

Ботанический журнал, 2020, T. 105, № 5, стр. 479-496
ОНТОГЕНЕТИЧЕСКАЯ СТРУКТУРА ЦЕНОПОПУЛЯЦИЙ BULBOCODIUM VERSICOLOR (MELANTHIACEAE) В НИЖНЕМ ПОВОЛЖЬЕ
А. С. Кашин 1, *, Л. В. Куликова 1, Н. А. Петрова 1, И. В. Шилова 1, А. С. Пархоменко 1, М. В. Лаврентьев 1
1 ФГБОУ ВО “СГУ имени Н.Г. Чернышевского”
410010 г. Саратов, ул. Астраханская, 83, Россия
* E-mail: kashinas2@yandex.ru
Поступила в редакцию 22.01.2020
После доработки 17.02.2020
Принята к публикации 18.02.2020
Аннотация
С 2014 по 2019 гг. в 23-х местообитаниях Нижнего Поволжья проведен мониторинг состояния ценопопуляций охраняемого вида Bulbocodium versicolor (Ker Gawl.) Spreng. (Melanthiaceae), который представлен здесь разобщенными локусами или единичными особями. При этом ценопопуляции проявляют существенную гетерогенность онтогенетических спектров субвыборок, что связано с неоднородностью пространственного размещения особей. Как правило, онтогенетические спектры ценопопуляций B. versicolor имели максимум на генеративных растениях. Установлена умеренная отрицательная корреляция (r = –0.49) между общим проективным покрытием и индексом восстановления. B. versicolor выпадает из сообщества при зарастании длиннокорневищными злаками и увеличении общего проективного покрытия. Условия сильной засухи провоцируют элиминацию ювенильных особей и переход растений разного онтогенетического состояния в круглогодичный покой. Большое количество осадков приводит к выходу из состояния круглогодичного покоя клубнелуковиц, пребывавших в нем в предыдущий год (или годы).
Брандушка разноцветная – Bulbocodium versicolor (Ker Gawl.) Spreng. (syn. Colchicum bulbo-codium subsp. versicolor (Ker Gawl.) K. Perss.) (Melanthiaceae) – редкий европейский вид с дизъюнктивным ареалом, распространенный преимущественно в лесостепной и степной зонах на Восточно-Европейской равнине от Подольской до Приволжской возвышенности, охраняемый на территории России и на Украине (Krasnaya…, 2008; Chervona…, 2009). Отдельные небольшие эксклавы расположены в Молдове, Румынии, Венгрии, Сербии и Италии (Csapody, 1982; Oltean et al., 1994; Conti et al., 1997; Conti, Bartolucci, 2012). На горных лугах в Пиренеях и Альпах, произрастает другой вид – B. vernum L. (Valentine, 1980). Однако, результаты морфологического и молекулярно-генетического анализа показали, что они представляют собой формы одного вида (Sramkó et al., 2008).
В России B. versicolor встречается преимущественно на Среднерусской и Приволжской возвышенностях (Tzvelev, 1979). В Нижнем Поволжье проходит юго-восточная и восточная (Sagalaev, Shantser, 2006), а в Саратовской обл. – частично еще и северная границы (Petrova et al., 2015) его ареала. Лимитирующими численность B. versicolor факторами в регионе принято считать антропогенное нарушение мест обитания (Krasnaya…, 2006) и аридизацию климата (Kulikova et al., 2019).
Большая часть исследований, посвященных отдельным аспектам экологии B. versicolor, а также структуре его популяций, проводилась на Украине (Pogrebennik et al., 1987; Goncharenko, Karpenko, 1998; Krivoruchko, 2005; Melnik et al., 2006а, b, 2007; Sokolov et al., 2011 и др.).
Известно, что выявление оптимальных эколого-климатических условий и факторов, снижающих численность редких видов, является важной составляющей изучения их биологии, в частности онтогенетической структуры популяций. Методика сбора материала для характеристики онтогенетического спектра ценопопуляции (ЦП), как правило, заключается в том, что каждая выборка включает несколько субвыборок, описанных в разных частях ЦП или в разные годы. Затем данные по всем субвыборкам суммируются или усредняются (Tsenopopulyatsii…, 1976, 1988). Однако суммирование материалов субвыборок (учетных площадок) и анализ данных по выборке в целом правомочно только в случае однородности (гомогенности) выборки, когда распределения разных субвыборок в пределах выборки статистически значимо не различаются, т. е. они могут быть описаны одним полиномиальным распределением (Ivanov, 2014). Данное условие однородности выборки при изучении ЦП B. versicolor очень часто не выполняется. Структура ЦП луковичных эфемероидов склонна претерпевать существенные колебания по годам. Для решения задач оценки и сравнения параметров онтогенетической структуры гетерогенных выборок предложен алгоритм статистических методов (Glotov, Ivanov, 2014), реализованный в программе для оценки и сравнения параметров онтогенетических спектров популяций растений и лишайников при гетерогенности выборки “OntoParam” в статистической среде R (Statisticheskaya…, 2019).
Целью данной работы являлся анализ онтогенетической структуры ЦП B. versicolor в различных эколого-ценотических условиях Нижнего Поволжья, а также анализ динамики онтогенетической структуры ЦП во времени, с учетом однородности (или неоднородности) выборок.
МАТЕРИАЛЫ И МЕТОДЫ
Сбор материала проводился в 2014–2019 гг. в 23-х местообитаниях на территории Саратовской и Волгоградской областей (табл. 1, рис. 1). Характеристика погодных условий на территории Саратовской обл. в данный период приведена на рис. 2. Во всех местообитаниях определено общее проективное покрытие, установлены доминанты травостоя и видовой состав растительного сообщества. Анализ местообитаний по растительному покрову с использованием экологических шкал Л.Г. Раменского (Ramenskii et al., 1956) проведен в программе EcoScaleWin (Zubkova et al., 2008; Grokhlina, Khanina, 2015).
Таблица 1.
Характеристика исследованных ценопопуляций в 2018 г. Table 1. Characteristics of the surveyed coenopopulations in 2018
| № ЦП № of CP |
Локалитет Locality |
Доминанты и
субдоминанты сообщества* Community dominants and subdominants* |
Общее проективное
покрытие, % General projective cover, % |
Характер степного войлока и нарушен-ности фитоценоза The nature of steppe felt and phytocenosis |
|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 |
| 1 | Саратовская обл., Балашовский р-н, окр. с. Тростянка/Saratov Region, Balashov District, Trostyanka | Alopecurus pratensis – Festuca valesiaca – Poa pratensis – Galium verum – Rumex acetosella – Plantago urvillei | 100 | Густая ветошь, зарастание злаками/Dense dead grasses, overgrown with grasses |
| 2 | Саратовская обл., Балашовский р-н, окр. с. Ключи/Saratov Region, Balashov District, Klyuchi | Alopecurus pratensis – Poa pratensis – Ficaria stepporum – Ranunculus acris – Fritillaria meleagroides – Stemmacantha serratuloides – Bulbocodium versicolor | 70 | Выпас, сенокос/Gra-zing, haying |
| 3 | Саратовская обл., Татищевский р-н, окр. с. Широкое/Saratov Region, Tatishchevo District, Shirokoye | Galatella villosa – Artemisia austriaca – Poa bulbosa – Bulbocodium versicolor | 45 | Интенсивный выпас, рекреация/Intensive grazing, recreation |
| 4 | Саратовская обл., Татищевский р-н, окр. с. Курдюм/Saratov Region, Tatishchevo District, Kurdyum | Festuca valesiaca – Valeriana tuberosa – Artemisia austriaca – Poa bulbosa | 90 | Густая ветошь, рекреация/Thick dead grasses, recreation |
| 5 | Саратовская обл., Саратовский р-н, окр. п. Красный Текстильщик/ Saratov Region, Saratov District, Krasnyy Tekstilshchik | Elytrigia intermedia – Bromopsis riparia – Fragaria viridis | 99 | Густая ветошь, сильное задернение/Thick dead grasses, strong sodding |
| 6 | Саратовская обл., Красноармейский р-н, окр. ст. Паницкая/Saratov Region, Krasnoarmeysk District, Panitskaya | Festuca valesiaca – Koeleria cristata – Elytrigia repens – Artemisia austriaca – Bulbocodium versicolor | 45 | Осенний пожар, выпас/Autumn fire, grazing |
| 7 | Саратовская обл., Красноармейский р-н, окр. ст. Паницкая/Saratov Region, Krasnoarmeysk District, Panitskaya | Festuca valesiaca – Koeleria cristata – Elytrigia repens – Bromus squarrosus – Artemisia campestris – Bulbocodium versicolor | 45 | Осенний пожар, выпас/Autumn fire, grazing |
| 8 | Саратовская обл., Энгельсский р-н, окр. пос. Красноармейское/Saratov Region, Engels District, Krasnoarmeyskoye | Festuca valesiaca – Artemisia campestris – Agropyron cristatum – Bulbocodium versicolor – Gagea pusilla – Erophila verna | 30 | Стерни и ветоши почти нет, умеренный выпас/Almost no stubble or rags, moderate grazing |
| 9 | Саратовская обл., Энгельсский р-н, окр. п. Прилужный/Saratov Region, Engels District, Priluzhnyy | Festuca valesiaca – Koeleria cristata – Artemisia austriaca – Bulbocodium versicolor | 40 | Ветоши мало, выпас/ Little dead grasses, grazing |
| 10 | Саратовская обл., Энгельсский р-н, окр. с. Новочарлык/Saratov Region, Engels District, Novocharlyk | Festuca valesiaca – Koeleria cristata – Artemisia austriaca – Bulbocodium versicolor – Poa bulbosa – Valeriana tuberosa | 50 | Войлок тонкий, выпас/Thin felt, grazing |
| 11 | Саратовская обл., Ровенский р-н, окр. п. Лиманный/Saratov Region, Rovnoye District, Limanny | Agropyron cristatum – Elytrigia lolioides – Festuca valesiaca – Bulbocodium versicolor | 75 | Ветоши мало, выпас/ Little dead grasses, grazing |
| 12 | Волгоградская обл., Еланский р-н, окр. с. Большой Морец/Volgograd Region, Yelan’ District, Bolshoy Morets | Festuca valesiaca – Alopecurus pratensis – Poa pratensis | 90 | Густой войлок, выпас/Thick felt, grazing |
| 13 | Волгоградская обл., Жирновский р-н, окр. с. Бородачи/Volgograd Region, Zhirnovsk District, Borodachi | Festuca valesiaca – Stipa capillata – Bulbocodium versicolor | 50 | Ветоши нет, обнажения почвы, интенсивный выпас/ Dead grasses missing, gaps with bare soil, intensive grazing |
| 14 | Волгоградская обл., Камышинский р-н, окр. с. Верхняя Добринка/Volgograd Region, Kamyshin District, Verkhnyaya Dobrinka | Elytrigia repens – Poa bulbosa – Bulbocodium versicolor | 50 | Ветоши мало, выпас, сенокос/Little dead grasses, grazing, haymaking |
| 15 | Волгоградская обл., Старополтавский р-н, окр. с. Валуевка/Volgograd Region, Staraya Poltavka District, Valuyevka | Festuca valesiaca – Bulbocodium versicolor – Tanacetum achilleifolium – Allium angulosum – Gagea pusilla | 65 | Ветоши мало, выпас/ Little dead grasses, grazing |
| 16 | Волгоградская обл., Нехаевский р-н, окр. хут. Нестеровский/Volgograd Region, Nekhaevskaya District, Nesterovskiy | Stipa capillata – Festuca valesiaca – Artemisia austriaca – Bulbocodium versicolor | 30 | Осенний пожар, выпас/Autumn fire, grazing |
| 17 | Волгоградская обл., Нехаевский р-н, окр. ст. Нехаевская/Volgograd Region, Nekhaevskaya District, Nekhaevskaya | Stipa capillata – Bulbocodium versicolor – Poa angustifolia – Centaurea jacea – Potentilla argentea | 75 | Сенокос, выпас/ Haymaking, grazing |
| 18 | Волгоградская обл., Даниловский р-н, окр. ст. Сергиевская/Volgograd Region, Danilovka District, Sergievskaya | Festuca valesiaca – Elytrigia repens – Poa bulbosa – Bulbocodium versicolor – Gagea pusilla | 95 | Ветоши мало,
интенсивный выпас/ Little dead grasses, intense grazing |
| 19 | Волгоградская обл., Котовский р-н, окр. хут. Попки/Volgograd Region, Kotovo District, Popki | Stipa lessingiana – Festuca valesiaca – Bulbocodium versicolor | 75 | Ветоши мало, стерня густая, выпас/Little dead grasses, thick stubble, grazing |
| 20 | Волгоградская обл., Клетский р-н, окр. х. Венцы/Volgograd Region, Kletskaya District, Ventsy | Stipa lessingiana – Bulbocodium versicolor | 95 | Густая ветошь, выпас/ Thick dead grasses, grazing |
| 21 | Волгоградская обл., Иловлинский р-н, окр. ст. Сиротинской/Volgograd Region, Ilovlya District, Sirotinskaya | Artemisia austriaca – Festuca valesiaca – Galatella villosa – Bromopsis riparia – Poa angustifolia | 70 | Густой войлок, выпас/Thick felt, grazing |
| 22 | Волгоградская обл., Иловлинский р-н, окр. х. Хмелевской/Volgograd Region, Ilovlya District, environs of Khme-levskoy | Poa angustifolia – Bulbocodium versicolor | 90 | Густая ветошь/ Thick dead grasses |
| 23 | Волгоградская обл., г. Волгоград, балка Отрада/Volgograd Region, Volgograd, Otrada ravine | Poa bulbosa – Bulbocodium versicolor – Gagea pusilla | 70 | Ветоши мало, выпас/ Little dead grasses, grazing |
Рис. 1.
Локалитеты ценопопуляций Bulbocodium versicolor. Белыми кругами отмечены ценопопуляции, исследованные в 2018 г., черными – не исследованные в 2018 г. Здесь и далее обозначения ценопопуляций те же, что в табл. 1.
Fig. 1. Localities of Bulbocodium versicolor coenopopulations. White circles indicate cenopopulations studied in 2018, black – not studied in 2018. Here and further, see Table 1 for cenopopulation designations.
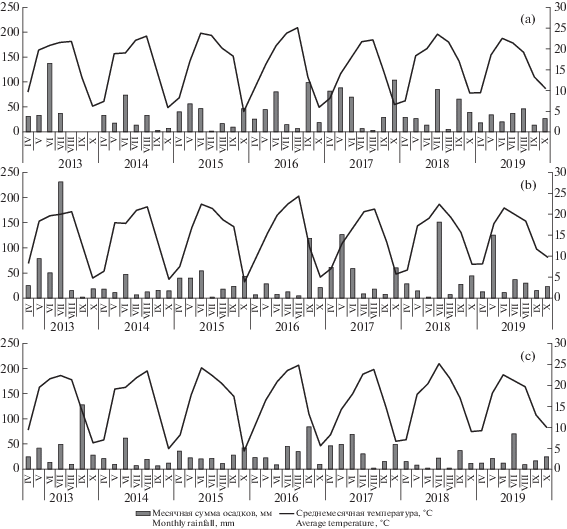
Рис. 2.
Среднемесячная температура и количество осадков с апреля по октябрь в годы исследований в местах расположения ценопопуляций Bulbocodium versicolor в Саратовской области по данным ближайших метеостанций (a – Саратов (аэропорт Южный); b – Сплавнуха; c – Красный Кут). По левой оси ординат – количество осадков, мм; по правой оси ординат – температура, ° С.
Fig. 2. Average monthly temperature and amount of precipitation from Аpril to Оctober during the years of research in the localities of cenopopulations Bulbocodium versicolor in the Saratov Region according to the nearest weather stations (a – Saratov (Yuzhnyy airport); b – Splavnukha; c – Krasnyy Kut). Left Y-axis – the amount of precipitation, mm; right Y-axis – temperature, ° С.
В каждой ЦП в пределах постоянной пробной площади 100 м2 случайным образом закладывали 10 учетных площадок 1 м2, на которых подсчитывали особи B. versicolor с учетом онтогенетических состояний и дальнейшим построением онтогенетических спектров (Tsenopopulyatsii…, 1976; Zhukova et al., 2014). Рассчитывали средние значения плотности особей, сравнивая эти значения при p ≤ 0.05.
Онтогенетическое состояние особи определялось без уничтожения растений, на основе размеров, числа и соотношения надземных органов (Pravila…, 1981). За основу брали описания онтогенетических состояний, приведенные для B. versicolor (Melnik et al., 2007; Metodychni…, 2016). Принимая во внимание специфические особенности вида, выделяли ювенильные, имматурные, виргинильные и генеративные растения. Поскольку онтогенетические спектры не содержат информацию о проростках и растениях постгенеративного периода, для анализа онтогенетических спектров использовался только индекс восстановления (Glotov, 1998), рассчитанный как отношение суммы прегенеративных (без проростков) к сумме прегенеративных и генеративных растений. Считали, что при значении индекса восстановления больше 0.5 ЦП способна к самообновлению и самовосстановлению численности, а при значении меньше 0.5 находится в критическом состоянии.
На первом этапе проводили анализ гетерогенности субвыборок из ЦП с помощью критерия χ2 Пирсона. Затем для каждой площадки в пределах ЦП был вычислен индекс восстановления. По данному параметру проводили дисперсионный анализ и множественное попарное сравнение с помощью критерия Шеффе. Количество итераций составило 10 000. Достоверными приняты различия выборок при p ≤ 0.05. Для оценки доли влияния изменчивости между онтогенетическими спектрами ЦП в общей изменчивости проводили дисперсионный анализ, используя бутстреп. При анализе данных использовали среду R (Statisticheskaya…, 2019) и реализованную в ней программу OntoParam (Ivanov, 2014). Для построения регрессионной модели был использован пакет Statistica 13.0 и программа PAST 3.13.
Латинские названия растений приведены согласно сводке С.К. Черепанова (Czerepanov, 1995).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Изученные сообщества с B. versicolor в Нижнем Поволжье произрастают чаще всего на склонах балок южной или восточной экспозиции, реже – на южных и северных склонах холмов, равнинных участках, днищах балок и лиманов, заливных пойменных лугах. Почвы – чернозем южный, темно-каштановые и каштановые, в отдельных случаях – темно-каштановые карбонатные или ареносоли, последние порой с выходами песчаника. Анализ местообитаний по растительному покрову показал, что почвы чаще всего богатые, реже – довольно богатые или слабосолонцеватые. Аллювиальность их чаще слабая, чем умеренная. Увлажнение происходит за счет атмосферных осадков, изредка за счет паводковых вод. Древесно-кустарниковые растения, как правило, в сообществах с B. versicolor не встречаются. Лишь в ЦП 5, располагающейся в непосредственной близости от зарослей кустарников и дубравы, обнаружены единичные экземпляры Quercus robur (подрост), Prunus spinosa и Rosa sp. В ЦП 21 и 22 присутствовали единично Spiraea crenata и Rosa sp. Полукустарнички представлены Artemisia santonica, Kochia prostrata, Ephedra distachya, Astragalus ucrainicus и A. varius. Общее проективное покрытие трав и полукустарничков в большинстве сообществ колеблется от 45 до 95%, снижаясь в некоторых сообществах в отдельные годы до 20–40% вследствие степных пожаров, сенокоса или интенсивного выпаса. В разные годы в 70–80% сообществ доминировала B. versicolor, в 50–60% сообществ доминантом являлась Festuca valesiaca, в несколько меньшем числе сообществ доминировали Poa bulbosa, Stipa capillata, Elytrigia repens, Agropyron cristatum, Galatella villosa, Artemisia aust-riaca, в некоторых – Koeleria cristata, Gagea pusilla, в единичных сообществах в число доминантов и содоминантов входили еще до 30 видов злаков и разнотравья (табл. 1).
На территории исследования ЦП B. versicolor зачастую существуют в виде разобщенных локусов или единичных растений. За пределами Нижнего Поволжья популяции брандушки значительной площади (10–30 и более га) известны в Воронежской области (Agafonov et al., 2009) и на Украине (Melnik et al., 2007). Из наблюдаемых нами ЦП только в Ровенском районе Саратовской области B. versicolor относительно равномерно произрастает на площади более 50 га по периферии пересыхающего лимана со слабосолонцеватыми почвами.
ЦП B. versicolor зачастую проявляют существенную гетерогенность онтогенетических спектров субвыборок и, соответственно, по индексу восстановления. Так при анализе с помощью критерия χ2 выборок ЦП, изученных в 2018 г., гетерогенными оказались 11 из 18 ЦП. Гетерогенность онтогенетических спектров достоверно не подтверждена только для ЦП 4, 5, 13, 16, 17, 19 и 21. Гетерогенность субвыборок в пределах одной ЦП отражает неоднородность ее пространственной структуры, связанную с особенностями семяношения, микрорельефом и структурой фитоценоза.
Значения индекса восстановления в разных ЦП также достоверно различаются (табл. 2). Доля влияния межпопуляционной изменчивости индекса восстановления в общей изменчивости всех онтогенетических спектров составила 0.7.
Таблица 2.
Средняя плотность особей и индекс восстановления в ценопопуляциях Bulbocodium versicolor Нижнего Поволжья в 2018 г. Table 2. Average population density and recovery indices in the Lower Volga cenopopulations of Bulbocodium versicolor in 2018
| № ЦП № of CP |
Число растений на площадке (min-max), экз. Number of plants on site (min-max), pcs |
Средняя плотность особей на площадке, экз./м2 Average density of individuals on site, pcs/m2 |
Среднее значение индекса восстановления
и 95% доверительный интервал среднего Average recovery index and 95% confidence interval average |
|---|---|---|---|
| 3 | 82–114 | 97.8 ± 4.41 | 0.58–0.72–0.85 |
| 4 | 1–19 | 8.3 ± 2.04 | 0.11–0.20–0.30 |
| 5 | 2–6 | 3.2 ± 0.72 | 0.19–0.29–0.44 |
| 6 | 20–187 | 97.9 ± 17.60 | 0.74–0.81–0.87 |
| 7 | 7–285 | 89.8 ± 30.03 | 0.72–0.78–0.84 |
| 8 | 58–161 | 89.4 ± 12.36 | 0.83–0.87–0.91 |
| 10 | 7–43 | 26.8 ± 6.08 | 0.35–0.50–0.65 |
| 11 | 6–113 | 44.2 ± 14.89 | 0.32–0.40–0.49 |
| 13 | 10–53 | 18.5 ± 3.98 | 0.56–0.69–0.80 |
| 14 | 22–194 | 97.4 ± 18.20 | 0.73–0.78–0.83 |
| 15 | 6–77 | 64.3 ± 8.16 | 0.41–0.47–0.53 |
| 16 | 3–15 | 7.0 ± 1.32 | 0.12–0.23–0.34 |
| 17 | 3–36 | 13.4 ± 3.39 | 0.54–0.69–0.82 |
| 18 | 19–87 | 53.9 ± 6.64 | 0.23–0.33–0.43 |
| 19 | 18–64 | 34.9 ± 4.20 | 0.21–0.28–0.36 |
| 20 | 7–103 | 39.2 ± 10.28 | 0.31–0.38–0.46 |
| 21 | 10–37 | 19.9 ± 2.62 | 0.09–0.15–0.20 |
| 23 | 19–85 | 50.3 ± 6.62 | 0.37–0.45–0.53 |
Онтогенетические спектры большинства ЦП имеют хорошо выраженный пик на численности генеративных особей (рис. 3а). Доля генеративных растений составляет при этом более 40%. В ЦП 6, 7, 13 и 14, находящихся на юге Приволжской возвышенности, пик приходится на ювенильные особи (рис. 3b). В ЦП 3, 8 и 16 преобладают растения имматурного возрастного состояния (рис. 3c).
Рис. 3.
Онтогенетические спектры некоторых ценопопуляций Bulbocodium versicolor Нижнего Поволжья в 2018 г.: a – с пиком на генеративных особях, b – с пиком на ювенильных особях, c – с пиком на имматурных особях. По оси ординат – доля особей в процентах; по оси абсцисс – онтогенетические группы.
Fig. 3. Ontogenetic spectra of some Bulbocodium versicolor cenopopulations of the Lower Volga in 2018: a – with a peak on generative individuals, b – with a peak on juvenile individuals, c – with a peak on immature individuals. Y-axis – the percentage of individuals; X-axis – ontogenetic groups.
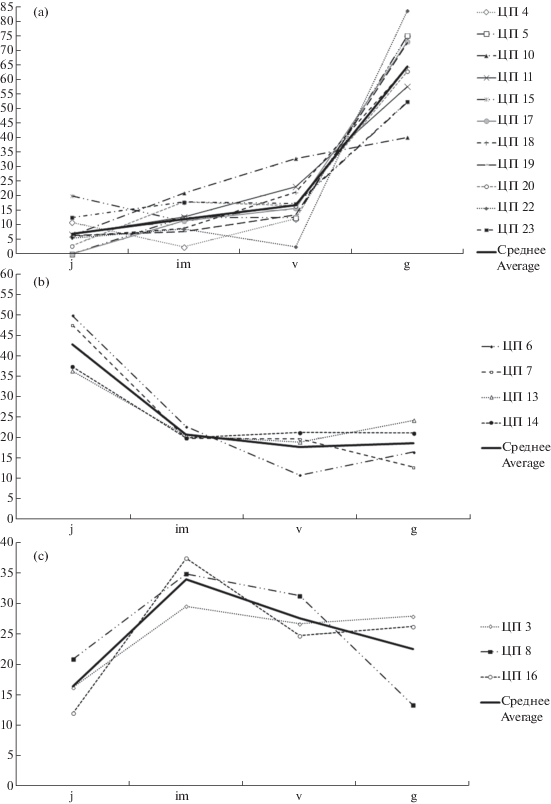
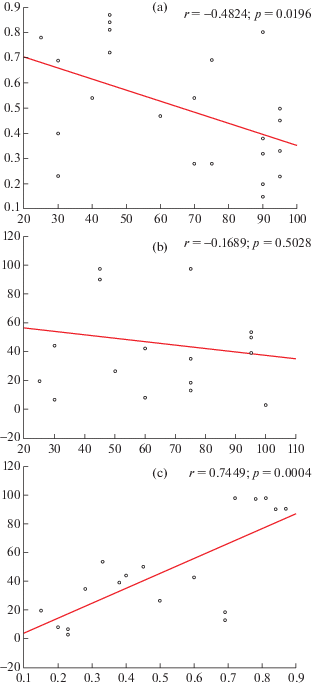
Рис. 4.
Корреляционные отношения индекса восстановления и общего проективного покрытия (a), общего проективного покрытия и средней плотности растений (b), средней плотности растений и индекса восстановления (c) в ценопопуляциях Bulbocodium versicolor Нижнего Поволжья, исследованных в 2018 г.
Fig. 4. Correlations between the restoration index and total projective cover (a), total projective cover and average plant density (b), average plant density and restoration index (c) in cenopopulations of Bulbocodium versicolor of the Lower Volga region studied in 2018.
Индекс восстановления ЦП B. versicolor в 2018 г. варьировал от 0.15 до 0.87 (табл. 2). Минимальный индекс восстановления имели ЦП 5 и 21 с низкой общей плотностью растений на сильно задернованных участках с густой ветошью. Максимальный индекс восстановления был характерен для ЦП, в которых максимум в онтогенетическом спектре приходился на ювенильные (ЦП 6, 7) или одновременно на имматурные и виргинильные особи (ЦП 8). В половине исследованных ЦП индекс восстановления был равен или превосходил значение 0.5, что свидетельствует о возможности их самовосстановления.
Попарные сравнения индекса восстановления на основе теста Шеффе (табл. 3), рассчитанного для ЦП B. versicolor Нижнего Поволжья в 2018 г. показали, что статистически значимо индекс восстановления различался в 89 парах, то есть исследованные популяции вида характеризуются значительной неоднородностью этого параметра. При этом для ЦП 5 и 21 с низкой общей плотностью растений на сильно задернованных участках с густой ветошью характерно максимальное число парных сочетаний с другими ЦП, по которым индекс восстановления достоверно отличается.
По нашему мнению, наиболее вероятной причиной низких значений индекса восстановления в ЦП B. versicolor является слабая конкурентоспособность растений ранних стадий развития. Известно, что семена этого вида прорастают только после длительной стратификации (Kulikova et al., 2017). При этом прорастание семян подземное. Семядольный листок отмирает в июне, в почве формируется небольшая луковица. На следующий год растение уже ювенильного онтогенетического состояния дает один узколинейный нитевидный лист длиной 2–3 см и формирует клубнелуковицу длиной 0.3–0.4 см и диаметром 0.5–0.8 см (Melnik et al., 2006а, b; 2007). Именно на этой стадии, растения, характеризующиеся слабым развитием листа и клубнелуковицы, вероятно, являются наиболее уязвимыми, по крайней мере, по отношению к сильной задернованности места произрастания и густой ветоши. На третий год растения переходят в имматурное состояние, в котором они имеют уже более сильное развитие вегетативных органов (лист 4–6 см дл., клубнелуковица 1.0–1.5 см в диам. (Melnik et al., 2006а, b; 2007).
При снижении конкурентных отношений, например после пожара, интенсивного выпаса или сенокоса (ЦП 6, 7, 13, 14) или произрастания на слаборазвитых почвах (ЦП 3, 8, 16), доля прегенеративных особей B. versicolor существенно возрастает. В зарастающих сообществах с высоким общим проективным покрытием и густой ветошью (ЦП 1, 4, 5) доля растений ранних онтогенетических состояний и средняя плотность растений в них снижаются.
Положительное влияние многолетнего выкашивания травостоя и умеренного выпаса на численность B. versicolor отмечалось и ранее (Sokolov et al., 2011; Rodіnka, Shevchenko, 2014; Solnyshkina, 2018). Неконтролируемые весенние палы степей ведут к резкому снижению численности вида и к замедлению темпов его развития (Shelegeda, Shelegeda, 2001; Kuznetsov et al., 2011; Sokolov et al., 2011). Однако если пал возник до начала вегетации, то в онтогенетическом спектре B. versicolor весной максимум приходился на ювенильные и виргинильные особи при уменьшении их общей плотности (Krivoruchko, 2008).
Результаты нашего исследования также указывают на положительное влияние палов в период покоя B. versicolor на состояние его популяций. При этом снижение общего проективного покрытия в сообществе ведет к увеличению доли растений прегенеративного периода и средней плотности растений B. versicolor. Различие состоит в том, что в случае разового воздействия на ЦП, приводящего к резкому снижению общего проективного покрытия в сообществе (осенний пожар, сенокос в предыдущий наблюдению год), происходит, прежде всего, увеличение численности растений B. versicolor ювенильного состояния (рис. 3b), в то время как постоянное произрастание в условиях низкого общего проективного покрытия (на слаборазвитых почвах) приводит к большей доле растений B. versicolor имматурного и виргинильного состояний (рис. 3c).
Результаты исследования ЦП B. versicolor в 2018 г. показали наличие умеренной отрицательной корреляции (r = –0.49) между общим проективным покрытием и индексом восстановления (рис. 4а), в то время как между плотностью растений и общим проективным покрытием корреляция не выявлена (рис. 4b). Это указывает на зависимость доли растений прегенеративного периода в ЦП от общего проективного покрытия, т. е. свидетельствует о низкой конкурентной способности растений этого онтогенетического периода. Высокая положительная корреляция выявлена между средней плотностью растений на учетной площадке и индексом восстановления (r = = 0.74) (рис. 4с), из чего следует, что средняя плотность растений B. versicolor определяется количеством особей прегенеративного периода.
Учитывая выявленную гетерогенность ЦП B. versicolor по индексу восстановления и существенную межпопуляционную изменчивость онтогенетической структуры в Нижнем Поволжье в один год наблюдения, был проведен мониторинг онтогенетической структуры и гетерогенности по индексу восстановления в течение 5–6 лет в ЦП Саратовской обл. (ЦП 3–8, 11) (рис. 5).
Рис. 5.
Средняя плотность растений различных онтогенетических состояний и индекс восстановления на постоянных мониторинговых площадках в ценопопуляциях Bulbocodium versicolor из Саратовской области в 2014–2019 гг.: по оси абсцисс – годы мониторинга, по левой оси ординат – плотность особей (экз./м2), по правой оси ординат – индекс восстановления. a – ЦП 3; b – ЦП 4; c – ЦП 5; d – ЦП 6; e – ЦП 7; f – ЦП 8; g – ЦП 11; j – ювенильные, im – имматурные, v – виргинильные, g – генеративные особи.
Fig. 5. The average density of plants of various ontogenetic states and the recovery index at constant monitoring sites in Bulbo-codium versicolor cenopopulations from the Saratov Region in 2014–2019: X-axis – years of monitoring, left Y-axis – the density of individuals (pcs/m2), right Y-axis – recovery index. a – CP 3; b – CP 4; c – CP 5; d – CP 6; e – CP 7; f – CP 8; g – CP 11; j – juvenile, im – immature, v – virginile, g – generative individuals.
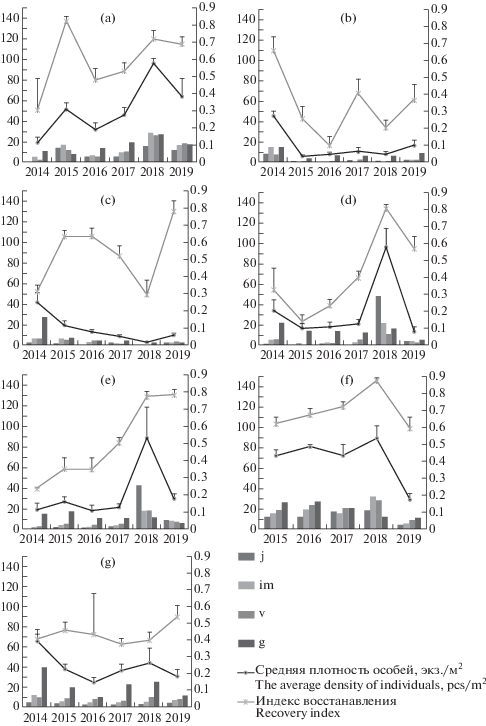
ЦП 3 расположена на крутом склоне холма южной экспозиции. Увлажнение сухостепное, от умеренного до сильно переменного. Почва – ареносоль с выходами песчаника, богатая, слабо аллювиальная. Испытывает умеренное влияние пастбищной дигрессии.
C использованием критерия χ2 показано, что субвыборки онтогенетических спектров в 2014, 2018 и 2019 гг. гетерогенны при уровне значимости p ≤ 0.005. Для выборок 2015–2017 гг. гетерогенность не установлена. ЦП 3 отличается высокой и стабильной способностью к самовосстановлению и высокой средней плотностью особей на 1 м2 (рис. 5а). В целом индекс восстановления здесь был выше 0.5, достигая в 2015 г. 0.8, а в 2018 г. – 0.7. Исключение составил только 2014 г., когда индекс восстановления был отмечен на уровне 0.35. Это связано, вероятно, с тем, что она находится на слаборазвитых почвах (ареносоли с выходом песчаника) и отличается относительно низким уровнем общего проективного покрытия во все годы мониторинга. Как следствие, растения вида имеют конкурентное преимущество, сохраняя год от года относительно высокую плотность и большую долю в онтогенетическом спектре растений прегенеративного периода. Низкий уровень средней плотности растений вида в ЦП в 2014 г. может быть связан с тем, что в августе – сентябре предыдущего года место расположения ЦП характеризовалось фактическим отсутствием осадков (рис. 2a) при нахождении ЦП в условиях сухостепного увлажнения, что и могло привести к круглогодичному покою части клубнелуковиц особей разных онтогенетических состояний весной 2014 г.
ЦП 4 расположена на пологом склоне южной экспозиции. Увлажнение средне- и сухостепное, умеренно переменное. Почва – чернозем солонцеватый, богатая, слабо аллювиальная. Испытывает умеренное влияние пастбищной дигрессии.
В 2018 и 2019 гг. субвыборки особей были гетерогенными при p ≤ 0.005. Для выборок 2014–2017 гг. гетерогенность не установлена. Индекс восстановления ЦП 4 различных лет наблюдения достоверно различим (при p = 0.0035). Доля вклада изменчивости между выборками в общую изменчивость составляет 0.34. Парные сравнения индекса восстановления, основанные на индексе Шеффе, показали, что из 15 возможных пар семь пар имеют достоверные отличия. В целом для ЦП 4 (рис. 5b) в годы мониторинга характерно колебание общего проективного покрытия на довольно высоком уровне (зарастание злаками и другими видами). Последствием этого является снижение уровня и усиление нестабильности индекса восстановления, а также последовательное снижение средней плотности растений в ЦП.
ЦП 5 приурочена к склону северной экспозиции. Увлажнение лугово-степное (влажностепное), умеренно переменное. Почва – чернозем южный, довольно богатая, умеренно аллювиальная. Испытывает умеренное влияние пастбищной дигрессии.
Достоверная гетерогенность онтогенетических спектров выборок данной ЦП не была выявлена. Однако значения индекса восстановления разных лет наблюдения достоверно различались (при р = 0.0001). Доля влияния изменчивости индекса восстановления между годами наблюдения в общей изменчивости составила 0.6. В ЦП 5 (рис. 5с) средняя плотность растений на 1 м2 последовательно снижалась с 2014 по 2018 гг., что связано, вероятно, с интенсивным зарастанием местообитания B. versicolor разнотравьем и длиннокорневищными злаками. В 2019 г. средняя плотность растений в ЦП незначительно возросла до уровня 2017 г. Сравнительная однородность онтогенетических спектров субвыборок ЦП 5 может быть объяснена однородностью микрорельефа и структуры фитоценоза на всей площади ее произрастания в условиях влажностепного увлажнения на склоне северной экспозиции.
ЦП 6 расположена на равнине у подножия заброшенной железнодорожной насыпи. Увлажнение средне- и сухостепное, умеренно и сильно переменное. Почва – чернозем южный, богатая и довольно богатая, слабо аллювиальная. Испытывает умеренное влияние пастбищной дигрессии.
Выборки 2014 и 2018 гг. были гетерогенными при p = 0.00099 и p = 2.2 × 10–16, соответственно. Для выборок 2015–2017 гг. гетерогенность не установлена. В 2015 г. в ЦП 6 (рис. 5d) наблюдалось снижение индекса восстановления по сравнению с 2014 г. и его показатель составил 0.14. С 2016 г. отмечается тенденция плавного увеличения индекса восстановления. В 2018 г. он составил 0.81, а в остальные годы находился в диапазоне от 0.14 до 0.39. Осенний пожар в предыдущий год стал причиной увеличения численности ювенильных и, в меньшей мере, имматурных и виргинильных, и даже генеративных особей, что привело к резкому увеличению средней плотности растений в 2018 г., а это сказалось на существенном изменении ее онтогенетической структуры. Однако всплеск численности в 2018 г. особей прегенеративной группы, которая является наиболее уязвимой к действию факторов окружающей среды, не обеспечил ни роста, ни сохранения численности особей данной ЦП в 2019 г. Напротив, в 2019 г. плотность особей всех онтогенетических состояний даже резко снизилась (до уровня, уступающего таковой в 2014–2017 гг.). Это может быть объяснено тем, что в ЦП вида колебание численности особей зачастую связано со способностью клубнелуковицы любого онтогенетического состояния переходить в состояние круглогодичного покоя и, соответственно, выходить из него через разное количество лет (Melnik et al., 2007). Высокое количество осадков в апреле–мае–июне 2017 г. (рис. 2b) могло сказаться на выходе из круглогодичного покоя клубнелуковиц разного онтогенетического состояния и их активной вегетации в 2018 г. на освободившемся после пожара пространстве. Чрезвычайно малое количество осадков в период активного роста и развития проростков B. versicolor в апреле–мае–июне 2018 г. и быстрое восстановление надземной вегетативной массы видов сообщества привело или к сильному выпаду, или, что более вероятно, к переходу части особей B. versicolor всех онтогенетических состояний к круглогодичному покою и снижению индекса восстановления в 2019 г. до 0.64.
ЦП 7 находится на склоне балки южной экспозиции. Увлажнение среднестепное, умеренно переменное. Почва – чернозем южный, богатая и довольно богатая, слабо и умеренно аллювиальная. Испытывает умеренное влияние пастбищной дигрессии.
Гетерогенность субвыборок была достоверной только для выборки 2018 г. Однако индекс восстановления ЦП различных лет наблюдения оказался достоверно различим. Доля вклада изменчивости между выборками в общую изменчивость составила 0.74. В ЦП 7 колебание индекса восстановления в период с 2014 по 2017 гг. было незначительным и имело тенденцию увеличения с 0.24 до 0.50. Затем отмечен его резкий скачок в 2018 г. до 0.83. Высоким, в отличие от ЦП 6, он оставался и в 2019 г. (рис. 5e), однако, как и в ЦП 6, средняя плотность растений в ЦП 7 в 2019 г. резко снизилась до уровня, уступающего таковому в 2014–2017 гг., хотя доля растений прегенеративного периода в онтогенетическом спектре оставалась высокой. Осенью 2017 г. эта ЦП также подверглась степному пожару, с чем, вероятно, и связан всплеск численности особей ювенильного периода в 2018 г. Обе ЦП расположены относительно недалеко друг от друга, близки друг к другу по экологическим условиям. Различие в динамике их онтогенетической структуры, но не в средней плотности растений, в период с 2018 по 2019 г. может быть связано с особенностями микрорельефа: ЦП 7 находится на склоне балки, а ЦП 6 – на выровненном участке. Соответственно первая из них в 2019 г. имела среднестепное (не имела дефицита почвенной влаги), а вторая – сухостепное увлажнение, испытывая дефицит почвенной влаги.
ЦП 8 расположена на днище балки. Увлажнение сухостепное, от умеренного до сильно переменного. Почва – ареносоль, слабо или умеренно аллювиальная. Испытывает умеренное влияние пастбищной дигрессии.
Использование критерия χ2 показало гетерогенность субвыборок для выборок всех лет мониторинга при p ≤ 0.005. Индекс восстановления ЦП 8 различных лет наблюдения также достоверно различим. Доля вклада изменчивости между выборками в общую изменчивость составляет 0.37. Колебание индекса восстановления и средней плотности растений в ЦП 8 в различные годы мониторинга незначительно, а уровень их во все годы, кроме 2019 г., высок (рис. 5f). При этом наблюдается незначительный, но устойчивый рост индекса восстановления с 2014 по 2018 гг. Однако в 2019 г. индекс восстановления снизился до уровня, несколько уступающего даже уровню 2014 г., хотя и оставался выше 0.5. Средняя плотность растений после повышения в 2018 г. с 70 до 90 экз./м2 на следующий год резко (в три раза) снизилась (с 90 до 30 экз./м2). За годы мониторинга данная ЦП была самой стабильной по параметрам средней плотности растений и индексу восстановления. Это, как и в случае с ЦП 3, связано с ее расположением на слаборазвитых почвах (ареносоли) при низком уровне общего проективного покрытия в сообществе (50%) (рис. 2c). В 2019 г. снижение плотности и индекса восстановления произошло за счет резкого снижения доли растений прегенеративного периода, скорее всего, по причине засухи в апреле–мае–июне 2018 г. К 2019 г. значительная часть растений прегенеративного периода или элиминовала, или осталась в состоянии покоя.
ЦП 11 расположена на днище лимана. Увлажнение лугово-степное (влажностепное), сильно переменное. Почва каштановая солонцеватая, слабо аллювиальная. Испытывает умеренное влияние пастбищной дигрессии.
Все субвыборки этой ЦП оказались гетерогенными (р ≤ 0.05). Однако парные сравнения индекса восстановления, основанные на тесте Шеффе, достоверных различий между выборками не показали. Оценка доли влияния изменчивости между выборками составила 0.09. Таким образом, несмотря на гетерогенность выборок, индекс восстановления этой ЦП в разные годы достоверно не различался и составлял около 0.4–0.5. ЦП 11 существует при равномерной пастбищной нагрузке, других существенных внешних воздействий разового характера, кроме разной степени затопления паводковыми водами, за годы мониторинга не наблюдалось. Для фитоценоза характерна пространственная неоднородность, связанная с микропонижениями и, соответственно, с неравномерностью затопления при наполнении лимана вешними водами, которая и привела к пространственной гетерогенности распределения особей B. versicolor. Общее проективное покрытие составляет 30–50%, при практическом отсутствии ветоши, что говорит о слабой задернованности ЦП и благоприятных условиях для развития особей прегенеративной группы, так как растения именно этой группы не способны конкурировать с другими растениями сообщества. В связи с этим ЦП является относительно стабильной: в ней около половины особей относятся к прегенеративному периоду. При этом средняя плотность особей на 1 м2 также сохраняет относительную стабильность во все годы наблюдения на уровне 30–40 экз.м2, за исключением 2014 г., когда она достигала 60 и более особей на 1 м2 (рис. 5g). Высокая средняя плотность растений в 2014 г. может быть связана с высоким уровнем осадков в июле и сентябре предыдущего года, что могло спровоцировать выход из покоя клубнелуковиц растений генеративного периода, пребывавших в покоящемся состоянии.
Как видно, возможны разные сценарии изменения онтогенетической структуры ЦП B. versicolor в зависимости от складывающихся в них экологических условий и особенностей мест произрастания. Плотность пространственного распределения особей разных онтогенетических состояний также существенно различается и кардинально изменяется при изменении условий произрастания, например, при зарастании, после пожара или сенокоса. В отдельные годы всходы B. versicolor могут не появляться вовсе. Массовое их выпадение или переход в состояние круглогодичного покоя происходит, например, при наступлении ранневесенней засухи или угнетении другими растениями при сильной задернованности участка. В отсутствии изменения внешних факторов, воздействующих на ЦП, их онтогенетические спектры остаются относительно стабильными и обладают слабой степенью динамичности.
Лабильность популяционных показателей B. versicolor указывает на то, что он является реактивным эксплерентом, способным очень быстро захватывать освободившиеся площади, но и освобождать их при вытеснении конкурентными видами. Этот вид способен массово развиваться на территориях с катастрофически снизившимся общим проективным покрытием благодаря запасу семян и покоящихся клубнелуковиц в почве.
ВЫВОДЫ
Онтогенетические спектры субвыборок B. versicolor чаще всего неоднородны в пределах выборки (ценопопуляции), что связано с неоднородностью пространственного размещения особей по причинам особенности распространения семян, неоднородности микрорельефа или фитоценоза и требует адекватного статистического анализа.
Онтогенетические спектры ЦП B. versicolor, как правило, имеют максимум на генеративных растениях. Редко встречаются спектры с пиком на растениях ювенильного онтогенетического состояния. В этом случае всплеск численности прегенеративных растений связан с нарушениями в фитоценозе, приводящими к низкому общему проективному покрытию. Пик на растениях имматурного и (или) виргинильного онтогенетического состояния свойственен ЦП, расположенным на слаборазвитых почвах, на которых сообщество также имеет низкое общее проективное покрытие.
Средняя плотность растений вида в ЦП определяется теми же экологическими факторами и той же направленности, что и формирование ее онтогенетической структуры. При зарастании фитоценоза длиннокорневищными злаками и последовательном увеличении общего проективного покрытия B. versicolor выпадает из сообщества.
Погодные условия, прежде всего, в период активной вегетации, цветения, формирования и перехода в состояние покоя клубнелуковиц B. versicolor, сказываются на численности, плотности и соотношении особей различного онтогенетического состояния в последующий год. Условия сильной засухи в этот период провоцируют переход растений разного онтогенетического состояния в круглогодичный покой, а большое количество осадков – приводит к выходу из состояния круглогодичного покоя клубнелуковиц, пребывавших в нем в предыдущий год (или годы).
Список литературы
[Agafonov et al.] Агафонов В.А., Кузнецов Б.И., Негробов В.В. 2009. К характеристике растительного покрова степных местообитаний брандушки разноцветной (Bulbocodium versicolor (Ker-Gawl.) Spreng.) в Воронежской области. – Поволжский экологический журнал. 3: 258–262.
[Chervona…] Червона книга України. Рослинний світ. 2009. Київ. 912 с.
Conti F., Aurelio M., Franco P. 1997. Liste Rosse Regio-nalli delle Piante d’Italia. Camerino. 139 p.
Conti F., Bartolucci F. 2012. Specie a rischio in Abruzzo. Elenco delle piante di interesse conservazionistico. – In: La Biodiversità vegetale in Abruzzo. Tutela e conservazione del patrimonio vegetale abruzzese. Abruzzo. P. 81–109.
Csapody I. 1982. Védett növényeink. Budapest. 349 p.
[Czerepanov] Черепанов С.К. 1995. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). СПб. 992 с.
[Glotov] Глотов Н.В. 1998. Об оценке параметров возрастной структуры популяций растений. – В сб.: Жизнь популяций в гетерогенной среде. Ч. 1. Йошкар-Ола. С. 146–149.
[Goncharenko, Karpenko] Гончаренко І.В., Карпенко К.К. 1998. Брандушка різнокольорова (Bulbo-codium versicolor (Ker.-Gawl.) Spreng.) на Сумщині. – В сб.: Вакалівщина. До 30-річчя біологічного стаціонару Сумського педінституту: Збірник наукових праць. Суми. С. 211–215.
[Grokhlina, Khanina] Грохлина Т.И., Ханина Л.Г. 2015. О компьютерной обработке геоботанических описаний по экологическим шкалам. – В сб.: Математическое моделирование в экологии. Материалы четвертой национальной научной конференции с международным участием. Пущино. С. 63–64.
[Ivahov] Иванов С.М. 2014. Анализ онтогенетических спектров гетерогенных популяций: Дис. … канд. биол. наук. Нижний Новгород. 116 с.
[Krasnaya…] Красная книга Российской Федерации (растения и грибы). 2008. М. 855 с.
[Krasnaya…] Красная книга Саратовской области: Грибы. Лишайники. Растения. Животные. 2006. Саратов. 528 с.
[Krivoruchko] Криворучко Т.В. 2005. Особливості поширення, вікові спектри та морфологічні показники ценопопуляцій Bulbocodium versicolor (Ker.-Gavl.) Spreng. деяких заповідних територій у межах Полтавської області. – Збірник наук. праць Полтавського держ. пед. ун-ту ім. В.Г. Короленка. Серія “Екологія. Біологічні науки”. 4 (43): 45–52.
Kulikova L.V., Kashin A.S., Shilova I.V., Petrova N.A. 2019. Some ecological peculiarities of Bulbocodium versicolor (Ker-Gawl.) Spreng. (Colchicaceae, Magnoliophyta) in the Lower Volga region. – Biol. Bull. 47 (10): 96–103.
[Kulikova et al.] Куликова Л.В., Шилова И.В., Серова Л.А., Кашин А.С. 2017. Особенности прорастания семян брандушки разноцветной (Bulbocodium versicolor (Ker-Gavel.) Spreng.) в лабораторных условиях. – Бюл. бот. сада Сарат. гос. ун-та. 15 (2): 53–57. https://doi.org/10.18500/1682-1637-2017-15-2-53-57
[Kuznetsov et al.] Кузнецов Б.И., Моисеева Е.В., Глазнева О.С. 2011. Семенная продуктивность ранневесенних степных эфемероидов на примере Bulbocodium versicolor (Ker-Gavel.) Spreng. и Bellevalia sarmatica (Pall. ex Georgi) в природных условиях и в культуре. – Вестн. Воронежск. гос. ун-та. Серия: География. Геоэкология. 2: 104–106.
[Melnik et al.] Мельник В.И., Гриценко В.В., Шевченко Д.Ю. 2006а. Ареал, эколого-ценотические условия местообитаний и структура популяций Bulbo-codium versicolor (Ker.-Gavl.) Spreng. (Melanthiaceae). – Інтродукція рослин. 1: 23–35.
[Melnik et al.] Мельник В.И., Гриценко В.В., Шевченко Д.Ю. 2006b. Bulbocodium versicolor (Melanthiaceae) в Украине. – Бот. журн. 91 (10): 88–97.
[Melnik et al.] Мельник В.И., Гриценко В.В., Шевченко Д.Ю., Диденко С.Я. 2007. Bulbocodium versicolor (Melanthiaceae) – редкий вид флоры Европы. Киев. 44 с.
[Metodychni…] Методичні рекомендації щодо вивчення популяцій брандушки різнобарвної Bulbocodium versicolor (Ker Gawl.). 2016. Підготовлено Драбинюк Г.В. http://www.pryingul.inf.ua/articles/MR-brandushka_2016.pdf
Oltean M., Negreanu G., Popescu A., Roman N., Dihoru G., Sanda V., Mihailescu S. 1994. Lişta rosie a plantelor superioare din România. – In: Studii, Sinteze, Documentaţii de Ecologie. Vol. 1. Timişoara. P. 1–52.
[Petrova et al.] Петрова Н.А., Шилова И.В., Кашин А.С., Березуцкий М.А., Серова Л.А., Решетникова Т.Б. 2015. О распространении брандушки разноцветной в Саратовской области. – Бюл. бот. сада Сарат. гос. ун-та. 13: 25–31.
[Pogrebennik et al.] Погребенник В.П., Кучерява Л.Ф., Нечитайло В.А., Хоменко Ж.И. 1987. Редкие и исчезающие растения в урочище “Шандровский лес”. – В сб.: Охрана, изучение и обогащение растительного мира. Вып. 14. Киев. С. 13–18.
[Pravila…] Правила сбора редких и находящихся под угрозой исчезновения видов растений (для ботанических садов). 1981. – Бюл. ГБС. 119: 94–96.
[Ramenskii et al.] Раменский Л.Г., Цаценкин И.А., Чижикова О.Н., Антипин H.A. 1956. Экологическая оценка кормовых угодий по растительному покрову. М. 471 с.
[Sagalaev, Shantser] Сагалаев В.А., Шанцер И.А. 2006. Сем. 36. Colchicaceae DC. – Безвременниковые. – В кн.: Флора Нижнего Поволжья. Т. 1. М. С. 331–333.
[Shelegeda, Shelegeda] Шелегеда В., Шелегеда Е. 2001. Экспедиция “Первоцветы Запорожья”: Атлас – справочник для экологических некоммерческих организаций. Запорожье. 92 с.
[Sokolov et al.] Соколов И.Д., Соколова Е.И., Бутылкина Н.Ю., Бережной М.В. 2011. Классификация факторов, влияющих на численность наземных растений из Красной книги Украины (на примере Bulbocodium versicolor (Ker Gawl.) Spreng и Tulipa quercetorum Klokov et Zoz). – Науковий вісник ЛНАУ. Серія Біологічні науки. 28: 92–96.
[Solnyshkina] Солнышкина Е.Н. 2018. Брандушка разноцветная Bulbocodium versicolor (Ker-Gavel.) Spreng. на участке Ямская степь Государственного природного заповедника “Белогорье”. – В сб.: Региональные ботанические исследования как основа сохранения биоразнообразия. Материалы Всероссийской (с международным участием) научной конференции. Воронеж. С. 152–155.
Sramkó G., Gulyás G., Matus G., Rudnóy Sz., Illyés Z., Bratek Z., Molnár A. V. 2008. Leaf width, nrDNA and cpDNA its sequence variation within Central European Bulbocodium vernum and B. versicolor (Colchicaceae) populations: are there really two taxa? – Acta Biol. Hung. 59 (1): 103–114. https://doi.org/10.1556/ABiol.59.2008.1.9
[Statisticheskaya…] Статистическая среда R. https://cran.r-project.org (Дата обращения: 26.11.2019).
[Tsenopopulyatsii…] Ценопопуляции растений (основные понятия и структура). 1976. М. 217 с.
[Tsenopopulyatsii…] Ценопопуляции растений (развитие и взаимоотношения). 1977. М. 131 с.
[Tsenopopulyatsii…] Ценопопуляции растений (очерки популяционной биологии). 1988. М. 184 с.
[Tzvelev] Цвелев Н.Н. 1979. Брандушка – Bulbocodium L. – В кн.: Флора Европейской части СССР. Т. 4. Л. С. 218.
Valentine D.H. 1980. Bulbocodium L. – In: Flora Europaea. Vol. 5. Cambridge. P. 25.
Vinnersten A., Reeves G. 2003. Phylogenetic relationships within Colchicaceae. – Am. J. Bot. 90 (10): 1455–1462.
[Zhukova et al.] Жукова О.В., Иванов С.М., Глотов Н.В. 2014. Онтогенетические спектры ценопопуляций Alchemilla vulgaris L. s.l. – Вестн. Удмуртск. ун-та: Биология. Науки о Земле. 2: 14–20.
[Zubkova et al.] Зубкова Е.В., Ханина Л.Г., Грохлина Т.И., Дорогова Ю.А. 2008. Компьютерная обработка геоботанических описаний по экологическим шкалам с помощью программы EcoScaleWin. Учебное пособие. Йошкар-Ола. 96 с.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал