

Ботанический журнал, 2020, T. 105, № 5, стр. 438-457
МЕЗОТРОФНЫЕ ЕЛЬНИКИ ХВОЩОВЫЕ, КРУПНОПАПОРОТНИКОВО- И ХМЕЛЕНИЧНО-ХВОЩОВЫЕ СФАГНОВЫЕ СРЕДНЕЙ И ЮЖНОЙ ТАЙГИ ЕВРОПЕЙСКОЙ РОССИИ И УРАЛА
И. Б. Кучеров 1, *, **, С. А. Кутенков 2, ***
1 Ботанический институт им. В.Л. Комарова Российской академии наук
197376 Санкт-Петербург, ул. Проф. Попова, 2, Российская Федерация
2 Институт биологии Карельского научного центра РАН
185910 Республика Карелия, г. Петрозаводск, ул. Пушкинская, 11, Российская Федерация
* E-mail: atragene@mail.ru
** E-mail: IKucherov@binran.ru
*** E-mail: effort@krc.karelia.ru
Поступила в редакцию 23.12.2019
После доработки 10.02.2020
Принята к публикации 18.02.2020
Аннотация
На основе 114 геоботанических описаний, сделанных авторами в 1995–2019 гг. или взятых из литературы, проведена доминантно-детерминантная классификация мезотрофных ельников хвощовых, крупнопапоротниково-хвощовых и хмеленично-хвощовых сфагновых с доминированием Picea abies s.l.., Equisetum sylvaticum и Sphagnum girgensohnii в Европейской России и на Урале. Выделены 2 ассоциации с 5 субассоциациями и 6 вариантами. Разнообразие синтаксонов обусловлено как широтной зональностью, так и изменениями континентальности климата, для субассоциаций и вариантов также различиями во внутриландшафтной приуроченности. Ельники хвощовые сфагновые типичной и щитовниковой (с согосподством Equisetum sylvaticum и Dryopteris expansa s.l.) субассоциаций произрастают от южных границ северной тайги до подтайги в западном и центральном секторах Европейской России, известны также в горах Средней Европы. В Предуралье и на Урале на фоне роста континентальности климата их замещают ельники и пихто-ельники хмеленично-хвощовые сфагновые c согосподством Equisetum sylvaticum и Rubus humulifolius. Леса всех синтаксонов развиваются на почвах умеренного минерального богатства на начальных этапах мезотрофного заболачивания. Ельники хвощовые и крупнопапоротниково-хвощовые тяготеют к почвам тяжелого гранулометрического состава, хмеленично-хвощовые – произрастают на почвах разного состава.
Настоящая публикация завершает начатый ранее (Kucherov, Kutenkov, 2020) обзор ценотического разнообразия мезотрофных ельников хвощовых сфагновых (с доминированием Picea abies s.l., Equisetum sylvaticum и Sphagnum girgensohnii в соответствующих ярусах сообщества) в Европейской России и на Урале. Она посвящена ельникам крупнопапоротниково-хвощовым и собственно хвощовым, свойственным в основном средней и южной тайге, а также хмеленично-хвощовым ельникам Двино-Печорского региона.
ИСПОЛЬЗОВАННЫЕ ДАННЫЕ И МЕТОДЫ
В основу работы положена выборка из 114 геоботанических описаний мезотрофных ельников хвощовых сфагновых Европейской России и Урала. Из них 82 описания выполнены авторами в экспедициях 1996–2019 гг. Как и при описании средне- и северотаежных ельников морошково-хвощовых сфагновых (Kucherov, Kutenkov, 2020), на площади не менее 400 м2 глазомерно оценивались проективные покрытия (ПП) видов по ярусам, выполнялась почвенная прикопка или измерялась глубина залегания торфа. Четыре описания из Республики Коми взяты из рукописи докторской диссертации Ю.П. Юдина (Yudin, 1948), хранящейся в библиотеке БИН РАН. Еще 4 описания из заповедника “Кологривский лес” в Костромской обл. получены по запросу из фитоценария ЦЭПЛ РАН (Zaugolnova, Khanina, 1996; Lugovaya, 2010). Оставшиеся 24 описания опубликованы в иных литературных источниках (Smirnova Z., 1928; Korchagin, 1929, 1940; Sambuk, 1930; Smirnova A., 1954; Nepomilueva, Duryagina, 1985).
Классификация растительности выполнена доминантно-детерминантным методом с выделением синтаксонов по доминантам и последующим уточнением их объема по детерминантным группам экологически близких видов. Критерии разграничения ассоциаций, субассоциаций и вариантов, а также ссылки на детальное изложение методики классификации приведены в предыдущей статье (Kucherov, Kutenkov, 2020).
Данные о 9 синтаксонах разного ранга, выделенных при классификации, содержатся в фитоценотической таблице (табл. 1), содержащей также сведения об общем покрытии и высоте ярусов. Названия синтаксонов даны согласно традиции школы В.Н. Сукачева (Sukachev, 1931). Названия в форме, принятой в работах по доминантно-детерминантной классификации, приводятся как синонимы (Kucherov, Kutenkov, 2020).
Таблица 1.
Фитоценотическая характеристика мезотрофных ельников хвощовых, щитовниково- и хмеленично-хвощовых сфагновых Европейской России и Урала Table 1. Phytocoenotic record of mesotrophic horsetail, shield-fern-horsetail, and hop-leaved raspberry-horsetail peatmoss spruce forests of European Russia and the Urals
| Вид/Параметр Species/Parameter |
Ярус Layer |
Синтаксоны/Syntaxa | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | ||
| Детерминанты субассоциаций и вариантов/Determinants of subassociations and variants | ||||||||||
| Pinus sylvestris | a1 | 412 | 405 | 565 | 557 | 6+ | 11+ | 252 | 9+ | |
| Calamagrostis arundinacea | c | 371 | 8011 | 671 | 18+ | 221 | 10+ | 18+ | ||
| Solidago virgaurea | c | 33+ | 1002 | 1002 | 36+ | 6+ | 44+ | 10+ | 17+ | 18+ |
| Dicranum polysetum | d | 412 | 801 | 441 | 731 | 13+ | 11+ | 30+ | 171 | 9+ |
| Cladonia coniocraea | z | 41+ | 80+ | 44+ | 82+ | 38+ | 11+ | 35+ | 36+ | |
| Plagiothecium laetum | z | 30+ | 60+ | 44+ | 55+ | 31+ | 14+ | 5+ | 27+ | |
| Ptilidium pulcherrimum | z | 33+ | 80+ | 22+ | 27+ | 31+ | 11+ | 15+ | 27+ | |
| Sciuro-hypnum reflexum | z | 15+ | 20+ | 22+ | 36+ | 6+ | 9+ | |||
| Luzula pilosa | c | 701 | 1001 | 1001 | 73+ | 751 | 22+ | 551 | 17+ | 45+ |
| Rubus saxatilis | c | 44+ | 1003 | 1002 | 36+ | 251 | 11+ | 35+ | 331 | 551 |
| Populus tremula | b | 33+ | 401 | 22+ | 9+ | 5+ | ||||
| Convallaria majalis | c | 7+ | 1001 | 11+ | 9+ | |||||
| Anemonoides nemorosa | c | 601 | 22+ | 181 | ||||||
| Galeobdolon luteum | c | 401 | ||||||||
| Plagiomnium affine | d | 40+ | 271 | |||||||
| Salix caprea | a + b | 22+ | 60+ | 33+ | 13+ | 22+ | 5+ | 17+ | ||
| Quercus robur | b | 20+ | 22+ | |||||||
| Deschampsia cespitosa | c | 11+ | 601 | 781 | 18+ | 19+ | 15+ | 27+ | ||
| Carex canescens | c | 7+ | 60+ | 22+ | 22+ | 27+ | ||||
| Calamagrostis canescens | c | 71 | 401 | 33+ | 18+ | 61 | 171 | |||
| Galium palustre | c | 40+ | 33+ | 6+ | 5+ | |||||
| Carex brunnescens | c | 4+ | 20+ | 221 | 9+ | |||||
| Agrostis canina | c | 40+ | 33+ | |||||||
| Rhytidiadelphus triquetrus | d | 221 | 1004 | 898 | 271 | 131 | 331 | 15+ | 335 | 279 |
| Sphagnum squarrosum | d | 11+ | 801 | 566 | 271 | 191 | 224 | 10+ | 271 | |
| Rhodobryum roseum | d | 4+ | 60+ | 441 | 27+ | 27+ | ||||
| Peltigera praetextata | z | 7+ | 40+ | 33+ | 9+ | |||||
| Frangula alnus | b | 151 | 60+ | 892 | 862 | 5+ | 172 | 9+ | ||
| Plagiochila asplenioides | d | 4+ | 806 | 676 | 45+ | 6+ | 10+ | |||
| Sanionia uncinata | z | 19+ | 80+ | 33+ | 45+ | 19+ | 33+ | 15+ | 27+ | |
| Tetraphis pellucida | z | 7+ | 60+ | 56+ | 55+ | 31+ | 11+ | 10+ | 9+ | |
| Hylocomium splendens | z | 11+ | 401 | 441 | 18+ | 311 | 112 | 5+ | 27+ | |
| Dicranum polysetum | z | 15+ | 40+ | 56+ | 55+ | 13+ | 11+ | 5+ | ||
| Rubus idaeus | b | 11+ | 401 | 56+ | 36+ | 381 | 5+ | 552 | ||
| Dryopteris expansa s.l. | c | 26+ | 1001 | 789 | 10014 | 502 | 10017 | |||
| Pleurozium schreberi | z | 15+ | 601 | 441 | 641 | 191 | 221 | 10+ | 641 | |
| Pohlia nutans | z | 19+ | 40+ | 44+ | 45+ | 19+ | 22+ | 55+ | ||
| Athyrium filix-femina | c | 4+ | 801 | 895 | 36+ | 6+ | 332 | 15+ | 642 | |
| Crepis paludosa | c | 20+ | 67+ | 11+ | 5+ | 17+ | 64+ | |||
| Plagiomnium ellipticum | d | 7+ | 40+ | 44+ | 18+ | 6+ | 22+ | 15+ | 454 | |
| Milium effusum | c | 7+ | 40+ | 11+ | 9+ | 19+ | 11+ | 5+ | 451 | |
| Stellaria holostea | c | 4+ | 601 | 27+ | 25+ | 5+ | 361 | |||
| Carex vaginata | c | 11+ | 20+ | 44+ | 9+ | 6+ | 11+ | 25+ | 9+ | |
| Circaea alpina | c | 4+ | 782 | 13+ | 5+ | 18+ | ||||
| Ranunculus repens | c | 20+ | 44+ | 6+ | 33+ | 5+ | 18+ | |||
| Lysimachia vulgaris | c | 7+ | 33+ | 18+ | ||||||
| Mercurialis perennis | c | 1115 | ||||||||
| Sphagnum wulfianum | d | 15+ | 202 | 441 | 181 | 61 | 441 | 209 | ||
| Dicranum scoparium | d | 221 | 201 | 22+ | 821 | 311 | 351 | 452 | ||
| Sciuro-hypnum curtum | d | 15+ | 33+ | 457 | 191 | 11+ | 5+ | 273 | ||
| Abies sibirica | a1 + 2 | 504 | 6720 | 647 | ||||||
| Pinus sibirica | a1 + 2 | 191 | 5+ | 675 | 453 | |||||
| Abies sibirica | b | 11+ | 813 | 11+ | 10+ | 10014 | 647 | |||
| Pinus sibirica | b | 19+ | 501 | 18+ | ||||||
| Phegopteris connectilis | c | 11+ | 36+ | 563 | 331 | 15+ | 331 | 828 | ||
| Geranium sylvaticum s.l. | c | 4+ | 13+ | 11+ | 5+ | 50+ | 451 | |||
| Carex disperma | c | 7+ | 11+ | 13+ | 675 | |||||
| Sphagnum angustifolium | d | 192 | 6+ | 7811 | 181 | |||||
| S. riparium | d | 4+ | 111 | 115 | 182 | |||||
| Ptilium crista-castrensis | d | 221 | 11+ | 18+ | 25+ | 442 | 25+ | 502 | ||
| Sphagnum russowii | d | 223 | 22+ | 183 | 338 | 255 | 172 | |||
| Calamagrostis purpurea s.l. | c | 26+ | 9+ | 311 | 442 | 35+ | 17+ | 821 | ||
| Viola epipsila | c | 20+ | 271 | 13+ | 22+ | 30+ | 17+ | 733 | ||
| Listera cordata | c | 7+ | 9+ | 6+ | 44+ | 30+ | 17+ | 18+ | ||
| Rhizomnium pseudopunctatum | d | 19+ | 441 | 302 | 55+ | |||||
| Sphagnum flexuosum | d | 221 | 102 | 185 | ||||||
| Veratrum lobelianum | c | 13+ | 11+ | 10+ | 331 | 64+ | ||||
| Melica nutans | c | 7+ | 11+ | 13+ | 5+ | 361 | ||||
| Paris quadrifolia | c | 7+ | 22+ | 5+ | 17+ | 36+ | ||||
| Calamagrostis obtusata | c | 19+ | 5+ | 363 | ||||||
| Bistorta major | c | 6+ | 36+ | |||||||
| Viola biflora | c | 11+ | 27+ | |||||||
| Rhytidiadelphus subpinnatus | d | 11+ | 10+ | 361 | ||||||
| Plagiothecium denticulatum | d | 18+ | 6+ | 36+ | ||||||
| Pellia neesiana | d | 36+ | ||||||||
| Детерминанты ассоциаций в табл. 1/Determinants of associations in Table 1 | ||||||||||
| Maianthemum bifolium | c | 1003 | 1002 | 1006 | 914 | 1005 | 781 | 1006 | 833 | 1004 |
| Oxalis acetosella | c | 892 | 1006 | 10010 | 10014 | 1009 | 891 | 801 | 834 | 916 |
| Dryopteris carthusiana | c | 782 | 1003 | 1002 | 551 | 636 | 333 | 501 | 672 | 27+ |
| Gymnocarpium dryopteris | c | 371 | 60+ | 441 | 27+ | 816 | 674 | 601 | 10018 | 10012 |
| Rosa acicularis | b | 48+ | 20+ | 18+ | 38+ | 56+ | 451 | 672 | 821 | |
| Rubus humulifolius | c | 1003 | 1005 | 673 | 1002 | |||||
| R. arcticus | c | 26+ | 19+ | 441 | 25+ | 50+ | 55+ | |||
| R. chamaemorus | c | 783 | 352 | 678 | 18+ | |||||
| Детерминанты всех ассоциаций мезотрофных ельников хвощовых сфагновых/ Determinants of all the associations of mesotrophic horsetail-peatmoss spruce forests |
||||||||||
| Equisetum sylvaticum | c | 10025 | 1007 | 10018 | 917 | 10014 | 10027 | 10019 | 8315 | 10035 |
| Sphagnum girgensohnii | d | 8546 | 10052 | 10047 | 10052 | 10050 | 6734 | 7537 | 10040 | 8228 |
| Константные виды/Constants | ||||||||||
| Picea abies s.l. | a1 | 10047 | 10045 | 10049 | 10050 | 9449 | 10037 | 10049 | 10038 | 10039 |
| Betula pubescens s.l. | a1 | 675 | 402 | 675 | 643 | 815 | 10011 | 805 | 8312 | 915 |
| Picea abies s.l. | a2 | 637 | 8013 | 789 | 558 | 6311 | 5610 | 656 | 508 | 736 |
| Sorbus aucuparia s.l. | a2 + b | 893 | 10010 | 10010 | 914 | 1008 | 674 | 853 | 833 | 1007 |
| Picea abies s.l. | b | 1008 | 10013 | 10012 | 10026 | 10013 | 10011 | 1007 | 10018 | 10011 |
| Vaccinium myrtillus | c | 1008 | 10011 | 893 | 1008 | 944 | 1005 | 10012 | 10012 | 914 |
| Trientalis europaea | c | 1002 | 1001 | 1003 | 1002 | 1003 | 891 | 952 | 832 | 1003 |
| Vaccinium vitis-idaea | c | 967 | 1005 | 1005 | 824 | 442 | 1004 | 705 | 674 | 361 |
| Linnaea borealis | c | 742 | 40+ | 331 | 181 | 944 | 1002 | 801 | 837 | 913 |
| Carex globularis | c | 784 | 40+ | 67+ | 45+ | 502 | 785 | 755 | 677 | 552 |
| Pleurozium schreberi | d | 895 | 1006 | 785 | 1005 | 817 | 783 | 906 | 10016 | 915 |
| Hylocomium splendens | d | 855 | 10020 | 898 | 643 | 505 | 782 | 9510 | 10019 | 734 |
| Polytrichum commune | d | 8110 | 601 | 331 | 827 | 8116 | 786 | 504 | 675 | 451 |
| Прочие виды/Companions | ||||||||||
| Populus tremula | a1 + 2 | 302 | 40+ | 111 | 92 | 13+ | 22+ | 10+ | 93 | |
| Larix sibirica | a1 + 2 | 5+ | 335 | |||||||
| Betula pubescens s.l. | a2 | 221 | 403 | 44+ | 271 | 381 | 251 | 336 | 18+ | |
| B. pendula | a + b | 4+ | 601 | 27+ | 63 | 111 | 5+ | 176 | ||
| Alnus glutinosa | a + b | 221 | 272 | |||||||
| A. incana | a2 + b | 26+ | 40+ | 11+ | 9+ | 443 | 101 | 18+ | ||
| Betula pubescens s.l. | b | 561 | 601 | 561 | 551 | 691 | 33+ | 15+ | 671 | 641 |
| Juniperus communis | b | 19+ | 6+ | 22+ | 5+ | 17+ | ||||
| Amelanchier ovalis | b | 4+ | 201 | 331 | 271 | |||||
| Salix aurita | b | 14+ | 22+ | 9+ | ||||||
| Lonicera nigra | b | 361 | ||||||||
| Orthilia secunda | c | 48+ | 601 | 671 | 27+ | 6+ | 331 | 551 | 672 | 27+ |
| Lycopodium annotinum | c | 331 | 20+ | 22+ | 271 | 381 | 33+ | 401 | 673 | 911 |
| Goodyera repens | c | 26+ | 19+ | 22+ | 35+ | 17+ | ||||
| Avenella flexuosa | c | 22+ | 18+ | 13+ | 22+ | 151 | 17+ | 27+ | ||
| Melampyrum sylvaticum | c | 26+ | 31+ | 33+ | 10+ | 17+ | ||||
| M. pratense | c | 15+ | 40+ | 36+ | 19+ | 11+ | 5+ | 17+ | 9+ | |
| Chamaenerion angustifolium | c | 15+ | 11+ | 33+ | 5+ | 331 | 45+ | |||
| Angelica sylvestris | c | 4+ | 20+ | 33+ | 6+ | 11+ | 10+ | 33+ | 18+ | |
| Cirsium heterophyllum | c | 4+ | 20+ | 22+ | 6+ | 10+ | 271 | |||
| Equisetum palustre | c | 4+ | 11+ | 221 | 20+ | 9+ | ||||
| Eriophorum vaginatum | c | 22+ | 5+ | 171 | ||||||
| Hieracium agg. vulgatum | c | 20+ | 22+ | 5+ | ||||||
| Aulacomnium palustre | d | 301 | 40+ | 44+ | 18+ | 31+ | 33+ | 452 | ||
| Sphagnum centrale | d | 308 | 20+ | 444 | 271 | 333 | 402 | 171 | 18+ | |
| S. warnstorfii | d | 114 | 93 | 223 | 151 | 17+ | 9+ | |||
| Dicranum majus | d | 15+ | 11+ | 19+ | 11+ | 5+ | 91 | |||
| Calliergon cordifolium | d | 11+ | 22+ | 11+ | 10+ | 18+ | ||||
| Rhizomnium punctatum | d | 7+ | 331 | 9+ | 6+ | 222 | ||||
| Sciuro-hypnum reflexum | d | 20+ | 11+ | 9+ | 271 | |||||
| Dicranum scoparium | z | 26+ | 20+ | 22+ | 9+ | 19+ | 11+ | 45+ | 27+ | |
| D. fuscescens | z | 26+ | 40+ | 11+ | 9+ | 31+ | 22+ | 35+ | 45+ | |
| Sciuro-hypnum starkei | z | 15+ | 11+ | 9+ | 13+ | 11+ | 5+ | 36+ | ||
| Средние сомкнутость или покрытие ярусов, %/Average density or cover of layers, % | ||||||||||
| – 1-го яруса древостоя/1st tree layer | 0.6 | 0.6 | 0.6 | 0.7 | 0.7 | 0.5 | 0.6 | 0.7 | 0.6 | |
| – 2-го яруса древостоя/2nd tree layer | 0.2 | 0.2 | 0.1 | 0.2 | 0.2 | 0.1 | 0.1 | 0.3 | 0.1 | |
| – подроста и подлеска/regrowth and shrubs | 15 | 25 | 30 | 35 | 25 | 20 | 10 | 40 | 30 | |
| – кустарничково-травяного/field layer | 60 | 55 | 65 | 55 | 65 | 75 | 65 | 80 | 80 | |
| – мохового/ground layer | 90 | 90 | 90 | 85 | 80 | 95 | 90 | 95 | 75 | |
| Средняя высота ярусов, м/Average height of layers, m | ||||||||||
| – 1-го яруса древостоя/1st tree layer | 22 | 25 | 23 | 24 | 22 | 17 | 20 | 17 | 25 | |
| – 2-го яруса древостоя/2nd tree layer | 12 | 17 | 17 | 18 | 14 | 12 | 13 | 8 | 15 | |
| – 1-го подъяруса подроста и подлеска/1st sub-layer of regrowth and shrubs | 4.2 | 5.7 | 4.8 | 4.8 | 4.5 | 3.1 | 5.5 | 2.1 | 6.0 | |
| – 2-го подъяруса/2nd sub-layer | 1.3 | 1.7 | 1.8 | 1.7 | 1.4 | – | 1.5 | 1.0 | 1.7 | |
| – 3-го подъяруса/3rd sub-layer | 0.8 | 0.5 | 0.7 | 0.6 | – | – | 1.0 | – | – | |
| Бонитет древостоя/Stand quality class | III | II | III | II | III | IV | III | IV | II | |
| Мощность торфа/Peat thickness, cm | 30 | 15 | 25 | 20 | 15 | 45 | 30 | 35 | 20 | |
| Число описаний/Number of relevés | 27 | 5 | 9 | 11 | 16 | 9 | 20 | 6 | 11 | |
Примечания. Синтаксоны: 1–5 – Piceetum (P.) sphagno girgensohnii-equisetosum sylvaticae: 1 – typicum; 2–5 – dryopteridosum expansae: 2 – var. Convallaria majalis, 3 –var. Circaea alpina, 4 – var. typica, 5 – var. Abies sibirica; 6–9 – P. sphagno girgensohnii-humulifolii-ruboso-equisetosum sylvaticae: 6 – caricosum dispermae, 7 – typicum, 8–9 – abietosum sibiricae: 8 – var. typica, 9 – var. Dryopteris expansa.
Ярусы: a1 и a2 – 1-й и 2-й ярусы древостоя, b – подрост и подлесок, c – кустарничково-травяной, d – моховой, z – эпифитные и эпиксильные мхи и лишайники. Для видов приводятся постоянство (%) и (в надстрочном регистре) среднее проективное покрытие (%). Детерминантные группы выделены серым фоном и жирной рамкой; виды в их пределах сортированы по ярусам, далее по убыванию встречаемости во всем массиве описаний таблицы. Для доминирующих видов значения покрытия и постоянства даны полужирным шрифтом. Среднее проективное покрытие видов менее 0.5% отмечено плюсом “+”. Прочерк “–” – отсутствие подъяруса. Исключены сопутствующие виды со встречаемостью менее 20% во всех синтаксонах, всходы деревьев и кустарников.
Notes. Syntaxa: 1–5 – Piceetum (P.) sphagno girgensohnii-equisetosum sylvaticae: 1 – typicum; 2–5 – dryopteridosum expansae: 2 – var. Convallaria majalis, 3 –var. Circaea alpina, 4 – var. typica, 5 – var. Abies sibirica; 6–9 – P. sphagno girgensohnii-humulifolii-ruboso-equisetosum sylvaticae: 6 – caricosum dispermae, 7 – typicum, 8–9 – abietosum sibiricae: 8 – var. typica, 9 – var. Dryopteris expansa.
Layers: a1 and a2 – dominant and subordinate tree layers, b – regrowth and shrubs, c – field layer, d – ground layer, z – epiphytic and epixylic mosses and lichens. Constancy (%) and average cover (%) (in superscripts) are given for species. Determinant groups are marked by grey backgrounds and bold margins; species within them are sorted first by layer, then by descending occurrence in the whole table relevé set. The values of cover and constancy of dominants are shown in bold. Average species cover under 0.5% is given as “+”. Dash “–” corresponds to sub-layer absence. Species with occurrence under 20% in all of the syntaxa are excluded from the table together with tree and shrub seedlings.
Номенклатура сосудистых растений дана по С.К. Черепанову (Cherepanov, 1995), мхов – по М.С. Игнатову с соавторами (Ignatov et al., 2006), лишайников – по “Списку лихенофлоры России” (A checklist…, 2010). Европейские таежные популяции ели отнесены к единому комплексу Picea abies s.l. (Popov, 2005), березы пушистой – к комплексу Betula pubescens s.l. (incl. B. subarctica, B. aurata, etc. [Tzvelev, 2004]). Границы зон и подзон растительности приняты по Т.И. Исаченко и Е.М. Лавренко (Isachenko, Lavrenko, 1980) с уточнениями (Safronova, Yurkovskaya, 2015). Зональная обусловленность ареалов синтаксонов оценена с помощью сумм превышений среднесуточных температур над базовой температурой в 10°C (“сумм градусо-дней выше 10°C”) по данным сети спутниковой метеосъемки (NASA…, 2006). На основе этой же базы вычислены индексы континентальности Конрада (Tukhanen, 1980; Kucherov, Kutenkov, 2020) (табл. 2).
Таблица 2.
Амплитуды среднемноголетних (1983–2004) значений метеопараметров в пределах ареалов синтаксонов мезотрофных ельников хвощовых, щитовниково- и хмеленично-хвощовых сфагновых Table 2. Amplitudes of averaged (1983–2004) climatic parameter values within the ranges of mesotrophic horsetail, shield-fern-horsetail, and hop-leaved raspberry-horsetail peatmoss spruce forest syntaxa
| Синтаксоны/Syntaxa | Метеопараметры/Climatic parameters | |
|---|---|---|
| GDD > 10, °C | K | |
| 1. Piceetum (P.) sphagno girgensohnii-equisetosum subass. typicum | (471) 484–817 | 32.3–42.4 |
| 2. Subass. dryopteridosum expansae var. Convallaria majalis | 686 | 32.3 |
| 3. Var. Circaea alpina | 674–758 | 29.3–32.3 |
| 4. Var. typica | 619–769 (1075) | 31.2–40.1 (42.7) |
| 5. Var. Abies sibirica | 388–761 | 40.3–45.6 |
| 6. P. sphagno girgensohnii-humulifolii-ruboso-equisetosum subass. caricosum dispermae | 410–635 | 38.3–44.1 |
| 7. Subass. typicum | 451–738 | 36.9–43.5 |
| 8. Subass. abietosum sibiricae var. typica | 388–816 | 43.5–46.3 |
| 9. Var. Dryopteris expansa | 388–795 | 37.3–48.1 |
Примечания. GDD > 10 – сумма градусо-дней выше 10°С (NASA, 2006), K – коэффициент континентальности Конрада (Tukhanen, 1980). Амплитуды рассчитаны с учетом как имеющейся совокупности описаний, так и данных литературы. В скобках даны значения параметров для экстразональных местонахождений сообществ.
Notes. GDD > 10 – sum of growing degree-days above 10°С (NASA, 2006); K – Conrad continentality index (Tukhanen, 1980). Amplitude calculations are based upon both the examined relevé set and the published sources. Parameter values for extrazonal community localities are given in brackets.
Общая характеристика “южных” сообществ мезотрофных ельников хвощовых сфагновых
Если в средней тайге ельники хвощовые сфагновые чаще всего встречаются по долинам ручьев и малых рек, то в южной характерным местом их произрастания становятся пониженные участки водоразделов с тяжелыми почвами, проточные в период снеготаяния, но лишь слабопроточные летом. Также подобные ельники произрастают по окраинам мезотрофных болот, а на Урале – на шлейфах горных склонов.
В южной тайге ельники хвощовые сфагновые, казалось бы, занимают еще меньшие площади, чем в северной и средней. На леса майниково-сфагновой серии типов, к которой относит их В.Н. Федорчук, приходится лишь 0.2–0.5% от общей площади ельников Ленинградской обл. (Fedorchuk et al., 2005). Однако в ландшафтах водоразделов и котловин с покровными суглинками данные ельники выступают одной из самых обычных лесных ассоциаций, как, например, в заказнике “Лисинский” в центральной части Ленинградской обл. (наблюдения И.Б. Кучерова 2017 г.).
Нанорельеф обычно двухкомпонентный из кочек (вкупе с приствольными повышениями) и сфагновых ковров; последние часто преобладают. При ветровалах образуются западины под вывалами. Почвы торфянисто- и торфяно-подзолисто-глеевые (Shaposhnikov, 1988) или торфяно-глеевые (Nitsenko, 1960; Gavrilov, Karpov, 1962; Dmitrieva, 1973), чаще суглинистые или супесчаные, в Двино-Печорском регионе также песчаные. Мощность торфа на покровных суглинках в депрессиях водоразделов обычно не более 10–30, лишь по окраинам болот изредка возрастает до 60–120 см.
Кислотность (${\text{p}}{{{\text{H}}}_{{{{{\text{H}}}_{{\text{2}}}}{\text{O}}}}}$) торфа в ельниках хвощовых сфагновых Ярославской обл. составляет 4.8 (Orlov et al., 1974), что сопоставимо с мезотрофными ельниками черничными сфагновыми (Kucherov, Kutenkov, 2019 b). Плодородие почв в лесах первого синтаксона при этом выше из-за утяжеления гранулометрического состава. Это позволяет мезотрофным видам расти на всех элементах нанорельефа (Kucherov, Kutenkov, 2020). Считается даже, что в ельниках хвощовых сфагновых плодородие почв сопоставимо с таковым в кисличных и выше, чем в черничных зеленомошных. Накопление подвижных форм N и P происходит из-за их поступления с поверхностным стоком с вышележащих склонов, а также в силу анаэробных условий в почвенной толще, способствующих аммонификации. Имеет место, однако, и угнетение поглощения элементов питания из-за снижения мощности корнеобитаемого слоя (Orlov et al., 1974).
Бонитет 1-го яруса древостоя в южной тайге и подтайге может достигать III и даже II при сомкнутости 0.6–0.7. К Picea abies s.l. обычно примешивается Betula pubescens s.l., в западном и центральном секторах Европейской России также Pinus sylvestris. Разреженный 2-й ярус древостоя и подрост сформированы елью. В средней тайге подрост групповой, в южной смыкающийся, с ПП 20% и более. Изредка может развиваться высокий подлесок из Sorbus aucuparia; в этом случае подрост ели не столь обилен. В нижнем подъярусе подлеска обычна Frangula alnus. На востоке Русской равнины и на Урале, как в средней, так и в южной тайге ельники замещаются пихто-ельниками с примесью Abies sibirica к Picea obovata и преимущественно пихтовым подростом, т. е. на дифференциацию синтаксонов влияют как широтная зональность, так и изменения континентальности климата (Kucherov, Kutenkov, 2020).
Вместе с Equisetum sylvaticum в кустарничково-травяном ярусе часто доминируют крупные папоротники (Dryopteris expansa s.l. и/или Athyrium filix-femina в зависимости от синтаксона). В Двино-Печорском регионе и на Урале к субдоминантам яруса относится также Rubus humulifolius, индикатор особой субконтинентальной ассоциации. При стволах и на кочках обычны и обильны Vaccinium myrtillus и V. vitis-idaea, на коврах также виды таежного мелкотравья, в первую очередь Maianthemum bifolium и Oxalis acetosella. При стволах к ним добавляются лесные мезофиты, нехарактерные для ельников морошково-хвощовых, – Calamagrostis arundinacea и Rubus saxatilis. В моховом ковре выражено господство Sphagnum girgensohnii, нередко с малочисленной примесью S. squarrosum. На микроповышениях преобладают таежные зеленые мхи: Hylocomium splendens, Pleurozium schreberi, Dicranum polysetum. К ним примешиваются Sphagnum russowii, S. wulfianum и Polytrichum commune.
Константами всех синтаксонов, как и в ельниках морошково-хвощовых (Kucherov, Kutenkov, 2020), выступают Picea abies s.l. и Betula pubescens s.l. в древостое, Picea abies в составе подроста, Sorbus aucuparia, Vaccinium myrtillus, V. vitis-idaea, Trientalis europaea, Linnaea borealis, Carex globularis, Pleurozium schreberi, Hylocomium splendens, Polytrichum commune. Доминирующие в приземных ярусах Equisetum sylvaticum и Sphagnum girgensohnii одновременно входят в число детерминантов всех выделенных ассоциаций (табл. 1).
Характеристика выделенных синтаксонов
Piceetum (P.). sphagno girgensohnii-equisetosum (Sphagno girgensohnii-Equiseto-P.) – собственно ельник (е.) хвощовый сфагновый (табл. 1, рис. 1: № 1–5). Леса, относящиеся к этой ассоциации, распространены от южных границ северной тайги до подтайги, чаще всего встречаясь в средней и южной тайге. На севере средней тайги ареал ассоциации частично перекрывается с таковым ельников морошково-хвощовых сфагновых (Kucherov, Kutenkov, 2020). Леса произрастают преимущественно на тяжелых почвах. Picea abies s.l. в большинстве подчиненных синтаксонов представлена смешанными популяциями P. × fennica и P. abies s.str. с увеличением доли последнего вида в южной тайге, особенно к юго-западу. Сравнительно с большинством субассоциаций ельников морошково-хвощовых возрастает бонитет древостоя (до II–III против IV). Подрост и подлесок стратифицированы по высоте на 2–3 подъяруса.
Рис. 1.
Распространение мезотрофных ельников хвощовых (1), лесновейниково- (2) и кочедыжниково-щитовниково-хвощовых (3), щитовниково-хвощовых (4) и пихто-ельников щитовниково-хвощовых (5) сфагновых в Европейской России и на Урале.
Зоны и подзоны: I – тундра, II – лесотундра и подгольцовые редколесья, III – северная, IV – средняя, V – южная тайга, VI – подтайга, VII – широколиственные леса, VIII – лесостепь. Картооснова (Isachenko, Lavrenko, 1980; Safronova, Yurkovskaya, 2015) объединяет зональные выделы и их высотно-поясные аналоги.
Fig. 1. Distribution of Piceetum (P.) sphagno girgensohnii-equisetosum subass. typicum (1), subass. dryopteridosum expansae var. Convallaria majalis (2), var. Circaea alpina (3), var. typica (4), and var. Abies sibirica (5) in European Russia and the Urals.
Zones and subzones: I – tundra, II – forest-tundra and subalpine open woodlands, III – northern-boreal, IV – middle-boreal, V – southern-boreal, VI – hemiboreal forests, VII – broadleaved forests, VIII – forest-steppe. The base map (Isachenko, Lavrenko, 1980; Safronova, Yurkovskaya, 2015) is generalized by the association of zonal / subzonal units together with their altitudinal analogs.
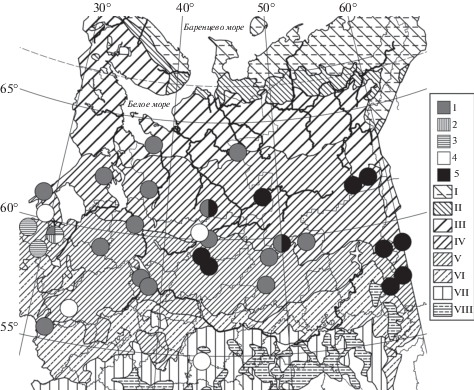
Собственные детерминанты ассоциации – бореальные лесные мезофиты Luzula pilosa и Rubus saxatilis. Для большинства подчиненных синтаксонов также характерен диагностический блок лесных мезофитов – как олиготрофных (эпифит Ptilidium pulcherrimum) и олигомезотрофных бореальных (Pinus sylvestris в древостое, Dicranum polysetum) и бореонеморальных (Calamagrostis arundinacea), так и мезотрофных бореальных (Solidago virgaurea, эпифит Sciuro-hypnum reflexum). В состав блока входят и мезотрофные полизональные эпифиты (Plagiothecium laetum, Cladonia coniocraea). Дифференциальная группа мезотрофных лесных мезофитов (Maianthemum bifolium, Oxalis acetosella, Dryopteris carthusiana, Gymnocarpium dryopteris) объединяет ассоциацию с ельниками хмеленично-хвощовыми и рядом субассоциаций и вариантов ельников морошково-хвощовых. Но выражена и негативная дифференциация от названных синтаксонов по отсутствию Rubus chamaemorus, R. arcticus и R. humulifolius. Кроме того, детерминанты ельников морошково-хвощовых Orthilia secunda и Lycopodium annotinum в собственно хвощовых перестают быть константными (табл. 1).
По составу видов кустарничково-травяного яруса выделены две субассоциации.
Subass. typicum (табл. 1, рис. 1: № 1). В средней тайге ельники типичной субассоциации чаще встречаются в долинах рек, заболоченных межозовых и межсельговых понижениях, в южной – на пониженных участках водоразделов с тяжелыми почвами. Леса произрастают как на силикатных, так и на карбонатных породах, почти исключительно на супесях и суглинках, на песках очень редко. Мощность торфа 15–40, в средней тайге иногда до 65–120 см. Нанорельеф ровный или кочковатый с кочками высотой 20–50 см, на ветровалах с западинами. В 1-м ярусе древостоя примесь Betula pubescens s.l. к Picea abies s.l. незначительна, Pinus sylvestris – единична. В еловом 2-м ярусе столь же малая примесь Betula pubescens s.l. и Sorbus aucuparia. Ярус подроста и подлеска рябиново-еловый. Его верхний подъярус состоит из наиболее успешных экземпляров Picea abies и Sorbus aucuparia, промежуточный – с участием Betula pubescens s.l., нижний – с редкой примесью Rosa acicularis. Ярко выражено господство Equisetum sylvaticum в кустарничково-травяном ярусе. Vaccinium myrtillus и V. vitis-idaea – доминанты 2-го порядка при стволах и на коврах. Maianthemum bifolium, Oxalis acetosella, Trientalis europaea, Linnaea borealis, а также Carex globularis и Dryopteris carthusiana – их постоянные спутники. В моховом ярусе к Sphagnum girgensohnii активно примешиваются S. centrale и Polytrichum commune; Pleurozium schreberi и Hylocomium splendens – доминанты 2-го порядка при стволах. Дифференциация негативная, по отсутствию видов из групп Populus tremula–Convallaria majalis, Salix caprea–Deschampsia cespitosa, Frangula alnus–Plagiochila asplenioides, Rubus idaeus–Dryopteris expansa, Athyrium filix-femina–Crepis paludosa и др. (табл. 1).
Ареал субассоциации, как и ассоциации в целом, тянется от северной тайги до подтайги. В северной тайге Архангельской обл. близ границы со средней ельники данного синтаксона описаны в левобережных низовьях р. Онеги (экстразональное сообщество южного склона с обильным Calamagrostis arundinacea) и на Пинежско-Мезенском водоразделе по р. Нюхче. В средней тайге сообщества отмечены на северо-западе Карельского перешейка (P. equisetoso-sphagnosum (Dmitrieva, 1973)), в Южной Карелии, на юго-западе и юге Архангельской обл. и в Кайском р-не на севере Кировской обл. (P. equiseto-polytrichoso-sphagnosum (Smirnova A., 1954)).
В южной тайге описания ельников субассоциации выполнены в Ленинградской и Вологодской областях, в последней– на карбонатной морене левобережья р. Кемы и на пермских глинах по р. Илезе (P. equisetosum permense (Korchagin, 1929)). Сообщества отмечены также в Ярославской обл. (Orlov et al., 1974) и на севере Кировской обл. (P. equisetoso-sphagnosum, P. equiseto-polytrichoso-sphagnosum (Smirnova A., 1954)). По мнению В.И. Василевича, они произрастают по всей южной тайге этой области (“ельник сфагновый” (Vasilevich, 2003)). Восточнее они не найдены.
В подтайге рассматриваемые ельники были отмечены на севере Смоленской обл. (“ельник осоково-сфагновый” IV бонитета с Frangula alnus, Lonicera xylosteum, Equisetum sylvaticum, Carex globularis и C. nigra (Grozdov, 1951)). Видимо, они росли также на Молого-Шекснинском междуречье (P. equisetoso-myrtilloso-polytrichosum c Oxalis acetosella и Calamagrostis arundinacea (Korchagin, Senyaninova-Korchagina, 1957)), где нередко встречались флористически и физиономически близкие ельники хвощово-черничные (P. sphagno girgensohnii-myrtillosum), переходные к черничным сфагновым (Kucherov, Kutenkov, 2019b). Здесь уместно сказать о трудностях при разграничении этих двух синтаксонов, хотя в типичных ельниках хвощовых сфагновых в среднем и выше ПП Equisetum sylvaticum (25 против 10% в хвощово-черничных) и меньше Vaccinium myrtillus (8 против 25%). Также первый синтаксон дифференцируют от второго Calamagrostis arundinacea, Solidago virgaurea, Rubus saxatilis и Gymnocarpium dryopteris, а второй от первого – Salix aurita и Melampyrum pratense (Kucherov, Kutenkov, 2019 b). Однако постоянство всех этих видов в основном не превышает III класса. В то же время у синтаксонов наблюдается немало общих черт, вплоть до сходства примесей в древостое, одинаковых не только по составу, но и по пропорции. Возможно, будущие исследования покажут, что эти синтаксоны все же не стоит разделять.
Subass. dryopteridosum expansae – е. щитовниково-хвощовый сфагновый (табл. 1, рис. 1: № 2–5). Преимущественно южнотаежная субассоциация на западе и в центре Европейской России, распространенная шире – от средней тайги до подтайги – в Предуралье и на Урале. Представлена четырьмя вариантами разной топоэдафической приуроченности, одни из которых вдобавок замещают другие с запада на восток.
Примесь Betula pubescens s.l. к Picea abies s.l. различна в разных вариантах, Pinus sylvestris – незначительна в большинстве из них. Сравнительно с subass. typicum, возрастают ПП Sorbus aucuparia, подроста Picea abies, общее ПП яруса подроста и подлеска. В лесах “западных” вариантов в заказнике “Лисинский” во все подъярусы последнего активно внедряется инвазионный орнитохорный вид Amelanchier ovalis. В нижний подъярус входит один из детерминантов синтаксона, Rubus idaeus.
В кустарничково-травяном ярусе Equisetum sylvaticum делит господство с другим детерминантом субассоциации, Dryopteris expansa s.l. (D. assimilis, на Урале и в Предуралье также D. expansa s.str.), в ряде вариантов также с другими видами крупных папоротников. Постоянны, но не обильны Trientalis europaea, Linnaea borealis, Luzula pilosa, Rubus saxatilis, Solidago virgaurea, Carex globularis. В моховом ярусе Sphagnum girgensohnii сопровождает большое число доминантов 2-го порядка и видов-спутников, набор которых тоже зависит от варианта.
Var. Convallaria majalis – е. лесновейниково-хвощовый сфагновый (табл. 1, рис. 1: № 2). Леса произрастают на силикатных супесях и суглинках в понижениях и на пологих склонах водоразделов. Мощность оторфованной подстилки 15–20 см; переувлажнение местообитаний в значительной степени сезонное. Поверхность ровная или кочковатая с преобладанием ковров; кочки не выше 20, редко 30–50 см. Древостой несколько разрежен. В его 2-м ярусе и в верхнем подъярусе подроста и подлеска согосподствуют Picea abies и Sorbus aucuparia с примесью Betula pubescens s.l. и Salix caprea. В нижележащих подъярусах к ели и рябине добавляются Frangula alnus, Populus tremula, Amelanchier ovalis, Rubus idaeus, единично Quercus robur. Ведущую роль в кустарничково-травяном ярусе играет синузия микроповышений с обильными Calamagrostis arundinacea, Oxalis acetosella и Vaccinium myrtillus. Доминант 2-го порядка – V. vitis-idaea, вместе с последними двумя видами частично спускающаяся c кочек на сфагновый ковер. В числе сопутствующих видов синузии – Maianthemum bifolium и Convallaria majalis. На участках ковров Equisetum sylvaticum лишь умеренно обилен. Ему постоянно сопутствуют Dryopteris expansa s.l., D. carthusiana, Athyrium filix-femina, а также Calamagrostis canescens и Deschampsia cespitosa. В моховом ярусе при стволах доминирует Hylocomium splendens с примесью Pleurozium schreberi и Rhytidiadelphus triquetrus. При основании микроповышений пятнами растет Plagiochila asplenioides. В сфагновом ковре к Sphagnum girgensohnii единично примешивается S. squarrosum. S. centrale нехарактерен. Polytrichum commune – сопутствующий вид.
Ельники этого варианта – наиболее неморализованные и флористически богатые среди всех рассматриваемых синтаксонов. Их собственные детерминанты – неморальные (Anemonoides nemorosa, Galeobdolon luteum) и бореонеморальные (Convallaria majalis, подрост Populus tremula) мезотрофные лесные мезофиты; к ним примыкает мезотрофный гигромезофит Plagiomnium affine. Бореонеморальные мезотрофные (Stellaria holostea) и мезоэвтрофные (Milium effusum) мезофиты сближают вариант также с уральскими пихто-ельниками щитовниково-хмеленично-хвощовыми.
С var. Circaea alpina вариант объединяет группа гигромезофитов и гигрофитов, растущих на участках с истонченной подстилкой и близким залеганием суглинка, от олигомезотрофов (Agrostis canina) и мезотрофов (Carex canescens, Sphagnum squarrosum) до мезоэвтрофов (Calamagrostis canescens, Deschampsia cespitosa, Galium palustre). К этой группе, однако, примыкают и мезофиты, в основном мезотрофные (Salix caprea, подрост Quercus robur, Carex brunnescens на вывалах, Rhyti-diadelphus triquetrus, эпифит Peltigera praetextata), отчасти также мезоэвтрофные (Rhodobryum roseum). Еще одна группа мезотрофных (Athyrium filix-femina, Plagiomnium ellipticum) и мезоэвтрофных (Crepis paludosa) гигромезофитов и мезогигрофитов сближает вариант как с var. Circaea alpina, так и с упомянутыми выше пихто-ельниками Урала.
Группу, объединяющую вариант не только с var. Circaea alpina, но и с var. typica, формируют бореонеморальные мезотрофные (Frangula alnus) и неморальные мезоэвтрофные (Plagiochila asplenioides) гигромезо- и мезогигрофиты вместе с мезофильными мхами, растущими на валеже (Tetraphis pellucida, Dicranum polysetum, Hylocomium splendens, Sanionia uncinata). Pohlia nutans и Pleurozium schreberi в составе эпиксильной синузии также объединяют вышеперечисленные три варианта с щитовниково-хмеленично-хвощовыми пихто-ельниками.
Леса рассматриваемого варианта пока известны авторам лишь на территории заказника “Лисинский” в южной тайге Ленинградской обл. Но они должны встречаться и в других южнотаежных регионах Северо-Запада России, а также в Эстонии.
Var. Circaea alpina – е. кочедыжниково-хвощовый сфагновый (табл. 1, рис. 1: № 3). Как и леса предыдущего варианта, эти сообщества приурочены к тяжелым почвам в понижениях силикатных водоразделов. Лишь иногда они встречаются также на надпойменных террасах или при основании склонов на контакте с краевой частью пойменных болот. Заболочены сильнее, чем ельники var. Convallaria majalis; развита неглубокая (10–40 см) торфяная залежь. Нанорельеф кочковатый; на кочки высотой 20–50 см приходится 30–70% площади ценоза; обычны вывалы. Бонитет 1‑го яруса древостоя ниже, чем у предыдущего варианта; 2-й ярус разрежен, иногда с примесью Betula pubescens s.l. и Alnus glutinosa, но в остальном как в var. Convallaria majalis. То же касается и верхнего подъяруса подроста и подлеска. В промежуточном подъярусе возрастает ПП Frangula alnus, а Populus tremula, напротив, “выпадает”. Rubus idaeus растет в основном на вывалах. В кустарничково-травяном ярусе выраженно возрастает ПП как Equisetum sylvaticum, так и доминантов 2‑го порядка – Dryopteris expansa s.l. и Athyrium filix-femina. В описании с р. Облы (Sambuk, 1930) обилен также Mercurialis perennis. На микроповышениях согосподствуют Oxalis acetosella, Maianthemum bifolium и Vaccinium vitis-idaea. В число видов-спутников переходят V. myrtillus и Calamagrostis arundinacea. В моховом ярусе микроповышений доминируют Pleurozium schreberi, Hylocomium splendens и Rhytidiadelphus triquetrus. При основании кочек и стволов по-прежнему много Plagiochila asplenioides. В ковре обильна примесь Sphagnum squarrosum к S. girgensohnii; на кочках растут S. wulfianum, S. centrale и Polytrichum commune.
Группа детерминантов варианта объединяет мезотрофных мезофитов (Circaea alpina) и олигомезотрофных гигромезофитов (Carex vaginata) микроповышений с мезоэвтрофными и эвтрофными мезофитами (Mercurialis perennis) и гигромезофитами (Ranunculus repens, Lysimachia vulga-ris), растущими на коврах при близком залегании водоупора. О группах, объединяющих синтаксон с var. Convallaria majalis, сказано выше. В свою очередь, мезотрофный гигромезофит Sphagnum wulfianum сближает его c ельниками хмеленично-хвощовыми осочковой и типичной субассоциаций.
Леса этого варианта пока тоже известны лишь из южной тайги Ленинградской обл.: из заказника “Лисинский”, а также на склонах Крикковского плато в бассейне р. Луги (P. oxalidosum humidum (Smirnova Z., 1928)) и в пойме ее притока р. Облы (P. oxalidoso-herbosum (Sambuk, 1930)). Но, как и сообщества предыдущего синтаксона, они могут быть найдены и в других местностях Северо-Запада Европейской России.
Именно к этому, а также к следующему вариантам в первую очередь относится указание о необходимости разграничения мезотрофных ельников хвощовых и мезоэвтрофных таволгово-хвощовых сфагновых (Kucherov, Kutenkov. 2020).
Ельники var. typica (табл. 1, рис. 1: № 4) отмечены в скрытопроточных понижениях на водоразделах, в ложбинах и распадках при основании “длинных” склонов озов и сельг, вдоль северного побережья Финского залива – при основании Литторинового уступа. В зависимости от региона они произрастают как на силикатных, так и на карбонатных породах, равно на тяжелых почвах и на песках. Торфяная залежь маломощная, как и в лесах предыдущего варианта. Кочки достигают высоты 20–80 см, но занимают не более 20–40% площади ценоза. Обычны западины под вывалами. Бонитет древостоя вновь возрастает. Второй ярус также гуще и выше, чем в предыдущем варианте, но тоже бывает с примесью Alnus glutinosa. Общее ПП подроста и подлеска максимально для ассоциации за счет подроста Picea abies; ПП Sorbus aucuparia при этом снижено. Для нижних подъярусов подлеска по-прежнему характерна Frangula alnus. Вдоль берега Финского залива в число инвазионных видов подлеска вместе с Amelanchier ovalis входит Lonicera nigra, одичавшая в финский период истории Карельского перешейка. Сравнительно с двумя предыдущими вариантами, кустарничково-травяной ярус беднее видами. На фоне выраженного господства Dryopteris expansa s.l. и Oxalis acetosella Equisetum sylvaticum переходит на роль доминанта 2-го порядка, так же как Maianthemum bifolium и Vaccinium myrtillus; V. vitis-idaea и Dryopteris carthusiana – сопутствующие виды. Gymnocarpium dryopteris, Linnaea borealis и Carex globularis становятся редки. Athyrium filix-femina, Crepis paludosa, Circaea alpina “выпадают”. В моховом ковре к Sphagnum girgensohnii примешивается Sciuro-hypnum curtum, а другие виды рода Sphagnum нехарактерны. Доминирующие мхи при стволах – Pleurozium schreberi и Polytrichum commune; Hylocomium splendens – сопутствующий вид. Постоянна примесь Dicranum scoparium. Plagiochila asplenioides малообильна, Rhytidiadelphus triquetrus редок. Мезотрофные мезофиты Dicranum scoparium и Sciuro-hypnum curtum объединяют вариант с пихто-ельниками щитовниково-хмеленично-хвощовыми. Собственная дифференциация его негативная, по отсутствию или нетипичности видов из групп Salix caprea–Deschampsia cespitosa, Athyrium filix-femina–Crepis paludosa и Carex vaginata–Circaea alpina (табл. 1).
Леса типичного варианта распространены от средней тайги до подтайги в западном и центральном секторах Европейской России. В средней тайге Вологодской обл. они описаны на лессовидных суглинках по р. Сухоне (P. filicosum var. oxalidosum, P. humidum (Korchagin, 1929)), а на западе Карельского перешейка – на границе средней и южной тайги в заказниках “Линдуловский” и “Комаровский берег”. Собственно в южной тайге Ленинградской обл. ельники варианта обычны в заказнике “Лисинский”. В подтайге Центрально-Лесного заповедника в Тверской обл. также нередок “ельник хвощово-сфагново-черничный” с Equisetum sylvaticum, Dryopteris expansa s.l., D. carthusiana, Rubus saxatilis, Solidago virgaurea, Sphagnum girgensohnii, S. wulfianum, Rhytidiadelphus triquetrus, Plagiochila asplenioides (Shaposhnikov, 1988). Экстразональный ельник папоротниково-хвощовый сфагново-зеленомошный найден у южных границ подтайги в Мордовском заповеднике на выходах холодных ключей при основании коренного берега сатиско-мокшанской поймы. В нем отмечены Equisetum sylvaticum, Dryopteris carthusiana, D. expansa, Athyrium filix-femina, Gymnocarpium dryopteris, Phegopteris connectilis и Listera cordata (Tereshkin, Tereshkina, 2006).
Восточнее синтаксон замещается следующим вариантом.
Var. Abies sibirica – пихто-ельник щитовниково-хвощовый сфагновый (табл. 1, рис. 1: № 5). Леса более характерны для долин рек и ручьев, на Урале – для шлейфов горных склонов. По окраинам болот встречаются очень редко. Произрастают как на силикатных, так и на карбонатных породах, чаще на тяжелых почвах, чем на песках. Торфяная залежь еще более истончается (15–20 см), а преобладание ковров в нанорельефе (70–80%) еще более выражено, чем в лесах var. typica. Второй ярус древостоя менее разрежен, но снижен. Picea abies s.l. в Архангельской и Вологодской областях представлена P. × fennica, в Заволжье также P. obovata, в Предуралье и на Урале только последним видом. Примесь Pinus sylvestris в древостое случайна, Betula pubescens s.l. – незначительна, Abies sibirica – существенна во 2-м и единична в 1-м ярусе. В Предуралье и на Урале в древостой входит и Pinus sibirica. Подрост пихтово-еловый умеренной густоты. В подлеске вновь возрастает ПП Sorbus aucuparia s.l. Участие других видов минимально, не считая Rubus idaeus на вывалах; инвазионные виды не отмечены. В кустарничково-травяном ярусе к Equisetum sylvaticum возвращается абсолютное господство; ПП Dryopteris expansa s.l. резко снижается. Доминантами 2-го порядка служат D. carthusiana, под пологом хвоща и крупных папоротников также Oxalis acetosella, Maianthemum bifolium и вновь (особенно в уральских сообществах) Gymnocar-pium dryopteris. Vaccinium myrtillus при стволах становится видом-спутником, постоянным наряду с V. vitis-idaea, Trientalis europaea, Linnaea borealis и Carex globularis. Практически “выпадают” Calamagrostis arundinacea и Solidago virgaurea. Ковер Sphagnum girgensohnii с выраженным участием Polytrichum commune. При стволах согосподствуют Pleurozium schreberi и Hylocomium splendens.
Детерминантную группу варианта, общую с уральскими пихто-ельниками хмеленично-хвощовыми, формируют лесообразователи сибирской тайги (Abies sibirica, Pinus sibirica) в древостое и в подлеске. С ними сопряжены мезотрофные (Geranium sylvaticum s.l. (incl. G. krylovii)) и мезоэвтрофные (Phegopteris connectilis) травянистые мезофиты. Нетипичны виды из групп, детерминирующих более “западные” варианты: Pinus sylvestris–Calamagrostis arundinacea, Frangula alnus–Plagiochila asplenioides, Dicranum scoparium–Sciuro-hypnum curtum, Carex disperma–Sphagnum angustifolium, Calamagrostis purpurea–Viola epipsila, Ptilium crista-castrensis–Sphagnum russowii (табл. 1).
Леса варианта распространены в средней и южной тайге. В первой из подзон они известны от юга и юго-востока Архангельской обл. до Печорского Предуралья и предгорий Северного Урала. На Среднем Урале отмечены также в горной средней тайге хребта Басеги в Пермском крае и от средней (Качканар) и южной (Висим) тайги до подтайги (Шамары) в Свердловской обл. (Koles-nikov et al., 1973). В южной тайге Костромской обл. вариант выступает самым распространенным типом заболоченных ельников в заповеднике “Кологривский лес” (P. filicoso-sphagnosum c господством Dryopteris carthusiana и D. expansa (Abaturov et al., 1988; Lugovaya, 2010)). Он описан также в южной тайге Вологодской (по р. Иде; P. filicosum var. sphagnosum (Korchagin, 1929)) и севера Кировской (P. equisetoso-sphagnosum (Smirnova A., 1954)) областей.
Без деления на субассоциации и варианты ельники хвощовые сфагновые приводятся для всей территории Ленинградской обл., где произрастают на торфяно-глеевых суглинистых почвах в понижениях рельефа (Zinserling, 1932; Nitsenko, 1960).
Ельники хвощовые сфагновые предгорий и гор Средней Европы описаны как ассоциация Equiseto sylvatici-Piceetum abietis Šmarda 1950. В их древесном ярусе к Picea abies примешивается Abies alba, реже Acer pseudoplatanus, Alnus glutinosa, Fagus sylvatica. Доминанты травяного яруса – Equisetum sylvaticum и Calamagrostis villosa, мохового – Polytrichum commune, P. formosum, Sphagnum girgensohnii, S. palustre. В числе диагностических видов Circaea alpina, Crepis paludosa, Phegopteris connectilis, Dryopteris dilatata (D. expansa s.l.), Lycopodium annotinum, Sphagnum squarrosum сочетаются с Soldanella montana, Luzula sylvatica, Bazzania trilobata, Mylia taylori, Mnium hornum. Почвы подзолисто-глеевые либо оглеенные грубогумусные с маломощным слоем торфа. Сообщества отмечены от Западных Карпат до Рудных гор, Чешского леса (Jirásek, 1996; Chytrý et al., 2013) и Северных Альп (Willner, Grabherr, 2007). В Шварцвальде, видимо, такие же ельники, развитые в условиях проточного переувлажнения, отнесены к синтаксону Bazzanio-Piceetum Br.-Bl. et Siss. in Br.-Bl. et al. 1939 subass. typicum var. Equisetum sylvaticum Oberd. 1992 (Oberdorfer, 1992).
Крупнопапоротниково-хвощовые ельники гор Норвегии (bregne-snelle-gran-utforming) с моховым покровом из Rhytidiadelphus subpinnatus и Eurhynchium angustirete (Fremstad, 1998) следует относить не к сфагновой, а к травяной группе ассоциаций. В системе Ж. Браун-Бланке они описаны как Eu-Piceetum abietis (Caj. 1921) K.-Lund 1962 subass. athyrietosum K.-Lund 1981, Equisetum-variant (Kielland-Lund, 1981).
P. sphagno girgensohnii-humulifolii-ruboso-equisetosum (Sphagno girgensohnii-Rubo humulifolii-P.) – е. хмеленично-хвощовый сфагновый (табл. 1, рис. 2: № 6–9). Характерная ассоциация таежной зоны Двино-Печорского и Уральского регионов. Отчасти замещает предыдущую ассоциацию к востоку, отчасти перекрывается с ней в своем распространении, демонстрируя при этом и некоторые черты произрастающих севернее ельников морошково-хвощовых. Леса развиваются на почвах разного гранулометрического состава равно на силикатных и на карбонатных породах. Бонитет древостоя ближе к таковому в ельниках морошково-хвощовых (III–IV), примесь Pinus sylvestris не выражена. Аспектирующий детерминант ассоциации – евросибирский бореальный мезотрофный гигромезофит Rubus humulifolius (малина хмелелистная, она же хмеленика по словарю В.И. Даля). Входя в “свиту” Abies sibirica, она, тем не менее, распространилась к западу от границы ее ареала, в основном в бореальном периоде голоцена (Minyaev, 1966). В группу детерминантов также входят гипоарктические гигромезофиты (Rubus arcticus, R. chamaemorus) и бореальные мезофиты (Rosa acicularis). Кроме олигомезотрофной Rubus chamaemorus, все они тоже мезотрофы. Наряду с этими видами постоянны Maianthemum bifolium, Oxalis acetosella и другие детерминанты, общие с ельниками хвощовыми сфагновыми (табл. 1), в большинстве подчиненных синтаксонов также Rubus saxatilis и Luzula pilosa. Dryopteris carthusiana и Trientalis europaea всегда играют роль спутников. Моховой ярус обычно сомкнут. Преобладание Sphagnum girgensohnii менее выражено, чем в лесах предыдущей ассоциации; ему сопутствуют другие виды рода. Выделяются три субассоциации.
Рис. 2.
Распространение мезотрофных ельников осочково-хмеленично-хвощовых (6) и хмеленично-хвощовых (7), пихто-ельников хмеленично-хвощовых (8) и щитовниково-хмеленично-хвощовых (9) сфагновых в Европейской России и на Урале. Обозначения как на рис. 1.
Fig. 2. Distribution of Piceetum (P.) sphagno girgensohnii-humulifolii-ruboso-equisetosum subass. caricosum dispermae (6), subass. typicum (7), and subass. abietosum sibiricae var. typica (8) and var. Dryopteris expansa (9) in European Russia and the Urals. For other notes and symbols see Fig. 1.
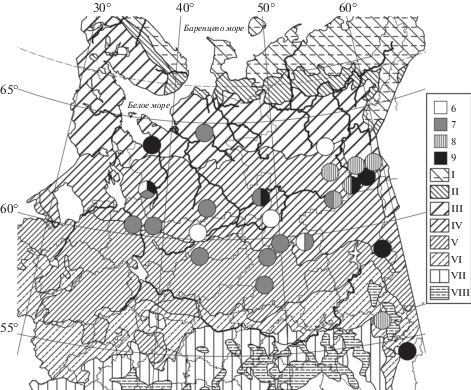
Subass. caricosum dispermae – е. осочково-хмеленично-хвощовый сфагновый (табл. 1, рис. 2: № 6). Сообщества заболоченных понижений на водоразделах, окраин болот и сплавин. Мощность торфа от 15 до 70–120 см. Нанорельеф ровный или сглаженный мелкокочковатый с примерно равной долей кочек и ковров. Примесь Be-tula pubescens s.l. к Picea abies s.l. в 1-м ярусе древостоя умеренна; 2-й ярус и подрост чисто еловые. Подрост групповой, умеренно высокий; деление на подъярусы не выражено. В подлеске Sorbus aucuparia s.l. и единичная Rosa acicularis. В кустарничково-травяном ярусе отчетливо преобладает Equisetum sylvaticum. Доминанты 2-го порядка – Vaccinium myrtillus и Carex globularis. Сопутствующие виды – Gymnocarpium dryopteris, Vaccinium vitis-idaea, Rubus chamaemorus, R. arcticus, Maianthemum bifolium, Oxalis acetosella; к ним здесь относится и Rubus humulifolius. Нетипичны R. saxatilis и Luzula pilosa. Моховой ковер вместе со Sphagnum girgensohnii формируют S. angustifolium, S. russowii и S. riparium – три доминанта 2-го порядка. На кочках и при стволах преобладает Polytrichum commune в сопровождении Pleurozium schreberi, Hylocomium splendens и Sphagnum wulfianum.
Детерминанты субассоциации – болотно-лесные гигромезо- и мезогигрофиты от мезотрофных (Carex disperma, Sphagnum riparium) до олиготрофных (S. angustifolium). Олигомезотрофные мхи, гигромезофиты (S. russowii) и мезофиты (Pti-lium crista-castrensis), объединяют ее с остальными субассоциациями за вычетом пихто-ельников щитовникового варианта. С упомянутыми пихто-ельниками и с ельниками хмеленично-хвощовыми subass. typicum синтаксон сближают мезотрофные (Calamagrostis langsdorffii (C. purpurea s.l.), Viola epipsila, Listera cordata, Rhizomnium pseudo-punctatum) и олигомезотрофные (Sphagnum flexuosum) гигромезофиты. С ельниками subass. typicum, а также крупнопапоротниково-хвощовыми var. Circaea alpina ее объединяет и S. wulfianum, а с хвощовыми типичной субассоциации – Solidago virgaurea.
Ельники этой субассоциации встречаются в основном в средней, реже в северной (Нижний Одес) тайге Двино-Печорского региона. Они описаны из НП “Кенозерский” на юго-западе Архангельской обл., в Тотемском р-не Вологодской обл. (P. sphagnosum var. caricosum на лессовидных суглинках (Korchagin, 1929)) и на междуречье Лузы, Сысолы и Вычегды на юге Коми (P. equisetoso-sphagnosum (Yudin, 1948, 1954)). К югу от упомянутого региона сообщества известны в Кайском р-не на севере Кировской обл. (P. equisetoso-sphagnosum (Smirnova A., 1954)).
Subass. typicum (табл. 1, рис. 2: № 7). Сообщества долин ручьев и речных террас, реже встречаются по окраинам болот. Торфяная залежь мощностью 15–50, вдоль болот до 70 см. Выражена кочковатость; высота кочек 15–80 см. Примесь Betula pubescens s.l. к Picea abies s.l. невелика. Подрост P. abies умеренно обильный, групповой, стратифицирован на три подъяруса с примесью Sorbus aucuparia s.l. (S. × gorodkovii, S. sibirica) в каждом из них. Кустарничково-травяной ярус средней густоты, в нем согосподствуют Equisetum sylvaticum и Vaccinium myrtillus. Rubus humulifolius пополняет список доминантов 2-го порядка, в который входят также Maianthemum bifolium, Vaccinium vitis-idaea и Carex globularis. Oxalis acetosella и Gymnocarpium dryopteris – их спутники. Постоянство Rubus arcticus и R. chamaemorus снижено. В сфагновом ковре и на низких кочках к Sphagnum girgensohnii иногда примешиваются S. wulfianum и/или S. russowii. Hylocomium splendens и Pleurozium schreberi – доминанты 2-го порядка при стволах; Polytrichum commune – сопутствующий вид.
С осочковой субассоциацией и с типичным вариантом пихто-ельников хмеленично-хвощовых субассоциацию сближает группа Ptilium crista-castrensis–Sphagnum russowii, со щитовниковым вариантом – группа Calamagrostis purpurea–Viola epipsila. С осочковой субассоциацией и с ельниками var. Circaea alpina синтаксон объединяет Sphagnum wulfianum. Собственная дифференциация негативная, по отсутствию видов из группы Carex disperma–Sphagnum angustifolium (табл. 1).
Ельники синтаксона встречаются в средней и южной тайге Двино-Печорского региона, а также территорий, граничащих с ним с юга. Они описаны по всему югу Архангельской обл. от НП “Кенозерский” (Rubo humulifolii-Piceetum (Kucherov et al., 2010)) до низовий Вычегды, в средней тайге Вологодской обл. известны из Чарондского края (“P. polytrichosum” с Equisetum sylvaticum, Gymnocarpium dryopteris и Rubus humulifolius на карбонатах (Shimanyuk, 1931)) и заказника “Ярбозерский бор” на правобережье Кемы. В Коми они найдены в бассейне р. Локчим (P. equisetoso-sphagnosum (Yudin, 1948, 1954)), а на северо-западе Пермского края – в Гайнском р-не (“P. polytrichosum” (Igoshina, 1930)). В южной тайге сообщества отмечены по р. Иде в Тотемском р-не Вологодской обл. (P. equisetosum var. sphagnosum (Korchagin, 1929)), на севере Кировской обл. (P. equisetoso-sphagnosum, P. equisetoso-sphagnosum var. sorbosum (Smirnova A., 1954)) и в центральной ее части (“ельник сфагновый” c Equisetum sylvaticum и Rubus humulifolius, без Abies sibirica (Vasilevich, 2003)). Единственная находка в северной тайге – в бывшем заказнике “Голубинский” на Средней Пинеге.
Subass. abietosum sibiricae – пихто-ельник хмеленично-хвощовый сфагновый (табл. 1, рис. 2: № 8–9). Леса заболоченных понижений и пологих склонов водоразделов, реже встречаются по долинам ручьев и берегам рек в восточной части Двино-Печорского региона и на Урале. В нано-рельефе преобладают участки ковров; кочки редкие, но высокие (20–80 см); всегда присутствуют вывалы с западинами под ними. Леса со смешанным древостоем из Picea abies s.l. (чаще всего P. obovata) и Abies sibirica с примесью Pinus sibirica, с обильным двухъярусным подростом и подлеском. В число доминантов кустарничково-травяного яруса, помимо Equisetum sylvaticum и Vaccinium myrtillus, всегда входит Gymnocarpium dryopteris. Rubus humulifolius – постоянный спутник этих видов. В моховом ярусе доминантом 2‑го порядка изредка становится Rhytidiadelphus triquetrus. Нехарактерен Sphagnum wulfianum. Детерминанты субассоциации – виды из группы Abies sibirica–Pinus sibirica, общие с пихто-ельниками щитовниково-хвощовыми (табл. 1). Выделены два варианта.
Var. typica (табл. 1, рис. 2: № 8). Преимущественно равнинные и предгорные леса. Мощность торфяной залежи 20–50 см. Древостой пихтово-еловый, густой, но высота его снижена. Примесь Betula pubescens s.l. умеренна, Pinus sibi-rica – мала; изредка есть единичная примесь Larix sibirica в 1-м и Betula pubescens s.l. во 2-м ярусе. Ярус подроста и подлеска тоже сильно снижен. Подрост пихтовый с примесью ели до 1/3 по составу; единичен подрост Pinus sibirica. Sorbus aucuparia s.l. малообильна, как и в лесах subass. typicum. В нижнем подъярусе постоянна Rosa acicularis. В кустарничково-травяном ярусе Gymnocarpium dryopteris превосходит по покрытию Equisetum sylvaticum; на 3-м месте Vaccinium myrtillus. В число доминантов 2-го порядка входят Rubus chamaemorus и Carex globularis, а также Linnaea borealis. Постоянные сопутствующие виды яруса – Maianthemum bifolium, Oxalis acetosella и Vaccinium vitis-idaea. Существенно возрастает покрытие Pleurozium schreberi и Hylocomium splendens, в сумме равное ПП Sphagnum girgensohnii; фактически это сфагново-зеленомошный пихто-ельник. Polytrichum commune – доминант 2-го порядка.
Детерминантная группа Ptilium crista-castrensis–Sphagnum russowii объединяет вариант с subass. typicum. В остальном дифференциация сугубо негативная – по отсутствию видов из групп, свойственных следующему синтаксону (табл. 1).
Леса варианта встречаются от северной тайги до подтайги в бассейне Печоры и на Урале. В северной тайге они описаны в долине р. Илыч (P. linnaeani-dryopteridoso-sphagnosum, P. myrtilloso-sphagnosum, P. chamaemoroso-equisetoso-sphagnosum (Korchagin, 1940)), а в средней – в бассейне р. Северной Кельтмы (P. linnaeani-dryopteridoso-sphagnosum (Yudin, 1948, 1954)), на Южном Тимане (массив Потчурк (Nepomilueva, Duryagina, 1985)) и в Якшинском участке Печоро-Илычского заповедника. Синтаксон приводится как сравнительно частый для всей средней и северной тайги Республики Коми (“ельник мелкотравно-сфагновый” (Yudin, 1948, 1954)). В подтайге Свердловской обл. есть указания для Уфимского плато (Kolesnikov et al., 1973).
Var. Dryopteris expansa – пихто-ельник щитовниково-хмеленично-хвощовый сфагновый (табл. 1, рис. 2: № 9). Наряду с равнинами и предгорьями леса характерны также для шлейфов горных склонов. Торфяная залежь маломощная, до 30 см. Древостой преимущественно еловый. Он несколько разрежен, но его высота и бонитет заметно возрастают. Примесь Abies sibirica и Betula pubescens s.l. незначительна, Pinus sibirica – единична. Около 1/2 ПП яруса подроста и подлеска приходится на Picea abies s.l., примерно по 1/4 – на Abies sibirica и Sorbus aucuparia s.l. Верхний подъярус высокий; в нижнем в составе низкого подлеска помимо Rosa acicularis присутствует Rubus idaeus. В кустарничково-травяном ярусе согосподствуют очень обильный Equisetum sylvaticum, Gymnocarpium dryopteris и Dryopteris expansa s.l. Доминанты 2-го порядка – Phegopteris connectilis и Oxalis acetosella. В числе сопутствующих видов как Athyrium filix-femina и Rubus saxatilis, так и Vaccinium myrtillus, V. vitis-idaea, Maianthemum bifolium, Linnaea borealis, Carex globularis. Постоянство Rubus chamaemorus вновь резко снижается. Ярус мхов разрежен. ПП Sphagnum girgensohnii снижено; к нему иногда примешиваются S. flexuosum или Plagiomnium ellipticum. При стволах константным доминантом 2-го порядка выступает лишь Pleurozium schreberi; Hylocomium splendens и Polytrichum commune – сопутствующие виды.
Детерминантную группу варианта формируют евразиатские и евросибирские мезотрофные (Calamagrostis obtusata, Melica nutans) и мезоэвтрофные (Veratrum lobelianum, Paris quadrifolia) лесные и лугово-лесные мезофиты, отчасти также мезоэвтрофные гигромезофиты (Bistorta major, Viola biflora). К группе примыкают и мезотрофные мхи с более протяженными ареалами, от мезофитов (Plagiothecium denticulatum, Rhytidiadel-phus subpinnatus) до мезогигрофитов (Pellia neesiana). Следует отметить, однако, что высоким постоянством в составе группы отличается лишь Veratrum lobelianum, прочие виды встречаются реже. В то же время группа Calamagrostis purpurea–Viola epipsila сближает вариант с осочковой и типичной субассоциациями. Вдобавок группа Rubus idaeus–Dryopteris expansa отличает вариант от типичного и сближает со щитовниковой субассоциацией ельников хвощовых, взятой в целом. Группа Dicranum scoparium–Sciuro-hypnum curtum объединяет его с типичным вариантом щитовниковой субассоциации, а группа Athyrium filix-femina–Crepis paludosa – c вариантами Circaea alpina и Convallaria majalis, наконец, группа Milium effusum–Stellaria holostea – только с последним (табл. 1).
Наиболее обычны пихто-ельники данного варианта в горной средней тайге Северного (Верхняя Печора) и Среднего (хребет Басеги) Урала. Из Басег синтаксон описан и ранее (P. equisetoso-sphagnosum с Rubus humulifolius, R. arcticus, R. chamaemorus, Crepis paludosa, Bistorta major, Cala-magrostis langsdorffii (Balandin, Ladygin, 2001)). Редколесья без Pinus sibirica и Dryopteris expansa, но с Rubus arcticus, Milium effusum и обильным Athyrium filix-femina отмечены также на склоне г. Большой Иремель в горной северной тайге Южного Урала (Rubo arctici-Piceetum obovatae Ishbirdin et al. 1996 приручьевого варианта (Ishbirdin et al., 1996)).
Реже сообщества встречаются в равнинной средней тайге от низовий Вычегды до Предуралья. Изолированные находки отмечены и западнее – в Лекшмозерье и (в северной тайге) в низовьях Онеги. Оба участка леса находятся за пределами ареала Abies sibirica, но в их травяном покрове присутствуют виды ее “свиты”, вероятные бореальные мигранты (Minyaev, 1966).
ЗАКЛЮЧЕНИЕ
Всего в Европейской России и на Урале нами описано 2 ассоциации мезотрофных ельников хвощовых сфагновых с 5 субассоциациями и 6 вариантами.
Распределение синтаксонов по градиенту теплообеспеченности соответствует их зональным ареалам (табл. 2), как и в ельниках черничных сфагновых (Kucherov, Kutenkov, 2019 b) и морошково-хвощовых (Kucherov, Kutenkov, 2020). Ельники хвощовые сфагновые типичной субассоциации распространены от южных границ северной тайги до подтайги, но главным образом в средней и южной тайге. Ельники щитовниково-хвощовые типичного и пихтового вариантов также тяготеют к средней и южной, вариантов Convallaria majalis и Circaea alpina – пока найдены лишь в южной тайге. Ельники осочково-хмеленично-хвощовые и пихто-ельники щитовниково-хмеленично-хвощовые произрастают от южной полосы северной тайги до средней тайги, ельники хмеленично-хвощовые типичные – в средней и южной тайге, пихто-ельники типичного варианта – также в подтайге.
Всем синтаксонам свойствен тот или иной тип секторальной приуроченности; их распространение соответствует изменениям континентальности климата (табл. 2). Ельники щитовниково-хвощовые вариантов Convallaria majalis и Circaea alpina пока отмечены лишь в Ленинградской обл. Ареал ельников хвощовых сфагновых subass. typicum прослежен от Западной Двины до Заволжья, а щитовниково-хвощовых subass. typicum – от Карельского перешейка до Среднего Поволжья. С другой стороны, ельники хмеленично-хвощовые осочковой и типичной субассоциаций встречаются в Двино-Печорском регионе, пихтовой – также на Урале, щитовниково-хвощовые var. Abies sibirica – от Заволжья до Среднего и Южного Урала.
В своем долготном распространении ельники хвощовые сфагновые большинства синтаксонов не схожи с черничными сфагновыми, чьи ареалы не обусловлены изменениями континентальности (Kucherov, Kutenkov, 2019 b). Как и в ельниках морошково-хвощовых (Kucherov, Kutenkov, 2020), произрастание сообществ на богатых почвах усиливает влияние континентализации климата на их флористический состав, тяготение же к бедным почвам сглаживает это влияние.
Ельники щитовниково-хвощовые var. Convallaria majalis и var. Circaea alpina приурочены к силикатным породам. Леса всех прочих синтаксонов успешно произрастают как на силикатах, так и на карбонатах, т.е. нейтральны по отношению к фактору богатства почвообразующей породы.
При дифференциации южнотаежных ельников хвощовых сфагновых обращает на себя внимание та диагностическая роль, которую наравне с эпигейными видами играют эпифитные мхи и лишайники (табл. 1), ранее отмеченная также в ельниках черничных сфагновых (Kucherov, Kutenkov, 2019 b) и морошково-хвощовых (Kucherov, Kutenkov, 2020). Принято рассматривать эпифитные криптогамные синузии как независимые фитоценозы и классифицировать их отдельно от лесов, в которых они произрастают (Barkman, 1958; Mucina et al., 2016). Однако виды, слагающие эти синузии, в то же время могут служить дифференциальными и при выделении лесных синтаксонов в целом. Вероятно, дифференцирующая роль криптогамных эпифитов и эпиксилов свойственна большинству синтаксонов лесов умеренной зоны, учитывая высокую степень специфичности и одновременно однородности условий микроклимата в пределах лесного сообщества (Geiger, 1957; Stoutjesdijk, Barkman, 1992).
Следует отметить также факт активного проникновения инвазионных видов кустарников в сообщества заболоченных ельников южной тайги Северо-Запада России. Еще недавно эти леса служили примером сообществ со стабильной ценофлорой.
Список литературы
[A checklist…] Список лихенофлоры России. 2010. СПб. 194 с.
[Abaturov et al.] Абатуров Ю.Д., Зворыкина К.В., Орлов А.Я., Письмеров А.В. 1988. Типы леса. – В кн.: Коренные темнохвойные леса южной тайги (резерват “Кологривский лес”). М.: с. 48–129.
[Balandin, Ladygin] Баландин С.В., Ладыгин И.В. 2002. Флора и растительность хребта Басеги (Средний Урал). Пермь. 191 с.
Barkman J.J. 1958. Phytosociology and ecology of cryptogamic epiphytes. Assen. 628 p.
[Cherepanov] Черепанов С.К. 1995. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). СПб. 991 с.
Chytrý M., Zelený D., Návratilová J., Sádlo J. 2013. Boreokontinentální jehličnaté lesy. – In: Vegetace České republiky. T. 4: Lesní a křovinná vegetace. Praha. S. 380–432.
[Dmitrieva] Дмитриева Е.В. 1973. Ельники северо-западной части Карельского перешейка. – Лесоведение. 2: 51–64.
[Fedorchuk et al.] Федорчук В.Н., Нешатаев В.Ю., Кузнецова М.Л. 2005. Лесные экосистемы северо-западных районов России: Типология, динамика, хозяйственные особенности. СПб. 382 с.
Fremstad E. 1998. Vegetasjonstyper i Norge. 2. oppl. – NINA Temahefte 12: 1–279.
[Gavrilov, Karpov] Гаврилов К.А., Карпов В.Г. 1962. Главнейшие типы леса и почвы Вологодской области в районе распространения карбонатной морены. – Тр. ин-та леса и древесины. 52: 5–119.
Geiger R. 1957. The climate near the ground. Cambridge (Mass.). 494 p.
[Grozdov] Гроздов Б.В. 1950. Типы леса Брянской, Смоленской и Калужской областей. Брянск. 55 с.
Ignatov M.S., Afonina O.M., Ignatova E.A. 2006. Check-list of mosses of East Europe and North Asia. – Arctoa: Бриологический журнал. 15: 1–130.
[Igoshina] Игошина К.Н. 1930. Растительность северной части Верхне-Камского округа Уральской области. – Тр. биол. ин-та Перм. ун-та. 3 (2): 73–176.
[Isachenko, Lavrenko] Исаченко Т.И., Лавренко Е.М. 1980. Ботанико-географическое районирование. – В кн.: Растительность европейской части СССР. Л. С. 10–22.
[Ishbirdin et al.] Ишбирдин А.Р., Муллагулов Р.Ю., Янтурин С.И. 1996. Растительность горного массива Иремель: синтаксономия и вопросы охраны. Уфа. 108 с.
Jirásek J. 1996. Společenstva přirozenych smrčin Česke republiky. – Preslia. 67 (3–4): 225–259.
Kielland-Lund J. 1981. Die Waldgesellschaften SO Norwegens. – Phytocoenologia. 9 (1–2): 53–250.
[Kolesnikov et al.] Колесников Б.П., Зубарева Р.С., Смологонов Е.П. 1973. Лесорастительные условия и типы леса Свердловской области. Свердловск. 176 с.
[Korchagin] Корчагин А.А. 1929. К вопросу о типах леса по исследованию в Тотемском уезде Вологодской губернии. – В кн.: Очерки по фитосоциологии и фитогеографии. М. С. 287–327.
[Korchagin] Корчагин А.А. 1940. Растительность северной половины Печорско-Ылычского заповедника. – В кн.: Труды Печорско-Ылычского заповедника. Т. 2. С. 5–415.
[Korchagin, Senyaninova-Korchagina] Корчагин А.А., Сенянинова-Корчагина М.В. 1957. Леса Молого-Шекснинского междуречья (дубняки, черноольшанники и ельники). – Тр. Дарвинского гос. заповедника. 4: 291–402.
[Kucherov et al.] Кучеров И.Б., Разумовская А.В., Чуракова Е.Ю. 2010. Еловые леса национального парка “Кенозерский” (Архангельская область). – Бот. журн. 95 (9): 1268–1301.
[Kucherov, Kutenkov] Кучеров И.Б., Кутенков С.А. 2019a. Олиготрофные сфагновые и сфагново-зеленомошные ельники Европейской России и Урала. – Бот. Журн. 104 (1): 12–40. https://doi.org/10.1134/S000681361901006X
[Kucherov, Kutenkov] Кучеров И.Б., Кутенков С.А. 2019b. Мезотрофные кустарничковые сфагновые и сфагново-зеленомошные ельники Европейской России и Урала. – Бот. журн. 104 (6): 859–887. https://doi.org/10.1134/S0006813619060115
[Kucherov, Kutenkov] Кучеров И.Б., Кутенков С.А. 2020. Мезотрофные ельники морошково-хвощовые сфагновые северной и средней тайги Европейской России и Урала. – Бот. журн. 105 (3): 55–72. https://doi.org/10.31857/S0006813620030060
[Lugovaya] Луговая Д.Л. 2010. Влияние экотопических факторов и природопользования на разнообразие лесных сообществ севера Костромской области. – Экология. 1: 9–16.
[Minyaev] Миняев Н.А. 1966. История развития флоры северо-запада европейской части СССР с конца плейстоцена: Автореф. дис. … докт. биол. наук. Л. 38 с.
Mucina L., Bültmann H., Dierßen K., Theurillat J.-P., and 28 more participants. 2016. Vegetation of Europe: hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. – Appl. Veg. Sci. 19 (Suppl. 1): 3–264.
NASA surface meteorology and solar energy. 2006. http://eosweb.larc.nasa.gov (Accessed 15.05.2006).
[Nepomilueva, Duryagina] Непомилуева Н.И., Дурягина Г.А. 1985. Пространственная и временная изменчивость темнохвойных лесов Южного Тимана. – В кн.: Структура и видовой состав растительных сообществ Европейского Севера СССР. Сыктывкар. С. 5–18.
[Nitsenko] Ниценко А.А. 1960. Еловые леса Ленинградской области. – Вестн. Ленингр. ун-та. Сер. 3: Биол. 2 (21): 5–16.
Oberdorfer E. 1992. Süddeutsche Pflanzengesellschaften. Teil 4: Wälder und Gebüsche. 2 Aufl. Jena. A. Textband. 282 S. – B. Tabellenband. 580 S.
[Orlov et al.] Орлов А.Я., Кошельков С.П., Осипов В.В., Соколов А.А. 1974. Типы лесных биогеоценозов южной тайги. М. 230 с.
[Popov] Попов П.П. 2005. Ель европейская и сибирская: структура, интерградация и дифференциация популяционных систем. Новосибирск. 231 с.
[Safronova, Yurkovskaya] Сафронова И.Н., Юрковская Т.К. 2015. Зональные закономерности растительного покрова равнин Европейской России и их отображение на карте. – Бот. журн. 100 (11): 1121–1141.
[Sambuk] Самбук Ф.В. 1930. Наблюдения над сосновыми борами и ключевыми болотами долины и бассейна р. Облы, притока р. Луги. – Тр. Бот. музея АН СССР. 22: 277–310.
[Shaposhnikov] Шапошников Е.С. 1988. Ассоциации еловых лесов Центрально-Лесного государственного заповедника: Дис. … канд. биол. наук. Л. 216 с.
[Shimanyuk] Шиманюк А.П. 1931. Опыт изучения северных лесов. М.; Л. 104 с.
[Smirnova A.] Смирнова А.Д. 1954. Типы еловых лесов крайнего севера Кировской области. Ч. II. – Учен. записки Горьковск. ун-та. 25: 191–226.
[Smirnova Z.] Смирнова З.Н. 1928. Лесные ассоциации северо-западной части Ленинградской области. – Тр. Петергоф. естеств.-науч. ин-та. 5: 119–258.
Stoutjesdijk P., Barkman J.J. 1992. Microclimate, vegetation and fauna. Uppsala. 205 p.
[Sukachev] Сукачев В.Н. 1931. Руководство к исследованию типов леса. 3-е изд. М. 328 с.
[Tereshkin, Tereshkina] Терешкин И.С., Терешкина Л.В. 2006. Растительность Мордовского заповедника. Последовательные ряды сукцессий. – Тр. Мордовского гос. природн. заповедника им. П.Г. Смидовича. 7: 186–287.
Tukhanen S. 1980. Climatic parameters and indices in plant geography. – Acta Phytogeogr. Suec. 67: 1–105.
[Tzvelev] Цвелев Н.Н. 2004. Род Береза – Betula L. – В кн.: Флора Восточной Европы. М.; СПб. Т. 11. С. 65–85.
[Vasilevich] Василевич В.И. 2003. Еловые леса Кировской области. – Бот. журн. 88 (7): 1–11.
Willner W., Grabherr G. (eds.) 2007. Die Wälder und Ge-büsche Österreichs. Ein Bestimmungswerk mit Tabellen. Im 2 Bände. München. 608 S.
[Yudin] Юдин Ю.П. 1948. Темнохвойные леса Коми АССР (геоботаническая характеристика): Дис. … докт. биол. наук. Сыктывкар. 323 с.
[Yudin] Юдин Ю.П. 1954. Темнохвойные леса. – В сб.: Производительные силы Коми АССР. М.; Л. Т. 3. Вып. 1. С. 42–125.
[Zaugolnova, Khanina] Заугольнова Л.Б., Ханина Л.Г. 1996. Опыт разработки и использования баз данных в лесной фитоценологии. – Лесоведение. 1: 76–83.
[Zinserling] Цинзерлинг Ю.Д. 1932. География растительного покрова северо-запада европейской части СССР. Л. 376 с.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал