T.C., Mitchell J.F.B., Wood R.A. 2000. The simulation of SST, sea-ice extent and ocean heat transport in a version of the Hadley Centre coupled model without flux adjustments. – Climatic Dyn. 16 (2): 147–168. https://doi.org/10.1007/S003820050010
Groisman P., Kicklighter D., Henebry G., Shugart H., Tchebakova N. et al. 2017. Northern Eurasia Future Initiative (NEFI): Facing the Challenges and Pathways of Global Change in the 21st Century. – Progress in Earth and Planetary Science. 4: 41. https://doi.org/10.1186/s40645-017-0154-5
Hulme M., Mitchell J.F.B., Ingram W., Lowe J.A., Johns T.C., New M.G., Viner D. 1999. Climate change scenarios for global impacts studies. – Glob. Environ. Change. 9 (1): P. 3–19. https://doi.org/10.1016/S0959-3780(99)00015-1
Kalashnik N.A., Yasovieva S.M. 2012. Analysis of meiotic chromosome aberrations in siberian spruce (Picea obovata Ledeb.) under conditions of natural and technogenic stress. – Russ. J. Ecol. 43 (6): 440–447. https://doi.org/10.1134/S1067413612060057
[Khvostova, Yachevskaya] Хвостова В.В., Ячевская Г.Л. 1975. Перестройки хромосом в мейозе. – В кн.: Цитология и генетика мейоза. М. С. 232–262.
[Khromov] Хромов С.П. 1983. Метеорология и климатология для географических факультетов. 3-е изд. Л. 456 с.
[Kozubov] Козубов Г.М. 1974. Биология плодоношения хвойных на Севере. Л. 136 с.
[Loskutov] Лоскутов Р.И. 1991. Интродукция декоративных древесных растений в южной части Средней Сибири. Красноярск. 189 с.
[Lucznik] Лучник З.И. 1976. Изменчивость ели сибирской (Picea obovata Ledeb.) на Алтае. – Новости сист. высш. pаст. 13: 5–8.
Luomajoki A. 1993. Climatic adaptation of Norway spruce (Picea abies (L.) Karsten) in Finland based on male flowering phenology. – Acta For. Fenn. 242: 1–28.
[Mamayev, Popov] Мамаев С.А., Попов П.П. 1989. Ель сибирская на Урале (внутривидовая изменчивость и структура популяций). М. 104 с.
Mehra P.N., Rai K.S. 1970. Cytogenetic studies of meiotic abnormalities in Collinsia tinctoria. I. Chromosomal stickiness. – Can. J. Genet. Cytol. 12 (3): 560–569. https://doi.org/10.1139/G70-075
Messaud Y., Bergeron Y., Asselin H. 2007. Reproductive potential of balsam fir (Abies balsamea), white spruce (Picea glauca), and black spruce (P. mariana) at the ecotone between mixedwood and coniferous forests in the boreal zone of western Quebec. – Am. J. Bot. 94 (5): 746–754. https://doi.org/10.3732/AJB.94.5.746
Moir R.B., Fox D.P. 1975. Male meiosis in Sitka spruce Picea sitchensis (Bong.) Carr. – Silvae Genet. 24 (5–6): 187–192.
Mok D.W.S., Peloquin S.J. 1975. Three mechanisms of 2n pollen formation in diploid potatoes. – Can. J. Genet. Cytol. 17 (2): 217–225. https://doi.org/10.1139/G75-029
[Muratova] Муратова Е.Н. 1995. Особенности мейоза сосны обыкновенной около северной границы ее ареала. – Онтогенез. 26 (2): 128–139.
[Muratova, Vladimirova] Муратова Е.Н., Владимирова О.С. 2001. Добавочные хромосомы кариотипа ели сибирской P. obovata. – Цитология и генетика. 35 (4): 38–44.
[Muraya et al.] Мурая Л.С., Буторина А.К., Дудецкая Е.М. 1988. Мейоз при микроспорогенезе и развитие мужского гаметофита у лжетсуги в условиях интродукции. – Лесоведение. 5: 37–44.
[Nekrasov] Некрасов В.И. 1971. Некоторые теоретические вопросы формирования интродукционных популяций лесных древесных пород. – Лесоведение. 5: 26–30.
Nikkanen T., Aronen T., Häggman H., Venäläinen M. 2000. Variation in pollen viability among Picea abies genotypes – potential for unequal paternal success. – Theor. Appl. Genet. 101: 511–518. https://doi.org/10.1007/S001220051510
Owens J.N., Molder M. 1985. The reproductive cycles of true firs. – Information Services Branch Ministry of Forests. Victoria, British Columbia. 35 p.
[Pausheva] Паушева З.П. 1980. Практикум по цитологии растений. М. 304 с.
[Pravdin] Правдин Л.Ф. 1975. Ель европейская и ель сибирская в СССР. М. 176 с.
[Rozhdestvensky] Рождественский Ю.Ф. 1981. О развитии мужских генеративных органов ели сибирской в районах Крайнего Севера. – Лесоведение. 3: 35–42.
Sax K., Sax H.J. 1933. Chromosome number and morphology in the conifer. – J. Arnold Arboretum. 14 (4): 356–374.
[Shkutina] Шкутина Ф.М. 1975. Мейоз отдаленных гибридов и амфиплоидов. – В кн.: Цитология и генетика мейоза. М. С. 292–311.
[Shkutko] Шкутко Н.В. 1991. Хвойные Беларуссии: эколого-биологические исследования. Минск. 264 с.
Simanovsky S.A., Bogdanov Yu.F. 2018. Genetic control of meiosis in plants. – Russ. J. Genet. 54 (4): 389–402. https://doi.org/10.1134/S1022795418030122
Sirois L. 2000. Spatiotemporal Variation in Black Spruce Cone and Seed Crops along a Boreal Forest-Tree Line Transect. – Can. J. For. Res. 30: 900–909. https://doi.org/10.1139/x00-015
Skroppa T. 1994. Growth rhythm and hardiness of Picea abies progenies of high altitude parents from seed produced at low elevation. – Silvae Genetica. 43 (2–3): 95–100.
Sosnikhina S.P., Fedotova Yu.S., Smirnov V.G., Mikhaylova E.I., Bogdanov Yu.F. 1994. The study of genetic control of meiosis in Rye. – Russ. J. Genet. 30 (8): 909–920.
[Spravochnik…] Справочник по климату СССР. В. 21. Ч. II. 1967. Л. С. 87.; В 21. Ч. IV. 1969. Л. С. 189.
Tchebakova N.M., Parfenova E.I., Korets M.A., Conard S.G. 2016. Potential change in forest types and stand heights in central Siberia in a warming climate. – Environmental Research Letters. 11: 03501. https://doi.org/10.1088/1748-9326/11/3/035016
[Varpholomeev, Maltsev] Варфоломеев И.И., Мальцев Ю.М. 2006. Государственный доклад о состоянии и защите окружающей среды в Красноярском крае в 2006 г. Красноярск. https://krasecology.ru/storage/Index?guid=3ee21b10-16af-418a-b9d7-06d831e815a7
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал