

Ботанический журнал, 2020, T. 105, № 6, стр. 523-541
ЖИЗНЕННЫЕ ФОРМЫ ЛИШАЙНИКОВ БЕЛАРУСИ
1 Гомельский государственный университет имени Франциска Скорины
246019 г. Гомель, ул. Советская, 104, Беларусь
2 Самарский национальный исследовательский университет имени академика С.П. Королева
443086 Самара, Московское шоссе, 34, Россия
* E-mail: tsurykau@gmail.com
Поступила в редакцию 05.12.2019
После доработки 21.01.2020
Принята к публикации 28.02.2020
Аннотация
В статье представлена система жизненных форм лишайников лесных сообществ, основанная на интеграции иерархической системы жизненных форм и морфолого-анатомического подхода. Всего выделено 30 групп биоморф, объединенных в 4 класса, 3 типа и 2 отдела. В рамках каждого класса показана эволюция в ряду групп жизненных форм, отражающая адаптацию лишайников к существованию в более освещенных и сухих условиях. Проведен биоморфологический анализ лихенобиоты Беларуси с использованием предложенной системы, выявивший преобладание эвритопных (42.7%) и мезофитных лесных таксонов (41.8%). Число лишайников, предпочитающих ксерофитные условия, относительно невелико (15.5%).
Возникновение симбиотических отношений между грибом и водорослью считается основным источником появления эволюционных инноваций, приведших к возникновению огромного разнообразия морфологических форм среди лишайник-образующих представителей отдела Ascomycota. Морфологические структуры такого уровня сложности не отмечены ни в одном из других таксонов грибных организмов (Honegger, 1991). Такое разнообразие морфологических и анатомических форм обусловлено адаптацией лишайников к разнообразным условиям окружающей среды, поскольку независимо от условий произрастания необходимо обеспечить фотобионту необходимое количество света, воздуха и влаги для эффективного осуществления процесса фотосинтеза (Hawksworth, 1988).
Несмотря на то, что первые попытки классификации разнообразия морфологических форм талломов лишайников были предприняты еще в конце XIX века (Zukal, 1891), а первые детализированные системы жизненных форм были созданы в 1920-х гг. (Frey, 1923), в настоящее время не существует общепринятой системы классификации жизненных форм лишайников. Наиболее широко используются две: принятая за рубежом неиерархическая система, основанная на морфолого-анатомическом подходе (Ahti et al., 1999, 2002, 2007, 2013; Nash et al., 2002, 2004, 2007; Smith et al., 2009; Thell, Moberg, 2011) и иерархическая система жизненных форм Н.С. Голубковой (Go-lubkova, 1983), в основу которой был положен эволюционно-экобиоморфологический принцип. Преимуществом последней является отражение эволюционных отношений разных жизненных форм лишайников (Muchnik, 2015), однако разработанная для анализа лихенобиоты Монголии система Н.С. Голубковой не охватывает все многообразие жизненных форм лишайников других природных территорий. Определенные шаги по доработке иерархической системы жизненных форм лишайников были сделаны С.А. Пристяжнюком применительно к территории субарктических тундр (Pristyazhnyuk, 1996а, 1996б). В частности, им были введены новые группы и подгруппы биоморф лишайников, характерных для п-ова Ямал. Однако, некоторые жизненные формы лишайников, характерные, в том числе и для лесных сообществ, до настоящего времени не отражены в иерархической системе Н.С. Голубковой (Himelbrant, Kuznetsova, 2014). Целью данной работы явилось дополнение вышеназванной системы для максимального охвата существующих жизненных форм лишайников, характерных для лесных сообществ, а также биоморфологический анализ лихенобиоты Беларуси с использованием предложенной системы.
Настоящая работа основана на анализе опубликованного ранее списка видов лишайников Беларуси (Tsurykau, 2018), составленного на основании результатов собственных полевых исследований 2003–2017 гг., проведенных ревизий отдельных систематических групп лишайников (роды Cetrelia, Hypotrachyna, Lepraria, Parmotrema, Punctelia, Xanthoparmelia, группа Cladonia chlorophaea-pyxidata) (Bely et al., 2014a; Tsurykau, Golubkov, 2015; Tsurykau et al., 2015, 2016a, 2018a), а также анализе содержания 386 статей и материалов конференций, опубликованных за почти 240-летнюю историю изучения лишайников Беларуси (1781–2017). Названия таксонов лишайников приведены согласно вышеупомянутому списку (Tsurykau, 2018).
Следуя современному представлению о жизненной форме растений (Mirkin, Naumova, 2014), в настоящей работе под жизненной формой лишайника понимается общий его облик (габитус), обусловленный своеобразием морфологической и анатомической структуры таллома, формирующейся в результате роста и развития в определенных условиях среды. При оценке жизненной формы лишайника учитывали два тезиса, ранее принятых для систем жизненных форм сосудистых растений (Serebryakov, 1962; Khokhryakov, 1981) и обычно учитываемых в лихенологии (Pristyazhnyuk, 1996а; Muchnik, 2015): один вид может быть представлен разными жизненными формами в различных частях ареала или в разных экологических условиях; индивидуум в процессе онтогенеза может сменить несколько жизненных форм. В качестве примера можно привести морфологическое описание Arthonia exilis (Flörke) Anzi из определителя лишайников СССР (Makarevicz, 1977) (в квадратных скобках указаны жизненные формы из предлагаемой системы, соответствующие вышеприведенной части описания) – слоевище тонкое до очень тонкого, цельное [плотнокорковый] или слегка трещиноватое [трещиноватый], иногда состоящее из слившихся мелких пятен, мелкозернистое до мелкобугорчатого [зернисто-бородавчатый], иногда мучнисто-зернистое [лепрозный], изредка заметное в виде мелких зерен только вокруг апотециев [аталлический]. Поэтому для видов с вариабельной морфологией таллома в качестве основной (учитываемой в анализе) жизненной формы принимали наиболее часто встречаемую на территории региона исследований (для A. exilis – плотнокорковую).
Характеристики жизненных форм с некоторыми изменениями следуют описаниям, предложенным Д.Е. Гимельбрантом и Е.С. Кузнецовой (Himelbrant, Kuznetsova, 2014).
Степени сухости (аридности) и освещенности (инсоляции) местообитаний лишайников оценивались по 5-балльной шкале согласно методике, разработанной P.L. Nimis, S. Martellos (2017).
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Принятые для лихенобиоты Беларуси ранги жизненных форм представлены в табл. 1. Ниже приводим характеристику выделенных рангов. В качестве примеров приведены наиболее типичные для данной жизненной формы представители лихенобиоты Беларуси.
ОТДЕЛ I. ЭНДОГЕННЫЕ. Слоевище развивается внутри субстрата.
ТИП 1. Плагиотропные. Слоевище ориентировано по отношению к субстрату горизонтально.
Класс 1.1. Накипные.
Группа 1.1.1. Эндосубстратные. Характеризуются полностью погруженной в субстрат вегетативной частью таллома (Acrocordia gemmata (Ach.) A. Massal., Chaenotheca xyloxena Nádv., Leptorhaphis atomaria (Ach.) Szatala, Sarcogyne regularis Körb., Xylographa paralella (Ach. : Fr.) Fr. и др.).
В своей системе жизненных форм Н.С. Голубкова (Golubkova, 1983) выделяет группы эндофлеодных и эндолитных жизненных форм. Однако, обоснование данных групп основано на субстратной приуроченности лишайника и является скорее предметом субстратно-экологического анализа лихенобиоты. С точки зрения биоморфологии жизненная форма у эндофлеодных и эндолитных лишайников идентична – талломы представлены пронизывающими субстрат грибными гифами, ассоциированными (постоянно или на отдельных этапах онтогенеза) с водорослевыми клетками, находящимися в поверхностном слое субстрата. Поэтому в представленной классификационной системе эндофлеодные и эндолитные лишайники объединены в одну группу жизненных форм – эндосубстратные (см. также Himelbrant, Kuznetsova, 2014).
В Беларуси большинство лишайников этой жизненной формы являются мезофитными таксонами. Единичные виды (Sarcogyne regularis Körb.) приурочены к достаточно засушливым условиям.
Вопрос о положении эндогенных биоморф в филогенетических рядах жизненных форм и их роли в эволюции лишайникового симбиоза поднимался ранее (Muchnik, 2015). Обычно эндогенные биоморфы рассматривают как более высокую эволюционную ступень по сравнению с эпигенными лишайниками, обосновывая это наличием у некоторых эндолитных представителей анатомических поверхностных структур (литокортекс), напоминающих коровой слой хорошо развитых стратифицированных биоморф (Büdel, Scheidegger, 2008). Следует, однако, отметить, что и у некоторых представителей с лепрозными талломами, признаваемыми обычно в качестве исходных форм при возникновении симбиотических отношений между грибом и водорослью, наблюдается тенденция к усложнению анатомии слоевища, в частности, появление гипоталлома, морфологически и функционально схожего с сердцевинным слоем стратифицированных талломов (Lendemer, 2011). Считается, что большинство эндосубстратных видов находится на пути к утрате фотобионта и возвращению к сапротрофному питанию (Lutzoni et al., 2001). Подобные экологические стратегии характерны для разных систематических групп грибов, например, многих представителей подклассов Ostropomycetidae (Lecanoromycetes) и Chaetothyriomycetidae (Eurotiomycetes) (Gueidan et al., 2008; Spribille et al., 2014). С таксономической точки зрения эндосубстратные лишайники относятся к разным классам отдела Ascomycota (Coniocybomycetes, Dothideomycetes, Eurotiomycetes, Lecanoromycetes) (Wijayawardene et al., 2018) и потому представляют собой полифилетическую группу грибов, имеющих различную степень интеграции с водорослевым компонентом. К сожалению, на сегодняшний день эндосубстратные лишайник-образующие грибы слабо изучены с применением молекулярных методов исследований, и потому вопросы об их происхождении и эволюции остаются открытыми.
ОТДЕЛ II. ЭПИГЕННЫЕ. Слоевище развивается на поверхности субстрата.
ТИП 2. Плагиотропные. Слоевище ориентировано по отношению к субстрату горизонтально.
Класс 2.1. Накипные. Включает представителей, плотно прикрепленных к субстрату всей нижней поверхностью (Pristyazhnyuk, 1996а) или большей ее частью, при этом не имеющих нижнего корового слоя.
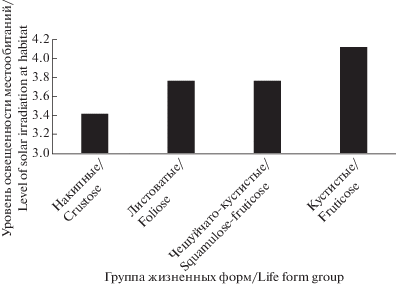
Н.С. Голубкова разделила класс накипных эпигенных лишайников на три группы – однообразно-накипных, диморфных и чешуйчатых жизненных форм – выделив в каждой от трех до пяти подгрупп. Однако, при детальном изучении неиерархической системы биоморф (Himelbrant, Kuznetsova, 2014) можно проследить плавную градацию анатомо-морфологических признаков в ряду накипных лишайников от однообразно-накипных до чешуйчатых через ряд промежуточных форм, отражающих постепенную смену их экологических адаптаций к высокому уровню инсоляции и аридности местообитаний (рис. 1). В представленной системе жизненных форм эти промежуточные формы представлены в виде групп. Таким образом, низшая таксономическая единица во всех классах системы имеет одинаковый ранг (подгруппы, характерные лишь для некоторых классов системы Н.С. Голубковой, не выделяются).
Рис. 1.
Приуроченность накипных жизненных форм лишайников к местообитаниям различного уровня освещенности и аридности.
Fig. 1. The association of crustose lichen life forms with habitats of different levels of solar irradiation and aridity.
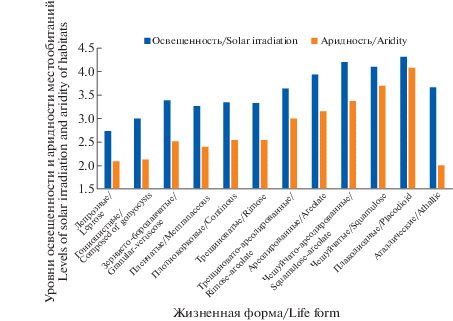
Группа 2.1.1. Аталлические. Характеризуются слабо развитым талломом, состоящим из отдельных мелких бугорков, обычно примыкающих к апотециям (некоторые виды рода Thelocarpon Nyl.).
Как и эндосубстратные, аталлические формы признаются большинством исследователей результатом вторичного упрощения слоевищ при переходе к сапротрофному питанию (Oxner, 1974; Pristyazhnyuk, 1996а; Muchnik, 2015). Все известные аталлические лишайники Беларуси (Thelocarpon impressellum Nyl., T. intermediellum Nyl., T. laureri (Flot.) Nyl.) обитают на разрушающейся древесине в лесных сообществах.
Таблица 1.
Состав жизненных форм лишайников Беларуси Table 1. Growth forms of lichens of Belarus
| Отдел/Division | Тип/Type | Класс/Class | Группа/Group |
|---|---|---|---|
| Эндогенные 19 (3.3%) Endogenous | Плагиотропные 19 (3.3%) Plagiotropic | Накипные 19 (3.3%) Crustose | Эндосубстратные 19 (3.3%)/Endosubstratal |
| Эпигенные 565 (96.7%) Epigenous | Плагиотропные 461 (78.9%) Plagiotropic | Накипные 345 (59.0%) Crustose | Аталлические 3 (0.5%)/Athallic (sheath around ascomata) |
| Лепрозные 20 (3.4%)/Leprose | |||
| Гониоцистные 16 (2.7%)/Composed of goniocysts | |||
| Зернисто-бородавчатые 102 (17.5%)/Granular-verrucose | |||
| Пленчатые 9 (1.5%)/Membranaceous | |||
| Плотнокорковые 82 (14.1%)/Continuous | |||
| Трещиноватые 28 (4.8%)/Rimose | |||
| Трещиновато-ареолированные 37 (6.3%)/Rimose-areolate | |||
| Ареолированные 23 (3.9%)/Areolate | |||
| Чешуйчато-ареолированные 12 (2.1)/Squamulose-areolate | |||
| Чешуйчатые 6 (1.0%)/Squamulose | |||
| Плакодиоидные 7 (1.2%)/Placodioid | |||
| Листоватые 116 (19.9%) Foliose | Умбиликатные 2 (0.3%)/Umbilicate | ||
| Широколопастные 32 (5.5%)/Broad-lobed | |||
| Среднешироколопастные 24 (4.1%)/Medium broad-lobed | |||
| Узколопастные 38 (6.5%)/Narrow-lobed | |||
| Вздутолопастные 5 (0.9%)/Hollow-lobed | |||
| Студенистые 15 (2.6%)/Gelatinous | |||
| Плагио-ортотропные 62 (10.6%) Plagio-orthotropic | Бородавчато- и чешуйчато-кустистые 62 (10.6%) Granular-fruticose or squamulose-fruticose |
Шиловидные 10 (1.7%)/Subulate | |
| Палочковидные 9 (1.5%)/Bacilliform | |||
| Сцифовидные 28 (4.8%)/Scyphose | |||
| Кустисто-разветвленные 15 (2.6%)/Richly branched | |||
| Ортотропные 42 (7.2%) Orthotropic | Листоватые 6 (0.9%) Foliose | Субфрутикозные 2 (0.3%)/Subfruticose | |
| Повисающие 2 (0.3%)/Pendent | |||
| Прямостоячие 4 (0.3%)/Erect | |||
| Кустистые 36 (6.3%) Fruticose | Филаментозные 1 (0.2%)/Filamentose | ||
| Повисающие 16 (2.8%)/Pendent | |||
| Прямостоячие 18 (3.1%)/Erect | |||
| Распростертые 1 (0.2%)/Decumbent |
Группа 2.1.2. Лепрозные. Характеризуются нестратифицированным талломом, состоящим из отдельных мелких элементов (соредий) и лишенным хорошо выраженных анатомических структур (представители рода Lepraria Ach., Chaenotheca furfuracea (L.) Tibell, Cliostomum leprosum (Räsänen) Holien et Tønsberg и др.).
Подробное изучение видового состава лишайников в 12 типах разновозрастных сосновых насаждений юго-востока Беларуси, отражающих весь трофогенный и гигрогенный ряд, показало, что лепрозные формы (Chaenotheca stemonea (Ach.) Müll. Arg., Lepraria elobata Tønsberg, L. incana (L.) Ach. и нек. др.) приурочены к свежим и влажным древостоям, обладающим сциофитными и мезофитными условиями (Tsurykau, Khramchankova, 2015). Приуроченность лепрозных лишайников к подобным местообитаниям отмечалась ранее (Golubkova, 1983; Olszewska et al., 2014) и, по-видимому, характерна для большинства представителей этой жизненной формы в Беларуси (Tsurykau et al., 2016b, 2016c, 2016d, 2016e). Открытые местообитания предпочитают лишь единичные виды, например, Lepraria rigidula (B. de Lesd.) Tønsberg (Kukwa, 2006; Tsurykau et al., 2016d). По-видимому, приуроченность лишайников с лепрозным типом таллома к закрытым от солнечного освещения, хорошо увлажненным микронишам (сформированные мезофитные лесные сообщества, трещины в корке деревьев, углубления в почве, затененные, обычно покрытые мхом участки каменистого субстрата) связана с отсутствием корового слоя, выполняющего фотопротекторную функцию (Büdel, Scheidegger, 2008).
Группа 2.1.3. Гониоцистные. Характеризуются нестратифицированным талломом, состоящим из дискретных, более или менее шаровидных гранул, представляющих собой группы клеток фотобионта, окруженных грибными гифами (некоторые представители родов Bacidina Vězda, Micarea Fr., Placynthiella Elenkin, Vezdaea Tsch.-Woess et Poelt и др.).
Поскольку в талломах лишайников с гониоцистным талломом доля водорослевых клеток по отношению к грибному компоненту достаточно велика, а также ввиду отсутствия развитого корового слоя, большинство видов приурочено к хорошо увлажненным местообитаниям. Некоторые виды (Micarea prasina Fr. s. str.) предпочитают разлагающуюся древесину в сомкнутых лесных сообществах (Launis et al., 2019), другие (Placynthiella dasaea (Stirt.) Tønsberg) часто встречаются между дерновинок мхов или чешуек первичного слоевища представителей рода Cladonia Hill ex P. Browne, долго удерживающих влагу. Отдельные представители являются эфемерными лишайниками (Thelidium zwackhii (Hepp) A. Massal.), заселяющими возникающие ненадолго микрониши в достаточно увлажненной почве по берегам ручьев или обочинам дорог (Orange, 2009).
Группа 2.1.4. Зернисто-бородавчатые. Характеризуются стратифицированным талломом, состоящим из дискретных зернистых или бородавковидных элементов, покрытых коровым слоем (многие представители родов Bacidia De Not., Calicium Pers., Candelariella Müll. Arg., Chaenotheca (Th. Fr.) Th. Fr., Trapeliopsis Hertel et Gotth. Schneid. и др.).
Виды с зернисто-бородавчатым талломом представляют собой экологически гетерогенную группу и заселяют любые виды доступных субстратов, часто проявляя широкую экологическую пластичность. Отдельные таксоны (Candelariella aurella (Hoffm.) Zahlbr., Cyphelium notarisii (Tul.) Blomb. et Forssell, Trapeliopsis granulosa (Hoffm.) Lumbsch и др.) предпочитают местообитания с явно выраженными ксерическими чертами, в то время как другие виды являются обитателями сомкнутых мезофитных лесных сообществ с постоянно влажным климатом (Catillaria croatica Zahlbr., Chaenotheca trichialis (Ach.) Th. Fr., Lichenomphalia umbellifera (L.: Fr.) Redhead, Lutzoni, Monclavo et Vilgalys и др.). Зернисто-бородавчатые лишайники широко представлены во всех типах растительных сообществ республики, обычны в городских и пригородных территориях, нередко колонизируют субстраты антропогенного происхождения (Tsurykau, 2013).
Группа 2.1.5. Пленчатые. Характеризуются нестратифицированным талломом, имеющим в увлажненном состоянии вид цельной слизистой пленки и тонкой слабозаметной корочки в сухом (Absconditella lignicola Vězda et Pišút, Jamesiella anastomosans (P. James et Vězda) Lücking, Sérus. et Vězda, Steinia geophana (Nyl.) Stein, Thrombium epigaeum (Pers.) Wallr. и др.).
Лишайники с пленчатым талломом встречаются в хорошо увлажненных, нередко заболоченных местообитаниях (Bely, 2012). Некоторые виды (Steinia geophana, Sarcosagium campestre (Fr.) Poetsch et Schied.) являются эфемерными видами, развивающимися в подходящих микронишах (расщелины в почве, разлагающиеся мхи) в осенний период. Ввиду особенностей их экологии эти виды лишайников редко собираются коллекторами и потому считаются редкими в большинстве стран Европы (Motiejūnaitė, 2006).
Группа 2.1.6. Плотнокорковые. Характеризуются стратифицированным цельным талломом (некоторые представители родов Arthonia Ach., Lecanora Ach., Opegrapha Ach. s. l., Pyrenula Ach. и др.).
Лишайники с плотнокорковым талломом являются типичными лесными таксонами, произрастающими преимущественно на коре хвойных и лиственных деревьев, часто доминируя над другими таксонами на стволах и ветвях древесных пород и кустарников с гладкой корой (Graphis scripta (L.) Ach., Phlyctis argena (Spreng.) Flot. и нек. др.). Некоторые виды (Arthonia fuliginosa (Schaer.) Flot., Phlyctis agelaea (Ach.) Flot., Zwackhia viridis (Ach.) Poetsch et Schied. и др.) приурочены к малонарушенным лесным сообществам и потому являются редкими на территории республики. Очень немногие представители плотнокорковых лишайников (например, Athallia pyracea (Ach.) Arup, Frödén et Søchting s. l., Arthrosporum populorum A. Massal., Lecanora pulicaris (Pers.) Ach.) встречаются на антропогенно нарушенных территориях и в других местообитаниях с относительно сухим микроклиматом.
Группа 2.1.7. Трещиноватые. Характеризуются стратифицированным талломом, пересеченным достаточно глубокими трещинами (достигающими сердцевинного слоя), не разделяющими его на обособленные элементы (ареолы) (некоторые представители родов Lecidella Körb., Lepra Scop., Ochrolechia A. Massal., Pertusaria DC.).
Появление глубоких трещин в талломе можно расценивать как первый шаг в адаптации лишайников к существованию в засушливых условиях. Несмотря на то, что некоторые лишайники этой группы отдают предпочтение мезофитным условиям (Lepra multipuncta (Turner) Hafellner, L. ophthalmiza (Nyl.) Hafellner, Ochrolechia alboflavescens (Wulfen) Zahlbr.), большинство представителей с трещиноватым типом таллома обладает широкой экологической пластичностью и встречается в различных местопроизрастаниях: естественных лесных сообществах, придорожных посадках, городских территориях (Amandinea punctata (Hoffm.) Coppins et Scheid., Buellia griseovirens (Turner et Borrer ex Sm.) Almb., Lecidella elaeochroma (Ach.) M. Choisy, L. euphorea (Flörke) Hertel, Rinodina exigua (Ach.) Gray, Verrucaria muralis Ach. и др.). Некоторые лишайники этой группы, являясь эпифитными видами, отдают предпочтение достаточно сухим местообитаниям, выбирая хорошо освещаемые стороны стволов часто одиночно стоящих деревьев (Cliostomum griffithii (Sm.) Coppins, Lecidella flavosorediata (Vezda) Hertel et Leuckert, Pertusaria coronata (Ach.) Th. Fr., P. flavida (DC.) J.R. Laundon и др.). Однако, лишайники с трещиноватым талломом не приурочены к типичным ксерическим условиям, уступая такие местообитания накипным видам с большей степенью расчлененности таллома (трещиновато-ареолированные, ареолированные) и/или более сложным уровнем морфологической организации слоевища (чешуйчато-ареолированные, чешуйчатые, плакодиоидные). В частности, многие эпилитные виды этой группы (например, Lecania hutchinsiae (Nyl.) A.L. Sm., Lecidella anomaloides (A. Massal.) Hertel et H. Kilias, Verrucaria aethiobola Wahlenb., Verrucaria praetermissa (Trevis.) Anzi) предпочитают полностью или частично затененные субстраты, часто расположенные в непосредственной близости от водных объектов.
Группа 2.1.8. Трещиновато-ареолированные. Характеризуются талломом, пересеченным глубокими трещинами, разделяющими его на обособленные элементы (ареолы), находящиеся на общем гипоталломе и имеющие не покрытые коровым слоем края (некоторые представители родов Brianaria S. Ekman et M. Svensson, Circinaria Link, Porpidia Körb., Rinodina (Ach.) Gray и др.).
Дальнейшее расчленение трещиноватого таллома на отдельные ареолы представляется следующим шагом в адаптации лишайников к ксерофитным условиям произрастания. По-видимому, формирование ареол явилось вынужденным приспособлением к существованию лишайников в переменных температурных условиях, в частности к линейному тепловому расширению субстрата произрастания в дневное время и его сжатию при остывании ночью. Типичным субстратом многих трещиновато-ареолированных лишайников являются хорошо освещаемые и прогреваемые силикатные и карбонатные каменистые породы. В частности, 27 видов, или 75% от общего числа известных в Беларуси лишайников с трещиновато-ареолированным типом таллома являются облигатными эпилитами. Как правило, это редкие лишайники, многие из которых известны из нескольких локалитетов на территории республики (Bacidina inundata (Fr.) Vězda, Circinaria sphaerothallina (J. Steiner) Sohrabi, Porpidia soredizodes (Lamy) J.R. Laundon, Protoparmelia badia (Hoffm.) Hafellner, Rinodina oxydata (A. Massal.) A. Massal., R. teichophila (Nyl.) Arnold, Varicellaria lactea (L.) I. Schmitt et Lumbsch и др.). Некоторые лишайники менее специфичны в своей субстратной селективности (Diploschistes muscorum (Scop.) R. Sant., D. scruposus (Schreb.) Norman и др.), однако, как и вышеуказанные виды, предпочитают хорошо прогреваемые и освещаемые местообитания.
Группа 2.1.9. Ареолированные. Характеризуются талломом, состоящим из обособленных ареол, находящихся на общем гипоталломе и имеющих покрытые коровым слоем края (некоторые представители рода Rhizocarpon Ramond ex DC., Lecidea fuscoatra (L.) Ach., Lecidella carpathica Körb., Trapelia involuta (Taylor) Hertel и др.).
Развитие корового слоя на краях ареол является результатом более высокой степени приспособленности лишайников к существованию на каменистом субстрате в переменных температурных условиях. Все лишайники Беларуси с ареолированным талломом являются облигатными эпилитными видами. Как и трещиновато-ареолированные виды, это редко встречающиеся в Беларуси лишайники, что связано, в первую очередь, с небольшой концентрацией валунного материала ледникового происхождения на значительной части территории страны (Golubkov, 1992, 1996; Tsurykau et al., 2018b).
Группа 2.1.10. Чешуйчато-ареолированные. Характеризуются талломом, состоящим из обособленных ареол (чешуек), прикрепленным к субстрату сужающимся основанием (вентрально) (некоторые виды рода Acarospora A. Massal., Buellia badia (Fr.) A. Massal., Caeruleum heppii (Nägeli ex Körb.) K. Knudsen et L. Arcadia и др.).
Появление края чешуек, обособленного от субстрата и покрытого коровым слоем, можно рассматривать в качестве важного шага в эволюции морфотипов лишайников, направленной на постепенный отрыв слоевища от субстрата произрастания (Elenkin, 1975; Golubkova, Byazrov, 1989) и формирование слоевищ более сложной анатомической структуры. Лишайники Беларуси с чешуйчато-ареолированным талломом произрастают на карбонатных, реже силикатных каменистых субстратах, единичные виды (Catapyrenium cinereum (Pers.) Körb., Endocarpon pusillum Hedw., Placidium squamulosum (Ach.) Breuss) колонизируют карбонатные хорошо прогреваемые почвы.
Группа 2.1.11. Чешуйчатые. Характеризуются талломом, состоящим из обособленных чешуек, прикрепленных к субстрату латерально (боковой стороной) (Carbonicola anthracophila (Nyl.) Bendiksby et Timdal, C. myrmecina (Ach.) Bendiksby etTimdal, Hypocenomyce scalaris (Ach. ex Lilj.) M. Choisy, Polycauliona ucrainica (S.Y. Kondr.) Frödén, Arup et Søchting, Xanthomendoza fulva (Hoffm.) Søchting, Kärnefelt et S.Y. Kondr., Xylopsora friesii (Ach.) Bendiksby et Timdal).
Чешуйчатые формы лишайников характеризуются гораздо большей степенью разобщения с субстратом произрастания по сравнению с чешуйчато-ареолированными за счет смещения области прикрепления с центрального на боковой участок таллома и одновременного уменьшения площади контакта. Вероятно, такая организация слоевища сформировалась в результате адаптации к произрастанию на вертикальных поверхностях (кора деревьев). Прикрепление чешуйки боковой стороной способствует сохранению ей горизонтальной ориентации в пространстве для более эффективного осуществления процесса фотосинтеза водорослевыми клетками. Все чешуйчатые виды лишайников Беларуси являются эпифитными видами, иногда заселяющими голую древесину. Некоторые лишайники (Polycauliona ucrainica, Xanthomendoza fulva) приурочены к антропогенным ландшафтам.
Группа 2.1.12. Плакодиоидные. Характеризуются талломом, имеющим трещиновато-ареолированную или ареолированную структуру в центральной части и лопастную по периферии (виды рода Calogaya Arup, Frödén et Søchting, Dimelaena oreina (Ach.) Norman, Dufourea ligulata (Körb.) Frödén, Arup et Søchting, Protoparmeliopsis muralis (Schreb.) M. Choisy, Variospora aurantia (Pers.) Arup, Søchting et Frödén).
По-видимому, развитие плакодиоидных талломов явилось результатом другого направления эволюции лишайников с ареолированными слоевищами, когда усложнение морфологической и анатомической структуры произошло не по краям каждой отдельной ареолы (путь развития ареолированные → чешуйчато-ареолированные → чешуйчатые), а по краям всего слоевища как единого целого. Вероятно, это направление эволюции явилось одним из магистральных путей развития лишайников на пути к формированию листоватых биоморф в результате развития нижнего корового слоя. Эта тенденция четко прослеживается в ряду некоторых представителей семейства Teloschistaceae Zahlbr.: Calo-gaya pusilla (A. Massal.) Arup, Frödén et Søchting (накипной плакодиоидный таллом без нижнего корового слоя, прикрепляющийся к субстрату нижней поверхностью) → Rusavskia elegans (Link) S.Y. Kondr. et Kärnefelt (листоватый таллом с нижним коровым слоем, прикрепляющийся к субстрату большей частью нижней поверхности) → Xanthoria parietina (L.) Th. Fr. (листоватый таллом с нижним коровым слоем, прикрепляющийся к субстрату отдельными участками нижней поверхности).
Плакодиоидные лишайники Беларуси являются ксерофитными эпилитными видами, заселяя в том числе бетонные и кирпичные постройки в крупных городах (Calogaya decipiens (Arnold) Arup, Frödén et Søchting, C. pusilla), часто доминируя в этих условиях над другими эпилитными видами.
Класс 2.2. Листоватые. Включает представителей с дорсовентральным строением таллома, прикрепленных к субстрату специальными органами прикрепления (гомфом, ризоидами, ризинами, гаптерами) или, реже, отдельными участками нижнего корового слоя.
Н.С. Голубкова выделила 3 класса плагиотропных лишайников – накипные, умбиликатные и листоватые (листовые) (Golubkova, 1983). Класс умбиликатных, в свою очередь, был разделен на 2 группы – умбиликатно-накипных и умбиликатно-листоватых жизненных форм. В Беларуси встречаются только представители с умбиликатно-листоватым типом таллома (виды родов Dermatocarpon Eschw. и Umbilicaria Hoffm.). Согласно данным анатомии, представители этих родов имеют идентичную со всеми остальными листоватыми лишайниками структуру (Krzewicka, 2004; Kummerová et al., 2006; Sonina et al., 2017) и их единственным отличием является наличие специфического органа прикрепления – гомфа. В представленной системе класс умбиликатных жизненных форм не выделяется и “умбиликатно-листоватые” виды рассматриваются в ранге группы умбиликатных класса листоватых лишайников, поскольку, на наш взгляд, наличие лишь специфического органа прикрепления при схожести всех остальных морфологических и анатомических признаков с таковыми класса листоватых лишайников недостаточно для выделения отдельной единицы высокого ранга.
Класс листоватых биоморф изначально был разделен на три группы: широколопастных ризоидальных, рассеченнолопастных ризоидальных и вздутолопастных неризоидальных жизненных форм (Golubkova, 1983). Такой подход не позволяет отразить все разнообразие морфологической структуры листоватых слоевищ и более четко обозначить экологические адаптации видов лишайников. Первые предложения по доработке системы листоватых жизненных форм были сделаны С.А. Пристяжнюком (Pristyazhnyuk, 1996а) (тем не менее, некоторые нововведения были подвергнуты критике (Muchnik, 2015)).
Одной из проблем вышеназванных систем является неопределенность понятий “широколопастные” и “узколопастные”, или “мелколопастные”. Например, согласно (Golubkova, 1983), представителями широколопастных ризоидальных лишайников являются виды родов Peltigera Willd., Lobaria (Schreb.) Hoffm. и Nephroma Ach., а к рассеченнолопастным ризоидальным относится род Cetrelia W.L. Culb. et C.F. Culb. Однако, ширина лопастей видов Peltigera didactyla (With.) J.R. Laundon, P. neckeri Hepp ex Müll. Arg., P. ponojensis Gyeln. и P. rufescens (Weiss) Humb. обычно не превышает 1 см (Vitikainen, 2007), в то время как ширина лопастей Cetrelia cetrarioides (Delise) W.L. Culb. et C.F. Culb., C. monachorum (Zahlbr.) W.L. Culb. et C.F. Culb. и C. olivetorum (Nyl.) W.L. Culb. et C.F. Culb. часто достигает 2 см (Bely et al., 2014b). Несколько иной подход к разграничению биоморф листоватых лишайников высказывается итальянскими лихенологами. Согласно (Nimis, Martellos, 2017), большинство пармелиоидных лишайников являются широколопастными, в то время как к узколопастным относятся фисциоидные и ксанториоидные лишайники.
Еще одной особенностью исходной системы биоморф является упоминание органов прикрепления талломов в названии групп. Например, лишайник Xanthoria parietina нельзя отнести ни к одной из групп системы Н.С. Голубковой, поскольку он является рассеченнолопастным неризоидальным.
В связи с вышеперечисленным, для наиболее полного охвата всех типов листоватых слоевищ лесной зоны (на примере Республики Беларусь) предлагается введение следующих групп.
Группа 2.2.1. Умбиликатные. Характеризуются моно-, реже полифильным талломом, прикрепленным к субстрату произрастания гомфом (Dermatocarpon miniatum (L.) W. Mann., Umbilicaria deusta (L.) Baumg.).
Представители умбиликатных жизненных форм являются исключительно эпилитными ксерическими видами (Golubkova, 1983). В Беларуси это крайне редкие лишайники, занесенные в Красную книгу (Krasnaya…, 2015).
Возможно, развитие умбиликатных жизненных форм связано с эволюцией ксерофитных эпилитных представителей с чешуйчато-ареолированным талломом, когда дальнейшее сужение основания чешуйки привело к формированию гомфа, а верхняя (плоская) поверхность увеличивалась в размерах. В пользу этого предположения свидетельствует филогенетическая близость многих родов c чешуйчато-ареолированными представителями и умбиликатных лишайников (роды Catapyrenium Flot., Endocarpon Hedw., Placidium A. Massal. и Dermatocarpon относятся к семейству Verrucariaceae Zenker).
Группа 2.2.2. Широколопастные. Характеризуются талломом с широкими, достигающими в ширину 10 мм и более, лопастями (представители родов Cetrelia, Lobaria, Nephroma, Parmotrema A. Massal., Peltigera, Punctelia Krog и др.).
Большинство широколопастных видов приурочены к старовозрастным лесам и встречаются на комлях и стволах старовозрастных деревьев, часто среди или поверх мхов, что положительно влияет на влагообеспеченность талломов (Bely et al., 2014a; Tsurykau et al., 2015). Эти виды обычно произрастают в пределах особо охраняемых природных территорий (Березинский биосферный заповедник, национальные парки “Беловежская пуща”, “Припятский”) и являются индикаторами старовозрастных лесов. Следует также отметить, что более половины широколопастных лишайников (18 видов, или 56% от числа видов группы) занесены в Красную книгу Республики Беларусь (I–IV категории, черный список, категория LC) или рекомендованы для включения в следующее ее издание (Cetrelia monachorum, C. olivetorum, Parmotrema perlatum (Huds.) M. Choisy, Punctelia jeckeri (Roum.) Kalb) (Krasnaya… 2015; Bely et al., 2014a; Tsurykau et al., 2015; Golubkov et al., 2017). Лишь немногие виды широколопастных лишайников являются распространенными в Беларуси. К таким видам можно отнести Peltigera didactyla (With.) J.R. Laundon, P. canina (L.) Willd., P. praetextata (Flörke ex Sommerf.) Zopf, P. rufescens (Weiss) Humb., Platismatia glauca (L.) W.L. Culb. et C.F. Culb. Однако следует отметить, что в южной части страны последний является достаточно редким видом и, учитывая современные тенденции климатических изменений и динамики видового состава лихенобиоты (Bertosh et al., 2014; Khomich et al., 2018; Tsurykau, 2019), этот бореальный лишайник является потенциально уязвимым.
Группа 2.2.3. Среднешироколопастные. Характеризуются талломом с лопастями средней ширины, обычно около 4–5 мм, редко шире, но не превышающими 10 мм (некоторые представители родов Melanelixia O. Blanco, A. Crespo, Divakar, Essl., D. Hawksw. et Lumbsch, Melanohalea O. Blanco, A. Crespo, Divakar, Essl., D. Hawksw. et Lumbsch, Nephromopsis Müll. Arg., Parmelia Ach., Parmelina Hale, Xanthoparmelia (Vain.) Hale и др.).
Большинство видов этой группы является распространенными лесными таксонами, предпочитающими мезофитные условия. Некоторые виды приурочены к переувлажненным местообитаниям и известны преимущественно из малонарушенных лесных сообществ (Hypotrachyna afrorevoluta (Krog et Swinscow) Krog et Swinscow, H. revoluta (Flörke) Hale). Единичные виды обитают на силикатных горных породах и предпочитают хорошо освещаемые ксерофильные условия произрастания (Xanthoparmelia conspersa (Ehrh. ex Ach.) Hale, X. pulla (Ach.) O. Blanco, A. Crespo, Elix, D. Hawksw. et Lumbsch и другие представители этого рода).
Группа 2.2.4. Узколопастные. Характеризуются талломом с узкими лопастями, обычно 1–2 мм, реже шире, но не превышающими 3 мм (представители родов Candelaria A. Massal., Parmeliopsis (Nyl.) Nyl., Phaeophyscia Moberg, Physcia (Schreb.) Michaux, Physconia Poelt, Scytinium (Ach.) Gray, Xanthomendoza S.Y. Kondr. et Kärnefelt, Xanthoria (Fr.) Th. Fr. и др.).
Большинство узколопастных лишайников приурочено к достаточно засушливым местообитаниям, одиночным деревьям, городским и урбанизированным промышленным территориям. Такие виды, как Phaeophyscia orbicularis (Neck.) Moberg, Physcia adscendens (Fr.) H. Olivier, P. dubia (Hoffm.) Lettau, P. stellaris (L.) Nyl., P. tenella (Scop.) DC., Polycauliona polycarpa (Hoffm.) Frödén, Arup, et Søchting, Xanthoria parietina составляют основное ядро лихенобиоты крупных городов (Tsurykau, 2013). Некоторые лишайники (Xanthomendoza coppinsii S.Y. Kondr. et Kärnefelt, X. huculica (S.Y. Kondr.) Diederich, X. ulophyllodes (Räsänen) Søchting, Kärnefelt et S.Y. Kondr.) встречаются исключительно на территории населенных пунктов, произрастая в парковых насаждениях или вдоль основных автомагистралей (Tsurykau, Kondratyuk, 2011; Tsurykau, 2013; Yatsyna, Kondratyuk, 2013; Golubkov, 2013). Узколопастные представители семейства Parmeliaceae Zenker произрастают преимущественно в светлых сосновых лесах (Cetraria sepincola (Ehrh.) Ach., Imshaugia aleurites (Ach.) S.F. Meyer, Parmeliopsis ambigua Nyl.) или заселяют типичные ксерические эпилитные субстраты (Arctoparmelia centrifuga (L.) Hale, Montanelia sorediata (Ach.) Divakar, A. Crespo, Wedin et Essl.). Лишь немногие узколопастные лишайники приурочены к хорошо увлажненным затененным мезофитным условиям (Heterodermia speciosa (Wulfen) Trevis., Parmeliopsis hyperopta (Ach.) Arnold).
Таким образом, в ряду широколопастные → → среднешироколопастные → узколопастные прослеживается плавное изменение экологической приуроченности видов лишайников. При сопоставлении максимальной ширины лопастей вида лишайника (согласно (Ahti et al., 1999, 2002, 2007, 2013; Nash et al., 2002, 2004, 2007; Smith et al., 2009; Thell, Moberg, 2011) с корректировкой на результаты собственных исследований) и степенью аридности его местообитаний (Nimis, Martellos, 2017) установлена четкая тенденция обратной связи между этими показателями (рис. 2). Данная тенденция подтверждает закономерности, установленные ранее для субарктических тундр (Pristyazhnyuk, 1996b): при увеличении сухости окружающей среды возрастает площадь контакта слоевища с воздушной средой за счет увеличения степени разветвленности и рассеченности таллома лишайника.
Рис. 2.
Связь между шириной лопастей листоватых лишайников Беларуси и степенью аридности их местообитаний.
Fig. 2. Relation between lobe width of foliose lichens of Belarus and aridity level of their habitats.
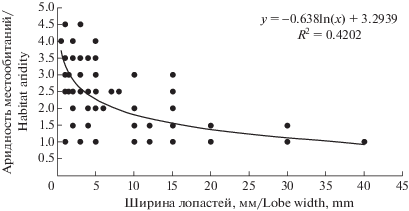
Группа 2.2.5. Вздутолопастные. Характеризуются талломом с узкими лопастями, имеющими воздушную полость внутри (виды рода Hypogymnia (Nyl.) Nyl., Menegazzia terebrata (Hoffm.) A. Massal.
Согласно (Kotlov, 1995; Pristyazhnyuk, 1996а) формирование группы вздутолопастных лишайников произошло в результате отдельного эволюционного пути развития слоевища. Разрыхление сердцевинного слоя и образование полости в талломе лишайника привело к увеличению отношения фотосинтезирующей поверхности к объему таллома. Вздутолопастные виды являются мезофитными лесными таксонами, некоторые представители включены в Красную книгу Республики Беларусь со статусом NT (Menegazzia terebrata) и DD (Hypogymnia farinacea Zopf, H. vittata (Ach.) Parrique).
Группа 2.2.6. Студенистые. Характеризуются гомеомерным талломом, имеющим в увлажненном состоянии вид студенистой листовидной массы, а в сухом виде становящимся очень тонким, хрупким (некоторые виды родов Blennothallia Trevis., Collema F.H. Wigg., Enchylium (Ach.) Gray, Lathagrium (Ach.) Gray, Leptogium (Ach.) Gray, Scytinium (Ach.) Gray).
Выделение студенистых (слизистых, желатинозных) жизненных форм является традиционным в лихенологии (Ahti et al., 2007; Smith et al., 2009; Tsurykau, Korchikov, 2018), хотя эта группа биоморф отсутствует в иерархических классификационных системах (Golubkova, 1983; Golubkova, Byazrov, 1989; Pristyazhnyuk, 1996а). Студенистые жизненные формы, как и вздутолопастные, четко отличаются от остальных групп листоватых лишайников своим анатомическим строением и узкой экологической селективностью. Подобно пленчатым накипным лишайникам, студенистые виды Беларуси приурочены к переувлажненным мезофитным, часто заболоченным местообитаниям, обычно поселяясь на покрытых мхом субстратах. Более трети видов студенистых лишайников Беларуси являются охраняемыми видами со статусом EN (Scytinium lichenoides (L.) Otálora, P.M. Jørg. et Wedin, S. subtile (Schrad.) Otálora, P.M. Jørg. et Wedin) или DD (Collema flaccidum (Ach.) Ach., Leptogium rivulare (Ach.) Mont., Leptogium cyanescens (Rabn.) Körb., Scytinium tenuissimum (Dickson) Otálora, P.M. Jørg. et Wedin). Помимо упомянутых видов, произрастание Leptogium saturninum (Dicks.) Nyl. было подтверждено (Yatsyna, 2017) спустя почти 100 лет после его последнего упоминания для территории республики (Błoński, 1889; Kreyer, 1913; Bachmann, Bachmann, 1920; Savicz, Savicz, 1924; Savicz, 1925). Среди всех студенистых лишайников, только Enchylium bachmanianum (Fink) Otálora, P.M. Jørg. et Wedin и E. tenax (Sw.) Gray встречаются в открытых хорошо прогреваемых местообитаниях, часто произрастая на городских территориях.
ТИП 3. Плагио-ортотропные. Представлены талломом, состоящим из горизонтальной (чешуйчатой или бугорчатой) и вертикально ориентированной в виде подециев или псеводоподециев частей.
Класс 3.1. Бородавчато- и чешуйчато-кустистые. Включает представителей с нередко исчезающим распростертым по субстрату плагиотропным накипным (зернисто-бородавчатым или чешуйчатым) и растущим перпендикулярно субстрату ортотропным кустистым слоевищем (подециями или псевдоподециями).
Традиционно класс подразделялся на группы шило- или сцифовидных, а также кустисто-разветвленных жизненных форм (Golubkova, 1983), однако позднее было предложено выделение четырех более дробных групп (Himelbrant, Kuznetsova, 2014; Muchnik, 2015), которые и приняты в настоящей классификационной системе.
Группа 3.1.1. Шиловидные. Характеризуются неразветвленными или слаборазветвленными подециями с заостренными окончаниями (Cladonia coniocraea (Flörke) Spreng., C. cornuta (L.) Hoffm., C. rei Schaer., C. subulata (L.) Weber ex F.H. Wigg. и др.).
Представители являются преимущественно мезофитными лесными таксонами. Некоторые лишайники приурочены к старовозрастным лесам, заселяя древесину или покрытые мхом субстраты (Cladonia cyanipes (Sommerf.) Nyl., C. norvegica Tønsberg et Holien, C. parasitica (Hoffm.) Hoffm.), однако большинство видов достаточно индифферентны к влажности и уровню освещенности и произрастают на опушках лесов и в открытых биотопах на сухой песчаной почве (Cladonia glauca Flörke, C. rei, C. subulata).
Группа 3.1.2. Палочковидные. Характеризуются неразветвленными или слаборазветвленными подециями с притупленными окончаниями (Cladonia botrytes (Hag.) Willd., C. cariosa (Ach.) Spreng., C. floerkeana (Fr.) Flörke, C. macilenta Hoffm., Pycnothelia papillaria Dufour и др.).
За исключением Cladonia incrassata Flörke, встречающейся исключительно в болотистых местообитаниях, все виды этой группы не требовательны к степени увлажнения среды обитания и встречаются как в лесных ценозах, так и на открытой местности.
Группа 3.1.3. Сцифовидные. Характеризуются неразветвленными или слаборазветвленными подециями с воронковидными окончаниями (Cladonia cenotea (Ach.) Schaer., C. chlorophaea (Flörke ex Sommerf.) Spreng., C. fimbriata (L.) Fr., C. verticillata (Hoffm.) Schaer. и др.).
Являясь преимущественно мезофитными видами, многие представители этой группы, тем не менее, предпочитают хорошо освещенные местообитания (Cladonia conista (Nyl.) Robbins, C. cryptochlorophaea Asahina, C. merochlorophaea Asahina, C. monomorpha Aptroot, Sipman et van Herk, C. novochlorophaea (Sipman) Brodo et Ahti и др.). Некоторые виды (Cladonia grayi G. Merr. ex Sandst.) проявляют очень широкую экологическую пластичность, одинаково часто встречаясь как в постоянно заболоченной местности, так и в сухих лишайниковых сосновых формациях (Tsurykau, Golubkov, 2015).
Группа 3.1.4. Кустисто-разветвленные. Характеризуются слабо- или сильноразветвленными подециями (Cladonia arbuscula (Wallr.) Flot., C. furcata (Huds.) Schrad., C. rangiformis Hoffm., C. uncialis (L.) F.H. Wigg., виды рода Stereocaulon Hoffm. и др.).
Большинство этих лесных видов предпочитают хорошо освещенные местообитания, встречаясь на полянах и опушках обычно сосновых светлых лесов, часто создавая сплошной лишайниковый покров на лесных песчаных дюнах (Gorbach, Kobzar, 1984). Некоторые виды (Cladonia portentosa (Dufour) Coem., C. stygia (Fr.) Ruoss) требовательны к влажности и произрастают в заболоченных открытых местообитаниях.
Таким образом, несмотря на то что большая часть бородавчато- и чешуйчато-кустистых лишайников является мезофитными лесными видами, и представители разных групп заселяют схожие по степени аридности местообитания (2.70–2.88 согласно (Nimis, Martellos, 2017)), в ряду шиловидные → палочковидные → сцифовидные → → кустисто-разветвленные прослеживается тенденция изменения экологической селективности видов в пользу предпочтения ими хорошо освещенных мест произрастания (рис. 3). Данная тенденция хорошо согласуется с положением о роли света как основного стимула биоморфологической эволюции лишайников (Golubkova, Byazrov, 1989; Kotlov, 1995; Muchnik, 2015). Как уже отмечалось ранее (Pristyazhnyuk, 1996б), формирование обильно разветвленных талломов способствует созданию особого микроклимата внутри лишайниковой дерновинки с целью снижения скорости испарения влаги с поверхности таллома и более эффективного осуществления процесса фотосинтеза фотобионтом.
Рис. 3.
Приуроченность жизненных форм бородавчато- и чешуйчато-кустистых лишайников к местообитаниям различного уровня освещенности.
Fig. 3. The association of verrucose- and squamulose-fruticose lichen life forms with habitats of different levels of solar irradiation.
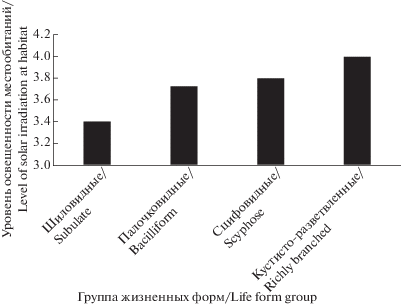
ТИП 4. Ортотропные. Слоевище ориентировано по отношению к субстрату вертикально.
Класс 4.1. Листоватые. Включает представителей с дорсовентральным строением таллома, имеющим один (дорсальный) водорослевый слой.
Группа 4.1.1. Субфрутикозные. Характеризуются талломом с очень узкими (до 0.3 мм) лопастями, образующим дерновинки небольшого размера, обычно до 1 см в диаметре (Phaeophyscia nigricans (Flörke) Moberg, Polycauliona candelaria (L.) Frödén, Arup et Søchting).
Субфрутикозные лишайники приурочены к засушливым, часто нитрофильным местообитаниям и являются постоянным компонентом лихенобиоты городов (Tsurykau, 2013).
Группа 4.1.2. Повисающие. Характеризуются талломом с разветвленными дорсовентральными лопастями, формирующими свисающее вдоль субстрата слоевище (Evernia prunastri (L.) Ach., Pseudevernia furfuracea (L.) Zopf).
Листоватые повисающие лишайники являются мезофитными лесными видами, произрастающими в хвойных, реже лиственных лесах.
Группа 4.1.3. Прямостоячие. Характеризуются талломом с разветвленными дорсовентральными лопастями, формирующими вертикально ориентированное относительно субстрата слоевище (Cetraria ericetorum Opiz, C. islandica (L.) Ach.).
Листоватые прямостоячие лишайники заселяют гораздо более сухие местообитания по сравнению с листоватыми повисающими лишайниками, в частности, песчаные почвы сухих типов сосновых лесов.
Класс 4.2. Кустистые. Включает представителей с лопастями радиальной структуры или реже уплощенными, но не дорсовентрального строения и в таком случае имеющими два водорослевых слоя.
Группа 4.2.1. Филаментозные. Характеризуются талломом, состоящим из очень тонких, нитевидных элементов, представляющих собой нити фотобионта, окруженные гифами микобионта (Reichlingia leopoldii Diederich et Scheid.).
Единственный представитель группы филаментозных жизненных форм, Reichlingia leopoldii, приурочен к мезофитным условиям старых парков, встречаясь на коре старых лиственных деревьев (Motiejūnaitė, 2009; Yatsyna, 2014).
Группа 4.2.2. Повисающие. Характеризуются талломом с радиальными или плоскими, но не дорсовентральными лопастями, формирующими свисающее вдоль субстрата слоевище (некоторые представители родов Bryoria Brodo et D. Hawksw., Evernia Ach., Ramalina Ach., Usnea Dill. ex Adans.).
Все повисающие кустистые лишайники являются эпифитными мезофитными кустистыми таксонами. Многие виды произрастают в достаточно хорошо освещенных условиях, обильно встречаясь в парковых посадках, дубравах (Evernia mesomorpha Nyl., Ramalina farinacea (L.) Ach., R. fraxinea (L.) Ach.), или заселяя верхние участки стволов деревьев (Usnea barbata (L.) Weber ex F.H. Wigg., U. dasopoga (Ach.) Nyl.).
Группа 4.2.3. Прямостоячие. Характеризуются талломом с радиальными или плоскими, но не дорсовентрального строения лопастями, формирующими вертикально ориентированное относительно субстрата слоевище (Bryoria furcellata (Fr.) Brodo et D. Hawksw., Cetraria aculeata (Schreb.) Fr., C. muricata (Ach.) Eckfeldt, большинство представителей родов Ramalina и Usnea).
Как и представители повисающих кустистых жизненных форм, прямостоячие эпифитные лишайники произрастают в хорошо увлажненных мезофитных лесных ценозах, однако оказываются более терпимыми к уровню инсоляции местообитаний (рис. 4). Исключением являются представители рода Cetraria, заселяющие ксерофитные песчаные склоны террас и эоловых речных дюн (Golubkov, 1992).
Рис. 4.
Приуроченность жизненных форм кустистых лишайников к местообитаниям различного уровня освещенности.
Fig. 4. The association of fruticose lichen life forms with habitats of different levels of solar irradiation.
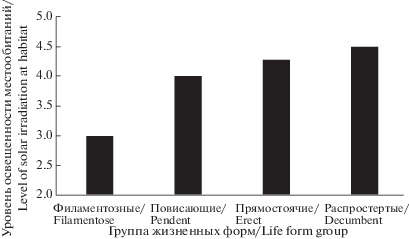
Группа 4.2.4. Распростертые. Характеризуются талломом, состоящим из округлых или уплощенных лопастей, формирующих распростертое по субстрату слоевище (Anaptychia ciliaris (L.) Körb.).
Единственный представитель распростертых жизненных форм, встречающийся в Беларуси, произрастает в хорошо освещаемых условиях в лесных фитоценозах и старых парках.
ЗАКЛЮЧЕНИЕ
Таким образом, в ряду групп каждого класса жизненных форм лишайников прослеживается тенденция изменения структуры таллома за счет его расчленения на более мелкие и/или короткие элементы (ареолы, чешуйки, лопасти) в связи с адаптацией к существованию в более освещенных и сухих условиях (рисунки 1–4). Более того, подобная тенденция может быть отмечена и для классов жизненных форм в целом (рис. 5) несмотря на то, что отдельные группы в пределах каждого класса обладают различной экологической приуроченностью.
Рис. 5.
Приуроченность представителей разных классов жизненных форм к местообитаниям различного уровня освещенности.
Fig. 5. The association of various classes of lichen life forms with habitats of different levels of solar irradiation.
На территории Республики Беларусь преобладают эвритопные виды лишайников (249; 42.7%), высока доля мезофитных лесных таксонов (244; 41.8%). Число лишайников, предпочитающих ксерофитные условия, относительно невелико (91; 15.5%). Они, как правило, приурочены к встречающимся достаточно редко каменистым субстратам ледникового происхождения или урбанизированным территориям. Такая структура лихенобиоты вполне закономерна для изучаемого региона, лесистость которого в настоящее время составляет 39.8% (Gosudarstvennyj… 2019).
Список литературы
Athi T., Jørgensen P.M., Kristinsson H., Moberg R., Søchting U., Thor G. 1999. Nordic lichens flora. Vol. 1. Introductory parts. Calicioid lichens and fungi. Uddevalla. 94 p.
Athi T., Jørgensen P.M., Kristinsson H., Moberg R., Søchting U., Thor G. 2002. Nordic lichens flora. Vol. 2. Physciaceae. Uddevalla. 115 p.
Athi T., Jørgensen P.M., Kristinsson H., Moberg R., Søchting U., Thor G. 2007. Nordic lichens flora. Vol. 3. Cyanolichens. Uddevalla. 219 p.
Athi T., Stenroos S., Moberg R. 2013. Nordic lichens flora. Vol. 4. Cladoniaceae. Göteborg. 117 p.
Bachmann E., Bachmann F. 1920. Litauische Flechten. – Hedwigia. 61 (6): 308–342.
Bely P. 2012. Absconditella lignicola (Stictidaceae) – lichen species new to Belarus. – Bot. Lith. 18 (2): 164–165. https://doi.org/10.2478/v10279-012-0018-2
Bely P., Golubkov V., Tsurykau A., Sidorovich E. 2014a. The lichen genus Cetrelia in Belarus: distribution, ecology and conservation – Bot. Lith. 20 (2): 69–76. https://doi.org/10.2478/botlit-2014-0010
[Bely et al.] Белый П.Н., Цуриков А.Г., Голубков В.В., Сидорович Е.А. 2014b. Новые данные о лишайниках рода Cetrelia (Lecanorales, Ascomycota) в Беларуси. – Доклады Национальной академии наук Беларуси. 58 (6): 83–88.
[Bertosh et al.] Бертош Е., Русаков Д., Лукашевич Т. 2014. Национальный доклад: Уязвимость и адаптация к изменению климата в Беларуси. Минск. 45 с.
Błoński F. 1889. Spis roślin zarodnikowych zebranych lub zanotowanych w lecie w r. 1888 w puszczach: Białowieskiéj, Świsłockiéj i Ladzkiéj. Pamiętnik fizyjograficzny. 9: 63–101.
Büdel B., Scheidegger C. 2008. Thallus morphology and anatomy. – In: Lichen Biology, 2nd ed. New York. P. 40–68. https://doi.org/10.1017/CBO9780511790478.005
[Elenkin] Еленкин А.А. 1975. Понятие лишайник и лишайниковый симбиоз. – Новости сист. низш. раст. 12: 1–81.
Frey E. 1923. Die Berücksichtigung der Lichenen in der soziologischen Pflanzengeographile, speziell in den Alpen. – Verb. Natf. Ges. Basel 35: 303–320.
[Golubkov] Голубков В.В. 1992. Лишайники охраняемых природных территорий Беларуси (флористическая и эколого-географическая характеристика): Дис. … канд. биол. наук. Минск. 503 с.
[Golubkov] Голубков В.В. 1996. Влияние антропогенной трансформации ландшафтов на особенности распространения и разнообразия лишайников в Белорусском Поозерье. – В сб.: Тез. докл. регион. науч.-практ. конф. “Сохранение биологического разнообразия Белорусского Поозерья”. Витебск. С. 81–82.
[Golubkov] Голубков В.В. 2013. Новые, редкие и малоизвестные таксоны лишайников, обнаруженные на территории Беларуси (краткое сообщение). – В сб.: Материалы IX междунар. науч.-практ. конф. “Актуальные проблемы экологии” Ч. 1. Гродно. С. 22–24.
[Golubkov et al.] Голубков В.В., Белый П.Н., Цуриков А.Г. 2017. Ревизия и анализ лишайников рода Punctelia (Parmeliaceae, lichenized Ascomycota) в Беларуси. – Ботаника (исследования). 76: 313–321.
[Golubkova] Голубкова Н.С. 1983. Анализ флоры лишайников Монголии. Л. 281 с.
[Golubkova, Byazrov] Голубкова Н.С., Бязров Л.Г. 1989. Жизненные формы лишайников и лихеносинузии. – Бот. журн. 74 (6): 794–805.
[Gorbach, Kobzar] Горбач Н.В., Кобзарь Н.Н. 1984. Формирование напочвенного лишайникового покрова в сосняках лишайниковых Белоруссии. – В сб.: Тез. докл. VII конф. по споровым растениям Средней Азии и Казахстана. Алма-Ата. С. 330–331.
[Gosudarstvennyj…] Государственный лесной кадастр Республики Беларусь по состоянию на 01.01.2019. 2019. Минск. 62 с.
Gueidan C., Villaseñor C.R., de Hoog G.S., Gorbu-shina A.A., Untereiner W.A., Lutzoni F. 2008. A rock-inhabiting ancestor for mutualistic and pathogen-rich fungal lineages. – Stud. Mycol. 61: 111–119. https://doi.org/10.3114/sim.2008.61.11
Hawksworth D.L. 1988. The variety of fungal-algal symbioses, their evolutionary significance, and the nature of lichens. – J. Linn. Soc. Bot. 96: 3–20. https://doi.org/10.1111/j.1095-8339.1988.tb00623.x
[Himelbrant, Kuznetsova] Гимельбрант Д.Е., Кузнецова Е.С. 2014. Таллом и репродуктивные структуры лишайников. – В кн.: Флора лишайников России: Биология, экология, разнообразие, распространение и методы изучения лишайников. М., СПб. С. 61–123.
Honegger R. 1991. Fungal evolution: Symbioses and morphogenesis. – In: Symbiosis, a Source of Evolutionary Innovation. Cambridge. P. 319–340.
[Khokhryakov] Хохряков А.П. 1981. Эволюция биоморф растений. М. 108 с.
[Khomich et al.] Хомич В.С., Логинов В.Ф., Мельник В.И., Табальчук Т.Г., Семенченко В.П., Кулак А.В., Степанович И.М. 2018. Признаки остепнения южной части Беларуси. – В сб.: Материалы VIII междунар. симпозиума “Степи Северной Евразии”. Оренбург. С. 1051–1054.
[Kotlov] Котлов Ю.В. 1995. О моделировании эволюции основных жизненных форм лишайников. – Бот. журн. 80 (3): 26–30.
[Krasnaya…] Красная книга Республики Беларусь: Редкие и находящиеся под угрозой исчезновения виды дикорастущих растений. 2015. Минск. 448 с.
[Kreyer] Крейер Г.К. 1913. К флоре лишайников Могилевской губернiи. Сборы 1908–1910 годов. – Труды Импер. Ботан. Сада. 31 (2): 263–440.
Krzewicka B. 2004. The lichen genera Lasallia and Umbilicaria in the Polish Tatra Mts. – Pol. Bot. Stud. 17: 1–88.
Kukwa M. 2006. The lichen genus Lepraria in Poland. – Lichenologist 38 (4): 293–305. https://doi.org/10.1017/S0024282906005962
Kummerová M., Barták M., Dubová J., Tríska J., Zubrová E., Zezulka S. 2006. Inhibitory effect of fluoranthene on photosynthetic processes in lichens detected by chlorophyll fluorescence. – Ecotoxicology. 15 (2): 121–131. https://doi.org/10.1007/s10646-005-0037-1
Launis A., Malíček J., Svensson M., Tsurykau A., Sérusiaux E., Myllys L. 2019. Sharpening species boundaries in the Micarea prasina group, with a new circumscription of the type species M. prasina. – Mycologia. 111 (4): 574–592. https://doi.org/10.1080/00275514.2019.1603044
Lendemer J.C. 2011. A standardized morphological terminology and descriptive scheme for Lepraria (Stereocaulaceae). – Lichenologist 43: 379–399. https://doi.org/10.1017/S0024282911000326
Lutzoni F., Pagel M., Reeb V. 2001. Major fungal lineages are derived from lichen symbiotic ancestors. – Nature. 41: 937–940. https://doi.org/10.1038/35082053
[Makarevicz] Макаревич М.Ф. 1977. Сем. Arthoniaceae. – В кн.: Определитель лишайников СССР. Вып. 4. Веррукариевые – Пилокарповые. Л. С. 279–290.
[Mirkin, Naumova] Миркин Б.М., Наумова Л.Г. 2014. Краткий энциклопедический словарь науки о растительности. Уфа. 288 с.
Motiejūnaitė J. 2006. Lichens of neglected habitats in eastern and eastern-central European lowlands. – Acta Mycol. 41 (1): 145–154. https://doi.org/10.5586/am.2006.018
Motiejūnaitė J. 2009. Lichens and allied fungi of two Regional Parks in Vilnius area (Lithuania). – Acta Mycol. 44 (2): 185–199. https://doi.org/10.5586/am.2009.017
[Muchnik] Мучник Е.Э. 2015. Жизненные формы лишайников: взгляды, системы, эволюция. – В сб.: Материалы VII Всерос. микол. школы-конф. “Биотические связи грибов: мосты между царствами”. М. С. 173–186.
Nash T.H.III, Gries C., Bungartz F. 2007. Lichen Flora of the Greater Sonoran Desert Region. Vol. 3. Tempe. 567 p.
Nash T.H.III, Ryan B.D., Diederich P., Gries C., Bungartz F. 2004. Lichen Flora of the Greater Sonoran Desert Region. Vol. 2. Tempe. 742 p.
Nash T.H.III, Ryan B.D., Gries C., Bungartz F. 2002. Lichen Flora of the Greater Sonoran Desert Region. Vol. 1. Tempe. 532 p.
Nimis P.L., Martellos S. 2017. ITALIC – The Information System on Italian Lichens. Version 5.0. http://dryades.units.it/italic.
Olszewska S., Zwolicki A., Kukwa M. 2014. Chemistry and morphology of Chrysothrix candelaris in Poland, with notes on the taxonomy of C. xanthina. – Mycotaxon 128: 165–172. https://doi.org/10.5248/128.165
Orange A. 2009. Thelidium A. Massal. (1855). – In: The lichens of Great Britain and Ireland. London. P. 879–883.
[Oxner] Окснер А.Н. 1974. Определитель лишайников СССР. Морфология, систематика и географическое распространение. Вып. 2. Л. 283 с.
[Pristyazhnyuk] Пристяжнюк С.А. 1996а. Жизненные формы лишайников субарктических тундр полуострова Ямал. I. Система жизненных форм. – Бот. журн. 81 (3): 34–41.
[Pristyazhnyuk] Пристяжнюк С.А. 1996b. Жизненные формы лишайников субарктических тундр полуострова Ямал. II. Связь с экологическими факторами. – Бот. журн. 81 (4): 48–55.
[Savicz] Савич В.П. 1925. Результаты лихенологических исследований 1923 года в Белоруссии. – Зап. Инст. С. Х. 4: 1–33.
[Savicz, Savicz] Савич В.П., Савич Л.И. 1924. Краткий предварительный отчет об исследовании флоры мхов и лишайников Белоруссии летом 1923 г. – Зап. Инст. С. Х. 3: 57–72.
[Serebryakov] Серебряков И.Г. 1962. Экологическая морфология растений. М. 378 с.
Smith C.W., Aptroot A., Coppins B.J., Fletcher A., Gilbert O.L., James P.W., Wolseley P.A. 2009. The lichens of Great Britain and Ireland. London. 1046 p.
Sonina A.V., Rumjantseva A.D. Tsunskaya A.A., Androsova, V.I. 2017. Adaptations of epilithic lichens to the microclimate conditions of the White Sea coast. – Czech Polar Rep. 7 (2): 133–143. https://doi.org/10.5817/CPR2017-2-13
Spribille T., Resl P., Ahti T., Pérez-Ortega S., Tønsberg T., Mayrhofer H., Lumbsch H.T. 2014. Molecular systematics of the wood-inhabiting, lichen-forming genus Xylographa (Baeomycetales, Ostropomycetidae) with eight new species. – Acta Univ. Ups. Symb. Bot. Ups. 37: 1–87.
Thell A., Moberg R. 2011. Nordic lichens flora. Vol. 4. Parmeliaceae. Göteborg. 184 p.
[Tsurykau] Цуриков А.Г. 2013. Лишайники юго-востока Беларуси. Гомель. 276 с.
Tsurykau A. 2018. A provisional checklist of the lichens of Belarus. – Opuscula Philolichenum. 17: 374–479.
[Tsurykau] Цуриков А.Г. 2019. Динамика географической структуры лихенобиоты Беларуси как индикатор современных биоклиматических условий. – Бот. журн. 104 (8): 1167–1188. https://doi.org/10.1134/S000681361908012X
Tsurykau A., Golubkov V. 2015. The lichens of the Cladonia pyxidata-chlorophaea complex in Belarus. – Folia Cryptog. Estonica. 52: 63–71. https://doi.org/10.12697/fce.2015.52.08
[Tsurykau, Khramchankova] Цуриков А.Г., Храмченкова О.М. 2015. Лишайники сосновой формации Гомельского района: видовой состав и особенности распространения. – В сб.: Материалы Междунар. науч.-практ. конф. “Геоботанические исследования естественных экосистем: проблемы и пути их решения”. Гомель. С. 159–163.
[Tsurykau, Kondratyuk] Цуриков А.Г., Кондратюк С.Я. 2011. Новые для Беларуси виды лишайников семейства Teloschistaceae. – Наука и инновации. 100 (6): 72.
[Tsurykau, Korchikov] Цуриков А.Г., Корчиков Е.С. 2018. Определитель лишайников Самарской области. Ч. 1. Листоватые, кустистые и слизистые виды. Самара. 128 с.
Tsurykau A., Golubkov V., Bely P. 2015. The genera Hypotrachyna, Parmotrema and Punctelia (Parmeliaceae, lichenized Ascomycota) in Belarus. – Herzogia. 28 (2): 736–745. https://doi.org/10.13158/heia.28.2.2015.736
Tsurykau A., Golubkov V., Bely P. 2016a. The genus Lepraria (Stereocaulaceae, lichenized Ascomycota) in Belarus. – Folia Cryptog. Estonica. 53: 43–50. https://doi.org/10.12697/fce.2016.53.06
[Tsurykau et al.] Цуриков А.Г., Голубков В.В., Белый П.Н. 2016b. Ревизия лишайников рода Lepraria в Беларуси: L. eburnea, L. ecorticata и L. vouauxii. – Вестник БГУ. Сер. 2. 2: 55–59.
[Tsurykau et al.] Цуриков А.Г., Голубков В.В., Белый П.Н. 2016c. Ревизия лишайников рода Lepraria в Беларуси: L. elobata и L. finkii. – Веснiк ВДУ. 2: 22–27.
[Tsurykau et al.] Цуриков А.Г., Голубков В.В., Белый П.Н. 2016d. Ревизия лишайников рода Lepraria в Беларуси: Lepraria incana. – Веснiк ГрДУ iмя Янкi Купалы. Серыя 5. 2: 133–140.
[Tsurykau et al.] Цуриков А.Г., Голубков В.В., Белый П.Н. 2016e. Ревизия лишайников рода Lepraria в Беларуси: L. jackii и L. Rigidula. – Веснiк Магiлеýскага дзяржаýнага ýнiверсiтэта iмя А.А. Куляшова. Серыя В. 1 (47): 91–97.
Tsurykau A., Golubkov V., Bely P. 2018a. The lichen genus Xanthoparmelia (Parmeliaceae) in Belarus. – Folia Cryptog. Estonica. 55: 125–132. https://doi.org/10.12697/fce.2018.55.13
[Tsurykau et al.] Цуриков А.Г., Голубков В.В., Белый П.Н. 2018b. Ревизия лишайников рода Xanthoparmelia Беларуси: X. delisei и X. pulla. – Журнал Белорусского государственного университета. Биология. 3: 21–27.
Vitikainen O. 2007. Peltigeraceae. – In: Nordic lichens flora. Vol. 3. Cyanolichens. Uddevalla. P. 113–131.
Wijayawardene N.N., Hyde K.D., Lumbsch H.T., Liu J.K., Maharachchikumbura S.S.N., Ekanayaka A.H., Tian Q., Phookamsak R. 2018. Outline of Ascomycota: 2017. – Fungal Divers. 88: 167–263. https://doi.org/10.1007/s13225-018-0394-8
Yatsyna A. 2014. Lichens from manor parks in Minsk region (Belarus). – Bot. Lith. 20 (2): 159–168. https://doi.org/10.2478/botlit-2014-0016
[Yatsyna] Яцына А.П. 2017. Лихенобиота спелых еловых насаждений двух особо охраняемых природных территорий Витебской области. – Веснік ВДУ. 3: 74–79.
[Yatsyna, Kondratyuk] Яцына А.П., Кондратюк С.Я. 2013. Новые данные о ксанториоидных лишайниках Беларуси. – Веснiк МДПУ iмя I. П. Шамякiна. 3: 29–33.
Zukal H. 1891. Halbflechten. – Flora (Regensburg). 74: 92–107.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал