

Ботанический журнал, 2021, T. 106, № 1, стр. 22-42
ДИНАМИКА МИКОРИЗООБРАЗОВАНИЯ У НЕКОТОРЫХ ВИДОВ РАСТЕНИЙ В ХОДЕ ВОССТАНОВИТЕЛЬНОЙ СУКЦЕССИИ НА ПЕСЧАНЫХ КАРЬЕРАХ (ЛЕНИНГРАДСКАЯ ОБЛАСТЬ)
А. О. Горбунова 1, 2, *, О. И. Сумина 1, **
1 Санкт-Петербургский государственный университет
199034 Санкт-Петербург, Университетская наб., 7/9, Россия
2 Всероссийский научно-исследовательский институт сельскохозяйственной микробиологии
196608 Пушкин, Санкт-Петербург, шоссе Подбельского, 3, Россия
* E-mail: st057575@student.spbu.ru
** E-mail: o.sumina@spbu.ru
Поступила в редакцию 11.05.2020
После доработки 15.09.2020
Принята к публикации 29.09.2020
Аннотация
Изучены изменения в микоризообразовании у 5 видов трав (Agrostis capillaris, Artemisia vulgaris, Chamaenerion angustifolium, Deschampsia cespitosa, Tussilago farfara), обычных в сообществах последовательных стадий восстановительной сукцессии. На двух разновозрастных песчаных карьерах Ленинградской области (Всеволожский район) описаны растительные сообщества четырех стадий сукцессии: пионерной, злаковой, кустарниковой и лесной. Впервые проведено сравнение ценотической роли и виталитета изученных видов с параметрами (встречаемостью и интенсивностью) микоризации их корней. В корнях всех видов имеется арбускулярная микориза в форме несептированного мицелия, арбускул и везикул. У каждого вида ее параметры наиболее сильно варьируют в пионерных сообществах. При переходе к злаковой стадии наблюдается слабо выраженная тенденция роста интенсивности микоризации, одновременно увеличивается проективное покрытие видов и, в некоторых случаях, виталитет. В то же время, высокая степень микоризации не всегда обеспечивает высокие виталитет и ценотическую роль вида. На лесной стадии сукцессии параметры микоризации снижаются у всех пяти видов. Вероятно, на песчаных карьерах, особенно на пионерной стадии сукцессии, наиболее сильное влияние на жизненность и ценотическую роль травянистых растений оказывают богатство субстрата и другие абиотические факторы.
Более 90% семейств сосудистых растений формируют арбускулярную микоризу (АМ) с грибами подотдела Glomeromycotina отдела Mucoromycota (Spatafora et al., 2016). Роль АМ в фитоценозе огромна, она может влиять как на рост и конкурентоспособность отдельных особей растений, так и на биоразнообразие и продуктивность всего растительного сообщества. Растения-микоризообразователи доминируют почти во всех экосистемах, за исключением некоторых сообществ ранних стадий первичных сукцессий, интенсивно возделываемых пахотных земель, сильно засоленных почв и почв с чрезвычайно низким содержанием фосфора, а также экстремально холодных областей (Öpik et al., 2006; Lambers et al., 2008).
Интерес к изучению микоризных связей растений в сообществах восстановительных сукцессий объясняется тем, что микоризные взаимодействия обуславливают способность растений к конкуренции и преодолению неблагоприятных эдафических условий (van der Heijden et al., 1998; Lambers et al., 2008). Таким образом, наличие микоризы может быть важным фактором, способствующим успешному освоению растениями свободных минеральных субстратов.
Микоризные симбионты могут существенно улучшить жизненность растений-первопоселенцев, так как грибы поглощают питательные вещества даже из твердых неорганических частиц очень бедных почв. Наличие микоризы является важным фактором адаптации растений в условиях недостатка азота, фосфора и других элементов минерального питания, неблагоприятных водного и воздушного режимов, обедненности субстрата органическими формами углерода (Aikio, 2000; Lukina, Ryazanova, 2012). Однако в пионерных сообществах часто доминируют виды, не образующие микоризного симбиоза: однолетники-эксплеренты из семейств Chenopodiaceae, Brassicaceae, Caryophyllaceae и Polygonaceae (Smith, Read, 2008). Показано, что пионерные виды растений по-разному восприимчивы к микоризному воздействию (Daft, Nicolson, 1974; Rydlová, Vosátka, 2001; Püschel et al., 2007). Низкое обилие растений, образующих микоризу на инициальной стадии сукцессии, отчасти связано с тем, что колонизация грибами проростков растений затруднена из-за отсутствия развитой мицелиальной сети, для поддержания функционирования которой необходимо наличие достаточно плотного и постоянного растительного покрова (van der Heijden et al., 2015). В пионерных сообществах растения сравнительно слабо взаимодействуют между собой, их надземные части не соприкасаются (конкуренция за свет отсутствует), хотя подземные органы могут перекрываться (но конкуренция за минеральное питание еще недостаточно интенсивна). Ф. Ривз с соавторами (Reeves et al., 1979) выдвигали гипотезу, что пионерные немикотрофные растения слабо конкурентоспособны и быстро замещаются видами, ассоциированными с грибами, как только инокулюм микоризных грибов попадает на зарастающий участок.
В зарастании антропогенно-нарушенных территорий, как и на начальных стадиях некоторых естественных сукцессий, активно участвуют немикоризные виды растений, а доля облигатных микотрофов оказывается пониженной (Veselkin, Betekhtina, 2011), но с течением сукцессии ситуация обычно меняется на противоположную (Allen, 1988; Gemma, Koske, 1990; Cázares et al., 2005). Конкурентные взаимоотношения между растениями с развитием фитоценоза усиливаются, первыми из него выпадают немикотрофные виды-эксплеренты и факультативные микотрофы, которые можно считать низко специализированными видами. На более поздних этапах сукцессии условия стабилизируются, что выражается в близком соотношении видов разного микоризного статуса. В работах Д.В. Веселкина с соавторами (Veselkin, Betekhtina, 2011; Veselkin, 2012a, b; Veselkin et al., 2015) было показано, что по мере приближения сукцессионных сообществ по составу и структуре к зональным, в них возрастает участие облигатно микоризных видов и, соответственно, снижаются доли факультативно микоризных и немикоризных видов. Такие данные подводят к выводу о том, что АМ грибы, вероятно, способствуют ходу сукцессии, но до сих пор прямых доказательств такой их роли в природных экосистемах очень мало (Smith, Read, 2008).
АМ-симбионты потенциально могут влиять на распределение ресурсов в растении-хозяине, что в свою очередь отражается на развитии надземных частей растений (и на их взаимодействии с другими видами сообщества). При положительном влиянии АМ на надземную биомассу растений (Wolfe et al., 2005) особи лучше развиваются, имеют более высокий виталитет и могут с большей вероятностью стать доминантами или содоминантами в сообществах.
Целью нашей работы было выяснение роли арбускулярной микоризы как фактора, способного улучшить жизненность и ценотический статус видов в растительных сообществах разных стадий восстановительной сукцессии.
МАТЕРИАЛЫ И МЕТОДЫ
Сбор материала проведен в первой половине июля 2018 и 2019 гг. на двух разновозрастных песчаных карьерах, расположенных во Всеволожском районе Ленинградской области. Карьер Калелово приостановил свою работу в декабре 2018 г. А.О. Горбуновой были описаны пионерные растительные сообщества (пионерная стадия), сообщества с преобладанием злаков (злаковая стадия) и сообщества с подростом деревьев и кустарников (кустарниковая стадия). Все они были приурочены к донным участкам карьера с умеренным увлажнением. Формирование растительности шло по типу первичной сукцессии.
Песчаный карьер Кузьмолово закончил работу в 2013 г. На нем описаны сообщества злаковой и более поздних стадий сукцессии, включая лесную стадию. Сообщества последней встречались в периферической части карьера, где мелколесные заросли Alnus incana11 формировались на вскрыше, содержащей перемещенную почву и зачатки растений, так что сукцессия на этих участках шла по типу вторичной.
Таблица 1.
Основные характеристики сообществ разных стадий сукцессии (фрагмент сводной таблицы описаний пробных площадок) Table 1. The main characteristics of plant communities of different stages of succession (part of the table with relevés of the studied plots)
| Название карьера/Quarry name | Калелово Kalelovo | Ку Ku |
Калелово Kalelovo | Кузьмолово Kuzmolovo | Кa Ka |
Кузьмолово Kuzmolovo | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Стадия сукцессии/Stage of succession | Пс | Пс | Пс | Пс | Пс | Пс | Пс | Зс | Зс | Зс | Зс | Кс | Кс | Кс | Кс | Лс | Лс |
| Номер площадки/Plot number | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 |
| Общее проективное покрытие, % Total cover, % |
1 | 4 | 5 | 7 | 15 | 17 | 7 | 40 | 60 | 60 | 85 | 80 | 60 | 85 | 70 | 80 | 95 |
| Сомкнутость крон древесного яруса, % Density of tree stand canopy, % |
- | <1 | <1 | - | - | <1 | 3 | <1 | 4 | 4 | 1 | 10 | 40 | 60 | 45 | 80 | 25 |
| ПП травяного яруса, % Coverage of grass layer, % | 1 | 4 | 1 | 7 | 13 | 17 | 2 | 30 | 37 | 51 | 80 | 2 | 3 | 17 | 27 | 70 | 87 |
| ПП мохового яруса, % Coverage of moss layer, % | - | - | 4 | <1 | 5 | <1 | 2 | 10 | 20 | 20 | 45 | 75 | 23 | 35 | 2 | 15 | 45 |
| Число видов трав Number of herbaceous species | 10 | 6 | 9 | 11 | 9 | 20 | 20 | 12 | 17 | 24 | 32 | 15 | 6 | 15 | 32 | 23 | 26 |
| Число видов древесных Number of tree species | 0 | 2 | 1 | 0 | 0 | 1 | 7 | 2 | 5 | 3 | 2 | 6 | 7 | 8 | 2 | 7 | 3 |
| Число видов мхов Number of moss species | 0 | 0 | 3 | 1 | 3 | 2 | 2 | 1 | 1 | 3 | 4 | 2 | 6 | 3 | 4 | 3 | 2 |
| Общее число видов Total number of species | 10 | 8 | 13 | 12 | 12 | 23 | 29 | 15 | 23 | 30 | 38 | 23 | 19 | 26 | 38 | 33 | 31 |
| Травы, в том числе: Herbs and grasses, including: |
|||||||||||||||||
| модельные виды/model species | |||||||||||||||||
| Tussilago farfara L. | ++ | ++ | +r | 3 | 3 | ++ | ++ | ++ | +1 | ++ | 2 | +1 | |||||
| Chamaenerion angustifolium (L.) Scop. | ++ | 3 | ++ | 4 | +1 | ++ | 1 | 5 | 3 | ++ | +1 | 3 | ++ | 1 | |||
| Deschampsia cespitosa (L.) P. Beauv. | +1 | 2 | 1 | ++ | 3 | 3 | +r | 20 | ++ | +1 | 8 | 5 | 45 | ||||
| Agrostis capillaris L. | ++ | ++ | 7 | 5 | ++ | 23 | 20 | 4 | 25 | ++ | +1 | 6 | 13 | ++ | ++ | ||
| Artemisia vulgaris L. | 3 | ++ | +1 | 5 | 1 | ++ | +1 | ++ | +1 | 3 | |||||||
| виды с максимальным ПП в описании: species with the highest percentage cover in relevé: |
|||||||||||||||||
| Galeopsis tetrahit L. | +r | ++ | 2 | ++ | +1 | +1 | ++ | ||||||||||
| Calamagrostis epigeios (L.) Roth | 3 | 25 | ++ | 2 | |||||||||||||
| Chenopodium album L. | 3 | ++ | |||||||||||||||
| Luzula multiflora (Ehrh.) Lej. | +1 | 1 | ++ | ||||||||||||||
| Stellaria holostea L. | 40 | ||||||||||||||||
| Деревья и кустарники: Trees and shrubs: |
|||||||||||||||||
| Betula pendula Roth/Betula pubescens Ehrh. | ++ | ++ | 1 | ++ | 3 | 2 | 17 | 10 | |||||||||
| Pinus sylvestris L. | +r | ++ | ++ | +1 | 5 | 3 | |||||||||||
| Salix caprea L. | +1 | +1 | 4 | +1 | 5 | 7 | 4 | ||||||||||
| Salix myrsinifolia Salisb. | ++ | ++ | ++ | 2 | 15 | +r | 3 | ||||||||||
| Alnus incana (L.) Moench | 1 | ++ | +r | +1 | ++ | 25 | 45 | 80 | 23 | ||||||||
| Populus tremula L. | +r | ++ | 5 | 13 | 15 | ||||||||||||
| Rubus idaeus L. | ++ | 4 | 3 | ||||||||||||||
| Sorbus aucuparia L. | ++ | +1 | 3 | ||||||||||||||
| Sambucus racemosa L. | 4 | ||||||||||||||||
| Мхи:/Mosses: | |||||||||||||||||
| Pogonatum urnigerum (Hedw.) P. Beauv. | ++ | ++ | 2 | ++ | 10 | 20 | 1 | ||||||||||
| Ceratodon purpureus (Hedw.) Brid. | 4 | 3 | ++ | 1 | 15 | 10 | 70 | 17 | 34 | 1 | |||||||
| Polytrichum commune Hedw. | 1 | 10 | 5 | ||||||||||||||
| Bryum lonchocaulon Müll. Hal. | ++ | ||||||||||||||||
| Polytrichum juniperinum Hedw. | 7 | 3 | +1 | ||||||||||||||
| Oxyrrhynchium hians (Hedw.) Loeske | 25 | 35 | |||||||||||||||
| Sciuro-hypnum oedipodium (Mitt.) Ignatov et Huttunen | 3 | 10 | |||||||||||||||
| Atrichum undulatum (Hedw.) P. Beauv. | 3 | 5 | |||||||||||||||
| Rhytidiastrum squarrosum (Hedw.) Ignatov et Ignatova | 7 | ||||||||||||||||
В сообществах разных стадий восстановительной сукцессии, на участках с относительно однородным растительным покровом были заложены пробные площади 5 × 5 м (Sumina, 2013). Все они размещались на ровных поверхностях (Калелово) или уклонах крутизной не более 25° (Кузьмолово). Всего было описано 17 площадей: 7 – для пионерной стадии, 4 – для злаковой, 4 – для кустарниковой и 2 – для лесной, сообщества которой встречаются редко. Сообщества пионерной стадии были слабо сомкнуты, общее проективное покрытие (ОПП) не превышало 20%. На злаковой стадии в сообществах, преимущественно образованных травами, господствовали злаки. На кустарниковой – верхний ярус сообществ был образован кустарниками и подростом деревьев. Заключительная – лесная стадия – была представлена молодняками лесного типа растительности.
В ходе геоботанического описания отмечали физиономические и другие характеристики сообщества: аспект, максимальную и господствующую высоту растений каждого яруса, характер размещения растений по площади, ОПП растительности (в процентах), проективное покрытие (ПП) по группам (травы, кустарники, деревья, мхи и лишайники), видовой состав и покрытие каждого вида. При ПП вида менее 1% использовали модифицированные (Barkman, 1991) оценки шкалы обилия-покрытия Браун-Бланке (Westhoff, van der Maarel, 1978): “+r” – 1–2 особи на пробную площадь, “++” – несколько особей, “+1” – до 100 особей, занимающих менее 1% площади.
На большинстве пробных площадей (5 – пионерной стадии, 3 – злаковой, 3 – кустарниковой и 2 – лесной) и на участке незадернованного голого грунта делали почвенные прикопки и отбирали пробы с глубин 5 и 20 см, подстилку или опад (при наличии). Почвенные образцы высушивали и в камеральный период определяли кислотность (pH в солевом растворе KCl, GOST 26483-85, 1985), содержание органического вещества (GOST 26213-91, 1993), общего азота (GOST 26107-84, 1984) и доступного фосфора (GOST P 54650-2011, 2013).
Пять видов травянистых растений (Agrostis ca-pillaris, Artemisia vulgaris, Chamaenerion angustifolium, Deschampsia cespitosa, Tussilago farfara), согласно литературным данным (Wang, Qiu, 2006; Akhmetzhanova et al., 2012) образующих АМ, были избраны в качестве модельных. Они присутствовали в сообществах практически всех стадий сукцессии и встречались на пробных площадях в числе более 5 особей. По каждому модельному виду все данные собирали в 10 сообществах, где он был наиболее обилен.
Оценка ценотического статуса модельных видов дана по шкале господства (Ipatov, Mirin, 2008) на основании их относительного покрытия. Относительное покрытие (ОП) вида выражается в процентах от общего проективного покрытия яруса. Шкала включает следующие классы: вид присутствует (ОП < 1%), является редким (ОП = = 1–5%) или “наполнителем” (ОП = 5%–1/3), согосподствует (ОП = 1/3–2/3), господствует (ОП = 2/3 и более), где 1/3 и 2/3 – доля от общего проективного покрытия травяного яруса.
Для определения жизненности (виталитета)22 использовали шкалу Ж. Браун-Бланке и И. Павийяра, дополненную А.А. Гроссгеймом (Westhoff, van der Maarel, 1978), включающую следующие балльные оценки: 1 – прорастание, отсутствие у особи развития вегетативных органов; 2 – особи ослаблены, недостаточно вегетативно развиты и не проходят полностью весь жизненный цикл; 3 – особи хорошо вегетативно развиты, но не проходят весь жизненный цикл; 4 – вегетативное развитие хорошее, цветение и плодоношение обильное; 5 – пышное развитие и повышенные плодоношение и цветение.
Для оценки степени микоризации растений собирали особи модельных видов: объем выборки для каждого из них составлял не менее 5 особей с пробной площади. Для каждой особи отмечали высоту, балл виталитета и дополнительные сведения (например, присутствие фитофагов). При сборе материала предпочтение отдавали особям, соответствующим состоянию большинства растений данного вида в сообществе, как правило, это были экземпляры (у корневищных видов – парциальные побеги), находящиеся в стадии цветения/плодоношения. При отсутствии таковых (например, у Tussilago farfara, цветущей ранней весной) собирали хорошо развитые вегетирующие парциальные побеги.
У собранных растений в свежем состоянии отделяли корни и помещали в крафт-пакеты на просушку. После высушивания отбирали тонкие неодревесневшие корни на анализ.
При подготовке микропрепаратов микоризованных корней пользовались методикой мацерации и окрашивания корней растений в трипановом синем (Phillips, Hayman, 1970). Мацерация проводилась в 10% KOH на водяной бане в течение 1 часа. Для окрашивания применяли смесь из 10% раствора молочной кислоты, глицерина, дистиллированной воды и красителя “трипановый синий” в соотношениях 62 мл: 62 мл: 875 мл: 0.3 г – соответственно.
Для расчета показателей микоризации использовали метод световой микроскопии, предложенный A. Trouvelot (Trouvelot et al., 1986). У 5 особей каждого вида брали усредненную выборку из фрагментов корней длиной около 1 см. От 50 до 80 таких фрагментов плотно укладывали на покрытое глицерином предметное стекло и накрывали другим предметным стеклом. Так подготавливали 3 предметных стекла. На каждом предметном стекле исследовали 100 полей зрения, визуально определяя процент корня (как объем, спроецированный на площадь), занятый структурами АМ грибов. Баллы присваивали трем классам степени обилия структур АМ:
1) класс степени микоризации (от 1 до 5 баллов): 1 – 0–1% микоризы в корне, 2 – 2–10%, 3 – 11–50%, 4 – 51–90%, 5 – 91–100%;
2) класс степени обилия арбускул (от 1 до 3 баллов): 1 – 1–20% арбускул в микоризованной части корня (обычно 1–4 арбускулы), 2 – 21–80%, 3 – 81–100%;
3) класс степени обилия везикул (от 1 до 3 баллов): 1 – 1–20% везикул в микоризованной части корня (обычно 1–4 везикулы), 2 – 21–80%, 3 – 81–100%.
Балльные оценки заносили в компьютерную программу вычисления индексов микоризации корней растений на базе Microsoft Excel (Vorobiev et al., 2016) и по формулам, приведенным в литературе (Trouvelot et al., 1986; Mycorrhiza Manual, 2001; Yurkov et al., 2010; Yurkov, Semenov, 2019), делали расчет следующих параметров микоризации:
встречаемость микоризной инфекции в исследуемых фрагментах корней (F, %);
интенсивность микоризной инфекции во всех исследованных фрагментах корней (M, %);
интенсивность микоризной инфекции в микоризованных фрагментах корней (m, %);
обилие арбускул во всех исследованных фрагментах корней (A, %);
обилие везикул во всех исследованных фрагментах корней (V, %).
Статистический анализ проведен в программе Microsoft Excel 2016 с помощью пакета Real Statistics. Проведен однофакторный дисперсионный анализ (ANOVA), различия в параметрах микоризации на разных стадиях сукцессии сравнивали, используя критерий достоверно значимой разности Тьюки (P < 0.05).
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Стадии восстановительной сукцессии
Пионерная стадия. Сообщества начала сукцессии разнообразны, поэтому описаны на большем числе пробных площадей (далее – площадки), которые располагались преимущественно на карьере Калелово, где сделано 6 описаний (табл. 1, № 1–6) на участках с сомкнутостью растительности от 1 до 17%. На 4 площадках ОПП не превышало 7%, общее число видов – 8–13. Первыми на песчаном субстрате карьера поселяются растения, как правило, не образующие микоризный симбиоз (Galeopsis tetrahit, Chenopodium album, Persicaria scabra (Moench) Moldenke и др.). В слабосомкнутых пионерных сообществах наибольшее ПП (никогда не превышавшее 3%) имели травы: два однолетника Chenopodium album и Galeopsis tetrahit или один из трех видов – Alopecurus aequalis Sobol., Chamaenerion angustifolium, Deschampsia cespitosa. В последнем случае в сообществе с покрытием 4% встречался мох Ceratodon purpureus.
На двух площадках, где ОПП было выше (15–17%), общее число видов различалось почти вдвое: 12 и 23. В обоих сообществах максимальное ПП было у Agrostis capillaris (5–7%), с покрытием более 1% встречались Tussilago farfara (3%) и Deschampsia cespitosa (1–2%). Кроме них в сообществе с бóльшим числом видов значимое ПП имели Chamaenerion angustifolium (4%) и Artemisia vulgaris (3%).
На карьере Кузьмолово описана одна площадка (табл. 1, № 7) со слабосомкнутой растительностью (ОПП 7%) и общим числом видов – 29, причем ПП каждого вида не превышало 1%. Наиболее обильны были: Betula pubescens (1%), Alnus incana (1%), Salix caprea (+1), Chamaenerion angustifolium (+1), Luzula multiflora (+1), а также мхи Ceratodon purpureus (1%) и Polytrichum commune (1%).
В целом для пионерных сообществ характерно участие трав, растущих на значительном удалении друг от друга и не образующих единого яруса. Высота их варьирует от 2 до 90 см. Немногие виды древесных представлены отдельными всходами и редким подростом, его господствующая высота у большинства деревьев составляла 15 см, у подроста березы – 30 см. Напочвенный ярус не выражен, так как покрытие мхов нигде не превышало 5%.
Грунты на глубинах 5 и 20 см характеризуются кислой реакцией: рН солевой (рНKCl) 4.48–4.86 и 4.27–5.26, соответственно (табл. 2); низким уровнем содержания общего углерода, азота и отчасти – фосфора (сравнительно бóльшее количество последнего отмечено на площадке № 7 в Кузьмолово).
Таблица 2.
Агрохимические характеристики грунтов пробных площадок Table 2. Agrochemical characteristics of grounds on studied plots
| Название карьера Name of quarry | Стадия сукцессии Stage of succession | № площадки Plot number |
рНKCl | P подвижный
в P2O5, мг/100 г Mobile P inP2O5, mg/100 g |
С общий, % Total C, % |
N общий, % Total N, % |
||||
|---|---|---|---|---|---|---|---|---|---|---|
| 5 см/5 cm | 20 см/20 cm | 5 см/5 cm | 20 см/20 cm | 5 см/5 cm | 20 см/20 cm | 5 см/5 cm | 20 см/20 cm | |||
| Калелово Kalelovo |
Голый грунт Bare ground |
0 | 4.37 | 4.04 | 7.5 | 6.3 | 0.065 | 0.076 | 0.020 | 0.024 |
| Пс | 1 | 4.67 | 4.88 | 4.0 | 4.5 | 0.153 | 0.117 | 0.029 | 0.027 | |
| Пс | 3 | 4.73 | 5.26 | 6.1 | 3.7 | 0.071 | 0.078 | 0.025 | 0.030 | |
| Пс | 4 | 4.86 | 5.21 | 4.5 | 4.6 | 0.086 | 0.030 | 0.028 | 0.029 | |
| Пс | 6 | 4.51 | 4.27 | 4.4 | 10.5 | 0.064 | 0.078 | 0.026 | 0.029 | |
| Кузьмолово Kuzmolovo |
Пс | 7 | 4.48 | 4.57 | 21.5 | 19.7 | 0.170 | 0.100 | 0.051 | 0.048 |
| Калелово Kalelovo |
Зс | 8 | 4.11 | 4.65 | 9.4 | 5.7 | 0.203 | 0.170 | 0.030 | 0.026 |
| Кузьмолово Kuzmolovo |
Зс | 10 | 4.31 | 4.31 | 15.1 | 14.0 | 0.320 | 0.750 | 0.063 | 0.100 |
| Зс | 11 | 4.17 | 4.24 | 24.5 | 51.7 | 1.570 | 1.230 | 0.156 | 0.146 | |
| Кс | 12 | 4.34 | 4.80 | 15.8 | 19.6 | 0.160 | 0.100 | 0.049 | 0.024 | |
| Кс | 13* | 2.83 | 2.55 | 13.0 | 42.2 | 0.370 | 0.360 | 0.065 | 0.063 | |
| Кс | 14** | 3.70 | 3.77 | 8.0 | 9.5 | 0.260 | 0.180 | 0.065 | 0.056 | |
| Лс | 16 | 3.61 | 3.70 | 7.9 | 6.1 | 1.270 | 0.770 | 0.147 | 0.104 | |
| Лс | 17 | 3.95 | 3.84 | 6.5 | 6.4 | 1.460 | 1.850 | 0.192 | 0.207 | |
В пионерных сообществах встречаются все модельные виды: Tussilago farfara (отмечен на 6 площадках из 7 с ПП от +r до 3%), Chamaenerion angustifolium (на 5 площадках, ПП ++ – 4%), Agrostis capillaris (на 5 площадках, ПП ++ – 7%), Deschampsia cespitosa (на 4 площадках, ПП ++ – 2%), Artemisia vulgaris (на 2 площадках, ПП ++ –3%).
Злаковая стадия. Сообщества второй стадии сукцессии описаны на 4 площадках в карьерах Калелово (табл. 1, № 8–9) и Кузьмолово (табл. 1, № 10–11). В Калелово ОПП растительности составляло 40 и 60%, общее число видов – 15 и 23. В сообществах доминировал Agrostis capillaris (20–23%), в напочвенном покрове – Pogonatum urnigerum (10–20%). ПП прочих видов не превышало 5%. В Кузьмолово на площадках сомкнутость растительности и общее число видов были больше: 60 и 85%; 30 и 38, соответственно. В одном сообществе доминировали Calamagrostis epigeios (25%) и мхи Ceratodon purpureus (15%) и Polytrichum commune (10%). В другом – Agrostis capillaris (25%), Deschampsia cespitosa (20%) и мхи Oxyrrhynchium hians (25%) и Ceratodon purpureus (10%).
В сообществах данной стадии преобладают злаки. Увеличиваются господствующая высота растений (до 30–50 см) и покрытие мхов; травяной и моховой ярусы частично перекрываются. На карьере Калелово травяной ярус сообществ еще фрагментарен, подъярусы не выражены, тогда как в Кузьмолово выделяются 3 подъяруса: высотой от 56 до 110 см (Chamaenerion angustifolium, Deschampsia cespitosa, Agrostis capillaris); от 31 до 55 см (Artemisia vulgaris, Leucanthemum vulgare Lam., Lathyrus pratensis L.), от 2 до 30 см (Achillea millefolium L., Equisetum arvense L., Tussilago farfara). Древесно-кустарниковый подрост единичен.
Грунты на глубинах 5 и 20 см кислые: рНKCl 4.11–4.31 и 4.24–4.65, соответственно (табл. 2). Содержание общего углерода, азота и фосфора заметно выше в карьере Кузьмолово, для Калелово эти показатели мало отличаются от показателей пионерной стадии.
На злаковой стадии присутствуют все модельные виды. На всех площадках встречались Agrostis capilla-ris (ПП 4–25%), Deschampsia cespitosa (+r–20%), Chamaenerion angustifolium (++–5%) и Artemisia vulgaris (++–5%). На 3 площадках из 4 отмечен Tussilago farfara (++–+1).
Кустарниковая стадия. Сообщества этой стадии описаны на 4 площадках (табл. 1, № 12–15), из которых 3 расположены на карьере Кузьмолово, и 1 – в Калелово. Во всех сообществах ОПП варьирует от 60 до 85%, общее число видов – от 19 до 38. В Кузьмолово верхний ярус состоит из подроста деревьев: Alnus incana (+1–25%), Betula pendula (2–17%), Populus tremula (5–15%), Pinus sylvestris (+1–5%), а также кустарников Salix myrsinifolia (2–15%) и S. caprea (+1–7%). Травяной ярус сообществ представлен слабо (ПП 2–17%), но значительно развит моховой покров (23–75%), в котором господствует Ceratodon purpureus (17–70%). Сообщество в Калелово отличается доминированием Alnus incana (45%), отсутствием других древесных пород, бóльшим покрытием трав (27%), из которых преобладает Agrostis capillaris, и слабым развитием мохового покрова (2%).
В сообществах этой стадии верхний ярус образован подростом деревьев и немногих кустарников и имеет неодинаковую высоту на разных площадках. На двух из них он формирует полог со средней высотой 50 см (максимальная высота осины – 115 см, ольхи – 145 см). На других площадках средняя высота верхнего яруса 110 см (максимальная высота осины 350 см на одной площадке и 520 см – на второй). Верхний ярус во всех сообществах более сомкнут (10–60%), чем фрагментарный ярус трав (2–27%), высота которого варьирует от 2 до 120 см.
Грунты на глубинах 5 и 20 см – от сильно- до среднекислых: рНKCl 2.83–4.34 и 2.55–4.80, соответственно (табл. 2); содержание общего углерода и азота снижается по сравнению со злаковой стадией, так как эти элементы концентрируются в основном в подстилке. Содержание общего фосфора на разных площадках сильно варьирует – примерно так же, как на злаковой стадии.
На всех площадках кустарниковой стадии встречались 3 модельных вида: Agrostis capillaris (++–13%), Deschampsia cespitosa (++–8%), Chamaenerion angustifolium (++–3%). Tussilago farfara отмечена на трех площадках (++– 2%), Artemisia vulgaris – на двух (++–+1).
Лесная стадия. Сообщества этой стадии встречаются редко, поэтому описаны на двух площадках (табл. 1, № 16–17), расположенных в краевой части карьера Кузьмолово. Здесь растительность формировалась на вскрыше, включавшей остатки старой дернины и почв, т.е. восстановление шло по типу вторичной сукцессии. Площадка № 16 располагается в ольшанике щучковом: сомкнутость крон 0.8, ПП травяного яруса – 70%, мохового – 15%; общее число видов – 33. В верхнем ярусе господствует Alnus incana (80%), в травяном – Deschampsia cespitosa (45%). Площадка № 17 описана на поляне среди более старых деревьев ольшаника звездчаткового: сомкнутость крон 0.3, ПП травяного яруса – 87%, мохового – 45%; общее число видов – 31. В травяном ярусе (господствующая высота 30–60 см) доминирует Stellaria holostea (40%), в моховом – Oxyrrhynchium hians (35%).
В ольшанике щучковом верхний ярус образуют молодые деревья Alnus incana высотой 10–15 м; в подлеске (высотой 50–210 см) встречаются Salix caprea, S. myrsinifolia, Sorbus aucuparia, Rubus idaeus, Sambucus racemosa и Padus avium Mill. На поляне в ольшанике звездчатковом верхний ярус образуют корневые отпрыски Alnus incana высотой 100–210 см. Травяной ярус обоих сообществ включает до 3 подъярусов: от 61 до 130 см (Artemi-sia vulgaris, Anthriscus sylvestris (L.) Hoffm., Geum urbanum L.), от 31 до 60 см (Deschampsia cespitosa, Equisetum sylvaticum L., Hieracium umbellatum L.), от 2 до 30 см (Veronica chamaedrys L., Stellaria holostea, S. media (L.) Vill).
Сильнокислые почвы на глубинах 5 и 20 см имеют рНKCl 3.61–3.95 и 3.70–3.84, соответственно (табл. 2), и заметно отличаются от почв кустарниковой стадии по содержанию общего углерода и азота, которое увеличивается в 2 и более раз. Содержание подвижного фосфора снижается практически до значений, наблюдаемых на пионерной стадии.
Из модельных растений на лесной стадии встречаются только 4: в обоих сообществах – Agrostis capillaris ( ++) и Artemisia vulgaris (+1–3%). Deschampsia cespitosa (45%) доминирует в одном из сообществ, вместе с ней участвует и Chamaenerion angustifolium (1%).
Таким образом, от стадии к стадии в сообществах увеличивается сомкнутость покрова, растет число видов, усложняется вертикальная структура за счет развития верхних ярусов из подроста древесно-кустарниковых пород, а также формирования наземного мохового покрова. Если для сообществ пионерной стадии характерно значительное участие рудеральных и немикотрофных растений, то на злаковой стадии наблюдается снижение их ценотической роли и замещение сначала факультативно-микотрофными злаками, а затем – на кустарниковой стадии – древесными растениями, для которых характерны актинориза и эктомикориза. На лесной стадии развивается сомкнутый травостой из видов, обычных для лесных сообществ, а пионерные виды присутствуют лишь спорадически. Параллельно с развитием растительности почвы становятся более кислыми, увеличивается содержание в них общего углерода и азота. Особенно это заметно при сравнении с голым субстратом, полностью лишенным растительности (табл. 2).
Модельные виды на разных стадиях сукцессии
Если оценить общую встречаемость модельных видов на всех пробных площадях, они сформируют следующий ряд: Agrostis capillaris (88%), Chamaenerion angustifolium (82%), Deschampsia cespitosa (76%), Tussilago farfara (71%), Artemisia vulgaris (59%). Более точное представление об участии каждого из модельных видов в сообществах разных стадий дает таблица 1.
Tussilago farfara и Chamaenerion angustifolium, обычно первыми заселяющие свободный субстрат, встречаются и на следующих этапах сукцессии, но не достигают заметного обилия (их ПП не превосходит 5%). На лесной стадии T. farfara выпадает из состава сообществ.
Tussilago farfara обычно растет на эрозионных и антропогенно нарушенных участках (берега водоёмов, склоны оврагов, оползни, свалки, пустыри и т.п.). Ценотическая роль этого вида в сукцессионном ряду исследованных сообществ незначительна и меняется мало. Исключение – пионерные более сомкнутые (ОПП 15–17%) сообщества в Калелово и наиболее сомкнутое (ОПП 85%) кустарниковое сообщество в Кузьмолово, где зафиксировано максимальное ПП вида, и он становится “наполнителем”.
Chamaenerion angustifolium – опушечно-лесной вид, который доминирует на пирогенных и иных нарушенных местообитаниях на ранних стадиях сукцессии, а в сукцессионно стабильных сообществах в течение долгого времени сохраняется с низким обилием (Broderick, 1990). В исследованных нами сообществах пионерной стадии C. angustifolium местами господствует, хотя в основном присутствует как “наполнитель” или “редкий”. В сообществах злаковой стадии ценотическая роль вида снижается, так как он не может конкурировать с интенсивно развивающимися злаками. По мере разрастания древесного подроста и кустарников C. angustifolium сохраняет статус от “редкого” до “наполнителя”.
Рудеральный вид Artemisia vulgaris, как и два вышеописанных модельных вида, встречается с небольшим ПП (не более 5%) на всех стадиях сукцессии, однако реже всего – на пионерной. На карьере Калелово A. vulgaris иногда становится “наполнителем” в сообществах пионерной и злаковой стадий.
Злаки Agrostis capillaris и Deschampsia cespitosa появляются на пионерной стадии, достигают максимального ПП на следующей – злаковой – стадии, а затем снижают свое обилие в сообществах. D. cespitosa – вид влажных или заболоченных лугов и сыроватых лесов. В сообществах пионерной, злаковой и кустарниковой стадий на карьере Калелово он имеет ценотическую роль “наполнителя”. В ольшанике щучковом на лесной стадии D. cespitosa господствует в травяном покрове, высокое ПП объясняется экологическими особенностями вида, характерного как для луговых, так и для лесных сообществ (Doronina, 2007), и, возможно, развитием ольшаника щучкового на материале вскрыши, богатом органикой и изначально содержавшем диаспоры растений.
Agrostis capillaris – вид лугов, опушек, песков и других местообитаний с бедными почвами. В некоторых пионерных сообществах на карьерах этот вид занимает положение согосподствующего. На злаковой стадии A. capillaris доминирует, лишь в одном случае (площадка № 10), уступая Calamagrostis epigeios. На кустарниковой стадии A. capillaris согосподствует или же становится “наполнителем” сообществ, на лесной – присутствует с ПП менее 1%.
Арбускулярная микориза и виталитет модельных видов
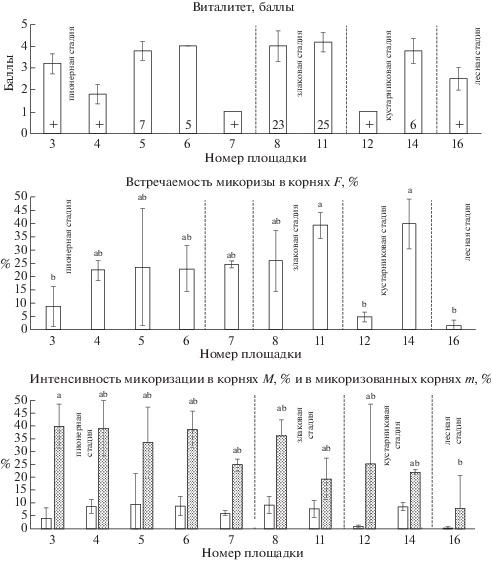
У Tussilago farfara не прослеживается четкого тренда изменения показателей микоризации по стадиям сукцессии33 (рис. 1). На пионерной стадии встречаемость микоризы варьирует сильнее всего, от полного отсутствия гриба в корнях до высоких значений (F = 59 ± 3.7%). Оба показателя интенсивности микоризации (M, m) также сильно варьируют, как и встречаемость микоризы. Виталитет T. farfara, как правило, выше в тех сообществах, где вид имеет наибольшее покрытие (рис. 1). Максимальный виталитет отмечен на площадке № 6 (пионерная стадия). В этом же сообществе достоверно более высокие значения интенсивности микоризации корней (M = 36 ± 2.2%) и обилие арбускул и везикул в корневой системе достоверно выше, чем во всех других сообществах (A = 23.4 ± 1.7%; V = 7.1 ± 1.2%). Особенностью развития АМ у данного вида в исследованных сообществах является то, что обилие арбускул всегда превышает обилие везикул в 2 раза и более (рис. 2).
Рис. 1.
Tussilago farfara: виталитет, встречаемость АМ, интенсивность микоризации в корнях (белые столбцы) и в микоризованных корнях (серые столбцы).
Обозначения. На графике виталитета в основании столбцов показано ПП вида в %, + означает ПП < 1%. На графиках интенсивности микоризации буквами a, b, A, B и т.д. обозначены группы значимо различающихся параметров согласно ANOVA и тесту Тьюки (P < 0.05). Планки погрешностей соответствуют стандартному отклонению.
Fig. 1. Tussilago farfara: vitality, arbuscular mycorrhiza frequency (F), intensity of root mycorrhization (M, white columns) and intensity of mycorrhization in mycorrhized roots (m, gray columns).
Footnote. On the graph of vitality, the species cover (%) is shown at the base of columns; + indicates the cover less than 1%. On the other graphs, letters a, b, A, B etc. represent significant differences (P < 0.05) using ANOVA and Tukey post-hoc tests. Bars represent ±1 SD.
пионерная стадия – pioneer stage
злаковая стадия – grassy stage
кустарниковая стадия – shrubby stage
номер площадки – plot number
баллы – points of vitality
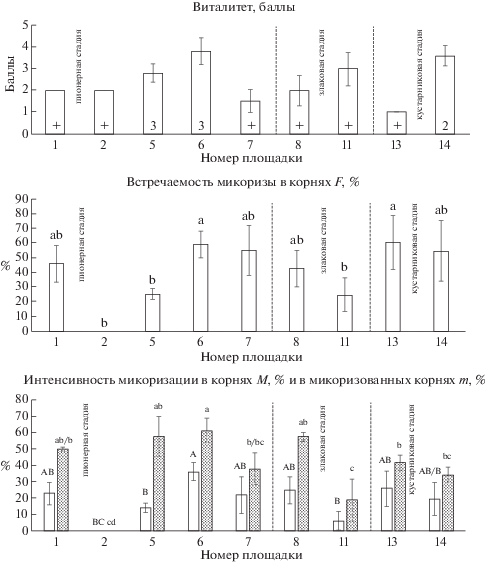
Рис. 2.
Обилие (%) арбускул (белые столбцы) и везикул (серые столбцы) в корнях модельных видов
Обозначения. Буквами a, b и т.д. обозначены группы значимо различающихся параметров согласно ANOVA и тесту Тьюки (P < 0.05). У Agrostis capillaris обилие арбускул и везикул в корнях достоверно не различается в разных сообществах. Планки погрешностей соответствуют стандартному отклонению.
Fig. 2. The abundance (%) of arbuscules (white columns) and vesicles (gray columns) in the roots of model plant species
Footnote. On the graphs letters a, b, c etc. represent significant differences (P < 0.05) using ANOVA and Tukey post-hoc tests. The abundance of arbuscules and vesicles in roots of Agrostis capillaris does not differ significantly in all communities. Bars represent ± 1 SD.
лесная стадия – forest stage
For other designations see the footnote to Fig. 1.
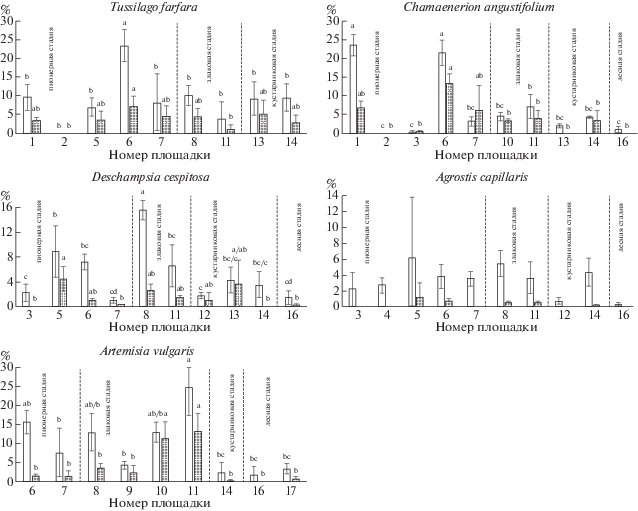
Микоризация Chamaenerion angustifolium сильно варьирует на пионерной стадии (рис. 3), в сообществах которой наблюдается как минимальная (F = 1.1 ± 1.1%), так и максимальная (F = 66 ± ± 1.1%) встречаемость микоризы. На злаковой и кустарниковой стадиях встречаемость АМ несколько ниже, на лесной стадии – достоверно ниже, чем в пионерном сообществе на площадке № 6, где этот показатель максимален. Интенсивность микоризации (M, m), так же как встречаемость, сильно варьирует на начальной стадии, причем М на всех последующих стадиях достоверно ниже, чем показатели, отмеченные в пионерных сообществах на площадках № 1 и № 6. На этих же площадках отмечено максимальное обилие арбускул (A = 23.6 ± 1.7% и 21.5 ± 1.9%, соответственно) и везикул (V = 13.3 ± 1.5% на площадке № 6), достоверно отличающееся от показателей сообществ других стадий (рис. 2).
Рис. 3.
Chamaenerion angustifolium: виталитет, встречаемость АМ, интенсивность микоризации в корнях (белые столбцы) и в микоризованных корнях (серые столбцы). Обозначения см. на рис. 1.
Fig. 3. Chamaenerion angustifolium: vitality, arbuscular mycorrhiza frequency, intensity of root mycorrhization (white columns) and intensity of mycorrhization in mycorrhized roots (gray columns).
лесная стадия – forest stage
For other designations see the footnote to Fig. 1.
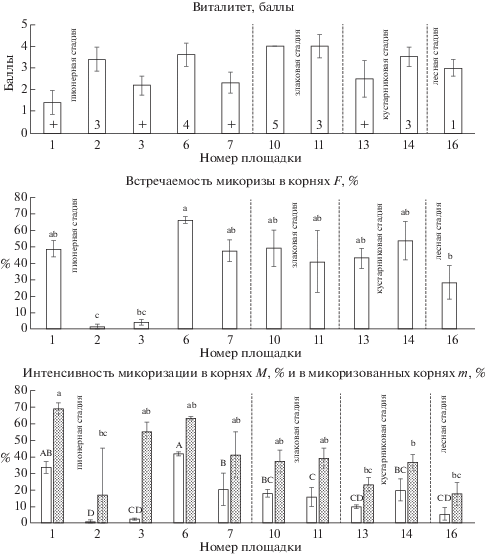
Виталитет C. angustifolium наиболее высок в сообществах, где вид имеет проективное покрытие более 1% и максимален на злаковой стадии (рис. 3). Интересно, что на площадке № 1 проективное покрытие и виталитет Chamaenerion angustifolium незначительны, оба эти показателя не достигают максимальных значений и на площадке № 6. В сообществах лесной стадии вид представлен редкими вегетирующими особями.
Очень низкая встречаемость АМ в корнях Chamaenerion angustifolium и Tussilago farfara в пионерном сообществе на площадке № 2, вероятно, объясняется отсутствием в грунте пропагул АМ-грибов. На площадке № 3, расположенной неподалеку, немногочисленные пропагулы образовали лишь слабую микоризу с C. angustifolium. Данные публикаций свидетельствуют о том, что AM-колонизация у данного вида в условиях сукцессии очень вариабельна: в интервале от 0 до 60% (Allen, 1988; Chapin, 1995), что связано с неравномерностью распределения грибных пропагул и, возможно, генетической изменчивостью растений по способности ассоциироваться с AM-грибами (Wolfe et al., 2005).
Для Deschampsia cespitosa характерен облигатный симбиоз с АМ-грибами (Mejstrik, 1972). Однако у этого вида наблюдается сильный разброс всех показателей микоризации в пределах каждой из стадий сукцессии (рис. 4). Так, в сообществах пионерной стадии встречаемость микоризы варьирует от низких (F = 9 ± 3.1%) до высоких значений (F = 42.1 ± 1%). Максимум встречаемости АМ (F = 58.3 ± 6.5%), достоверно отличающийся от значений на большинстве площадок других стадий, отмечен в сообществе злаковой стадии (площадка № 8). Здесь же зафиксировано максимальное обилие арбускул (A = 15.5 ± 0.9), достоверно отличное от других сообществ (рис. 2). Интенсивность микоризации корней D. cespitosa (M) показывает сходную со встречаемостью динамику от стадии к стадии, в то время как интенсивность микоризации в микоризованных фрагментах корней (m) достоверно не различается в разных сообществах. Обилие арбускул и везикул у D. cespitosa ниже, чем у двух ранее описанных видов. Виталитет варьирует в пределах каждой стадии, причем лучшие показатели соответствуют злаковой и лесной стадиям (рис. 4). Однако на площадке № 8, для которой характерна максимальная (по показателям встречаемости и интенсивности) микоризация корней D. cespitosa, отмечен худший виталитет и наименьшее проективное покрытие вида среди сообществ злаковой стадии. На кустарниковой стадии обилие и виталитет D. cespitosa несколько снижаются, а в лесном сообществе ольшаника на площадке № 16 повышаются: вид доминирует в травяном покрове (45%) и формирует маломощные дерновины, обильно развивающие генеративные побеги.
Рис. 4.
Deschampsia cespitosa: виталитет, встречаемость АМ, интенсивность микоризации в корнях (белые столбцы) и в микоризованных корнях (серые столбцы). Обозначения см. на рис. 1.
Fig. 4. Deschampsia cespitosa: vitality, arbuscular mycorrhiza frequency, intensity of root mycorrhization (white columns) and intensity of mycorrhization in mycorrhized roots (gray columns).
See the footnotes to Figs. 1 and 3.
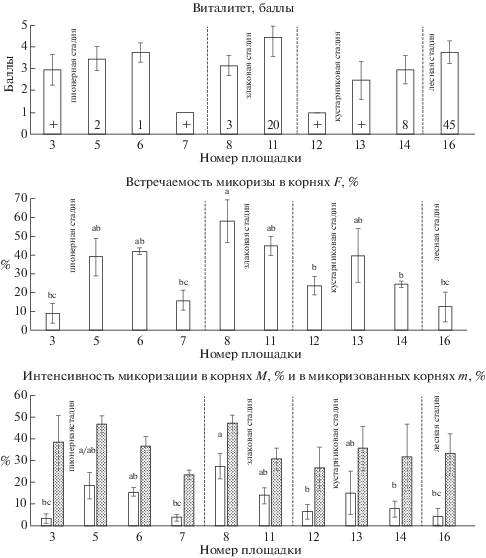
У Agrostis capillaris встречаемость АМ заметно варьирует в пределах пионерной и кустарниковой стадий сукцессии (рис. 5). Минимальное значение встречаемости микоризы отмечено в лесном сообществе (F = 1.3 ± 1.3%), а максимальные (F = = 39.3 ± 2.8% и 39.6 ± 5.4%) – в сообществах злаковой (площадка № 11) и кустарниковой (площадка № 14) стадий, соответственно. Интенсивность микоризации корней (M) невелика и достоверно не различается в сообществах разных стадий. В микоризованных фрагментах корней (m) интенсивность АМ наибольшая в сообществах пионерной стадии. Среди модельных видов у A. capillaris было наименьшее и достоверно не различающееся в разных сообществах обилие арбускул и везикул в корнях, причем обилие везикул редко превышало 1% (рис. 2). Лучший виталитет A. capillaris отмечен в сообществах, где ее проективное покрытие составляет 5% и более (рис. 5). Кроме того, в этих сообществах особи A. capillaris имеют в корнях везикулы, тогда как в других сообществах они отсутствуют. В сообществах кустарниковой (площадка № 12) и лесной (площадка № 16) стадий, где виталитет вида снижается, отмечены минимальные значения встречаемости и интенсивности микоризации его корней.
Рис. 5.
Agrostis capillaris: виталитет, встречаемость АМ, интенсивность микоризации в корнях (белые столбцы) и в микоризованных корнях (серые столбцы).
Обозначения см. на рис. 1.
Fig. 5. Agrostis capillaris: vitality, arbuscular mycorrhiza frequency, intensity of root mycorrhization (white columns) and intensity of mycorrhization in mycorrhized roots (gray columns).
See the footnotes to Figs. 1 and 3.
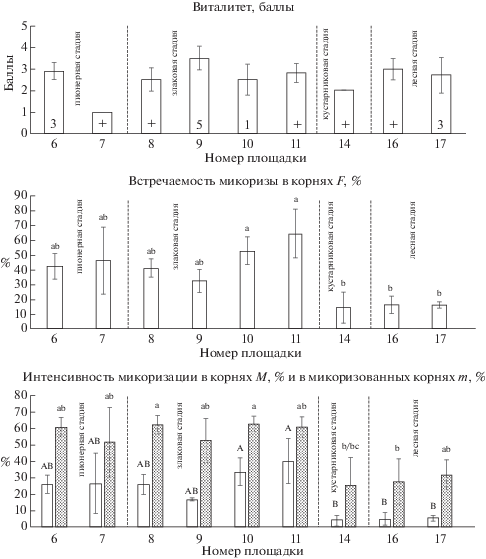
У Artemisia vulgaris встречаемость микоризы на двух первых и двух последних стадиях сукцессии достоверно различается44 (рис. 6). Максимальные значения встречаемости (F = 53 ± 5.5% и 64.7 ± ± 9.6%) отмечены в сообществах злаковой, а минимальные (F = 14.3 ± 6.1% и 16.2 ± 1.2%) – в сообществах кустарниковой и лесной стадий, соответственно. Аналогично встречаемости АМ в сообществах разных стадий различаются и оба показателя интенсивности микоризации корней. Обилие арбускул в корнях A. vulgaris больше, чем обилие везикул примерно в 2 раза. Достоверно более высокое обилие везикул наблюдается у особей, произрастающих в сообществах злаковой стадии на карьере Кузьмолово (V = 11.4 ± 2.6% и 13.1 ± 2.8%). Обилие арбускул в некоторых сообществах пионерной и злаковой стадий достоверно выше, чем на кустарниковой и лесной стадиях (рис. 2). Виталитет A. vulgaris не связан прямо с покрытием вида и параметрами микоризации. Однако в сообществе, где вид имел максимальное покрытие (площадка № 9), отмечен и его наилучший виталитет (рис. 6). В большинстве сообществ ценотическая роль A. vulgaris низкая, как и ее виталитет. В сообществе кустарниковой стадии (площадка № 14), где наблюдалось самое слабое развитие микоризы, A. vulgaris встречался с покрытием менее 1% и имел низкий виталитет.
Рис. 6.
Artemisia vulgaris: виталитет, встречаемость АМ, интенсивность микоризации в корнях (белые столбцы) и в микоризованных корнях (серые столбцы).
Обозначения см. на рис. 1.
Fig. 6. Artemisia vulgaris: vitality, arbuscular mycorrhiza frequency, intensity of root mycorrhization (white columns) and intensity of mycorrhization in mycorrhized roots (gray columns).
See the footnotes to Figs. 1 and 3.
Таким образом, микоризация модельных видов в ходе восстановительной сукцессии не возрастает, а иногда даже снижается. При этом у каждого вида она наиболее сильно варьирует в пионерных сообществах. Слабо выраженная тенденция увеличения интенсивности микоризации наблюдается у модельных видов при переходе к злаковой стадии, в сообществах которой увеличивается их проективное покрытие и, в некоторых случаях, виталитет.
Tussilago farfara и Chamaenerion angustifolium – обычные первопоселенцы свободных субстратов. Ценотическая роль T. farfara в сукцессионном ряду исследованных сообществ незначительна и мало меняется. Ее максимальное обилие и виталитет соответствуют пионерной стадии сукцессии, хотя сравнительно высокие встречаемость и интенсивность микоризации у данного вида наблюдались и на злаковой, и на кустарниковой стадиях.
Chamaenerion angustifolium местами господствует на пионерной стадии, в сообществах злаковой и кустарниковой стадий становится второстепенным, а в лесном сообществе – редким. На пионерной стадии у C. angustifolium отмечены как максимальные, так и минимальные показатели микоризации, хотя наилучший виталитет он имеет в сообществах злаковой стадии.
Виды, типичные для второй стадии – Deschampsia cespitosa и Agrostis capillaris. D. cespitosa чаще всего является “наполнителем” в исследованных сообществах. Наилучший виталитет вид имеет на злаковой стадии, в сообществах которой отмечена также наибольшая встречаемость АМ, хотя интенсивность микоризации корней D. cespitosa высока и на пионерной, и на кустарниковой стадиях. В лесном сообществе этот вид господствует в травяном покрове, но его виталитет ниже, чем на злаковой стадии, и встречаемость микоризы в корнях низкая.
Agrostis capillaris наибольшего господства и виталитета достигает на злаковой стадии, в лесных сообществах встречаемость вида не превышает нескольких особей на площадку. У A. capillaris высокая встречаемость микоризы отмечена и на злаковой, и на кустарниковой стадиях, а интенсивность микоризации корней почти не различается на всех стадиях, кроме лесной.
Artemisia vulgaris встречается на всех стадиях с небольшим проективным покрытием (не более 5%). Похожие значения проективного покрытия и виталитета A. vulgaris имеет в сообществах разных стадий (вплоть до лесной), однако наибольшие показатели микоризации вида соответствуют пионерной и злаковой стадиям.
Близкие результаты были получены в работах Д. Касовской, изучавшей пионерные сообщества на отвалах месторождения глины (Kasowska, 2002). Эдафические условия сильно отличались от исследованных нами: субстрат содержал больше глинистой и илистой фракций, его pH был более приближен к нейтральному. Были изучены две стадии сукцессии: инициальная (1–2 года) и продвинутая (8–9 лет). Инициальная стадия характеризовалась ОПП растительности около 15% и доминированием рудеральных видов, в первую очередь Polygonum aviculare L. На продвинутой стадии ОПП составляло около 20%, преобладала рудеральная и злаковая растительность с доминированием Tussilago farfara. На первой стадии у Agrostis capillaris АМ не была обнаружена, а у T. farfara и Artemisia vulgaris – выявлена АМ без везикул. На второй стадии у A. capillaris развилась АМ, а у T. farfara и A. vulgaris образовались везикулы в корнях. Д. Касовская заключает, что на инициальной стадии АМ симбиоз не был эффективным, что отражалось в малом обилии арбускул и везикул, тогда как на более долго зараставшем участке АМ грибы оказались более адаптированными к условиям среды и растениям-хозяевам.
Все изученные нами модельные виды на лесной стадии, как правило, имели слабую микотрофность. Это можно объяснить тем, что все они – светолюбивы и хуже развиваются в условиях затенения пологом леса, что негативно сказывается и на микоризации. Кроме того, возможно угнетение АМ грибов вследствие конкуренции с другими почвенными организмами и/или неподходящих условий в почвах лесных сообществ.
Предположение, что высокая степень микоризации обеспечивает высокие виталитет и ценотическую роль вида, не подтверждено полученными данными. Только в отдельных случаях (на отдельных площадках) виталитет, ценотическая роль и интенсивность микоризации модельных видов были связаны положительно. Например, у Tussilago farfara в пионерном сообществе на площадке № 6 отмечены максимальные значения всех перечисленных показателей, а у Agrostis capillaris в сообществе кустарниковой стадии (площадка № 12) наблюдались минимальные значения виталитета, проективного покрытия и интенсивности микоризации корней.
С другой стороны, были описаны сообщества пионерной (площадка № 6) и злаковой (площадка № 11) стадий, в которых все модельные виды имели наилучший виталитет и высокую встречаемость АМ в корнях (не менее 20%). В почве на площадке № 11 (табл. 2) выявлено высокое содержание фосфора, азота и органики, что благоприятно для развития растений и не подавляет развитие АМ грибов. На площадке № 6 на глубине 20 см также отмечено значительное для пионерной стадии содержание подвижного фосфора.
ЗАКЛЮЧЕНИЕ
Восстановительная сукцессия на песчаных карьерах представлена четырьмя последовательными стадиями зарастания свободного субстрата и восстановления древесной растительности: пионерной, злаковой, кустарниковой и лесной.
У всех модельных видов показатели микоризации наиболее сильно варьируют в пионерных сообществах, что можно объяснить неодинаковыми условиями, поскольку каждый первичный экотоп заселяется случайным набором видов, по-разному взаимодействующих друг с другом и изменяющих абиотическую среду. Слабо выраженная тенденция увеличения интенсивности микоризации наблюдается при переходе к злаковой стадии, в сообществах которой увеличивается проективное покрытие и, в некоторых случаях, виталитет модельных видов. Общим для последних оказалось значительное снижение показателей АМ в сообществах лесной стадии, сопровождавшееся уменьшением роли видов, наиболее характерных для начальных стадий сукцессии.
Полученные данные не подтверждают предположение, что высокая степень микоризации всегда обеспечивает высокие виталитет и ценотическую роль вида. Более того, богатство субстрата часто оказывается более важным фактором, определяющим жизненность растений. Для более точной оценки взаимосвязей видов трав и арбускулярной микоризы в сообществах восстановительной сукцессии на песчаных карьерах необходимы дальнейшие исследования.
Список литературы
Aikio S. 2000. Plant adaptive strategies in relation to variable resource availability, soil microbial processes and ecosystem development: Diss. … Doct. Sci. Oulu. 35 p.
Akhmetzhanova A.A., Soudzilovskaia N.A., Onipchenko V.G., Cornwell W.K., Agafonov V.A., Selivanov I.A., Cornelissen J.H.C. 2012. A rediscovered treasure: mycorrhizal intensity database for 3000 vascular plant species across the former Soviet Union. – Ecology. 93(2): 689–690. https://doi.org/10.2307/23143955
Allen M.F. 1988. Re-establishment of VA mycorrhizas following severe disturbance: comparative patch dynamics of a shrub desert and a subalpine volcano. – Proc. Roy. Soc. Edinburgh. 94(B): 63–71. https://doi.org/10.1017/S0269727000007132
[Barkman] Баркман Я. 1991. Верность и характерные виды: критическая оценка. – Бот. журн. 76(7): 936–949.
Broderick D.H. 1990. The biology of Canadian weeds. 93. Epilobium angustfolium L. (Onagraceae). – Can. J. Plant Sci. 70: 247–259. https://doi.org/10.4141/cjps90-027
Cázares E., Trappe J.M., Jumponnen A. 2005. Mycorrhiza-plant colonization patterns on a subalpine glacier forefront as a model system of primary succession. – Mycorrhiza. 15: 405–416. https://doi.org/10.1007/s00572-004-0342-1
Chapin D.M. 1995. Physiological and morphological attributes of two colonizing plant species on Mount St. Helens. – Am. Midl. Nat. 133: 76–87.
Daft M.J., Nicolson T.H. 1974. Arbuscular mycorrhizas in plants colonizing coal wastes in Scotland. – New Phytol. 73: 1129–1138. https://doi.org/10.1111/j.1469-8137.1974.tb02142.x
[Doronina] Доронина А.Ю. 2007. Сосудистые растения Карельского перешейка (Ленинградская область). М. 574 с.
Gemma J.N., Koske R.E. 1990. Mycorrhziae on recent Volcanic substrates in Hawaii. – Am. J. Bot. 79: 1193–1200. https://doi.org/10.2307/2444630
[GOST 26107-84] ГОСТ 26107-84. 1984. Почвы. Методы определения общего азота. М. 11 с.
[GOST 26483-85] ГОСТ 26483-85. 1985. Почвы. Приготовление солевой вытяжки и определение ее рН по методу ЦИНАО. М. 6 с.
[GOST 26213-91] ГОСТ 26213-91. Почвы. Методы определения органического вещества. 1993. М. 8 с.
[GOST Р 54650-2011] ГОСТ Р 54650-2011. 2013. Почвы. Определение подвижных соединений фосфора и калия по методу Кирсанова в модификации ЦИНАО. М. 8 с.
Heijden van der M.G.A., Klironomos J.N., Ursic M., Moutoglis P., Streitwolf-Engel R., Boller T., Wiemken A., Sanders I.R. 1998. Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. – Nature. 396: 72–75. https://doi.org/10.1038/23932
Heijden van der M.G.A., Martin F.M., Selosse M.-A., Sanders I.R. 2015. Mycorrhizal ecology and evolution: the past, the present, and the future. – New Phytol. 205: 1406–1423. https://doi.org/10.1111/nph.13288
[Ipatov, Mirin] Ипатов В.С., Мирин Д.М. 2008. Описание фитоценоза: Методические рекомендации. СПб. 71 с.
IPNI: The International Plant Names Index. 2020. http://www.ipni.org (Accessed 12.06.2020).
Kasowska D. 2002. Mycorrhizal status of plants in two successional stages on spoil heaps from fireloam mining in Lower Silesia (SW Poland). – Acta Soc. Bot. Pol. 71: 155–161. https://doi.org/10.5586/asbp.2002.018
Lambers H., Raven J.A., Shaver G.R., Smith S.E. 2008. Plant nutrient-acquisition strategies change with soil age. – Trends Ecol. Evol. 23(2): 95–103. https://doi.org/10.1016/j.tree.2007.10.008
[Lukina, Ryazanova] Лукина Н.В., Рязанова С.В. 2012. Особенности микоризообразования в техногенных экосистемах. – Экосистемы, их оптимизация и охрана. 7: 261–269.
Mejstrik V.J. 1972. Vesicular-arbuscular mycorrhizas of the species of a Molinietum coeruleae L.I. association: the ecology. – New Phytol. 71: 883–890. https://doi.org/10.1111/j.1469-8137.1972.tb01968.x
[Mirkin et al.] Миркин Б.М., Розенберг Г.С., Наумова Л.Г. 1989. Словарь понятий и терминов современной фитоценологии. M. 223 с.
Mycorrhiza Manual. 2001. https://www2.dijon.inrae.fr/mychintec/Protocole/protoframe.html (Accessed 12.06.2020).
Öpik M., Moora M., Liira J., Zobel M. 2006. Composition of root-colonizing arbuscular mycorrhizal fungal communities in different ecosystems around the globe. – J. Ecol. 94: 778–90. https://doi.org/10.1111/j.1365-2745.2006.01136.x
Phillips J.M., Hayman D.S. 1970. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. – Trans. Brit. Mycol. Soc. 55: 158–161. https://doi.org/10.1016/S0007-1536(70)80110-3
Püschel D., Rydlova J., Vosatka M. 2007. Mycorrhiza influences plant community structure in succession on spoil banks. – Basic Appl. Ecol. 8: 510–520. https://doi.org/10.1016/j.baae.2006.09.002
Reeves F.B., Wagner D., Moorman T., Kiel J. 1979. Role of endomycorrhizae in revegetation practices in the semi-arid west. 1. Comparison of incidence of mycorrhizae in severely disturbed vs natural environments. – Amer. J. Bot. 66: 6–13.
Rydlová J., Vosátka M. 2001. Associations of dominant plant species with arbuscular mycorrhizal fungi during vegetation development on coal mine spoil banks. – Folia Geobot. 36: 85–97. https://doi.org/10.1007/BF02803141
Smith S.E., Read D.J. 2008. Mycorrhizal symbiosis. Cambridge. 787 p.
Spatafora J.W., Chang Y., Benny G.L., Lazarus K., Smith M.E., Berbee M.L., Bonito G., Corradi N., Grigoriev I., Gryganskyi A., James T.Y., O’Donnell K., Roberson R.W., Taylor T.N., Uehling J., Vilgalys R., White M.M., Stajich J.E. 2016. A phylum-level phylogenetic classification of zygomycete fungi based on genome-scale data. – Mycologia. 108(5): 1028–1046. https://doi.org/10.3852/16-042
[Sumina] Сумина О.И. 2013. Формирование растительности на техногенных местообитаниях Крайнего Севера. СПб. 337 с.
The Plant List. 2013. Version 1.1. Published on the Internet; http://www.theplantlist.org/ (Accessed 12.06.2020).
Trouvelot A., Kough J.L., Gianinazzi-Pearson V. 1986. Mesure du taux de mycorhization VA d’un système radiculaire. Recherche de méthodes ayant une signification fonctionnelle. – In: Physiological and Genetical Aspects of Mycorrhizae. Paris. P. 217–221.
[Veselkin] Веселкин Д.В. 2012a. Стабилизация соотношения между числом видов растений разного микоризного статуса – один из аттрактантов прогрессивных сукцессий? – Известия Самарского научного центра РАН. 1206–1209.
[Veselkin] Веселкин Д.В. 2012b. Участие растений разного микотрофного статуса в сукцессии при формировании “агростепи”. – Экология. 4: 270–275.
[Veselkin, Betekhtina] Веселкин Д.В., Бетехтина А.А. 2011. Участие растений разного микотрофного статуса в техногенно обусловленных сукцессиях в степной зоне Урала – Вест. ОГУ. 131(2): 44–46.
[Veselkin et al.] Веселкин Д.В., Лукина Н.В., Чибрик Т.С. 2015. Соотношение микоризных и немикоризных видов растений в первичных техногенных сукцессиях. – Экология. 5: 345–353. https://doi.org/10.7868/S0367059715050200
[Vorobiev et al.] Воробьев Н.И., Юрков А.П., Проворов Н.А. 2016. Свидетельство № 2016612112 от 12.02.2016 о регистрации программы ЭВМ “Программа вычисления индексов микоризации корней растений”. М. Федеральная служба по интеллектуальной собственности.
Wang B., Qiu Y.L. 2006. Phylogenetic distribution and evolution of mycorrhizas in land plants. – Mycorrhiza. 16: 299–363. https://doi.org/10.1007/s00572-005-0033-6
Westhoff V., van der Maarel E. 1978. The Braun-Blanquet approach. – In: Classification of plant communities. The Hague. P. 287–399.
Wolfe B.E., Husband B.C., Klironomos J.N. 2005. Effects of a belowground mutualism on an aboveground mutualism. – Ecol. Letters. 8: 218–223. https://doi.org/10.1111/j.1461-0248.2004.00716.x
[Yurkov, Semenov] Юрков А.П., Семенов Д.Г. Биология. Особенности применения световой микроскопии в анализе биологических тканей. 2019. СПб. 56 с.
[Yurkov et al.] Юрков А.П., Шишова М.Ф., Семенов Д.Г. Особенности развития люцерны хмелевидной с эндомикоризным грибом. 2010. Саарбрюккен. 215 с.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал