

Ботанический журнал, 2020, T. 105, № 9, стр. 861-877
ДИНАМИКА ПОПУЛЯЦИЙ ANEMONASTRUM BIARMIENSE (RANUNCULACEAE) В ЮЖНО-УРАЛЬСКОМ ЗАПОВЕДНИКЕ (РЕСПУБЛИКА БАШКОРТОСТАН)
О. В. Юсупова 1, 2, 3, *, Л. М. Абрамова 2, ***, И. Р. Юсупов 1, **
1 Южно-Уральский государственный природный заповедник
453560 Белорецкий р-н, д. Реветь, Центральная усадьба, Россия
2 Южно-Уральский ботанический сад-институт – обособленное структурное подразделение
Уфимского федерального исследовательского центра РАН
450080 г. Уфа, ул. Менделеева, 195/3, Россия
3 Национальный парк “Зюраткуль”
456910 Саткинский р-н, г. Сатка, ул. Комсомольская, 13, Россия
* E-mail: yusupova_ov@mail.ru
*** E-mail: abramova.lm@mail.ru
** E-mail: i777yus@yandex.ru
Поступила в редакцию 27.02.2020
После доработки 06.05.2020
Принята к публикации 19.05.2020
Аннотация
Представлены результаты изучения разногодичной динамики морфометрических параметров и виталитетной структуры пяти ценопопуляций эндемичного вида Урала Anemonastrum biarmiense s. l. на территории Южно-Уральского государственного природного заповедника в Республике Башкортостан. Высокие средние значения морфометрических параметров отмечены у растений A. biarmiense в урочище Василевские поляны и на хребте Белятур, минимальные значения большинства параметров – на хребте Нараташ в подгольцовом поясе. Наибольшей изменчивостью характеризуются следующие параметры: число цветоносных побегов (Cv = 19.55–62.87%), число розеточных листьев (Cv = 17.61–40.34%), наименьшей – высота цветоносного побега (Cv = 9.76–21.48%), диаметр цветка (Cv = 4.41–28.12%). На межпопуляционном уровне амплитуда изменчивости для шести из десяти признаков A. biarmiense варьирует от средней до повышенной. Наиболее высокая изменчивость по всем изучаемым параметрам наблюдается в ЦП Нараташ (Cv = 20.32–49.88%) с самыми контрастными условиями произрастания. Низкая изменчивость отмечена в ценопопуляциях Дунан-сунган (Cv = 12.06–29.19%), Юша (Cv = 11.89–29.18%), Василевские поляны (Cv = 12.03–30.20%), расположенных на относительно выровненных участках в луговых ценозах. Условия экотопа оказывают более сильное воздействие на параметры растений по сравнению с погодными условиями. Анализ жизненности ценопопуляций A. biarmiense показал, что они неоднородны по своему составу: виталитетный тип их меняется от процветающего до депрессивного, в том числе и по годам исследований.
Уральские эндемичные виды длительное время являются объектами изучения в связи с уникальностью и уязвимостью их популяций. Среди приоритетных направлений стратегии сохранения редких видов является территориальная охрана “in situ”, которая существует благодаря развитию и совершенствованию сетей особо охраняемых природных территорий (Mirkin et al., 2004; Viktorov, 2018). На Южном Урале многолетние научные исследования, направленные на изучение популяций редких видов растений, ведутся в Южно-Уральском государственном природном заповеднике на территории Республики Башкортостан.
В составе флоры этой особо охраняемой природной территории, богатой редкими видами сосудистых растений, встречается высокогорно-луговой эндемичный вид ветреник пермский – Anemonastrum biarmiense (Juz.) Holub (Ranunculaceae). Представители рода Anemonastrum распространены в горных системах по всему миру, в том числе в Евразии и на Аляске, и образуют между собой близкие переходные виды.
Во флоре России известно девять видов, а на Урале встречается три – A. biarmiense, A. crinitum (Juz.) Holub, A. sibiricum (L.) Holub (Mamaev, Knyazev, 1995; Endemic plants …, 2013). На Северном и Приполярном Урале A. biarmiense замещается восточносибирско-дальневосточным A. sibiricum, а на Южном Урале образует смешанные популяции с южносибирским высокогорным A. crinitum (Tzvelev, 2001). Реликтовая популяция A. crinitum, по литературным данным, известна на вершине хребта Большой Нургуш (Mamaev, Knyazev, 1995).
Первые результаты изучения биологии A. biarmiense нами опубликованы ранее (Karimova et al., 2013; Yusupova et al., 2016; 2017; 2018). Учитывая высказывание Н.Н. Цвелева (Tzvelev, 2001) о том, что процесс взаимного поглощения и вторичного сближения видов рода Anemonastrum в уральской части ареала происходит и в настоящее время, мы рассматриваем A. biarmiense в широком понимании.
До настоящего времени ведутся исследования биологии и экологии, таксономии и филогении рода Anemonastrum, изучается распространение его представителей в разных горных системах по всему миру (Dutton et al., 1995; Malinovs’kij, 2002; Hoot et al., 2012; Abdaladze et al., 2015; Mosyakin, 2016; 2018; Mosyakin, Lange, 2018 и др.).
Для A. biarmiense в литературе имеются сведения об анатомо-морфологических особенностях строения растений, возрастной структуре популяций, изменчивости признаков, семенной продуктивности, распространении и экотопологической приуроченности на Северном и Приполярном Урале (Vernigor, 1981; Mineeva, 1985; Bobretsova, 2002; Degteva, 2008; Plotnikova, 2009; Poletaeva, 2015; Valuiskikh, Kanev, 2019). На территории Южно-Уральского заповедника особенности структуры популяций A. biarmiense и его биологии выявлены впервые.
Целью данной работы является исследование варьирования морфометрических признаков Anemonastrum biarmiense в пяти локалитетах Южно-Уральского заповедника в разные годы и характеристика виталитетной структуры ценопопуляций.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
A. biarmiense – эндемичный вид Урала, распространенный в горном поясе от Южного Урала до южной части Полярного Урала. Согласно классификации жизненных форм И.Г. Серебрякова (Serebryakov, 1959), вид является розеточным короткокорневищным травянистым поликарпиком с моноподиальным нарастанием осевого побега. Приурочен к приручейным разнотравным горно-тундровым лужайкам и травяно-моховым тундрам, способен выступать в качестве доминанта на участках, занимаемых горно-тундровыми сообществами. Заходит в горно-лесной пояс, где произрастает по лесным полянам и опушкам. По происхождению принадлежит к Уральской викарной расе полиморфных видовых комплексов, широко распространенных в высокогорьях различных горных систем Северной Евразии и Голарктики в целом, возникших в результате дифференциации и раздробления ареала общего предкового вида (Endemic plants…, 2013).
А. biarmiense мы относим к типично высокогорным растениям по причине его выраженных адаптационных черт к экстремальным условиям (укороченный вегетационный сезон, перепады температур с летними заморозками, морозобои, обедненные маломощные почвы, интенсивные инсоляция и ветер). Комплекс адаптаций, включающий компактную жизненную форму, миниатюризацию, отсутствие специализированного защитного покрова почек возобновления и их раннего формирования, геофитизацию, ксерофитизацию и психроморфность, раннее подснежное развитие, И.В. Волков (Volkov, 2007) относит к пассивному комплексу, способствующему увеличению устойчивости растений к неблагоприятным факторам среды.
На Южном Урале вид встречается на всех горных хребтах с отметками от 1000 до 1640 м над ур. м., также места его произрастания отмечаются в горно-лесном поясе, ниже 1000 м над ур. м. Как эндемичный вид, рекомендован к охране во всем Уральском регионе (Gorchakovsky, Shurova, 1982; Kucherov et al., 1987). С третьей категорией редкости вид включен в Красные книги Свердловской области (Krasnaya …, 2018), Республики Коми (Krasnaya …, 2019), Ханты-Мансийского (Krasnaya …, 2013) и Ямало-Ненецкого (Krasnaya …, 2010) автономных округов.
Исследования на территории Южно-Уральского государственного природного заповедника (Республика Башкортостан) проводились с 2015 по 2018 г. Мониторинговые наблюдения вели в пяти ценопопуляциях (рис. 1), расположенных в Центральной (Уфимско-Бельской) подпровинции горно-лесной провинции, к которой относятся Таганайско-Ямантауский гольцово-таежный округ и Инзерско-Масимский лугово-лесной увалисто-хребтовый округ (Fiziko-geograficheskoe…, 1964; Kadilnikov, 1966).
Рис. 1.
Карта-схема расположения Южно-Уральского заповедника на Южном Урале и локализации ценопопуляций Anemonastrum biarmiense на данном ООПТ.
Fig. 1. Schematic map of the South Ural Reserve location in the South Urals, and localization of Anemonastrum biarmiense coenopopulations in this protected area.
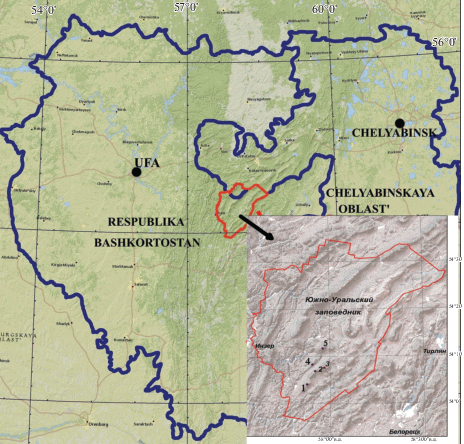
Таганайско-Ямантауский округ Уфимско-Бельской подпровинции представляет наиболее высокую часть среднегорий Южного Урала (горные массивы Большой Иремель и Большой Нургуш, гора Уван, хребты Таганай, Москаль, Зигальга, Машак, Нары, Нараташ). Хребты сложены кварцитами, межгорные понижения – сланцами и доломитами (Gorichev, 2008). В растительности выделяются три пояса – горно-таежный, подгольцовый и горно-тундровый. В горно-лесном поясе распространены горно-лесные серые, бурые и дерновоподзолистые почвы, а горно-тундровые, горно-луговые – в высокогорной области. В составе растительности преобладают горно-таежные пихтово-еловые леса. Среднемесячные температуры июля и января соответственно +17°С и –15.8°С.
В Инзерско-Масимский округ входят несколько районов, из которых нами обследован Караташско-Юрматауский р-н (хребты Белятур, Юша). В сложении горных пород участвуют сланцы, песчаники, конгломераты, алевролиты, аргиллиты, известняки и доломиты. Климат округа более континентальный. Среднегодовая температура воздуха на вершинах высоких хребтов колеблется от 1°С до –0.5°С. В связи с расчлененностью рельефа почвенный покров пестрый. На хребтах развиты светло-серые лесные и маломощные горные почвы, в межгорных понижениях почвы более богаты гумусом. В растительном покрове преобладают березовые, осиновые и сосновые леса.
Краткая характеристика местообитаний изученных ценопопуляций представлена в табл. 1.
Таблица 1.
Характеристика местообитаний изученных ценопопуляций (ЦП) Anemonastrum biarmiense Table 1. Habitat characteristics for the studied coenopopulations (CP) of Anemonastrum biarmiense
| Название ЦП/ Name of CP | Местонахож-дение ЦП/ Location of CP | Пояс расти-тельности/ Belt vegetation | Высота над ур. м., м; координаты/ Altitude, m a. s. l.; coordinates | Местообитание, экотоп/Habitat, ecotope | Синтаксономи-ческая единица растительности/ Syntactic unit of vegetation |
|---|---|---|---|---|---|
| Дунан-сунган/ Dunan-Sungan | Горный хребет Юша/ Yusha Ridge | Горно-лесной/ Montane forest | 943 м/m N 54.0686 E 57.8768 |
Горный высокотравный ксерофитный луг/ Montane tall- grass dry meadow | Сообщество/
Community Dracocephalum ruyschiana-Tephroseris integrifolia |
| Василевские поляны/ Vasilevskiye Glades | -"- | -"- | 1020 м/m N 54.1234 E 57.9578 |
Горный высокотравный луг/Tall-grass montane meadow | Ассоциация/
Association Anemonastro biarmiensis- Calamagrostietum arundinaceae |
| Юша/Yusha | -"- | -"- | 977 м/m N 54.1149 E 57.9212 |
-"- | -"- |
| Белятур/ Belyatur | Горный хребет Белятур/ Belyatur Ridge | -"- | 967 м/m N 54.1384 E 57.9059 |
Березово- лиственичное редколесье/ Birch-larch woodland | Ассоциация/
Association Anemonastro biarmiensis- Laricetum sukaczewii |
| Нараташ/ Naratash | Горный хребет Нараташ/ Naratash Ridge | Подгольцовый/ Subalpine | 1162 м/m N 54.1945 E 57.9625 |
Тундроподобное сообщество/ Tundra-like community | Ассоциация/
Association Diantho-Festucetum igoschiniae |
Согласно эколого-флористической классификации растительности Башкортостана (Yamalov et al., 2012), три из исследованных ценопопуляций (Дунан-сунган, Юша, Василевские поляны) произрастают в сообществах высокотравных послелесных лугов союза Polygonion krascheninnikovii Kashapov 1985 порядка Carici macrourae-Сrepidetalia sibiricae Ermakov et al. 1999 класса Molinio-Arrhenatheretea R. Tx. 1937, среди них ЦП Дунан-сунган приурочена к сообществу Dracocephalum ruyschiana-Tephroseris integrifolia ксерофитных лугов, а ЦП Василевские поляны и Юша – к ассоциации Anemonastro biarmiensis-Calamagrostietum arundinaceae Shirokikh et al. 2018 (Shirokih, 2018). ЦП Белятур приурочена к березово-лиственничному редколесью союза Trollio europaea-Pinion sylvestris Fedorov ex Ermakov et al. 2000 порядка Chamaecytiso ruthenici-Pinetalia sylvestris Solomeshch et Ermakov in Ermakov et al. 2000 класса Brachypodio рinnati-Betuletea рendulae Ermakov, Koroljuk et Latchinsky 1991 (aсс. Anemonastro biarmiensis-Laricetum sukaczewii Solomeshch et Martynenko 2009 prov.). ЦП Нараташ произрастает в тундроподобном сообществе елово-березового криволесья союза Anemonastro sibiricae-Festucion ovinae Chytry et al. 1993 порядка Juncetalia trifidi Daniels 1994 класса Caricetea сurvulae Br.-Bl. 1948 (асс. Diantho-Festucetum igoschiniae Ishbirdin et al. 1996).
На основании показаний термохронов (устройство 21A3EF2D0000008F DS1921G-F5: от –40°C до +85°C/0.5°C), установленных на территории Южно-Уральского заповедника в горной части, составлен график данных по минимальной температуре с мая по июль 2015–2018 гг. (рис. 2).
Рис. 2.
Показания минимальных температур в период вегетации Anemonastrum biarmiense в Южно-Уральском заповеднике с 2015 по 2018 гг.
Fig. 2. Values of minimum temperatures during the growing seasons of Anemonastrum biarmiense in the South Ural Reserve from 2015 to 2018.
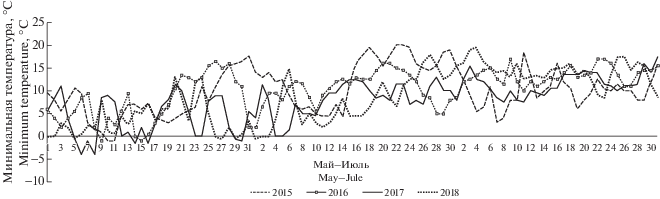
В дополнение к графику приведена краткая метеорологическая характеристика вегетационных сезонов с 2015 по 2018 гг. из материалов “Летописей природы Южно-Уральского государственного природного заповедника” (Letopis’ …, 2015–2018 гг.). Включены данные наблюдений, сделанных на центральной усадьбе заповедника – минимальные и максимальные суточные температуры, наличие осадков (табл. 2).
Таблица 2.
Метеорологическая характеристика вегетационных сезонов 2015–2018 гг. Table 2. Meteorological characteristics of vegetation seasons 2015–2018
| Год Year | Окончание весны Late spring | Лето Summer | ||||||
|---|---|---|---|---|---|---|---|---|
| Месяцы (дни) Months (days) | Число дней Number of days | Месяцы (дни) Months (days) | Число дней Number of days | |||||
| с заморозком with freeze | с осадками with precipitation | with t > 15°С | with t > 25°С | with t >30°С | с осадками with precipitation | |||
| 2015 | Май/May (1–28) | – | 17 | – | Май/May (29–31) | 3 | 3 | 2 |
| Июнь/June | 21 | 16 | 15 | |||||
| Июль/July | 10 | – | 22 | |||||
| Август/August (1–23) | 8 | – | 12 | |||||
| Всего/Total | – | 17 | – | Всего/Total | 42 | 19 | 51 | |
| 2016 | Май/May (1–19) | 6 | 15 | 6 | Май/May (20–31) | 8 | 1 | 3 |
| Июнь/June | 17 | 4 | 15 | |||||
| Июль/July | 28 | 8 | 14 | |||||
| Август/August (1–29) | 27 | 25 | 8 | |||||
| Всего/Total | 6 | 15 | 6 | Всего/Total | 80 | 38 | 40 | |
| 2017 | Май/May | 8 | 16 | 17 | Июнь/June (13–30) | 6 | – | 13 |
| Июнь/June (1–12) | 1 | 9 | 9 | Июль/July | 11 | 3 | 23 | |
| Август/August | 15 | 4 | 7 | |||||
| Сентябрь/September (1–7) | 4 | 1 | 2 | |||||
| Всего/Total | 9 | 25 | 26 | Всего/Total | 36 | 8 | 45 | |
| 2018 | Май/May | 7 | 22 | 16 | Июнь/June (19–30) | 7 | 1 | 6 |
| Июнь/June (1–18) | – | 11 | 14 | Июль/July | 21 | 7 | 12 | |
| Август/August (1–26) | 9 | – | 16 | |||||
| Всего/Total | 7 | 33 | 30 | Всего/Total | 37 | 8 | 34 | |
Влияние погодных условий на раннецветущие растения сильнее отражается в период их активного роста и цветения, преимущественно в мае-июне. Опираясь на полученные цифры, следует указать, что из совокупности дней с заморозком, наибольшее их число отмечено для 2017 г. (9 дней), и не отмечено вовсе для 2015 г. Из совокупности дней с осадками, выпавшими за время вегетационного сезона, дождливыми являются 2017 г. (70 дней), 2015 г. (68 дней), 2018 г. (67 дней). Меньше всего осадков отмечено для 2016 г. (55 дней). Из совокупности дней с температурой от 15 до 30°С наиболее теплым является 2016 г. (124 дня), наименее – 2015 г. (61 день).
Условия обитания ветреника пермского в горно-тундровой области Южного Урала характеризуются высокой влажностью субстрата за счет конденсации влаги, происходящей в результате восходящего движения воздушных масс, повышенной суммой осадков, а также укороченным вегетационным сезоном. В каждой конкретной ценопопуляции условия экотопа отличаются – например, в расположенной на высоте 1162 м над ур. м. ценопопуляции Нараташ средние температуры ниже, чем на высотах 943–967 м над ур. м.; имеют значение экспозиция склона, почвенный покров, затенение и прочие абиотические и биотические факторы.
Изучение морфометрии в природных условиях проводилось согласно методу В.Н. Голубева (Go-lubev, 1962) на 25 средневозрастных особях во всех ценопопуляциях A. biarmiense. Наблюдения и измерения проводились в фазе цветения, при этом учитывались следующие параметры: Ngs – число цветоносных побегов на одно растение; H – высота цветоносного побега (см); D – диаметр стебля (см); Nl – число розеточных листьев в розетке; Ll – длина листового сегмента, (см); Sl – ширина листового сегмента (см); Lp – длина черешка розеточного листа (см); Li – длина цветоножки (см); Nfl – число цветков в соцветии; Dfl – диаметр цветка (см).
Изучение виталитетного состава проводилась по методике Ю.А. Злобина (Zlobin, 1989), основанной на дифференциации растений одного онтогенетического состояния на классы виталитета. В качестве объектов виталитетного анализа использовались растения средневозрастного генеративного онтогенетического состояния, которое в наибольшей степени влияет на семенное самоподдержание ценопопуляций. Предварительно были проведены факторный и корреляционный анализы для определения детерминирующего комплекса признаков. После обработки данных были составлены виталитетные спектры, отражающие соотношения растений высшего (а), промежуточного (b) и низшего (с) классов виталитета, а также определен индекс качества (Q) ценопопуляции и виталитетные типы: процветающие, равновесные, депрессивные.
Статический анализ провели в MS Excel 2007 при помощи пакета статистических программ Statistica 6.0 c использованием стандартных показателей. При статистическом анализе количественных показателей рассчитывали средние арифметические значения и их ошибки (M ± m) (Zaytsev, 1990). Общую межпопуляционную и внутрипопуляционную изменчивость морфометрических параметров растений оценивали по абсолютным средним значениям изучаемых признаков и значениям коэффициента вариации (Cv, %) с учетом эмпирической шкалы изменчивости С.А. Мамаева (Mamaev, 1975): очень низкая – меньше 7%, низкая – 8–12%, средняя – 13–20%, повышенная – 21–30%, высокая – 31–40%, очень высокая – больше 40%.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Морфометрические методы являются одним из основных способов получения репрезентативного объема данных об особях и состоянии конкретных популяций, произрастающих в разных экологических условиях и находящихся под разной степенью антропогенного воздействия. С применением данных методов можно зафиксировать изменения габитуса растений в горно-лесном и горно-тундровом поясах и, таким образом, выявить адаптационные возможности A. biarmiense в условиях высокогорий.
В таблице 3 представлены значения параметров растений ценопопуляций A. biamiense в разные годы исследований. Установлено, что по максимальным средним значениям морфометрических показателей отличаются растения из ценопопуляций, произрастающие в березовом редколесье на хр. Белятур и в высокотравном мезофильном луговом сообществе в урочище Василевские поляны. Наиболее высокие средние значения параметров выявлены у растений из ценопопуляции Василевские поляны. Повышенные значения параметров у растений из лугового местопроизрастания обусловлены, на наш взгляд, обилием влаги, образующейся вследствие таяния мощных слоев снега, поступающей в виде дождей. Растения из ценопопуляции Белятур также отличаются высокими средними значениями по многим из линейных параметров, такими как высота цветоносных побегов, длина и ширина листового сегмента розеточного листа, длина черешка розеточного листа и др. Среди числовых параметров у растений этой ценопопуляции нет высоких значений, к примеру, число цветоносных побегов варьирует по годам от 2.64 до 3.60. Аналогичные результаты получены в Печоро-Илычском заповеднике, где наиболее крупные размеры особей наблюдались в горном березняке на Северном Урале (Bobretsova, 2002; Plotnikova, 2009). Вероятно, из-за присутствия в сообществе деревьев, которые определяют произрастание на данном участке многих опушечных и лесных видов растений, занимающих доминирующее положение по высоте и продукции (Crepis sibirica L., Aconitum nemorosum Bieb. ex Reichenb., Pleurospermum uralense Hoffm. и др.), особи A. biarmiense таким образом приспосабливаются к условиям с недостатком солнечного света, максимально высоко вынося соцветия и листья относительно травостоя.
Таблица 3.
Внутрипопуляционная изменчивость морфометрических признаков Anemonastrum biarmiense в ценопопуляциях Table 3. Intrapopulation variability of morphometrical characteristics of Anemonastrum biarmiense in the coenopopulations
| Годы наблюдений Years of observations |
Средние значения морфометрических параметров (n = 25) Average values of morphometric parameters |
||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Ngs | H | D | Nl | Ll | Sl | Lp | Li | Nfl | Dfl | ||
| Дунан-сунган/Dunan-Sungan | |||||||||||
| 2015 | M ± m | 4.48 ± 0.30 | 44.68 ± 0.94 | 0.68 ± 0.01 | 10.72 ± 0.40 | 7.59 ± 0.25 | 10.20 ± 0.37 | 20.64 ± 0.56 | 9.90 ± 0.31 | 5.56 ± 0.17 | 4.08 ± 0.07 |
| Cv,% | 32.93 | 10.54 | 10.91 | 18.56 | 16.21 | 17.91 | 13.61 | 15.71 | 15.64 | 8.00 | |
| 2016 | M ± m | 4.00 ± 0.20 | 52.41 ± 1.60 | 0.59 ± 0.02 | 9.72 ± 0.41 | 8.63 ± 0.27 | 9.23 ± 0.35 | 22.04 ± 0.74 | 10.45 ± 0.28 | 5.12 ± 0.15 | 3.78 ± 0.11 |
| Cv,% | 25.00 | 15.28 | 20.06 | 21.32 | 15.4 | 18.71 | 16.85 | 13.26 | 14.17 | 14.18 | |
| 2017 | M ± m | 3.60 ± 0.19 | 31.52 ± 1.02 | 0.71 ± 0.02 | 8.28 ± 0.29 | 6.46 ± 0.14 | 7.09 ± 0.25 | 14.52 ± 0.35 | 5.32 ± 0.18 | 4.88 ± 0.17 | 3.16 ± 0.07 |
| Cv,% | 26.59 | 16.18 | 16.77 | 17.61 | 10.73 | 17.46 | 12.06 | 17.24 | 17.06 | 11.3 | |
| 2018 | M ± m | 4.20 ± 0.27 | 57.45 ± 1.35 | 0.71 ± 0.04 | 8.24 ± 0.39 | 9.15 ± 0.30 | 8.95 ± 0.34 | 28.38 ± 0.83 | 10.68 ± 0.25 | 4.96 ± 0.15 | 3.76 ± 0.11 |
| Cv,% | 32.24 | 11.75 | 25.76 | 23.57 | 16.3 | 19.23 | 14.61 | 11.63 | 14.81 | 14.75 | |
| 2015–2018 | min max |
2.0 | 20.5 | 0.4 | 5 | 5.1 | 4.5 | 11.7 | 4.2 | 3.0 | 2.3 |
| 9.0 | 74.3 | 1.0 | 14 | 12.4 | 13.7 | 38.1 | 15.8 | 8.0 | 5.4 | ||
| Юша/Yusha | |||||||||||
| 2015 | M ± m | 4.10 ± 0.26 | 50.38 ± 1.06 | 0.65 ± 0.03 | 9.48 ± 0.51 | 8.81 ± 0.17 | 9.61 ± 0.27 | 25.20 ± 0.84 | 11.84 ± 0.20 | 5.28 ± 0.11 | 3.60 ± 0.08 |
| Cv,% | 33.07 | 10.51 | 24.85 | 26.74 | 9.87 | 13.86 | 16.57 | 8.25 | 10.26 | 10.85 | |
| 2016 | M ± m | 4.36 ± 0.29 | 40.78 ± 1.05 | 0.69 ± 0.03 | 9.56 ± 0.50 | 7.39 ± 0.26 | 7.73 ± 0.33 | 17.77 ± 0.69 | 7.47 ± 0.36 | 5.56 ± 0.17 | 3.47 ± 0.06 |
| Cv,% | 33.68 | 12.93 | 18.44 | 26.34 | 17.39 | 21.28 | 19.41 | 24.16 | 15.65 | 8.38 | |
| 2017 | M ± m | 4.48 ± 0.21 | 31.62 ± 0.85 | 0.74 ± 0.03 | 8.96 ± 0.37 | 6.48 ± 0.19 | 7.53 ± 0.29 | 14.65 ± 0.43 | 4.49 ± 0.17 | 5.20 ± 0.22 | 3.07 ± 0.10 |
| Cv,% | 23.34 | 13.46 | 16.95 | 20.51 | 14.35 | 19.18 | 14.57 | 18.73 | 21.5 | 15.54 | |
| 2018 | M ± m | 4.60 ± 0.24 | 40.81 ± 0.87 | 0.72 ± 0.03 | 8.84 ± 0.41 | 8.03 ± 0.30 | 8.13 ± 0.28 | 19.82 ± 0.60 | 6.40 ± 0.23 | 5.20 ± 0.16 | 3.41 ± 0.10 |
| Cv,% | 26.62 | 10.69 | 18.37 | 23.02 | 18.59 | 17.08 | 15.11 | 17.72 | 15.7 | 15.2 | |
| 2015–2018 | min max |
2.0 | 22.6 | 0.4 | 4.0 | 4.8 | 5.2 | 11.3 | 3.2 | 4.0 | 2.2 |
| 8.0 | 59.8 | 1.0 | 15.0 | 11.0 | 13.2 | 34.7 | 13.1 | 8.0 | 4.9 | ||
| Белятур/Belyatur | |||||||||||
| 2015 | M ± m | 2.64 ± 0.15 | 52.49 ± 1.51 | 0.62 ± 0.02 | 9.04 ± 0.54 | 8.66 ± 0.32 | 9.99 ± 0.34 | 30.38 ± 0.86 | 10.96 ± 0.21 | 5.56 ± 0.23 | 3.42 ± 0.09 |
| Cv,% | 28.68 | 14.39 | 19.24 | 29.87 | 18.32 | 16.95 | 14.17 | 9.48 | 20.82 | 13.31 | |
| 2016 | M ± m | 2.96 ± 0.19 | 48.66 ± 1.20 | 0.64 ± 0.02 | 9.00 ± 0.47 | 7.53 ± 0.25 | 8.02 ± 0.25 | 23.37 ± 0.98 | 9.18 ± 0.33 | 4.96 ± 0.19 | 3.25 ± 0.10 |
| Cv,% | 31.57 | 12.29 | 19.24 | 26.25 | 16.86 | 15.3 | 20.92 | 17.99 | 18.84 | 15.09 | |
| 2017 | M ± m | 3.21 ± 0.30 | 48.93 ± 1.75 | 0.62 ± 0.03 | 8.16 ± 0.44 | 8.81 ± 0.28 | 10.02 ± 0.44 | 25.55 ± 0.87 | 9.89 ± 0.38 | 5.04 ± 0.27 | 3.25 ± 0.09 |
| Cv,% | 46.87 | 17.91 | 22.79 | 27.09 | 16.15 | 22.07 | 17.01 | 19 | 26.54 | 13.85 | |
| 2018 | M ± m | 3.60 ± 0.34 | 49.38 ± 1.49 | 0.66 ± 0.03 | 9.44 ± 0.44 | 8.38 ± 0.31 | 9.22 ± 0.37 | 24.38 ± 1.04 | 9.37 ± 0.42 | 5.52 ± 0.24 | 3.37 ± 0.09 |
| Cv,% | 46.76 | 15.09 | 23.16 | 23.12 | 18.48 | 19.95 | 21.42 | 22.42 | 21.64 | 13.66 | |
| 2015–2018 | min max |
2.0 | 25.6 | 0.4 | 4.0 | 5.5 | 6.1 | 9.4 | 6.2 | 3.0 | 2.1 |
| 6.0 | 72.6 | 1.0 | 14.0 | 12.4 | 18.6 | 36.8 | 14.1 | 9.0 | 4.7 | ||
| Василевские поляны/Vasilevskiye Glades | |||||||||||
| 2015 | M ± m | 3.76 ± 0.26 | 45.37 ± 0.98 | 0.67 ± 0.03 | 9.36 ± 0.49 | 8.24 ± 0.26 | 8.81 ± 0.24 | 21.20 ± 0.77 | 8.38 ± 0.36 | 6.01 ± 0.24 | 3.54 ± 0.10 |
| Cv,% | 34.57 | 10.81 | 20.83 | 25.96 | 15.91 | 13.37 | 18.16 | 21.37 | 19.84 | 14.59 | |
| 2016 | M ± m | 4.08 ± 0.30 | 45.42 ± 0.89 | 0.64 ± 0.03 | 9.08 ± 0.41 | 7.54 ± 0.25 | 8.89 ± 0.37 | 22.97 ± 0.66 | 8.16 ± 0.25 | 5.44 ± 0.21 | 3.72 ± 0.07 |
| Cv,% | 36.71 | 9.76 | 20.74 | 22.68 | 16.79 | 20.89 | 14.44 | 15.43 | 19.23 | 9.94 | |
| 2017 | M ± m | 4.64 ± 0.18 | 57.54 ± 1.44 | 0.76 ± 0.03 | 10.08 ± 0.42 | 9.21 ± 0.23 | 10.64 ± 0.41 | 29.35 ± 0.91 | 9.96 ± 0.23 | 5.04 ± 0.17 | 3.82 ± 0.10 |
| Cv,% | 19.55 | 12.51 | 16.99 | 20.63 | 12.49 | 19.09 | 15.31 | 11.54 | 16.68 | 13.44 | |
| 2018 | M ± m | 4.12 ± 0.25 | 38.36 ± 1.15 | 0.76 ± 0.02 | 8.36 ± 0.35 | 7.30 ± 0.22 | 7.88 ± 0.31 | 17.93 ± 0.71 | 6.94 ± 0.25 | 4.92 ± 0.19 | 3.42 ± 0.10 |
| Cv,% | 29.99 | 15.03 | 16.24 | 20.68 | 15.29 | 19.61 | 19.82 | 17.85 | 19.39 | 14.68 | |
| 2015–2018 | min max |
2.0 | 30.2 | 0.5 | 5.0 | 5.2 | 4.6 | 10.5 | 4.9 | 3.0 | 2.4 |
| 8.0 | 75.4 | 1.1 | 15.0 | 11.5 | 17.7 | 36.6 | 12.7 | 6.0 | 4.8 | ||
| Нараташ/Naratash | |||||||||||
| 2015 | M ± m | 2.12 ± 0.18 | 33.49 ± 1.32 | 0.46 ± 0.02 | 7.28 ± 0.59 | 6.45 ± 0.30 | 6.84 ± 0.33 | 15.99 ± 0.86 | 5.28 ± 0.13 | 5.64 ± 0.26 | 3.30 ± 0.11 |
| Cv,% | 41.57 | 19.71 | 23.75 | 40.34 | 23.58 | 24.06 | 26.87 | 12.73 | 22.82 | 16.41 | |
| 2016 | M ± m | 2.36 ± 0.19 | 30.28 ± 1.46 | 0.40 ± 0.01 | 8.28 ± 0.50 | 6.01 ± 0.24 | 6.28 ± 0.28 | 12.84 ± 0.68 | 5.33 ± 0.44 | 5.16 ± 0.24 | 3.34 ± 0.12 |
| Cv,% | 40.35 | 24.08 | 18.19 | 29.89 | 20.14 | 22.55 | 26.55 | 41.16 | 22.85 | 18.00 | |
| 2017 | M ± m | 2.48 ± 0.27 | 20.27 ± 0.74 | 0.49 ± 0.02 | 8.16 ± 0.35 | 4.57 ± 0.17 | 4.59 ± 0.22 | 9.14 ± 0.47 | 3.79 ± 0.20 | 4.76 ± 0.19 | 2.09 ± 0.16 |
| Cv,% | 54.75 | 18.33 | 21.09 | 21.71 | 18.79 | 23.73 | 25.75 | 26.69 | 19.44 | 28.12 | |
| 2018 | M ± m | 2.52 ± 0.32 | 19.92 ± 0.78 | 0.49 ± 0.02 | 8.16 ± 0.35 | 4.57 ± 0.17 | 4.59 ± 0.22 | 9.14 ± 0.47 | 2.71 ± 0.19 | 4.76 ± 0.19 | 3.31 ± 0.14 |
| Cv,% | 62.87 | 19.69 | 21.09 | 21.71 | 18.79 | 23.73 | 25.75 | 34.65 | 19.44 | 20.81 | |
| 2015–2018 | min max |
1.0 | 11.7 | 0.3 | 4.0 | 3.1 | 2.7 | 6.1 | 1.1 | 2.0 | 1.1 |
| 7.0 | 46.9 | 0.7 | 18.0 | 9.7 | 10.7 | 18.6 | 9.1 | 9.0 | 4.6 | ||
| Средние значения/ Average values | 3.45 | 44.05 | 0.62 | 9.09 | 7.90 | 8.45 | 21.28 | 9.25 | 5.17 | 3.28 | |
Примечание. Ngs – число цветоносных побегов на одно растение; H – высота цветоносного побега, см; D – диаметр стебля, см; Nl – число розеточных листьев в розетке; Ll – длина листового сегмента, см; Sl – ширина листового сегмента, см; Lp – длина черешка розеточного листа, см; Li – длина цветоножки, см; Nfl – число цветков в соцветии; Dfl – диаметр цветка, см.
Note. Ngs – the number of flower-bearing shoots per plant, pcs.; H – height of flower-bearing shoot, cm; D – stem diameter, cm; Nl – the number of rosette leaves per rosette, pcs.; Ll – the length of leaf segment, cm; Sl – the width of leaf segment, cm; Lp – the length of rosette leaf petiole, cm; Li – peduncle length, cm; Nfl – the number of flowers per inflorescence, pcs.; Dfl – flower diameter, cm.
Менее высокие значения среди параметров отмечены у растений из ценопопуляций Дунан-сунган и Юша, при этом большинство параметров имеют близкие значения в каждой из них. Данные ценопопуляции приурочены к высокотравным лугам горно-лесного пояса. Они отличаются экспозицией склона и составом трав, но имеют сходство по высоте, на которой расположены (943 и 977 м над ур. м.).
Характерной особенностью растений из этих ценопопуляций является образование повышенного числа цветков и их большего диаметра. Так диаметр цветков на хр. Юша варьирует от 2.2 до 4.9 см, а на горе Дунан-сунган – от 2.3 до 5.4 см, у растений ЦП Дунан-сунган также отмечено наибольшее значение длины цветоножки (15.8 см), в сравнении с растениями из других ценопопуляций. По-видимому, более благоприятный термический режим позволяет растениям затрачивать больше энергетических ресурсов на репродукцию.
Минимальные средние значения среди параметров отмечены для ценопопуляции Нараташ, где сказываются наименее благоприятные по температурным характеристикам климатические условия для роста растений. Следует отметить, что на хр. Нараташ у растений возрастает вариабельность числа цветков в соцветии (от 2 до 9) и диаметр цветка варьирует от 1.1 до 4.6 см.
Общеизвестно, что чем выше амплитуда адаптивной изменчивости вида, тем больше у него возможностей противостоять стрессовому воздействию и тем выше его устойчивость в растительном сообществе (Rostova, 2002). Полученные данные свидетельствуют, что изменчивость признаков во всех исследуемых ценопопуляциях примерно однотипна. Наибольшей изменчивостью обладают следующие параметры: число цветоносных побегов (Cv = 19.55–62.87%), число розеточных листьев (Cv = 7.61–40.34%). Наименьшей изменчивостью – высота цветоносного побега (Cv = 9.76–21.48%), диаметр цветка (Cv = = 4.41–28.12%). Очень высокие и низкие коэффициенты вариации для перечисленных параметров обусловлены, на наш взгляд, высотным градиентом: чем выше высота над ур. м., тем вариабельнее становятся признаки.
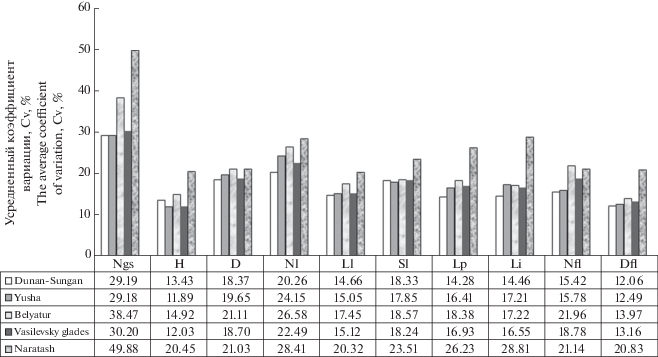
На рисунке 3 отражен график с показателями межпопуляционной изменчивости (Cv, %) морфометрических признаков в природных ценопопуляциях A. biarmiense. На межпопуляционном уровне амплитуда изменчивости исследуемых признаков A. biarmiense варьирует в следующих пределах: число цветоносных побегов – от повышенной до очень высокой (Cv = 29.18–49.88%); высота цветоносного побега – от низкой до средней (Cv = 11.89–20.45%); длина листового сегмента варьирует в средних пределах (Cv = 14.66–20.32%); диаметр цветка – от низкой до средней (Cv = 12.06–20.83%). Остальные параметры – диаметр стебля (Cv = 16.14–21.11%), число розеточных листьев (Cv = 20.26–28.56%), ширина листового сегмента (Cv = 17.27–23.51%); длина черешка (Cv = 14.28–26.23%); длина цветоножки (Cv = 14.46–28.81%); число цветков в соцветии (Cv = 15.42–21.96%) характеризуются изменчивостью от среднего до повышенного уровня. Самая высокая изменчивость по всем изучаемым параметрам наблюдается в ценопопуляции Нараташ (Cv = 20.32–49.88%) с самыми контрастными условиями, где растения сгруппированы между обломочными породами на мелкоземе с чередованием открытых участков и можжевеловых зарослей в еловом криволесье. Для большинства изучаемых параметров выявлена высокая изменчивость также в ценопопуляции Белятур (Cv = = 13.97–38.47%), где условия приближены к контрастным. Низкая изменчивость отмечена в луговых ценопопуляциях Дунан-сунган (Cv = 12.06–29.19%), Юша (Cv = 11.89–29.18%), Василевские поляны (Cv = 12.03–30.20%), произрастающих на относительно выровненных участках с менее контрастными условиями обитания.
Рис. 3.
Межпопуляционная изменчивость морфометрических параметров растений в природных ценопопуляциях (расшифровка аббревиатуры параметров в примечании к табл. 3).
Fig. 3. Interpopulation variability of morphometric parameters of plants in natural coenopopulations (see the note to Table 3 for a key to abbreviations).
Средние значения параметров различаются в каждой ценопопуляции по годам. В более благоприятном по температурным условиям 2015 г. большинство метрических и количественных признаков имели повышенные значения и наоборот, более низкие значения параметров растений отмечены в неблагоприятном 2017 г. По-видимому, растения как можно эффективнее используют ресурсы среды, путем увеличения габитуса.
Для оценки влияния погодных условий и условий экотопа ценопопуляции на морфометрические параметры A. biarmiense применен двухфакторный дисперсионный анализ. Результаты проведенного двухфакторного дисперсионного анализа массива данных по A. biarmiense представлены в табл. 4.
Таблица 4.
Оценка влияния комплекса экологических факторов на морфометрические параметры растений Anemonastrum biarmiense Table 4. Assessment of the influence of environmental factors complex on the morphometric parameters of Anemonastrum biarmiense plants
| Параметры/ Parametrs | Сила влияния факторов, %/ The power of influence of factors, % | Генеральные средние по градациям факторов/ General averages by gradation of factors | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| А | B | АB | А1 | А2 | А3 | А4 | B1 | B2 | B3 | B4 | B5 | |
| Ngs | 3.39 | 32.31** | 2.51 | 3.40 | 3.55 | 3.68 | 3.81 | 4.15 | 2.37 | 4.36 | 3.10 | 4.07 |
| H | 26.38*** | 71.71*** | 30.96*** | 45.28 | 43.51 | 37.97 | 41.18 | 46.67 | 25.99 | 40.89 | 49.86 | 46.51 |
| D | 11.27*** | 39.66** | 2.29 | 0.62 | 0.59 | 0.66 | 0.69 | 0.70 | 0.46 | 0.70 | 0.64 | 0.67 |
| Nl | 3.08 | 6.82** | 3.79* | 9.18 | 9.13 | 8.73 | 8.61 | 9.22 | 7.97 | 9.21 | 8.91 | 9.24 |
| Ll | 9.85*** | 48.48*** | 14.21*** | 7.95 | 7.42 | 7.10 | 7.49 | 8.07 | 5.40 | 7.68 | 8.34 | 7.96 |
| Sl | 16.01*** | 48.81*** | 10.73*** | 9.09 | 8.03 | 7.97 | 7.75 | 9.06 | 5.57 | 8.25 | 9.31 | 8.87 |
| Lp | 21.65*** | 67.97*** | 28.44*** | 22.68 | 19.79 | 18.72 | 19.93 | 22.96 | 11.78 | 19.36 | 25.92 | 21.39 |
| Li | 45.57*** | 70.84*** | 30.87*** | 9.27 | 8.11 | 6.69 | 7.22 | 8.36 | 4.27 | 7.55 | 9.85 | 9.09 |
| Nfl | 9.78** | 2.38* | 2.57 | 5.61 | 5.25 | 4.98 | 5.07 | 5.35 | 5.08 | 5.31 | 5.27 | 5.13 |
| Dfl | 21.11*** | 23.29*** | 9.10** | 3.59 | 3.51 | 3.08 | 3.45 | 3.62 | 3.01 | 3.39 | 3.33 | 3.70 |
Примечание. Фактор А – погодные условия года вегетации (A1 – 2015 г., A2 – 2016 г., А3 – 2017 г., А4 – 2018 г.); фактор В – условия экотопа ценопопуляции (B1 – Василевские поляны, B2 – Нараташ, B3 – Юша, В4 – Белятур, В5 – Дунан-сунган). Жирным шрифтом выделены максимальные значения параметров; * – влияние фактора достоверно при уровне значимости р < 0.05; ** – влияние фактора достоверно при уровне значимости р < 0.01; *** – влияние фактора достоверно при уровне значимости р < 0.001.
Note. Factor A – weather conditions of the growing year (A1 – 2015, A2 – 2016, A3 – 2017, A4 – 2018); factor B – ecotope conditions of coenopopulation (B1 – Vasilevskiye Glades, B2 – Naratash, B3 – Yusha, B4 – Belyatur, B5 – Dunan-Sungan). The maximum values of the parameters are shown in bold. * – the influence of the factor is reliable at a significance level p < 0.05; ** – the influence of the factor is reliable at a significance level p < 0.01; *** – the influence of the factor is reliable at a significance level p < 0.001.
Оценка влияния комплекса условий экотопа и погодных условий года вегетации и их совместного воздействия на морфометрические параметры растений A. biarmiense показала, что данные факторы статистически значимы для всех параметров, но в большей или меньшей степени (расшифровка аббревиатуры параметров в примечании к табл. 3). Более высокое в процентном соотношении воздействие фактора погодных условий года вегетации на параметры растений отмечено для некоторых линейных признаков – высота цветоносного побега, длина черешка, длина цветоножки, диаметр цветка (21.1–45.6%), а для количественных признаков влияние фактора почти не выражено (3.1–9.8%). Значимое влияние фактора погодных условий на упомянутые параметры можно объяснить с позиции лимитирующего воздействия пониженных температур дождливых и холодных вегетационных сезонов на процессы роста растений. В холодный весенний сезон 2017 г. с колебаниями минимальной и максимальной температуры от 0 до 15°С и с заморозком на почве, во время проведения измерений в луговых сообществах растения имели угнетенный вид и приземистую форму, и напротив, в 2015 г., когда не наблюдалось заморозка на почве и в более теплом 2016 г. растения были намного мощнее и выше.
Влияние условий экотопа на параметры растений выражено сильнее в сравнении с воздействием фактора погодных условий года вегетации. Значения варьируют от 2.38% до 71.71%, при этом максимальные значения выявлены для линейных признаков – высота цветоносного побега (71.71%), длина цветоножки (70.84%), длина черешка розеточного листа (67.97%), длина и ширина листового сегмента (48.48% и 48.81%). Среди количественных параметров наибольшее влияние отмечено для параметра число цветоносных побегов (32.31%). Совместное воздействие двух факторов неодинаковое, нередко они компенсируют друг друга. Изучаемые популяции, как уже говорилось, расположены в лесном и подгольцовом поясах, условия экотопа в них меняются от умеренных до экстремальных. Исследуемые параметры имеют бóльшие значения (высоту, длину и пр.) в горно-лесном поясе и меньшие – в подгольцовом поясе. Таким образом проявляется защитная приспособительная реакция растений к экстремальным условиям обитания – растения уменьшаются в размерах (миниатюризация), но при этом нормально цветут и плодоносят.
Максимальные значения средних по многим из параметров выделены для ценопопуляции Белятур, которая расположена в более благоприятных термических условиях (занимает южный прогреваемый склон в привершинной части хребта под пологом деревьев) и ценопопуляции Василевские поляны, характеризующейся условиями большей увлажненности (занимает высокотравный мезофильный луг). Минимальные значения показателей отмечены для ценопопуляции Нараташ, которая расположена в подгольцовом редколесье между скалами на высоте 1162 м над ур. м. Для разногодичных наблюдений максимальные значения факториальных средних отмечены в 2015 году, характеризующемся более дождливым и относительно теплым весенним периодом. Напротив, минимальные значения факториальных средних наблюдались в 2017 г. с наименее благоприятными погодными условиями для вегетации.
Важный показатель для оценки состояний ценопопуляций – виталитет, это характеристика жизненного состояния особей растений, выполняемая с опорой на морфометрические параметры, оценивающие рост, продукцию растений (Zlobin, 1989). Предварительно проведенный факторный и корреляционный анализы позволили выделить среди исследованных биометрических показателей A. biarmiense детерминирующий комплекс признаков: число генеративных побегов и число листьев, которые в дальнейшем были использованы для оценки виталитетного спектра ценопопуляций.
Распределение особей A. biarmiense по классам виталитета приведено в табл. 5. Среди луговых ценопопуляций следует выделить группу растений на хр. Юша, т.к. особям этой ценопопуляции свойственно устойчиво высокое жизненное состояние на протяжении четырех лет наблюдений. В ценопоппуляции Дунан-сунган жизненное состояние растений преимущественно процветающее, но становится депрессивным в неблагоприятном 2017 г. Проведение измерений морфометрических параметров ветреника в ЦП Дунан-сунган в 2017 г. состоялось 7 июня, на рис. 1 видно, что температура в этот момент времени не превышала 5°С, при этом местами наблюдался снег. В 2018 г. цветение ветреника было поздним и измерение параметров проведено уже в теплое время.
Таблица 5.
Распределение особей Anemonastrum biarmiense по классам виталитета Table 5. Distribution of Anemonastrum biarmiense plants by vitality classes
| Дата наблюдений/ Date of observation | Относительная частота размерных классов/ Relative frequency of dimensional classes | Качество популяции/ Population quality, Q | Виталитетный тип ЦП/ Vitality type of coenopopulation | ||
|---|---|---|---|---|---|
| с | b | a | |||
| Дунан-сунган/Dunan-Sungan | |||||
| 27.06.18 | 0.28 | 0.28 | 0.44 | 0.36 | Процветающая/Thriving |
| 07.06.17 | 0.40 | 0.12 | 0.48 | 0.30 | Депрессивная/Depressive |
| 31.05.16 | 0.24 | 0.24 | 0.52 | 0.38 | Процветающая/Thriving |
| 02.06.15 | 0.16 | 0.04 | 0.80 | 0.42 | -"- |
| Юша/Yusha | |||||
| 20.06.18 | 0.12 | 0.16 | 0.72 | 0.44 | Процветающая/Thriving |
| 11.06.17 | 0.15 | 0.11 | 0.84 | 0.47 | -"- |
| 01.06.16 | 0.25 | 0.28 | 0.50 | 0.39 | -"- |
| 10.06.15 | 0.28 | 0.36 | 0.36 | 0.36 | -"- |
| Белятур/Belyatur | |||||
| 15.06.18 | 0.28 | 0.36 | 0.36 | 0.36 | Процветающая/Thriving |
| 20.06.17 | 0.52 | 0.20 | 0.28 | 0.24 | Депрессивная/Depressive |
| 25.05.16 | 0.52 | 0.32 | 0.16 | 0.24 | -"- |
| 10.06.15 | 0.58 | 0.32 | 0.12 | 0.22 | -"- |
| Василевские поляны/Vasilevskiye Glades | |||||
| 20.06.18 | 0.32 | 0.20 | 0.44 | 0.32 | Равновесная/Equilibrium |
| 12.06.17 | 0.28 | 0.04 | 0.68 | 0.36 | Процветающая/Thriving |
| 01.06.16 | 0.20 | 0.48 | 0.32 | 0.40 | -"- |
| 06.06.15 | 0.52 | 0.12 | 0.36 | 0.24 | Депрессивная/Depressive |
| Нараташ/Naratash | |||||
| 21.06.18 | 0.32 | 0.36 | 0.32 | 0.34 | Равновесная/Equilibrium |
| 06.05.17 | 0.52 | 0.12 | 0.36 | 0.24 | Депрессивная/Depressive |
| 30.05.16 | 0.52 | 0.20 | 0.28 | 0.24 | -"- |
| 09.06.15 | 0.68 | 0.16 | 0.16 | 0.16 | -"- |
В ценопопуляции Василевские поляны виталитетный тип меняется от процветающей к равновесной и депрессивной. Во время проведения измерений в 2015 г. значение средней температуры зафиксировано около 7°С, и виталитетный тип ценопопуляции оказался депрессивным. В последующие годы исследований жизненное состояние растений меняется в сторону его повышения. Следовательно, жизненное состояние растений во многом зависит от погодных, прежде всего, температурных условий года вегетации и от воздействия лимитирующих факторов, свойственных местоположению на высоте 1020 м над ур. м.
Ценопопуляция растений A. biarmiense на хр. Белятур характеризуется низким жизненным состоянием. Местообитание данного вида расположено на облесенном участке склона и высоте более 900 м над ур. м., где растения ограничены в солнечном освещении, в отличие от луговых ценопопуляций. Как упоминалось ранее, растениям этой ценопопуляции свойственны увеличенные размеры листьев, побегов и соцветий, но расположены растения более разреженно. В 2018 г. популяция стала процветающей. Вероятно, во второй декаде июня растения восстановились после перенесенных заморозков этого года, и цветение уже окрепших растений продолжилось позднее обычного срока.
Ценопопуляция Нараташ локализована в поясе елового криволесья, на самой большой высоте из всех исследуемых ценопопуляций, что объясняет ежегодно угнетенное жизненное состояние растений с позиции неоптимальности их местообитания. В 2018 г. цветение продлилось до конца июня и растения к этому времени заметно окрепли, отличаясь более крупными листьями, высокими побегами, в связи с чем виталитетный тип данной ценопопуляции сменился на равновесный.
ЗАКЛЮЧЕНИЕ
Установлено, что наиболее высокие средние значения морфометрических показателей имеют растения A. biarmiense из березового редколесья на склоне хр. Белятур по причине произрастания среди лесного и опушечного крупнотравья, также из высокотравного мезофильного лугового сообщества в урочище Василевские поляны, вследствие обильного увлажнения, поступающего со сходом снега с горных вершин. В остальных горных высокотравных луговых сообществах Дунан-сунган и Юша средние значения параметров растений ниже, но число цветков и их диаметр больше, в связи с чем следует предполагать наличие здесь более благоприятных климатических условий для семенного размножения. Минимальные средние значения параметров отмечены у растений из елового криволесья на склоне хр. Нараташ в подгольцовом поясе, где сказываются менее благоприятные условия для роста растений.
Среди изученных параметров наибольшей изменчивостью обладают число цветоносных побегов, число розеточных листьев, наименьшей – высота цветоносного побега, диаметр цветка. Очень высокие и низкие коэффициенты вариации для перечисленных параметров, на наш взгляд, обусловлены расположением ценопопуляций на высотном градиенте.
На межпопуляционном уровне амплитуда изменчивости для шести из десяти признаков A. biarmiense варьирует от средней до повышенной. Самая высокая изменчивость по всем изучаемым параметрам наблюдается в ценопопуляции Нараташ с самыми контрастными условиями, а самая низкая – в луговых ценопопуляциях Дунан-сунган, Юша, Василевские поляны, расположенных на выровненных участках с более благоприятными условиями обитания.
Разногодичная внутрипопуляционная изменчивость по большинству признаков во всех ценопопуляциях слабо выражена, однако прослеживается повышение коэффициента вариации среди большинства параметров в более теплые, благоприятные по погодным условиям годы наблюдений.
Исследования показали, что условия экотопа оказывают более сильное воздействие на природные ценопопуляции, в сравнении с разногодичными погодными условиями. Приспособление к экстремальным условиям высокогорий проявляется в уменьшении размеров растений, при этом число цветков возрастает от луговых местообитаний к еловому криволесью подгольцового пояса. Неблагоприятные погодные условия года вегетации оказывают значимое влияние в период цветения растений, угнетая рост генеративных органов и, тем самым, снижают семенную продуктивность растений и ценопопуляций в целом, что отрицательно сказывается на семенном возобновлении данного вида.
В условиях достаточного увлажнения, несмотря на высокую конкуренцию со стороны других видов сообщества, в ценопопуляциях сохраняется высокий уровень жизненности отдельных особей. На фоне ухудшения погодных условий процессы роста особей A. biarmiense значительно подавляются. Среди луговых ценопопуляций в ЦП Юша отмечено преобладание особей высшего класса виталитета ежегодно, и она отнесена к категории процветающих. В остальных двух луговых ценопопуляциях данная категория выявлена с меньшим постоянством, однако особи высшего класса встречены почти ежегодно. Ценопопуляция растений A. biarmiense на хр. Белятур отличается более низким жизненным состоянием от предыдущих популяций. Ценопопуляция Нараташ расположена в подгольцовом поясе, что объясняет ежегодное угнетенное жизненное состояние растений с позиции неблагоприятности их местообитания.
Список литературы
Abdaladze O., Nakhutsrishvili G., Batsatsashvili K., Gigauri K., Jolokhava T., Mikeladze G. 2015. Sensitive Alpine Plant Communities to the Global Environmental Changes (Kazbegi Region, the Central Great Caucasus). – Am. J. Environ. Prot. 4 (3–1): 93–100. https://doi.org/10.11648/j.ajep.s.2015040301.25
[Bobretsova] Бобрецова И.А. 2002. Популяционная биология ветреницы пермской в Печоро-Илычском заповеднике. – В сб.: Материалы док. “Актуальные проблемы биологии и экологии”. Сыктывкар. С. 123–124.
[Degteva] Дегтева С.В. 2008. Сообщества травянистых растений Печоро-Илычского заповедника. – Вестник Института биологии Коми НЦ УрО РАН. 9 (131): 18–20.
Dutton B.E., Reveal J.L., Keener C.S., Gandhi K.N. 1995. Proposal to conserve the orthography of Anemone narcissiflora L. (Anemonastrum narcissiflorum; Ranunculaceae). – Taxon. 44 (3): 421–422. https://doi.org/10.2307/1223419
[Endemic plants…] Эндемичные растения Урала во флоре Свердловской области. 2013. Екатеринбург. 610 с.
[Fiziko-geograficheskoe…] Физико-географическое районирование Башкирской АССР. 1964. Уфа. 210 с.
[Golubev] Голубев В.Н. 1962. Основы биоморфологии травянистых растений Центральной лесостепи. Воронеж. 511 с.
[Gorchakovsky, Shurova] Горчаковский П.Л., Шурова Е.А. 1982. Редкие и исчезающие растения Урала и Приуралья. М. 208 с.
[Gorichev] Горичев Ю.П. 2008. Природные особенности Южно-Уральского государственного природного заповедника. – В сб.: “Труды Южно-Уральского государственного природного заповедника”. Уфа. С. 13–56.
Hoot S.B., Meyer K.M., Manning J.C. 2012. Phylogeny and reclassification of Anemone (Ranunculaceae), with an emphasis on Austral species. – Syst. Bot. 37 (1): 139–152. https://doi.org/10.1600/036364412X616729
[Kadilnikov] Кадильников И.П. 1966. Физико-географическое районирование Южного Урала. – Труды МОИП. 18: 107–120.
[Karimova et al.] Каримова О.А., Жигунов О.Ю., Голованов Я.М., Абрамова Л.М. 2013. Характеристика ценопопуляций редких горно-скальных видов в Зауралье Республики Башкортостан. – Вестник Томского гос. ун-та. 2 (22): 70–83.
[Krasnaya…] Красная книга Свердловской области: животные, растения, грибы. 2018. Екатеринбург. 450 с.
[Krasnaya…] Красная книга Республики Коми. 2019. Сыктывкар. 768 с.
[Krasnaya…] Красная книга Ханты-Мансийского автономного округа – Югры: животные, растения, грибы. 2013. Екатеринбург. 460 с.
[Krasnaya…] Красная книга Ямало-Ненецкого автономного округа: животные, растения, грибы. 2010. Екатеринбург. 308 с.
[Kucherov et al.] Кучеров Е.В., Мулдашев А.А., Галеева А.Х. 1987. Охрана редких видов растений на Южном Урале. М. 204 с.
[Letopis’…] Летопись природы Южно-Уральского государственного природного заповедника. 2016. Белорецк. С. 12–13; 2017, С. 13–14; 2018, С. 12–14; 2019, С.1 2–13.
[Malinovs’kij] Малиновський К.А. 2002. Рослинні угруповання високогір’я Українських. Ужгород. 244 с.
[Mamaev] Мамаев С.А. 1975. Основные принципы методики исследования внутривидовой изменчивости древесных растений. – В сб.: Индивидуальная и эколого-географическая изменчивость растений. Свердловск. С. 3–14.
[Mamaev, Knyazev] Мамаев С.А., Князев М.С. 1995. Ветреницы в природе и культуре. Екатеринбург. 18 с.
[Mineeva] Минеева О.Н. 1985. Анатомо-морфологические особенности ветреницы пермской на Северном Урале. – В сб.: Растительный мир Урала и его антропогенные изменения. Свердловск. С. 63–70.
[Mirkin et al.] Миркин Б.М., Мулдашев А.А., Мартыненко В.Б., Маслова Н.В. 2004. Охрана биологического разнообразия Башкортостана: современное состояние исследований и их перспективы. – Вестник Академии наук Республики Башкортостан. 9 (1): 38–47.
Mosyakin S.L. 2016. Nomenclatural notes on North American taxa of Anemonastrum and Pulsatilla (Ranunculaceae), with comments on the circumscription of Anemone and related genera. – Phytoneuron. 79: 1–12.
Mosyakin S.L. 2018. Further new combinations in Anemonastrum (Ranunculaceae) for Asian and North American taxa. – Phytoneuron. 55: 1–11.
Mosyakin S.L., Lange P.J. 2018. Anemonastrum tenuicaule and A. antucense (Ranunculaceae), new combinations for a New Zealand endemic species and its South American relative. – PhytoKeys. 99: 107–124. https://doi.org/10.3897 / phytokeys.99.26489
[Plotnikova] Плотникова И.А. 2009. Ветренник пермский. – В кн.: Биология и экология редких растений Республики Коми. Екатеринбург. С. 86–118.
[Poletaeva] Полетаева И.И. 2015. Структура популяций Аnemonastrum biarmiense в горной части Печоро-Илычского заповедника. – В сб.: Проблемы изучения и сохранения растительного мира Восточной Фенноскандии. Апатиты. С. 75–76.
[Rostova] Ростова Н.С. 2002. Корреляции: структура и изменчивость. СПб. 307 с.
[Serebryakov] Серебряков И.Г. 1959. Типы развития побегов у травянистых многолетников и факторы их формирования. – Вопросы биологии растений. 100 (5): 3–27.
[Shirokih] Широких П.С., Мартыненко В.Б., Баишева Э.З., Бикбаев И.Г. 2018. О новой ассоциации лугов на вырубках светлохвойных бореальных лесов Южного Урала. – Известия Уфимского научного центра РАН 3: 67–78.
[Tzvelev] Цвелев Н.Н. 2001. Род Anemonastrum Holub – Ветреник. – Флора Восточной Европы. 10: 77–79.
[Valuiskikh, Kanev] Валуйских О.Е., Канев В.А. 2019. Новые сведения о распространении редких видов сосудистых растений на хребте Поясовый камень (Северный Урал). – Бот. журн. 104 (9): 1475–1482. https://doi.org/10.1134/S000681361909014X
[Vernigor] Вернигор Р.А. 1981. Изменчивость и структура популяций высокогорного уральского эндемика ветреницы пермской (Anemone biarmiensis Jus.): Дис. … канд. биол. наук. Свердловск. 193 с.
[Viktorov] Викторов В.П. 2018. Стратегии сохранения редких видов растений. – Вестник Тверского гос. ун-та. 3: 106–129.
[Volkov] Волков И.В. 2007. Биоморфологические адаптации высокогорных растений. Томск. 412 с.
[Yamalov et al.] Ямалов С.М., Мартыненко В.Б., Абрамова Л.М., Голуб В.Б., Баишева Э.З., Баянов А.В. 2012. Продромус растительных сообществ Республики Башкортостан. Уфа. 100 с.
[Yusupova et al.] Юсупова О.В., Абрамова Л.М., Каримова О.В. 2016. К биологии и экологии эндема Аnemonastrum biarmiense (Juz.) Holub в Южно-Уральском государственном природном заповеднике. – Вестник Пермского ун-та. 3: 222–228.
[Yusupova et al.] Юсупова О.В., Абрамова Л.М., Мустафина А.Н. 2017. К биологии и экологии эндема Anemonastrum biarmiense (Juz.) Holub на Южном Урале. – Slovak international scientific journal. 7 (7): 7–13.
[Yusupova et al.] Юсупова О.В., Абрамова Л.М., Юсупов И.Р. 2018. Оценка влияния комплекса экологических факторов на морфометрические параметры эндемичного вида Anemonastrum biarmiense (Juz.) Holub в ценопопуляциях горного Урала. – Известия Уфимского научного центра РАН. 4: 65–70.
[Zaytsev] Зайцев Г.Н. 1990. Математика в экспериментальной биологии. М. 296 с.
[Zlobin] Злобин Ю.А. 1989. Принципы и методы изучения ценотических популяций растений: учебно-методическое пособие. Казань. 146 с.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал