

Ботанический журнал, 2020, T. 105, № 6, стр. 556-577
РАСПРЕДЕЛЕНИЕ РАСТИТЕЛЬНОСТИ В ДОЛИНЕ Р. БОЛЬШАЯ ХЭХЭГАНЪЯХА (БОЛЬШЕЗЕМЕЛЬСКАЯ ТУНДРА)
В. В. Нешатаев 1, *, И. А. Лавриненко 1, **
1 Ботанический институт им. В.Л. Комарова РАН
197376 Санкт-Петербург, ул. Проф. Попова, 2, Россия
* E-mail: xssa@mail.ru
** E-mail: lavrinenkoi@mail.ru
Поступила в редакцию 01.08.2019
После доработки 16.03.2020
Принята к публикации 17.03.2020
Аннотация
В долине р. Большая Хэхэганъяха, пересекающей возвышенность Вангуреймусюр в северо-западной части Большеземельской тундры, изучено распределение растительных сообществ на геоботанических профилях, заложенных на разнотипных участках – от бровки коренного склона до прирусловой части поймы. На 7-ми профилях выполнены геоботанические описания 49 сообществ, которые отнесены к 16 синтаксонам классификации Браун-Бланке ранга ассоциации и ниже. Ассоциация Salici hastatae–Caricetum stantis ass. nov. hoc. loco и вариант Hedysarum arcticum типовой субассоциации асс. Dryado octopetalae–Hylocomietum splendentis Andreyev ex Lavrinenko et Lavrinenko 2018 описаны впервые. Сообщества синтаксонов, которые сменяют друг друга в пределах эколого-динамических рядов долины, хорошо отражают экологические особенности местообитаний на геоботанических профилях.
В тундровой зоне речные долины относятся к элементам ландшафтной структуры со своеобразным микроклиматом и высоким уровнем биологического разнообразия растительного и животного мира, что связано с отепляющим эффектом водотоков, контрастно проявляющимся в Арктике. Долины являются транзитными элементами экологического каркаса (Stoyashcheva, 2007; Lavrinenko, Anufriev, 2009) и представляют собой “экологические коридоры”, обеспечивающие условия для расселения и миграции популяций и отдельных особей видов растений и животных (Pavleichik, 2009). По долинам рек на север продвигаются виды и сообщества растений с более южных районов, с ними связаны значительные флуктуации границы леса (Lavrinenko, Lavrinenko, 2004). Долины рек можно отнести к своеобразным резерватам, способствующим повышению видового и ценотического разнообразия.
Растительность речных долин можно представить, как совокупность обобщенных эколого-динамических рядов (Isachenko, 1967; Il’ina, 1968, 1991; Lipatova, 1971). Состав и структуру таких рядов определяют циклические флуктуационные изменения сообществ, связанные с колебаниями паводкового режима, сукцессионные смены сообществ и геоморфологические процессы, которые с разной скоростью и интенсивностью протекают на разных высотных уровнях долины. Мы придерживаемся мнения Б.М. Миркина, что в условиях динамичного ландшафта речной долины любой экологический ряд является фиксированным в пространстве динамическим рядом сукцессионной серии (Mirkin, 1971).
Своеобразие растительного покрова долин многих тундровых рек обусловлено почти повсеместным проявлением на склонах террас криогенных процессов (солифлюкция, пучение), которые протекают с разной интенсивностью, что определяется, прежде всего, крутизной склона, его экспозицией и подстилающим субстратом.
Важнейшей особенностью малых рек является тесная связь формирования стока с ландшафтом всего водосборного бассейна, что обусловливает их значительную уязвимость. Возрастание техногенной нагрузки при освоении территории (разработка и обустройство месторождений углеводородов, прокладка трубопроводов и дорожной сети, добыча твердых полезных ископаемых и т.п.) в пределах малого речного бассейна неизбежно приводит к трансформации режимов максимального и сезонного стоков (Makagonova, 2009), что ощутимо сказывается на экологическом состоянии малых рек. В связи с этим, изучение растительного покрова речных долин тундровой зоны в зависимости от геоморфологических особенностей и экологических факторов представляет особый интерес.
РАЙОН ИССЛЕДОВАНИЙ
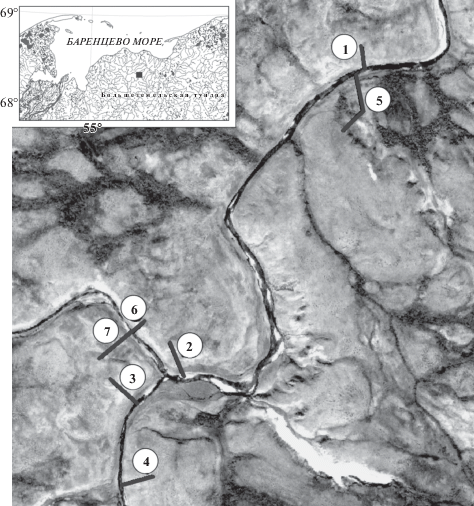
Исследования были проведены в июле–августе 2017 г. на возвышенности Вангуреймусюр в долине р. Большая Хэхэганъяха (с притоками), впадающей в р. Черную бассейна Баренцева моря на северо-западе Большеземельской тундры (рис. 1A). Территория расположена в подзоне северных гипоарктических (типичных) тундр и относится к Европейско-Западносибирской провинции, Канино-Печорской подпровинции, Западно-Большеземельскому геоботаническому округу, Вангурейскому району (Geobotanicheskoe…, 1989; Zony…, 1999a, b; Lavrinenko, 2013). На склонах террас, особенно в долинах водотоков, повсеместно развиты солифлюкционные процессы.
Рис. 1.
Район работ и расположение профилей в долине р. Большая Хэхэганъяха.
Fig. 1. Area of work and location of profiles in the Bolshaya Heheganyaha valley.
Зональным типом растительных сообществ в районе исследований являются пятнистые кустарничково (Dryas octopetala, Salix polaris, S. reticulata)-травяно (Carex bigelowii subsp. arctisibirica, Deschampsia glauca)-моховые (Aulacomnium turgidum, Hylocomium splendens, Ptilidium ciliare, Tomentypnum nitens) тундры, занимающие пологие склоны суглинистых холмов и отнесенные к асс. Dryado octopetalae–Hylocomietum splendentis Andreev ex Lavrinenko et Lavrinenko 2018 (Lavrinenko, Lavrinenko, 2018). На верхних частях склонов в долину и склонах сухих песчаных сопок сформированы ерниковые (Betula nana) кустарничково (Arctous alpina, Empetrum hermaphroditum, Vaccinium vitis-idaea, V. uliginosum subsp. microphyllum)-лишайниковые (Cladonia arbuscula, C. rangiferina, Flavocetraria nivalis, Sphaerophorus globosus) тундры (Andreyev, 1932) из союза Loiseleurio-Arctostaphylion Kalliola ex Nordhagen 1943. Для понижений на водоразделах обычны небольшие массивы деградирующих плоскобугристых торфяников с морошково (Rubus chamaemorus)-кустарничково (Ledum palustre subsp. decumbens, Empetrum hermaphroditum, Vaccinium vitis-idaea)-лишайниково (Cladonia arbuscula, C. rangiferina)-моховыми (Dicranum elongatum, Polytrichum strictum) сообществами асс. Rubo chamaemori–Dicranetum elongati Dedov ex Lavrinenko et Lavrinenko 2015 на буграх (Lavrinenko, Lavrinenko, 2015) и осоково-сфагновыми сообществами ассоциаций Carici rariflorae–Sphagnetum lindbergii Andreyev ex Lavrinenko, Matveyeva et Lavrinenko 2016 и Carici rariflorae–Sphagnetum baltici Andreyev ex Lavrinenko, Matveyeva et Lavrinenko 2016 в увлажненных и обводненных топях (Lavrinenko et al., 2016).
МАТЕРИАЛЫ И МЕТОДЫ
На берегах р. Большая Хэхэганъяха и ее безымянного притока было заложено 7 поперечных геоботанических профилей (рис. 1Б). Их закладывали на участках речных долин в пределах пешей доступности (8–10 км), учитывая геоморфологические особенности берегов, своеобразие микро- и нанорельефа, выраженность эколого-генетических зон поймы (прирусловой, центральной, притеррасной), разнообразие фитоценозов. По профилям от бровки склона коренной террасы до уреза воды (прирусловой части поймы) проводили геодезическую съемку. Геоботанические описания выполняли на площадках 5 × 5 м для травяных и кустарничковых сообществ (из-за малых размеров фитоценозов) и 10 × 10 м – для зарослей кустарников. Фитоценозы меньших размеров описывали в пределах естественных границ.
Для оценки почвенных условий, выявления признаков криогенеза и генетических типов отложений делали почвенные прикопки на глубину от 20 до 50 см. Всего было выполнено 49 описаний. Участие видов оценивали по шкале обилия–встречаемости Браун-Бланке: r – единично; + – менее 1%; 1 – 1–5%; 2а – 6–12%; 2b – 13–25%; 3 – 26–50%; 4 – 51–75%; 5 – 76–100%. Оценку проективного покрытия (%) давали в целом и по основным жизненным формам (кустарники, кустарнички, травы, мхи и лишайники). Обработку описаний выполняли с использованием программы “Excel”. В сводных таблицах для видов приведена константность, рассчитанная по шкале (%): I – 1–20, II – 21–40, III – 41–60, IV – 61–80, V – 81–100. Диагностическими видами синтаксона считали виды, которые отличают его от других синтаксонов в пределах всей выборки описаний. Номенклатура синтаксонов приведена в соответствии с “International code of phytosociological nomenclature” (ICPN) (Weber et al., 2000), таксонов сосудистых растений – со списком сосудистых растений Российской Арктики (Sekretareva, 2004), мхов – с перечнем мхов Российской Арктики (Afonina, Czernyadjeva, 1995), лишайников – со списком лихенофлоры России (A checklist…, 2010).
ГЕОМОРФОЛОГИЧЕСКИЕ ЭЛЕМЕНТЫ РЕЧНЫХ ДОЛИН
При описании растительности на профилях в долине р. Большая Хэхэганъяха (с притоками) мы используем следующие геоморфологические понятия и символы:
А. Коренной склон – это уступ, прилегающий к долине реки, и отделяющий ее от междуречий. Он представлен бровкой склона (А1) и его поверхностью (А2).
Бровка коренного склона – его резкий перелом, отделяющий вышележащую пологую часть или горизонтальную поверхность от нижележащего, более крутого участка склона. Бровка представляет собой верхний элемент речной долины, разделяющий собственно долину от местообитаний водораздела. В районе исследований на бровках зачастую отмечена дефляция песчаных отложений.
Поверхность коренного склона – наклонная поверхность пород, слагающих субгоризонтальные водораздельные территории. В зависимости от конкретных физико-географических условий склоны отличаются большим разнообразием.
В пределах обследованной территории в долине р. Большая Хэхэганъяха выявлено несколько типов коренных склонов разной крутизны с развитием различных эрозионных процессов. В фокус настоящей работы попали по большей части солифлюкционные склоны, на которых происходит постепенное массовое смещение чехла рыхлого материала.
В. Надпойменные террасы – естественные горизонтальные или слабонаклонные поверхности различного происхождения на склонах речных долин, ограниченные уступами. Террасы долины образуются как за счет врезания реки, так и за счет различных процессов денудации склонов: оползневых, морозного выветривания и солифлюкции.
Солифлюкционные террасы (В1) формируются вследствие значительного влияния на их генезис неравномерного течения насыщенного водой поверхностного слоя грунта в условиях близкого залегания многолетнемерзлых пород. В долине р. Большая Хэхэганъяха солифлюкционные террасы распространены повсеместно и через них был заложен ряд профилей.
Поверхность (В1.1) и склон (В1.2) таких террас обычно слабо дифференцированы. Если поверхность солифлюкционной террасы представлена почти горизонтальным участком, то при угле наклона более 2–3° она переходит в склон.
Солифлюкционный вал (В1.3) сложен рыхлыми породами, перемешанными с дерном, образуется чаще всего на пологих склонах (3–5°) у подножья террасы под влиянием солифлюкции, при участии морозного пучения. Протяженность вала может достигать нескольких сотен метров при ширине от 0.5 до 10.0 м и высоте до 1–2 м. В плане вал имеет дугообразные очертания.
С. Пойма, в пределах которой выделяли притеррасную (С1), центральную (С2) и прирусловую зоны. Последняя, в свою очередь, была подразделена на береговой вал (С3.1), прирусловый участок берега (С3.2) и островной участок или отмель (С3.3) (Kostenko, 1985).
На каждом из основных геоморфологических элементов профиля (А-В-С) встречались элементы нивального мерзлотного ландшафта (I), которые отличались длительным залеганием снежников и развитием процессов снежной эрозии (нивации).
К ним относятся нивальные ниши I(A-C)1, которые представляют собой незначительное углубление на склоне или у подножия возвышенности. И криопланационные террасы I(A-C)2 – выположенные поверхности с углами наклона 1–5°. Они имеют покров из щебнистых супесей и суглинков мощностью от нескольких десятков сантиметров до 1.5–2.0 м. Покров обычно разбит системой неявных полигонов различных размеров, часто сортированных, с разнообразными полигонально-жильными структурами (эпигенетическими повторно-жильными льдами, жилами, выполненными грубообломочным материалом и др.) (Kunitskiy, 2006).
СМЕНА РАСТИТЕЛЬНЫХ СООБЩЕСТВ НА ПРОФИЛЯХ
Профиль № 1. Левый пологий берег р. Большая Хэхэганъяха, склон юго-юго-восточной экспозиции. Длина профиля – 40 м, перепад высот – 0–7 м, геоморфологические элементы C – B1.3 – A2. Ряд сообществ: ивово-мохово-осоковое (Carex aquatillis subsp. stans, C. juncella, Philonotis tomentella, Salix hastata) → кустарничково-травяное (Poa arctica, Veratrum lobelianum, Salix hastata, S. reticulata) → мохово-кустарничково-разнотравное (Pachypleurum alpinum, Salix reticulata, Hylocomium splendens).
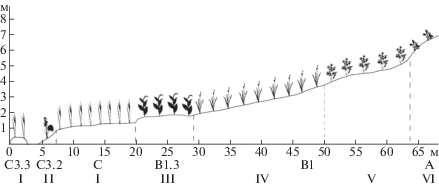
Профиль № 2. Левый берег р. Большая Хэхэганъяха, ниже по течению от места впадения безымянного притока, склон юго-восточно-восточной экспозиции. Длина профиля – 65 м, перепад высот – 0–7 м, геоморфологические элементы C3.3 – C3.2 – C – B1.3 – B1 – A (рис. 2). Ряд сообществ: ивово-мохово-осоковое на островной пойме (Carex aquatillis subsp. stans, Philonotis tomentella, Salix hastata) → белокопытниковое (Petasites radiatus) → ивово-мохово-осоковое (Carex aquatillis subsp. stans, Philonotis tomentella, Salix hastata) → ивняк мохово-разнотравный (Salix glauca, Poa palustris, Pyrola grandiflora, Hylocomium splendens) → осоково-кустарничково-моховое (Hylocomium splendens, Dryas octopetala, Carex bigelowii subsp. arctisibirica) → мохово-кустарничково-разнотравное (Pachypleurum alpinum, Salix reticulata, Hylocomium splendens) → травяно-кустарничковое (Salix nummularia, Luzula confusa).
Рис. 2.
Распределение синтаксонов на профиле № 2.
Здесь и на рис. 3–5:
по оси абсцисс – относительная длина профиля, по оси ординат – относительная высота профиля.
A – коренной склон; A1 – бровка склона; A2 – поверхность склона. B – надпойменные террасы; B1 – солифлюкционные террасы; B1.1 – поверхность террасы; B1.2 – склон террасы; B1.3 – солифлюкционный вал. C – пойма; C1 – притеррасная пойма; C2 – центральная пойма; C3 – прирусловая пойма; C3.1 – береговой вал; C3.2 – прирусловый участок (отмель); C3.3 – островной участок (отмель). I – элементы нивального мерзлотного ландшафта: A – прибровочные зоны; B – присклоновые зоны; C – притеррасные зоны; I(A-C)1 – нивальная ниша; I(A-C)2 – криопланационная терраса.
I – асс. Salici hastatae–Caricetum stantis фац. typicum; II – Petasites radiatus com.; III – Hylocomium splendens–Salix glauca com. type фац. Salix hastata; IV – асс. Dryado octopetalae–Hylocomietum splendentis субасс. typicum вар. Hedysarum arcticum фац. typicum; V – Hylocomium splendens–Pachypleurum alpinum com. type; VI – Luzula confusa–Salix nummularia com. type; VII – Silene acaulis–Salix reticulata com. type; VIII – Hylocomium splendens–Salix glauca com. type фац. typicum; IX – Empetrum hermaphroditum–Salix glauca com.; X – Hylocomium splendens–Salix glauca com. type фац. Carex juncella; XI – Sibbaldia procumbens–Veratrum lobelianum com. type; XII – Alchemilla murbeckiana com.
Fig. 2. Distribution of syntaxa on profile number 2.
Here and in fig. 3–5:
the abscissa is the relative length of the profile, the ordinate is the relative height of the profile.
A – root slope; A1 – edge of slope; A2 – the surface of the slope. B – terraces above floodplains; B1 – solifluction terraces; B1.1 – terrace surface; B1.2 – terrace slope; B1.3 – solifluction shaft. C – floodplain; C1 – inland floodplain; C2 – central floodplain; C3 – near-watercourse floodplain; C3.1 – coastal shaft; C3.2 – the near-bed area (shallow); C3.3 – island plot (shallow). I – elements of the nival permafrost landscape: A – near-edge zone; B – near-slope zones; C – near-terrace zones; I(A-C)1 – nival niche; I(A-C)2 – cryoplantation terrace.
I – ass. Salici hastatae–Caricetum stantis фац. typicum; II – Petasites radiatus com.; III – Hylocomium splendens–Salix glauca com. type fac. Salix hastata; IV – ass. Dryado octopetalae–Hylocomietum splendentis subass. typicum var. Hedysarum arcticum fac. typicum; V – Hylocomium splendens–Pachypleurum alpinum com. type; VI – Luzula confusa–Salix nummularia com. type; VII – Silene acaulis–Salix reticulata com. type; VIII – Hylocomium splendens–Salix glauca com. type fac. typicum; IX – Empetrum hermaphroditum–Salix glauca com.; X – Hylocomium splendens–Salix glauca com. type fac. Carex juncella; XI – Sibbaldia procumbens–Veratrum lobelianum com. type; XII – Alchemilla murbeckiana com.
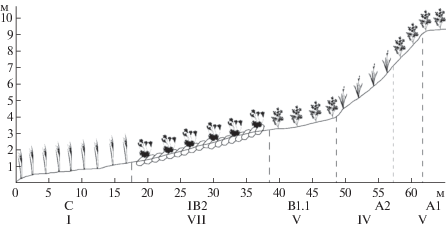
Профиль № 3. Левый берег р. Большая Хэхэганъяха, выше по течению от места впадения безымянного притока, склон юго-восточно-восточной экспозиции. Длина профиля – 60 м, перепад высот – 0.0–9.5 м, геоморфологические элементы C – IB2 – B1.1 – A2 – A1 (рис. 3). Ряд сообществ: ивово-мохово-осоковое (Carex aquatillis subsp. stans, Philonotis tomentella, Salix hastata) → ивово-травяно-моховое (Hylocomium splendens, Silene acaulis, Salix reticulata) → мохово-кустарничково-разнотравное (Pachypleurum alpinum, Salix reticulata, Hylocomium splendens) → осоково-кустарничково-моховое (Carex bigelowii subsp. arctisibirica, Salix reticulata, Hylocomium splendens) → мохово-кустарничково-разнотравное на бровке (Pachypleurum alpinum, Salix reticulata, Hylocomium splendens).
Рис. 3.
Распределение синтаксонов на профиле № 3.
Fig. 3. Distribution of syntaxa on profile number 3.
Профиль № 4. Правый крутой берег р. Большая Хэхэганъяха, склон западной экспозиции. Длина профиля – 45 м, перепад высот – 0–11 м, геоморфологические элементы C – B1.3 – A2 – A1. Ряд сообществ: ивово-мохово-осоковое (Carex aquatillis subsp. stans, Philonotis tomentella, Salix hastata) → мохово-кустарничково-разнотравное (Pachypleurum alpinum, Trollius europaeus, Salix reticulata, Hylocomium splendens) → мохово-кустарничково-разнотравное (Pachypleurum alpinum, Salix reticulata, Hylocomium splendens).
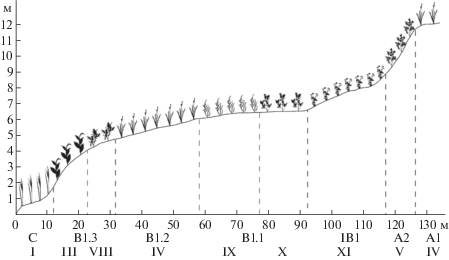
Профиль № 5. Правый берег р. Большая Хэхэганъяха, склон северной и северо-западной экспозиции. Проложен напротив профиля № 1. Длина профиля – 130 м, перепад высот – 0–12 м, геоморфологические элементы C – B1.3 – B1.2 – B1.1 – IB1 – A2 – A1 (рис. 4). Ряд сообществ: ивово-мохово-осоковое (Carex aquatillis subsp. stans, Philonotis tomentella, Salix hastata) → ивово-мохово-разнотравное (Poa palustris, Pyrola grandiflora, Hylocomium splendens, Salix glauca, S. hastata) → ивняк мохово-разнотравный (Salix glauca, Poa palustris, Pyrola grandiflora, Hylocomium splendens) → мохово-кустарничково-разнотравное (Pachypleurum alpinum, Salix reticulata, Hylocomium splendens) → ивняк кустарничково-моховый (Salix glauca, Hylocomium splendens, Empetrum hermaphroditum, Vaccinium vitis-idaea) → ивняк осоково-мохово-разнотравный (Salix glauca, Hylocomium splendens, Poa palustris, Pyrola grandiflora, Carex juncella) → нивальное разнотравно-ивково-моховое (Hylocomium splendens, Salix polaris, Sibbaldia procumbens, Veratrum lobelianum) → мохово-кустарничково-разнотравное (Pachypleurum alpinum, Salix reticulata, Hylocomium splendens).
Рис. 4.
Распределение синтаксонов на профиле № 5.
Fig. 4. Distribution of syntaxa on profile number 5.
Профиль № 6. Безымянный левый приток р. Большая Хэхэганъяха, крутой склон юго-западной экспозиции. Проложен перпендикулярно профилю № 2. Длина профиля – 20 м, перепад высот – 0–9 м, у русла обрыв около 1 метра выс., геоморфологические элементы A2 – A1 (рис. 5). Ряд сообществ: разнотравный луг (Alchemilla murbeckiana, Alopecurus pratensis subsp. alpestris, Achillea millefolium) → мохово-кустарничково-разнотравное (Pachypleurum alpinum, Salix reticulata, Hylocomium splendens) → травяно-кустарничковое (Salix nummularia, Luzula confusa).
Рис. 5.
Распределение синтаксонов на профиле № 6.
Fig. 5. Distribution of syntaxa on profile number 6.
Профиль № 7. Безымянный левый приток р. Большая Хэхэганъяха, склон северо-западной экспозиции. Проложен напротив профиля № 6, перпендикулярно профилю № 3. Длина профиля – 80 м, перепад высот – 0.0–8.5 м, геоморфологические элементы C – IB2 – IB1 – A2 – A1. Ряд сообществ: ивово-мохово-осоковое (Carex aquatillis subsp. stans, Philonotis tomentella, Salix hastata) → нивальные разнотравно-ивково-моховое (Hylocomium splendens, Salix polaris, Sibbaldia procumbens, Veratrum lobelianum) → ивово-осоково-хвощево-моховое (Philonotis tomentella, Equisetum arvense subsp. boreale, Carex aquatillis subsp. stans, Salix hastata) → нивальное разнотравно-ивково-моховое (Hylocomium splendens, Salix polaris, Sibbaldia procumbens, Veratrum lobelianum) → мохово-кустарничково-разнотравное (Pachypleurum alpinum, Salix reticulata, Hylocomium splendens).
ХАРАКТЕРИСТИКА СИНТАКСОНОВ
Растительность долины р. Большая Хэхэганъяха на основании различий во флористическом составе отнесена к 2 ассоциациям (включая 1 субассоциацию, 1 вариант и 5 фаций), 5 типам сообществ (с 3 фациями) и 4 сообществам (по 1 описанию).
Асс. Salici hastatae–Caricetum stantis ass. nov. hoc. loco (табл. 1; оп 1–11, номенклатурный тип (holotypus) – оп. 8 (возвышенность Вангуреймусюр, оп. 146, 21.07.2017, автор – В.В. Нешатаев); табл. 3, индекс Sh-Cs).
Таблица 1.
Ассоциация Salici hastatae–Caricetum stantis Table 1. Association Salici hastatae–Caricetum stantis
| Ассоциация/Association | Salici hastatae–Caricetum stantis | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Фация/Facies | typicum | C. j. | E. a. | ||||||||
| Площадь описания, м2/Relevé area, m2 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 |
| Проективное покрытие/Projective cover, %: | |||||||||||
| общее/total | 90 | 100 | 95 | 80 | 99 | 90 | 100 | 90 | 100 | 99 | 100 |
| кустарники/shrubs | <1 | <1 | <1 | <1 | <1 | <1 | 30 | <1 | 20 | 0 | <1 |
| кустарнички/dwarf shrubs* | 80 | 60 | 20 | 20 | 20 | 0 | 15 | 20 | 35 | 40 | 25 |
| травы/herbs | 30 | 50 | 80 | 60 | 90 | 80 | 75 | 65 | 55 | 70 | 45 |
| мохообразные/mosses | 10 | 60 | 70 | 55 | 60 | 60 | 20 | 40 | 30 | 50 | 70 |
| лишайники/lichens | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Число видов/Number of species: | |||||||||||
| общее/total | 21 | 24 | 15 | 17 | 16 | 16 | 19 | 28 | 24 | 22 | 9 |
| кустарников/shrubs | 4 | 3 | 4 | 4 | 4 | 1 | 4 | 4 | 5 | 2 | 0 |
| кустарничков/dwarf shrubs | 0 | 1 | 2 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 1 |
| трав/herbs | 16 | 20 | 8 | 12 | 10 | 14 | 15 | 23 | 18 | 18 | 7 |
| Cклон/Slope: | |||||||||||
| экспозиция/exposition | N | SEE | NE | NE | NE | W | SEE | SEE | SEE | S | NW |
| угол/angle | 0–3 | 0–3 | 0–3 | 0–3 | 0–3 | 2–7 | 0–2 | 0–20 | 0–2 | 5–10 | 2–5 |
| Дата/Date | 17.07.17 | 16.07.17 | 22.07.17 | 22.07.17 | 22.07.17 | 16.07.17 | 20.07.17 | 21.07.17 | 19.07.17 | 13.07.17 | 22.07.17 |
| Номер описания/Number of relevé: | |||||||||||
| авторский/by author | 134 | 131 | 153 | 151 | 152 | 128 | 143 | 146 | 142 | 118 | 154 |
| табличный/in table | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8** | 9 | 10 | 11 |
| Диагностические виды асс. Salici hastatae–Caricetum stantis и фац. typicum/Diagnostic species of ass. Salici hastatae–Caricetum stantis and fac. typicum | |||||||||||
| Carex aquatilis subsp. stans | 2a | 3 | 4 | 3 | 5 | 5 | 4 | 3 | 3 | + | 3 |
| Philonotis tomentella | 2a | 4 | 4 | 4 | 4 | 4 | 2b | 3 | 3 | 3 | 4 |
| Geum rivale | 1 | . | 1 | + | 1 | + | 1 | + | 1 | + | + |
| Equisetum variegatum | + | + | + | 1 | + | + | + | 1 | + | . | + |
| Caltha palustris | . | . | + | + | + | + | + | + | + | + | . |
| Comarum palustre | . | . | . | + | 1 | . | 2a | + | 2b | + | . |
| Salix lapponum | . | . | . | . | . | + | 1 | + | 1 | . | . |
| Диагностический вид асс. Salici hastatae–Caricetum stantis фац. Carex juncella/Diagnostic species of ass. Salici hastatae–Caricetum stantis fac. Carex juncella | |||||||||||
| Carex juncella | 1 | 2a | . | 2b | 1 | 1 | . | . | . | 3 | 1 |
| Диагностический вид асс. Salici hastatae–Caricetum stantis фац. Equisetum arvense/Diagnostic species of ass. Salici hastatae–Caricetum stantis fac. Equisetum arvense | |||||||||||
| Equisetum arvense | + | 1 | + | + | + | + | + | + | + | + | 2b |
| Константые виды/Constant species | |||||||||||
| Salix hastata | 5 | 2b | 2a | 2a | 2a | . | 2b | 2b | 2b | 3 | . |
| S. reticulata | . | 2b | 2a | . | 2a | . | . | + | 1 | + | 2b |
| S. glauca | + | . | + | + | + | . | 2a | . | 2a | + | . |
| Veratrum lobelianum | + | + | + | . | + | . | r | r | + | + | + |
| Bistorta vivipara | + | . | + | 1 | + | . | 1 | 1 | + | + | . |
| Trollius europaeus | + | + | . | + | . | . | + | + | r | 2a | . |
| Salix lanata | + | . | + | + | + | . | 2b | + | + | . | . |
| Ranunculus propinquus | + | + | . | . | r | . | . | + | + | . | . |
| Прочие виды/Other species | |||||||||||
| Viola biflora | . | . | . | . | . | . | + | . | + | + | . |
| Poa alpigena | . | + | . | . | . | . | + | + | + | . | . |
| Astragalus alpinus subsp. arcticus | . | + | . | . | . | . | . | + | . | + | . |
| Bistorta major | 2a | . | . | . | . | . | + | . | . | . | . |
| Equisetum scirpoides | + | + | . | . | . | . | . | . | + | + | . |
| Juncus arcticus | . | . | . | 1 | . | . | . | 1 | . | + | . |
| Salix phylicifolia | . | + | + | + | + | . | . | . | . | . | . |
| Hierochloё odorata | + | . | . | . | . | + | . | + | . | . | . |
| Salix myrsinites | + | + | . | . | . | . | . | . | + | . | . |
| Galium uliginosum | . | . | . | . | . | + | + | . | + | . | . |
| Veronica longifolia | . | r | . | . | . | . | . | + | + | . | . |
| Geranium albiflorum | + | . | . | . | . | + | . | + | . | . | . |
Примечание/Note. Виды, встреченные в 1–2 описаниях с обилием r, + или 1/Species found in 1–2 relevés with an abundance r, + or 1:
Achillea millefolium (8, 10), Alchemilla murbeckiana (8, 10), Alopecurus pratensis subsp. alpestris (3), Betula nana (8), Cardamine pratensis (6), Carex bigelowii subsp. arctisibirica (8, 9), C. capillaris (2), C. parallela subsp. redowskiana (2, 10), C. rariflora (2), Cerastium jenisejense (6), Chrysosplenium alternifolium (2), Conioselinum tataricum (2, 6), Eriophorum polystachion (7), Euphrasia frigida (8), Festuca rubra subsp. arctica (4, 8), Galium boreale (1), Hylocomium splendens (11), Lagotis glauca subps. minor (2), Luzula confusa (6), Parnassia palustris (8), Pedicularis compacta (9), Petasites frigidus (2), Pinguicula alpina (2, 9), Poa alpina (7), Ranunculus monophyllus (10, 11), Salix polaris (3), Saussurea alpina (1), Stellaria fennica (6), Tanacetum bipinnatum (4, 8), Thalictrum alpinum (2), Viola epipsila (10), V. epipsiloides (1).
GPS-координаты (WGS 84) (с.ш., в.д.)/GPS coordinates (WGS 84) (N, E): 1 – 68°21'20'', 56°29'31''; 2 – 68°20'58.6'', 56°28'46.7''; 3 – 68°21'3.5'', 56°28'45.8''; 4 – 68°21'3.2'', 56°28'47.3''; 5 – 68°21'3.1'', 56°28'46.9''; 6 – 68°20'52.2'', 56°28'45''; 7 – 68°20'59.1'', 56°28'47.8''; 8 – 68°21'0.1'', 56°28'56.2''; 9 – 68°20'58.4'', 56°28'46.9''; 10 – 68°21'20.1'', 56°29'32.2''; 11 – 68°21'2'', 56°28'44.1''.
C. j. – фац. Carex juncella/fac. Carex juncella, E. a. – фац. Equisetum arvense/fac. Equisetum arvense.
Ивово-мохово-осоковые сообщества сырых и увлажненных местообитаний.
Состав. Диагностические виды ассоциации и фац. typicum (индекс Sh-Cs-t): Caltha palustris, Carex aquatillis subsp. stans, C. juncella, Comarum palustre, Equisetum variegatum, Geum rivale, Philonotis tomentella, Salix hastata и S. lapponum. Число видов всего – 61, в сообществах варьирует от 9 до 28, в том числе кустарников – 0–5, кустарничков – 0–2, травянистых растений (трав) – 7–23. Высококонстантных 11 – 14 видов, или 23%; кроме диагностических это также Bistorta vivipara, Equisetum arvense, Salix glauca, S. lanata, S. reticulata, Trollius europaeus, Veratrum lobelianum.
Структура. Общее проективное покрытие (ОПП) – от 80 до 100%. Верхний ярус несомкнутый, 20–40 см выс. (некоторые кусты ив до 90 см выс.), образован содоминирующими травянистыми растениями (Carex aquatillis subsp. stans, Comarum palustre, Geum rivale, Carex juncella) с покрытием 30–90%, иногда и кустарниковыми ивами (Salix glauca, S. lanata), присутствующими единично или с покрытием до 30%. Во втором ярусе, 5–15 см выс., покрытие варьирует от 15 до 80% – преобладают кустарниковая (Salix hastata) и кустарничковая (S. reticulata) ивы. Напочвенный покров – моховой фрагментарный или сомкнутый (покрытие 10–70%), в основном из Philonotis tomentella.
Местообитания. Пойма, особенно ее прирусловая часть, иногда и наиболее пониженные участки на террасах. Структура фитоценоза определяется нанорельефом – сочетанием обводненных депрессий, поверхностным залеганием крупнообломочного материала. Для прирусловых участков характерно регулярное проточное увлажнение. На более высоких уровнях в течение вегетационного периода увлажнение изменяется от проточного (во время паводка) до застойного. В засушливые периоды экотопы могут пересыхать. Почвы аллювиальные серогумусовые и слоистые (примитивные).
Синтаксономия. Кроме типовой фации (табл. 1, оп. 1–9) выделены еще две – фац. Carex juncella (табл. 1, оп. 10; табл. 3, индекс Sh-Cs_cj), к которой отнесено эрозиофильное сообщество с доминированием одноименной осоки и незначительным участием Carex aquatillis subsp. stans, и фац. Equisetum arvense (табл. 1, оп. 11; табл. 3, индекс Sh-Cs_ea), сообщества которой занимают пониженные участки на криопланационных террасах.
Sibbaldia procumbens–Veratrum lobelianum com. type (табл. 3, индекс Sp-Vl).
Нивальные разнотравно-ивково-моховые сообщества.
Состав. Диагностические виды: Lagotis glauca subsp. minor, Oxyria digyna, Poa alpina, Pyrola minor, Salix polaris, Selaginella selaginoides, Sibbaldia procumbens, Tofieldia pusilla. Число видов всего – 58, в сообществах варьирует от 23 до 40, в том числе кустарников – 2–5, кустарничков – 1–2, травянистых растений – 19–32. Высококонстантных – 18 видов или 31% (Achillea millefolium, Anthoxanthum odoratum subsp. alpinum, Bistorta vivipara, Carex bigelowii subsp. arctisibirica, Equisetum arvense, Luzula multiflora subsp. frigida, Pachypleurum alpinum, Ranunculus propinquus, Salix glauca, S. hastata, S. reticulata, Tanacetum bipinnatum, Trollius europaeus, Vaccinium uliginosum subsp. microphyllum, Veratrum lobelianum).
Структура. ОПП – 80–100%. Травяно-кустарничковый ярус сомкнутый (от 60% до 85%, преобладают кустарнички Salix reticulata и S. polaris). В некоторых сообществах обильны травянистые растения Equisetum arvense, Ranunculus propinquus, Sibbaldia procumbens, Veratrum lobelianum. В напочвенном покрове преобладают мхи (40–70%). Доминанты мохового покрова – Hylocomium splendens и Sanionia uncinata. Лишайников значительно меньше (от менее 1% до 15%).
Местообитания. Пологие, слабовыпуклые солифлюкционные поверхности, вытянутые вдоль бортов долины (подножья бортов долины), с длительным залеганием снежного покрова, вследствие чего вегетационный период укорочен. Процессы нивации и солифлюкции сопутствуют образованию различных денудационных и аккумулятивных форм нанорельефа. Почвы – криоземы, петроземы.
Silene acaulis–Salix reticulata com. type (табл. 3, индекс Sa-Sr).
Ивково-травяно-моховые сообщества в местах выхода на поверхность крупнообломочного материала – сообщества “курумников” на криопланационной террасе.
Состав. Диагностические виды: Salix reticulata и Silene acaulis. Число видов в 2 описанных сообществах всего – 34 и 44, в том числе кустарников – 3 и 4, кустарничков – 2 и 5, трав – 27 и 33. Доминанты: Astragalus alpinus subsp. arcticus, Carex bigelowii subsp. arctisibirica, Equisetum arvense, Festuca ovina, Hylocomium splendens, Luzula multiflora subsp. frigida, Salix hastata, Saussurea alpina, Vaccinium uliginosum subsp. microphyllum.
Структура. Растительный покров (ОПП 90–95%) зачастую прерывается выходами крупнообломочного материала. Сомкнутый верхний ярус сложен кустарничками (50–60%) и травами (35–40%). Мохово-лишайниковый покров фрагментарный (покрытие мхов 25–50%, лишайников – до 15%). Присутствие видов Oxyria digyna, Poa alpina и Salix polaris сближает сообщества с нивальными, Astragalus alpinus subsp. arcticus, Potentilla crantzii – с луговыми, Carex bigelowii subsp. arctisibirica и Hylocomium splendens – с тундровыми зональными.
Местообитания. Солифлюкционный вал, солифлюкционные террасы. Пологие склоны с приповерхностным залеганием крупнообломочного материала, которое связано с оплыванием грунта, а также криогидрационным выветриванием пород из-за нивальных процессов. Особенности подстилающей поверхности приводят к наличию как гидрофильных, так и криоксерофильных экотопов. Почвы – криоземы, петроземы.
Poa arctica–Veratrum lobelianum com. (табл. 3, индекс Pa-Vl).
Кустарничково-травяное сообщество.
Состав. Диагностические виды: Poa arctica и Veratrum lobelianum. Число видов в 1 описании – 18, в том числе кустарников – 2, кустарничков – 2 и травянистых растений – 14. Среди доминантов наряду с диагностическими видами присутствуют Carex bigelowii subsp. arctisibirica, Salix hastata и S. reticulata.
Структура. Растительный покров сомкнутый (ОПП 90%). Покрытие травянистых растений выше (70%), чем кустарничков (20%). Напочвенный покров фрагментарный (20% мхов и менее 1% лишайников).
Местообитание. Солифлюкционный вал на границе с поймой. Выпуклый неявно бугорковый склон (3–5°). Почвы криометаморфические.
Асс. Dryado octopetalae–Hylocomietum splendentis Andreyev ex Lavrinenko et Lavrinenko 2018 субасс. typicum вар. Hedysarum arcticum var. nov. (табл. 2, оп. 1–15; табл. 3, индекс DH-Ha).
Таблица 2.
Ассоциация Dryado octopetalae–Hylocomietum splendentis субасс. typicum вар. Hedysarum arcticum Table 2. Association Dryado octopetalae–Hylocomietum splendentis subass. typicum var. Hedysarum arcticum
| Синтаксон / Syntaxa | Dryado octopetalae–Hylocomietum splendentis субасс. typicum вар. Hedysarum arcticum Dryado octopetalae–Hylocomietum splendentis subass. typicum var. Hedysarum arcticum | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Фация/Facies | typicum | Trollius europaeus | |||||||||||||
| Проективное покрытие Projective cover, %: | |||||||||||||||
| общее/total | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| кустарники/shrubs | <1 | <1 | 20 | 30 | <1 | <1 | 10 | 5 | 5 | <1 | <1 | <1 | 5 | 25 | <1 |
| кустарнички/dwarf shrubs | 40 | 65 | 60 | 60 | 50 | 40 | 60 | 65 | 50 | 50 | 40 | 50 | 60 | 70 | 40 |
| травы/herbs | 20 | 55 | 30 | 25 | 10 | 10 | 20 | 25 | 30 | 40 | 80 | 60 | 20 | 30 | 20 |
| мохообразные/mosses | 80 | 50 | 80 | 80 | 60 | 50 | 70 | 70 | 60 | 50 | 1 | 30 | 70 | 50 | 90 |
| лишайники/lichens | 10 | <1 | 0 | 20 | 15 | 10 | <1 | 15 | 1 | 1 | 0 | 0 | <1 | 0 | 0 |
| Число видов/Number of species: | 28 | 35 | 22 | 29 | 21 | 49 | 36 | 34 | 44 | 33 | 28 | 37 | 32 | 29 | 21 |
| общее/total | 17 | 23 | 16 | 19 | 12 | 24 | 24 | 23 | 25 | 23 | 23 | 31 | 25 | 21 | 15 |
| трав/herbs | 2 | 4 | 2 | 3 | 1 | 3 | 4 | 4 | 4 | 4 | 1 | 3 | 2 | 3 | 3 |
| кустарников/shrubs | 5 | 6 | 3 | 6 | 4 | 6 | 6 | 5 | 5 | 5 | 4 | 2 | 4 | 4 | 2 |
| Cклон/Slope: экспозиция/exposition | SSE | SEE | N | N | SSE | SSE | NEE | NE | SSW | W | SEE | W | W | W | W |
| угол/angle | 5–10 | 2–5 | 0–3 | 0–2 | 7–10 | 2–5 | 5–10 | 5–10 | 10–15 | 10–15 | 20 | 10–15 | 15–20 | 5–15 | 15–20 |
| Дата/Date | 07.07.17 | 20.07.17 | 17.07.17 | 18.07.17 | 07.07.17 | 07.07.17 | 17.07.17 | 22.07.17 | 13.07.17 | 14.07.17 | 11.07.17 | 16.07.17 | 14.07.17 | 14.07.17 | 14.07.17 |
| Номер описания Number of relevé: | |||||||||||||||
| авторский/by author | 104 | 144 | 136 | 137 | 105 | 106 | 132 | 149 | 119 | 120 | 115 | 127 | 121 | 122 | 123 |
| табличный/in table | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 |
| Диагностические виды асс. Dryado octopetalae–Hylocomietum splendentis/Diagnostic species of ass. Dryado octopetalae–Hylocomietum splendentis | |||||||||||||||
| Dryas octopetala | 1 | 2b | 3 | 2b | 1 | 2a | + | + | 2a | 2b | 3 | 2a | 2a | 1 | + |
| Salix reticulata | 2a | + | 2b | 2b | 2a | 2a | + | 2a | 2a | + | . | 2b | 2a | 1 | 2a |
| S. hastata | . | 1 | . | . | . | . | 2a | 2a | 2a | 1 | . | 2b | 2b | 3 | 2b |
| Equisetum scirpoides | + | + | . | . | + | + | . | + | + | + | . | + | + | + | 1 |
| Диагностические виды субасс. typicum/Diagnostic species of subass. typicum | |||||||||||||||
| Lagotis glauca subsp. minor | + | + | . | . | + | r | . | + | r | r | . | + | + | + | . |
| Thalictrum alpinum | + | + | + | . | . | r | . | + | . | . | + | + | . | + | + |
| Oxytropis sordida | + | . | . | . | + | + | r | . | 2a | + | 2a | . | + | . | . |
| Draba sibirica | . | . | . | . | . | . | + | . | + | + | + | . | + | + | + |
| Saussurea alpina | . | . | . | . | . | . | + | + | + | . | . | + | . | . | + |
| Astragalus frigidus | . | + | . | + | . | . | . | . | 1 | . | . | . | . | . | . |
| Диагностический вид вар. Hedysarum arcticum и фац. typicum/Diagnostic species of var. Hedysarum arcticum and fac. typicum | |||||||||||||||
| Hedysarum arcticum | . | 3 | 2b | . | + | + | + | . | 2a | 2b | . | 2a | 2a | 1 | + |
| Диагностические виды фац. Trollius europaeus/Diagnostic species of fac. Trollius europaeus | |||||||||||||||
| Trollius europaeus | + | . | . | . | . | . | . | . | . | . | + | 2b | + | 2b | + |
| Ranunculus monophyllus | . | r | . | . | . | . | . | . | + | + | . | + | + | 1 | + |
| R. propinquus | . | r | . | . | . | . | + | . | 1 | + | . | 2a | + | + | + |
| Характерные виды высших синтаксонов зональной растительности в восточноевропейских тундрах/Characteristic species of higher syntaxa of zonal vegetation in the East European tundra | |||||||||||||||
| Hylocomium splendens | 3 | 3 | 4 | 4 | 4 | 3 | 4 | 2a | 3 | 3 | + | 1 | 4 | 3 | 5 |
| Carex bigelowii subsp. arctisibirica | 1 | + | + | + | 2a | 2a | 2b | + | r | + | . | 2a | 1 | + | 2a |
| Salix glauca | + | + | 2a | 2a | . | + | 2a | + | 1 | + | . | + | 1 | 2b | + |
| Bistorta major | + | + | 1 | 2a | + | 1 | + | 1 | + | + | + | 2a | + | + | . |
| B. vivipara | + | + | 1 | 2a | + | + | + | + | + | + | + | 2a | + | + | + |
| Stellaria peduncularis | . | + | + | + | . | + | + | . | + | + | + | + | + | + | + |
| Poa arctica | + | + | + | + | . | r | + | + | + | + | . | . | + | . | . |
| Valeriana capitata | . | + | + | + | . | r | + | . | + | + | 2b | . | + | + | . |
| Pedicularis oederi | . | + | + | + | + | + | . | + | + | + | . | . | + | . | + |
| Aulacomnium turgidum | . | + | 2a | 1 | . | . | . | . | 2a | . | . | . | . | . | 1 |
| Константые виды/Constant species | |||||||||||||||
| Festuca ovina | 1 | 1 | + | 1 | 1 | + | + | 2a | + | + | 2b | . | + | . | . |
| Betula nana | + | + | 2b | 1 | + | + | + | 1 | + | + | + | . | . | + | . |
| Equisetum arvense | r | + | . | . | . | + | 1 | 1 | r | + | + | + | + | + | + |
| Vaccinium vitis-idaea | r | 1 | . | + | + | + | + | + | + | + | + | . | + | + | . |
| V. uliginosum subsp. microphyllum | . | 3 | . | 2a | . | 2a | 2b | 3 | 2b | 2b | 1 | . | 2b | 2b | . |
| Arctous alpina | + | 1 | + | 1 | . | + | + | + | + | + | 1 | . | . | . | . |
| Tephroseris integrifolia | . | + | + | r | . | . | + | + | + | + | . | + | + | . | . |
| Myosotis asiatica | . | . | . | + | . | . | + | . | + | + | + | + | + | + | . |
| Cerastium jenisejense | . | . | + | + | . | . | + | . | + | + | + | . | + | + | . |
| Silene acaulis | r | r | . | . | + | + | + | r | r | . | . | . | . | . | . |
| Astragalus alpinus subsp. arcticus | . | + | . | + | . | . | + | 1 | 2a | . | . | + | . | r | . |
| Luzula multiflora subsp. frigida | . | + | + | + | . | . | + | + | . | . | . | + | + | . | . |
| Poa alpigena | . | + | + | + | . | . | . | + | + | + | . | . | + | . | . |
| Прочие виды/Other species | |||||||||||||||
| Empetrum hermaphroditum | + | + | . | + | + | 1 | + | . | . | . | . | . | . | . | . |
| Carex vaginata subsp. quasivaginata | + | + | . | . | + | r | . | . | . | . | 2a | + | . | . | . |
| Pachypleurum alpinum | + | + | . | . | + | . | . | + | . | . | 2a | + | . | . | . |
| Dianthus superbus | . | . | . | . | . | . | + | . | r | . | + | + | + | + | . |
| Tanacetum bipinnatum | . | . | . | . | . | . | . | + | . | + | + | + | + | + | . |
| Salix lanata | . | + | . | 1 | . | . | + | + | . | . | . | + | . | . | + |
| Carex parallela subsp. redowskiana | + | . | . | . | + | + | . | . | . | . | . | + | . | . | . |
| Veratrum lobelianum | + | . | . | . | . | . | . | r | . | . | . | + | . | 1 | . |
| Potentilla crantzii | . | . | . | . | . | . | . | . | . | + | + | . | + | + | . |
| Tofieldia pusilla | . | r | . | . | . | r | + | + | . | . | . | . | . | . | . |
| Luzula confusa | . | . | . | . | . | . | . | . | + | + | 2b | . | . | . | . |
| Salix nummularia | . | . | . | . | . | + | . | . | + | + | . | . | . | . | . |
Примечание/Note. Виды, встреченные в 1–2 описаниях с обилием r, + или 1/Species found in 1–2 relevés with an abundance r, + or 1: Achillea millefolium (11, 12), Alchemilla murbeckiana (12), Alopecurus pratensis subsp. alpestris (12), Arctagrostis latifolia (6), Armeria maritima s. l. (6), Astragalus latifolia (4), Cardamine pratensis (12, 15), Cerastium regelii [incl. subsp. caespitosum] (4), Cortusa matthioli (11), Equisetum pratense (7), E. variegatum (6), Eutrema edwardsii (6), Festuca rubra subsp. arctica (4, 8), Gastrolychnis apetala (3), Geranium albiflorum (12), Geum rivale (12), Hierochloё alpina (11), H. odorata (11, 12), Luzula nivalis (3, 4), Parnassia palustris (9), Petasites frigidus (15), Pinguicula alpina (6, 8), Poa alpina (7), P. pratensis (7), Pyrola grandiflora (1, 6), Ranunculus repens (1), Rubus arcticus (8), Saxifraga hieracifolia (6), Solidago lapponica (11), Veronica longifolia (12), Viola biflora (7).
GPS-координаты (WGS 84) (с.ш., в.д.)/GPS coordinates (WGS 84) (N, E): 1 – 68°21'21.2'', 56°29'32''; 2 – 68°21'1.1'', 56°28'55.6''; 3 – 68°21'13.9'', 56°29'30.9''; 4 – 68°21'18.3'', 56°29'31.3''; 5 – 68°21'21.1'', 56°29'31.9''; 6 – 68°21'21.2'', 56°29'32''; 7 – 68°21'16.1'', 56°29'27.8''; 8 – 68°21'1.3'', 56°28'41.9''; 9 – 68°20'58.4'', 56°29'18.4''; 10 – 68°20'52.9'', 56°28'49.4''; 11 – 68°20'59.4'', 56°28'43.5''; 12 – 68°20'52.4'', 56°28'45.1''; 13 – 68°20'52.6'', 56°28'48.3''; 14 – 68°20'52.6'', 56°28'47.8''; 15 – 68°20'52.6'', 56°28'43.1''.
Таблица 3.
Синоптическая таблица синтаксонов долины р. Большая Хэхэганьяха Table 3. Synoptic table of syntaxa of the Bol’shaya Khekheganyakha valley
| Индекс синтаксона Syntaxon index | Sh-Cs_t | Sh-Cs_cj | Sh-Cs_ea | Sh-Cs | Sp-Vl | Sa-Sr | Pa-Vl | DH-Ha-te | DH-Ha-t | DH-Ha | Lc-Sn | Hs-Pa | Eh-Sg | Hs-Sg_t | Hs-Sg_cj | Hs-Sg_sh | Hs-Sg | Am |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Число видов/Number of species | 15–28 | 22 | 9 | 9–28 | 23–40 | 34–44 | 18 | 21–38 | 21–49 | 21–49 | 12–19 | 18–39 | 31 | 44 | 28 | 29–37 | 28–44 | 16 |
| Среднее число видов/Average number of species | 20 | 19 | 31 | 39 | 30 | 33 | 32 | 15 | 30 | 33 | 35 | |||||||
| Число описаний/Number of relevés | 9 | 1 | 1 | 11 | 5 | 2 | 1 | 4 | 11 | 15 | 2 | 7 | 1 | 1 | 1 | 2 | 4 | 1 |
| Диагностические виды асс. Salici hastatae–Caricetum stantis/Diagnostic species of ass. Salici hastatae–Caricetum stantis | ||||||||||||||||||
| Carex aquatilis subsp. stans | V3 | + | 3 | V3 | IIr | 2/+ | 2/+ | |||||||||||
| Philonotis tomentella | V3 | 3 | 4 | V3 | ||||||||||||||
| Geum rivale | V1 | + | + | V1 | II+ | 1/1 | I1 | + | 1/+ | 2/+ | + | |||||||
| Equisetum variegatum | V+ | + | V+ | 2/+ | + | I+ | I+ | |||||||||||
| Caltha palustris | IV+ | + | IV+ | |||||||||||||||
| Comarum palustre | III1 | + | III1 | |||||||||||||||
| Salix lapponum | III1 | II1 | I+ | |||||||||||||||
| Диагностические виды асс. S. h.–C. s. фац. Carex juncella/Diagnostic species of ass. S. h.–C. s. fac. Carex juncella | ||||||||||||||||||
| Carex juncella | III2a | 3 | 1 | IV2a | Ir | 4 | 1/4 | |||||||||||
| Диагностические виды Sibbaldia procumbens–Veratrum lobelianum com. type/Diagnostic species of Sibbaldia procumbens–Veratrum lobelianum com. type | ||||||||||||||||||
| Salix polaris | I+ | I+ | IV2a | 1/+ | ||||||||||||||
| Sibbaldia procumbens | IV1 | I+ | ||||||||||||||||
| Lagotis glauca subsp. minor | I+ | I+ | IV+ | 2/+ | 3/+ | IV+ | IV+ | 1/r | II+ | 1/+ | 1/r | |||||||
| Tofieldia pusilla | III+ | r | II+ | II+ | I+ | |||||||||||||
| Poa alpina | Ir | Ir | IIIr | 1/+ | I+ | I+ | I+ | |||||||||||
| Oxyria digyna | II+ | 1/+ | ||||||||||||||||
| Pyrola minor | II+ | |||||||||||||||||
| Selaginella selaginoides | II+ | 1/+ | 1/r | |||||||||||||||
| Диагностические виды Silene acaulis–Salix reticulata com. type/Diagnostic species of Silene acaulis–Salix reticulata com. type | ||||||||||||||||||
| Silene acaulis | 2/2a | IVr | IIIr | III+ | 1/+ | 1/r | ||||||||||||
| Диагностические виды асс. Dryado octopetalae–Hylocomietum splendentis субасс. typicum и вар. Hedysarum arcticum/Diagnostic species of ass. Dryado octopetalae–Hylocomietum splendentis subass. typicum and var. Hedysarum arcticum | ||||||||||||||||||
| Poa arctica | 2b | 1/+ | V+ | IV+ | 1/+ | II+ | + | + | 1/+ | |||||||||
| Hylocomium splendens | 1 | I1 | II3 | 2/2b | 4/3 | V3 | V3 | 1/2a | V2b | 5 | 3 | 2a | 1/2b | 3/2b | ||||
| Carex bigelowii subsp. arctisibirica | II+ | I+ | V+ | 2/2a | 2a | 4/1 | V1 | V1 | III2a | 2b | + | 1/+ | ||||||
| Dryas octopetala | 4/1 | V2a | V2a | 2/2a | I1 | |||||||||||||
| Hedysarum arcticum | I1 | 4/1 | IV2a | IV2a | 1/+ | I1 | 1/+ | 1/+ | ||||||||||
| Stellaria peduncularis | 2/+ | 4/+ | IV+ | IV+ | IIr | + | + | 1/+ | 2/+ | |||||||||
| Bistorta major | II1 | I1 | II1 | 1/+ | 3/1 | V+ | V1 | 1/+ | V+ | + | + | + | 1/+ | 3/+ | ||||
| Valeriana capitata | 2/+ | IV+ | IV+ | I+ | + | + | + | 2/+ | ||||||||||
| Pedicularis oederi | 2/+ | IV+ | IV+ | + | ||||||||||||||
| Oxytropis sordida | 1/+ | IV+ | III+ | II2a | + | 1/+ | ||||||||||||
| Draba sibirica | II+ | 3/+ | II+ | III+ | III+ | + | 1/+ | |||||||||||
| Aulacomnium turgidum | 1/1 | II1 | II1 | + | ||||||||||||||
| Диагностические виды Luzula confusa–Salix nummularia com. type/Diagnostic species of Luzula confusa–Salix nummularia com. type | ||||||||||||||||||
| Arctous alpina | 1/+ | V+ | IV+ | 2/2b | ||||||||||||||
| Salix nummularia | 1/+ | II+ | I+ | 2/2b | Ir | |||||||||||||
| Luzula confusa | I | I+ | II1 | I1 | 1/2a | II+ | ||||||||||||
| Empetrum hermaphroditum | + | III+ | II+ | 1/1 | 2b | |||||||||||||
| Диагностические виды Empetrum hermaphroditum–Salix glauca com./Diagnostic species of Empetrum hermaphroditum–Salix glauca com. | ||||||||||||||||||
| Vaccinium vitis-idaea | 2/+ | V+ | IV+ | 1/2a | V+ | 3 | + | 1/+ | ||||||||||
| Ledum palustre subsp. decumbens | + 2a |
|||||||||||||||||
| Sphagnum capillifolium | ||||||||||||||||||
| Диагностические виды Hylocomium splendens–Pachypleurum alpinum com. type/Diagnostic species of Hylocomium splendens–Pachypleurum alpinum com. type | ||||||||||||||||||
| Pachypleurum alpinum | IVr | 1/+ | 1/+ | III+ | II+ | V1 | + | 2/+ | 3/+ | 2a | ||||||||
| Viola biflora | II+ | + | II+ | III+ | 1/+ | I+ | I+ | 1/r | V1 | 2a | 2/1 | 3/1 | 1 | |||||
| Achillea millefolium | I+ | + | I+ | IV+ | 1/+ | 1/+ | I+ | I+ | IV+ | + | 2/+ | 3/+ | 2b | |||||
| Geranium albiflorum | II+ | II+ | II1 | + | 1/+ | I+ | IV1 | + | + | 1/+ | 3/+ | |||||||
| Potentilla crantzii | IIIr | 2/+ | 2/+ | I+ | II+ | III1 | + | 2/+ | 3/+ | |||||||||
| Carex vaginata subsp. quasivaginata | + | 1/+ | III+ | II+ | III1 | + | 1/+ | |||||||||||
| Диагностические виды Hylocomium splendens–Salix glauca com. type/Diagnostic species of Hylocomium splendens–Salix glauca com. type | ||||||||||||||||||
| Salix glauca | IV1 | + | IV1 | IV+ | 2/+ | 4/1 | V1 | V1 | IV+ | 3 | 3 | 5 | 2/+ | 4/2b | ||||
| S. lanata | IV+ | IV+ | III+ | 2/+ | 2/+ | II+ | II+ | II+ | 2b | 2b | 1 | 2/2a | 4/2a | |||||
| Sanionia uncinata | III2a | I2a | 1 | 1 | 1/2b | 3/2a | ||||||||||||
| Cortusa matthioli | I1 | I1 | I+ | + | + | 2/+ | ||||||||||||
| Pyrola grandiflora | II+ | 1/+ | I1 | I1 | + | + | 1 | 2/1 | ||||||||||
| Poa palustris | + | 1 | 2/1 | |||||||||||||||
| Диагностические виды Alchemilla murbeckiana com./Diagnostic species of Alchemilla murbeckiana com. | ||||||||||||||||||
| Alopecurus pratensis subsp. alpestris | I+ | I+ | II1 | 1/+ | 1/+ | I+ | III1 | 2a | ||||||||||
| Alchemilla murbeckiana | I+ | + | I+ | 1/+ | I+ | I+ | 1/+ | 1/+ | 2b | |||||||||
| Константные виды нескольких синтаксонов/Constant species of several syntaxa | ||||||||||||||||||
| Salix hastata | V2b | 3 | V2b | V+ | 2/1 | 2a | 4/2b | III2a | III2a | V1 | + | 2/2b | 2/2b | |||||
| S. reticulata | III2a | + | 2b | IV2a | V2a | 2/3 | 2a | 4/2a | V2a | V2a | IV2b | + | 2/2b | 2/2b | ||||
| Equisetum arvense | V+ | + | 2b | V+ | V1 | 2/1 | 1 | 4/+ | IV+ | IV+ | 2/+ | V+ | + | 2a | 1 | 2/1 | 4/1 | 2a |
| Bistorta vivipara | IV+ | + | IV+ | V+ | 2/+ | 4/1 | V+ | V+ | IV+ | + | + | r | 2/+ | 4/+ | 1 | |||
| Veratrum lobelianum | IV+ | + | + | V+ | V1 | 2/+ | 2a | 2/1 | I+ | II+ | V1 | + | + | 2/2a | 4/1 | + | ||
| Trollius europaeus | IV+ | 2a | IV+ | IV+ | 2/+ | 1 | 4/2a | I+ | II1 | IV1 | + | + | 2/1 | 4/1 | ||||
| Ranunculus propinquus | III+ | III+ | V1 | 2/+ | 4/1 | II+ | III+ | IV+ | + | + | 2/1 | 4/+ | 2a | |||||
| Equisetum scirpoides | II+ | + | II+ | III+ | + | 4/+ | IV+ | IV+ | 1/+ | IV+ | + | + | 2/+ | 3/+ | ||||
| Poa alpigena | III+ | II+ | II+ | 2/1 | 1/+ | III+ | III+ | I+ | + | + | 1/1 | 2/1 | ||||||
| Константные виды синтаксонов, за исключением пойменных сообществ/Constant species of syntaxa, excluding floodplain communities | ||||||||||||||||||
| Astragalus alpinus subsp. arcticus | II+ | + | II+ | II+ | 2/2a | + | 2/+ | III1 | III+ | 1/2a | III2a | 2/2a | 2/2a | 2a | ||||
| Betula nana | Ir | Ir | III+ | 1/r | + | 1/+ | V+ | IV+ | 1/2b | III+ | 2a | 1 | r | 1/+ | 3/+ | |||
| Festuca ovina | II1 | 2/1 | 1/+ | V1 | IV1 | 2/2a | IV1 | 1 | + | 1/1 | 2/1 | |||||||
| Vaccinium uliginosum subsp. microphyllum | IV+ | 2/1 | 2/2b | IV2b | IV2b | 1/2a | V2a | + | + | 1/+ | 2/+ | |||||||
| Tanacetum bipinnatum | II1 | I1 | IV+ | 2/+ | + | 3/+ | II+ | II+ | 2/+ | IV+ | + | 2/+ | 3/+ | |||||
| Luzula multiflora subsp. frigida | IV+ | 2/1 | + | 2/+ | III+ | III+ | 1/+ | IV+ | + | + | 2/+ | 3/+ | ||||||
| Tephroseris integrifolia | I+ | 2/+ | 2/+ | IV+ | III+ | 1/r | IV+ | 2/+ | 2/+ | r | ||||||||
| Thalictrum alpinum | I+ | I+ | I+ | 2/+ | + | 3/+ | III+ | III+ | III+ | + | 1/+ | 2/+ | ||||||
| Saussurea alpina | I+ | I+ | I+ | 2/1 | 2/+ | II+ | II+ | V+ | + | + | + | 1/+ | 3/+ | |||||
| Myosotis asiatica | I+ | 2/+ | 3/+ | III+ | III+ | III+ | + | 1/+ | 2/+ | + | ||||||||
| Dianthus superbus | Ir | 1/+ | 3/+ | II+ | II+ | 1/1 | III+ | 2/+ | 2/+ | |||||||||
| Cardamine pratensis | I+ | I+ | IIIr | 2/+ | 2/+ | I+ | I+ | + | + | 1/+ | 3/+ | |||||||
| Anthoxanthum odoratum subsp. alpinum | IV1 | IV2a | + | + | 1/2a | 3/1 | ||||||||||||
| Ranunculus monophyllus | + | + | I+ | 1/r | 4/+ | II+ | III+ | I+ | + | 1/+ | 2/+ | |||||||
| Carex parallela subsp. redowskiana | I1 | + | I1 | + | 1/+ | II+ | II+ | I2a | + | 1/+ | ||||||||
| Cerastium jenisejense | I+ | I+ | 2/+ | III+ | III+ | |||||||||||||
Осоково-кустарничково-моховые сообщества, производные (деривативные) от фитоценозов зональной асс. Dryado octopetalae–Hylocomietum splendentis Andreyev ex Lavrinenko et Lavrinenko 2018 субасс. typicum.
Состав. Постоянны или часто встречаются почти все диагностические виды ассоциации: Dryas octopetala, Equisetum scirpoides, Salix hastata, S. reticulata и типовой субассоциации: Astragalus frigidus, Draba sibirica, Lagotis glauca subsp. minor, Oxytropis sordida, Thalictrum alpinum, Saussurea alpina, а также характерные виды высших синтаксонов зональной растительности в восточноевропейских тундрах: Aulacomnium turgidum, Bistorta major, B. vivipara, Carex bigelowii subsp. arctisibirica, Hylocomium splendens, Pedicularis oederi, Poa arctica, Salix glauca, Stellaria peduncularis, Valeriana capitata. Вариант выделен на основе высокой константности Hedysarum arcticum (со средним обилием 2a), отсутствующего в типовой субассоциации. Число видов всего – 99, в сообществах варьирует от 21 до 37, в том числе кустарников – 1–4, кустарничков – 2–6, трав – 12–31. Среди содоминантов вместе с некоторыми диагностическими видами (Dryas octopetala, Hylocomium splendens, Carex bigelowii subsp. arctisibirica, Hedysarum arcticum) отмечены Salix hastata, S. reticulata и Vaccinium uliginosum subsp. microphyllum. Постоянно встречаются кустарнички Arctous alpina и Vaccinium vitis-idaea, но обилие их низкое.
Структура. Растительный покров сомкнут (ОПП 100%). Верхний ярус (проективное покрытие до 30%) составлен кустарниками (в основном Betula nana и Salix glauca). Травяно-кустарничковый ярус сплошной (проективное покрытие кустарничков варьирует от 40 до 70%, трав – от 10 до 80%). Напочвенный покров почти сомкнут (проективное покрытие мхов – до 90%, лишайников – до 20%).
Местообитания. Выпуклые склоны с выположенной или гофрировано-бугорковой (бугорки до 40 см выс.) поверхностью, структура которой зачастую обусловлена мерзлотными рельефообразующими процессами (полигонизация, морозобойные трещины).
Синтаксономия. Из-за различий в составе сообществ и их положения на разных элементах долины в варианте выделено 2 фации: typicum и Trollius europaeus.
Фация typicum (табл. 2, оп. 1–11; табл. 3, индекс DH-Ha-t).
Кустарничково-осоково-моховые тундры на солифлюкционных террасах, бровках и пологих склонах коренных террас.
Состав и структура, как в ассоциации.
Местообитания. Преимущественно на бровке коренных склонов, на границе с сообществами водоразделов. Участки хорошо дренированные, при наличии мерзлотных форм рельефа в пределах фитоценоза формируется мозаичность (кустарники зачастую приурочены к морозобойным трещинам). Почвы – криометаморфические.
Фация Trollius europaeus (табл. 2, оп. 12–15; табл. 3, индекс DH-Ha-te).
Олуговелые кустарничково-осоково-моховые тундровые сообщества на крутых склонах коренных террас.
Состав и структура. Сообщества отличаются от таковых типового варианта более высоким участием в покрове мезофитных травянистых растений – Ranunculus monophyllus, R. propinquus, Trollius europaeus, Veratrum lobelianum (покрытие травянистых растений – от 20 до 60%) и отсутствием некоторых кустарничков (Arctous alpina, Empetrum hermaphroditum) и ксерофитных видов травянистых растений (Festuca ovina, Silene acaulis).
Местообитания. Обычно в верхней трети коренных склонов разной экспозиции, поверхностный сток и защищенность бортами долины благоприятствуют высокой доле мезофитных растений. Почвы – криоземы.
Luzula confusa–Salix nummularia com. type (табл. 3, индекс Lc-Sn)
Травяно-кустарничковые сообщества на песчаных хорошо дренированных бровках и верхних частях коренных склонов долин.
Состав. Диагностические виды: Arctous alpina, Empetrum hermaphroditum, Luzula confusa, Salix nummularia, Vaccinium vitis-idaea. Число видов всего – 25, в 2 сообществах 12 и 19, в том числе кустарников 1 и 2, кустарничков – 2 и 5, травянистых растений – 9 и 11. Среди содоминантов, кроме диагностических видов, также Astragalus alpinus subsp. arcticus, Betula nana, Dryas octopetala, Festuca ovina, Vaccinium uliginosum subsp. microphyllum.
Структура. Растительный покров (ОПП 75 и 100%) иногда прерывается дефляционными песчаными участками с аккумулированной на поверхности галькой. Травяно-кустарничковый ярус хорошо выражен, но фрагментарен (проективное покрытие кустарничков – 45 и 55%, травянистых растений – 15 и 20%), напочвенный покров разрежен (проективное покрытие мхов – 10 и 20%, лишайников – 5 и 20%). В одном сообществе отмечена Betula nana (15%).
Местообитания. Ветрообдуваемые, хорошо дренированные, сухие летом и с неглубоким снежным покровом зимой, участки преимущественно на бровках и в верхних частях коренных склонов долин. Почвы – слабо развитые псаммоземы и подбуры тундровые.
Hylocomium splendens–Pachypleurum alpinum com. type (табл. 3, индекс Hs-Pa)
Склоновые мохово-кустарничково-разнотравные сообщества.
Состав. Диагностические виды: Achillea millefolium, Carex vaginata subsp. quasivaginata, Geranium albiflorum, Pachypleurum alpinum, Potentilla crantzii, Viola biflora. Число видов всего – 68, в сообществах варьирует от 18 до 39, в том числе кустарников – 1–4, кустарничков – 2–3, травянистых растений – 11–30. Высококонстантны 17 видов травянистых растений, кроме некоторых диагностических (Geranium albiflorum, Pachypleurum alpinum, Viola biflora) – также Anthoxanthum odoratum subsp. alpinum, Astragalus alpinus subsp. arcticus, Bistorta major, B. vivipara, Equisetum arvense, E. scirpoides, Festuca ovina, Luzula multiflora subsp. frigida, Ranunculus propinquus, Saussurea alpina, Tanacetum bipinnatum, Tephroseris integrifolia, Trollius europaeus, Veratrum lobelianum); явных доминантов нет. Постоянны кустарники – Salix hastata, S. glauca и кустарнички – Salix reticulata, Vaccinium uliginosum subsp. microphyllum и V. vitis-idaea, в напочвенном покрове – мох Hylocomium splendens.
Структура. Растительный покров сомкнут (ОПП 99–100%). В травяно-кустарничковом ярусе доминируют растения той или иной жизненной формы (проективное покрытие травянистых растений варьирует от 15 до 80%, кустарничков – от 5 до 60%). В напочвенном покрове преобладают мхи (от 20 до 60%), лишайники отсутствуют или их мало, редко до 30%. Кустарники единичны или редко имеют проективное покрытие до 25%.
Местообитания. Бровки и верхние части коренных склонов (5–45° ) долины. Часто сообщества постепенно переходят в прилегающие тундровые на водоразделе. Поверхность слабо бугристая. Неровности, связанные с действием мерзлотных и склоновых процессов, создают разнообразие экологических условий на уровне нанорельефа. Почвы – подбуры тундровые, криометаморфические.
Alchemilla murbeckiana com. (табл. 3, индекс Am)
Хионофитный склоновый разнотравный луг.
Состав. Диагностические виды: Alchemilla murbeckiana и Alopecurus pratensis subsp. alpestris. Описано одно сообщество, в котором зарегистрировано 16 видов. Наряду с диагностическими наиболее обильны Achillea millefolium, Astragalus alpinus subsp. arcticus, Equisetum arvense, Pachypleurum alpinum, Ranunculus propinquus.
Структура. Проективное покрытие травянистых растений – 100%. Напочвенный моховой покров разрежен (5%).
Местообитания. Крутые (10–30° ) склоны южной экспозиции, где снег не тает до середины июля. Глубокий снежный покров способствует слабому промерзанию склонов, которые по мере таяния снега хорошо обеспечены влагой. В вегетационный период местообитания теплые, поскольку поверхность склонов находится под сильным радиационным воздействием. Всё это создает благоприятные условия для развития мезофитного разнотравья. Почвы серогумусовые.
Hylocomium splendens–Salix glauca com. type (табл. 3, индекс Hs-Sg)
Ивняки мохово-разнотравные.
Состав. Диагностические виды: Cortusa matthioli, Poa palustris, Pyrola grandiflora, Salix glauca, S. lanata, Sanionia uncinata. Число видов всего – 66, в сообществах варьирует от 28 до 44, в том числе кустарников – 3–4, травянистых растений – 22–34. Доминанты: Equisetum arvense, Hylocomium splendens, Salix glauca, S. lanata.
Структура. Покров сомкнут (ОПП 100%). Кустарниковые ивы (Salix glauca, S. lanata), 1.0–1.3 м выс., имеют проективное покрытие 70–90%. Травяной ярус с покрытием 40–60% делится на 2 подъяруса: верхний (50 см выс.) сложен Bistorta major, Cortusa matthioli, Equisetum arvense, Poa palustris, Trollius europaeus, нижний (15–20 см выс.) – Adoxa moschatellina, Myosotis palustris, Viola biflora. Микрорельеф кочковатый. Напочвенный покров моховой (Climacium dendroides, Hylocomium splendens, Sanionia uncinata, Rhytidiadelphus triquetrus), фрагментарный (30–50%). Кустарничков и лишайников менее 1%.
Синтаксономия. Кроме типовой (фац. typicum, индекс Hs-Sg_t) выделены фац. Carex juncella (индекс Hs-Sg_cj) и фац. Salix hastata (индекс Hs-Sg_sh).
Местообитания. На разных участках долины (пойма и надпойменные террасы) в местах с повышенным увлажнением, связанным с временными водотоками, эрозионно-криогенными депрессиями, стоком со снежников. Почвы дерново-подбуры тундровые. Сообщества фац. H. s.–S. g. typicum описаны на бровке солифлюкционной террасы, фац. H. s.–S. g. Carex juncella – на присклоновой депрессии на террасе, фац. H. s.–S. g. Salix hastata – на солифлюкционном валу и склонах солифлюкционных террас на границе с поймой. Экотопы – от мезофильных в верхних и средних частях склонов до гигрофильных в нижних. Почвы криометаморфические глеевые, криоземы.
Empetrum hermaphroditum–Salix glauca com. (табл. 3, индекс Eh-Sg)
Ивняк кустарничково-моховой.
Состав. Диагностические виды, отличающие сообщество от других ивняков: Empetrum hermaphroditum, Ledum palustre subsp. decumbens, Vaccinium vitis-idaea. В одном описанном сообществе всего 30 видов, в том числе кустарников – 4, кустарничков – 4, травянистых растений – 21. Высоко обилие кустарниковых ив Salix glauca, S. lanata, кустарничков Empetrum hermaphroditum и Vaccinium vitis-idaea, осоки Carex bigelowii subsp. arctisibirica и мхов Hylocomium splendens и Sphagnum capillifolium.
Структура. Покров сомкнут (ОПП 100%). Проективное покрытие кустарников (кроме ив есть ерник Betula nana) – 60%, под ними сомкнутые: травяно-кустарничковый (кустарничков – 80%, травянистых растений – 20%) ярус и лишайнико-моховой (мхов – 80%, лишайников – 5%) покров.
Местообитание. Пологий участок присклоновой части солифлюкционной террасы с поверхностным увлажнением. Мелкобугристый полигонизированный микрорельеф. Хорошее увлажнение из-за стока воды с находящихся выше склонов и снежников благоприятствует развитию кустарниковой растительности. Почвы торфяно-глееземы глееватые.
Petasites radiatus com. (индекс Pr)
Белокопытник лучистый формирует разреженные сообщества или сомкнутые (до 70–90%) преимущественно моновидовые заросли на мелководьях по берегам рек, на перекатах, на песчаном или гравийном речном аллювии.
Состав. Petasites radiatus, являясь диагностическим видом синтаксона, формирует моновидовые заросли.
Местообитание. Прирусловая часть поймы, также на мелководьях, на перекатах, на песчаном или гравийном речном аллювии.
ОБСУЖДЕНИЕ
Условный полный эколого-динамический ряд растительных сообществ речной долины включает в себя фитоценозы разныx синтаксонов, которые были встречены в пределах обследованной территории (табл. 3). Распределение сообществ синтаксонов на геоморфологических элементах долины р. Большая Хэхэганъяха хорошо отражает матрица, в которой строки – синтаксоны, столбцы – геоморфологические элементы (табл. 4). Она показывает, что иногда сообщества, отнесенные к одному синтаксону, могут занимать несколько местообитаний на профиле (сходных по экологическим условиям, например, увлажнению) и наоборот, один и тот же геоморфологический элемент на разных профилях, в зависимости от почвенных условий или экспозиции склона, может быть занят сообществами разных синтаксонов.
Таблица 4.
Распределение синтаксонов по элементам долины Table 4. Distribution of syntaxa among the valley’s elements
| Синтаксоны Syntaxa | Элементы долины Elements of the valley | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Коренной склон Base rock slope | Надпойменные террасы Terraces above floodplains |
Нивальные элементы Nival elements |
Пойма Floodplain | |||||||
| A1 | A2 | B1.1 | B1.2 | B1.3 | IB1 | IB2 | C | C3.2 | C3.3 | |
| Ass. Dryado octopetalae–Hylocomietum splendentis subass. typicum var. Hedysarum arcticum fac. typicum | + | + | + | |||||||
| Luzula confusa–Salix nummularia com. type | + | |||||||||
| Ass. Dryado octopetalae–Hylocomietum splendentis subass. typicum var. Hedysarum arcticum fac. Trollius europaeus | + | |||||||||
| Hylocomium splendens–Pachypleurum alpinum com. type | + | |||||||||
| Alchemilla murbeckiana com. | + | |||||||||
| Hylocomium splendens–Salix glauca com. type fac. typicum | + | |||||||||
| Empetrum hermaphroditum–Salix glauca com. | + | |||||||||
| Hylocomium splendens–Salix glauca com. type fac. Carex juncella | + | |||||||||
| Hylocomium splendens–Salix glauca com. type fac. Salix hastata | + | |||||||||
| Poa arctica–Veratrum lobelianum com. | + | |||||||||
| Ass. Salici hastatae–Caricetum stantis fac. Equisetum arvense | + | |||||||||
| Sibbaldia procumbens–Veratrum lobelianum com. type | + | + | ||||||||
| Silene acaulis–Salix reticulata com. type | + | |||||||||
| Ass. Salici hastatae–Caricetum stantis fac. typicum | + | + | ||||||||
| Ass. Salici hastatae–Caricetum stantis fac. Carex juncella | + | |||||||||
| Petasites radiatus com. | + | + | ||||||||
Бровка (А1) и поверхность коренного склона (А2) чаще всего заняты фитоценозами асс. Dryado octopetalae–Hylocomietum splendentis субасс. typicum вар. Hedysarum arcticum, которые представляют собой производные сообщества от зональной для типичных тундр субасс. Dryado octopetalae–Hylocomietum splendentis typicum, впервые описанной на возвышенности Вангуреймусюр В.Н. Андреевым (1932). В них сохраняются многие диагностические виды зонального синтаксона, постоянно встречаются кустарнички Dryas octopetala, Salix reticulata, Vaccinium uliginosum subsp. microphyllum. Ценозы типовой фации обычны на бровке и преимущественно в верхних частях склона коренной террасы, а фац. Trollius europaeus, отличающиеся присутствием мезофильных травянистых растений – только на склоне. Сообщества Hylocomium splendens–Pachypleurum alpinum com. type распространены преимущественно на склонах южных экспозиций и Alchemilla murbeckiana com. – на крутых склонах южных экспозиций с длительным залеганием снежного покрова.
Исключительно на бровке склона выполнены описания сообществ Luzula confusa–Salix nummularia com. type, которые связаны с песчаными почвами и малоснежными ветрообдуваемыми местообитаниями.
На обследованном участке долины р. Большая Хэхэганъяха надпойменные террасы представлены солифлюкционными (B1), что обусловлено широким распространением на склонах долины криогенных процессов, протекающих с разной интенсивностью в зависимости от крутизны склона, его экспозиции и подстилающего субстрата. Несмотря на относительно размытые границы между элементами солифлюкционных террас, каждый из них отличается своеобразием синтаксонов. Так, поверхность относительно высоких и хорошо заросших террас (B1.1) часто покрыта сообществами асс. Dryado octopetalae–Hylocomietum splendentis субасс. typicum вар. Hedysarum arcticum фац. typicum, подобно бровкам и верхним частям коренных склонов долин, что свидетельствует о замедлении солифлюкционных процессов на таких участках и формировании относительно стабильных сообществ. Своеобразие поверхности солифлюкционных террас хорошо диагностируется сообществами ивняков с густым моховым покровом – Hylocomium splendens–Salix glauca com. type фац. typicum и Empetrum hermaphroditum–Salix glauca com. На пологих склонах солифлюкционной террасы (B1.2) описаны фитоценозы Hylocomium splendens–Salix glauca com. type фац. Carex juncella, а поверхность солифлюкционного вала (B1.3) занимают сообщества Hylocomium splendens–Salix glauca com. type фац. Salix hastata и Poa arctica–Veratrum lobelianum com.
К участкам с длительным сроком залегания снежного покрова (нивальным нишам и криопланационным террасам) приурочены нивальные сообщества Sibbaldia procumbens–Veratrum lobelianum com. type. В депрессии на границе с участком выхода на поверхность крупнообломочного материала, где из-за таяния долго лежащего снега создаются условия повышенной влажности, выполнено описание сообщества асс. Salici hastatae–Caricetum stantis фац. Equisetum arvense. Другой тип участков нивального мерзлотного ландшафта, длительнее пребывающих без снега в период вегетации, диагностируется сообществами Silene acaulis–Salix reticulata com. type.
Пойма реки слабо разделена на элементы, а иногда не выражена (профиль № 6). Растительность участков поймы, где нет ее отчетливой дифференциации на элементы, представлена сообществами асс. Salici hastatae–Caricetum stantis с 2 фациями – преобладающей фац. typicum и описанной единожды фац. Carex juncella. На отдельных участках выделяется прирусловая зона, которая хорошо диагностируется сообществами Petasites radiatus com., хотя здесь также обычны сообщества асс. Salici hastatae–Caricetum stantis фац. typicum.
Таким образом, большинство элементов геоморфологического профиля долины р. Большая Хэхэганъяха отличаются перечнем синтаксонов, которые отражают их положение на профиле и позицию в пределах конкретного эколого-динамического ряда. Разнообразие профилей характеризует высокую динамичность экологических рядов сообществ в долинах тундровых рек. Причем последняя обусловлена не только особенностями самой поймы (изменения вследствие колебания паводкового режима и сукцессионные смены сообществ, протекающие с различной скоростью на разных уровнях поймы), но и широким распространением в речных долинах криогенных процессов, из которых в обследованном районе преобладает солифлюкция. Каждый из эколого-динамических рядов изученных профилей отличается как перечнем сообществ разных синтаксонов, так и последовательностью их смены при продвижении от бровки коренного склона к прирусловой части поймы.
ЗАКЛЮЧЕНИЕ
Разнообразие местообитаний, обусловленное большим числом экзогенных процессов, и динамичный характер растительного покрова в пределах речных долин тундровых рек определили многообразие синтаксономических единиц низших рангов: 2 ассоциации (включая 1 субассоциацию, 1 вариант и 5 фаций), 5 типов сообществ (с 3 фациями) и 4 сообщества. Особенностью небольших речных долин тундровой зоны является наличие каменисто-щебнистых пляжей, почти лишенных растительности, а также слабая выраженность пойм с лугами.
Сообщества синтаксонов, которые сменяют друг друга в пределах эколого-динамических рядов речной долины, хорошо отражают экологические особенности местообитаний на геоботанических профилях. В этом отношении, видовой состав сообщества, которое в течение длительного времени формировалось на конкретной территории, является наиболее информативным показателем и индикатором, отражающим усредненные многолетние эколого-климатические параметры местообитания.
Список литературы
[A checklist…] Список лихенофлоры России. 2010. СПб. 194 с.
[Afonina, Czernyadjeva] Афонина О.M., Чернядьева И.В. 1995. Мхи Российской Арктики: список видов и библиография. – Arctoa: Бриол. журн. 5: 99–142.
[Andreyev] Андреев В.Н . 1932. Подзоны тундр Северного Края. – Природа. 10: 890–906.
[Geobotanicheskoe…] Геоботаническое районирование Нечерноземья европейской части РСФСР. 1989. Л. 64 с.
[Il’ina] Ильина И.С. 1968. Динамический принцип построения крупномасштабной геоботанической карты (на примере сухих степей Орского Зауралья). – Геоботаническое картографирование. Л. С. 21–37.
[Il’ina] Ильина И.С. 1991. Отражение пространственно-временных категорий растительности на крупномасштабных геоботанических картах. – Геоботаническое картографирование. СПб. С. 13–21.
[Isachenko] Исаченко Т.И. 1967. О картографировании серийных и микропоясных рядов в долинах и озерных котловинах (на примере р. Шарасун и оз. Большой Чандант Читинской области юго-восточного Забайкалья). – Геоботаническое картографирование. Л. С. 42–57.
[Kostenko] Костенко Н.П. 1985. Геоморфология. М. 312 с.
[Kunitskiy] Куницкий В.В. 2006. Нивальные мерзлотные ландшафты и образование экстранивитов. – Наука и образование. 4: 89–93.
[Lavrinenko] Лавриненко И.А. 2013. Геоботаническое районирование Большеземельской тундры и прилегающих территорий. – Геоботаническое картографирование. СПб. С. 74–92. https://doi.org/10.31111/geobotmap/2013.74
[Lavrinenko, Anufriev] Лавриненко И.А., Ануфриев В.В. 2009. Природный каркас восточноевропейских тундр. – В сб.: Материалы Международной научно-практической конференции “Сохранение биологического разнообразия наземных и морских экосистем в условиях высоких широт”. Мурманск. С. 150–153.
[Lavrinenko, Lavrinenko] Лавриненко О.В., Лавриненко И.А. 2004. Фитоиндикация изменений климата на северо-востоке европейской части России. – География и природные ресурсы. 2: 54–61.
[Lavrinenko, Lavrinenko] Лавриненко О.В., Лавриненко И.А. 2015. Сообщества класса Oxycocco-Sphagnetea Br.-Bl. Et r. Tx. 1943 в восточноевропейских тундрах. – Растительность России. 26: 55–84. https://doi.org/10.31111/vegrus/2015.26.55
[Lavrinenko, Lavrinenko] Лавриненко О.В., Лавриненко И.А. 2018. Зональная растительность равнинных восточноевропейских тундр. – Растительность России. 32: 35–108. https://doi.org/10.31111/vegrus/2018.32.35
[Lavrinenko et al.] Лавриненко О.В., Матвеева Н.В., Лавриненко И.А. 2016. Сообщества класса Scheuchzerio-Caricetea nigrae (Nordh. 1936) Tx. 1937 в восточноевропейских тундрах – Растительность России. 28: 55–88. https://doi.org/10.31111/vegrus/2016.28.55
[Lipatova] Липатова В.В. 1971. О показе растительности пойм на обзорных мелкомасштабных картах. – Геоботаническое картографирование. Л. С. 29–37.
[Makagonova] Макагонова М.А. 2009. Динамика параметров водообмена малых речных бассейнов в области Восточно-азиатского муссона. – География и природные ресурсы. 2: 139–145.
[Mirkin] Миркин Б.М. 1971. Статистический анализ экоклинов. – Бот. журн. 56 (12): 1772–1788.
[Pavleichik] Павлейчик В.М. 2009. Ландшафтно-экологический каркас Оренбургской области: методы идентификации и структура. – Вестник ВГУ. Серия: География. Геоэкология. 1: 5–12.
[Sekretareva] Секретарева Н.А. 2004. Сосудистые растения Российской Арктики и сопредельных территорий. М. 131 с.
[Stoyashcheva] Стоящева Н.В. 2007. Экологический каркас территории и оптимизация природопользования на юге Западной Сибири. Новосибирск. 140 с.
Weber H.E., Moravec J., Theurillat J.-P. 2000. International code of phytosociological nomenclature. 3rd ed. – J. Veg. Sci. 11 (5): 739–768. https://doi.org/10.2307/3236580
[Zony…] Зоны и типы поясности растительности России и сопредельных территорий. М: 1:8000000. 1999a. М. 2 с.
[Zony…] Зоны и типы поясности. Пояснительный текст и легенда к карте. 1999b. М. 64 с.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал